
Abstract
The federally endangered Schaus swallowtail butterfly (Heraclides aristodemus ponceanus) has reached critically low numbers. Exotic ants are a potential threat to H. a. ponceanus and other rare butterflies as they can attack immature stages. Ant surveys conducted in subtropical dry forests in Biscayne National Park documented ant species diversity and relative abundance. A caterpillar predator exclusion experiment using physical barriers in different combinations evaluated caterpillar survivorship of both early and late instar caterpillars exposed to different threats. Ant-caterpillar interactions were also documented by placing caterpillars on plants and observing physical interactions between caterpillars and ants. A total of 1418 ants comprising 25 ant species was captured and identified. In canopies of H. a. ponceanus host plants, 243 ants comprising 12 species were found. The four most common ants collected in the host plant canopies were Pseudomyrmex gracilis, Camponotus planatus, Cremastogaster ashmeadi, and Camponotus floridanus. The predator exclusion experiment revealed survivorship was significantly lower for early and late instar caterpillars without any physical barrier, as well as for early instars not protected by a mesh cage. Pseudomyrmex gracilis and C. floridanus were more aggressive towards caterpillars in comparison to other ant species; these two species ranked first and second in the “ant danger index” ranking predatory abilities of the four most common ant species. Pseudomyrmex gracilis is a common arboreal exotic ant in Biscayne National Park and presents a major threat to caterpillars during their earliest life stages.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Mortality at every life history stage is common for most invertebrates, but for species whose numbers are reduced or reproduction limited to a short time period, heavy predation on immatures can severely threaten their continued existence (Schoener et al. 2001; Beuzelin et al. 2009; Wagner and Van Driesche 2010). The federally endangered Schaus swallowtail butterfly (Heraclides aristodemus ponceanus) inhabits subtropical dry forest and is known from a limited number of locations. Its occurrence is associated with clumped distributions of torchwood (Amyris elemifera, Rutaceae), the primary larval host plant (Emmel et al. 1988; Jameson 2002; United States Fish and Wildlife Service 2008). Schaus swallowtail populations in Biscayne National Park (BNP) and North Key Largo (NKL) have declined precipitously over the last 15 years (Salvato 2008; United States Fish and Wildlife Service 2008; Fig. 1). In 2012, two projects were implemented to increase H. a. ponceanus population: a captive-breeding program at the University of Florida (Daniels 2014) and the National Park Service (NPS) Schaus swallowtail habitat enhancement project in BNP. The Schaus swallowtail habitat enhancement project capitalized upon ongoing projects that required the elimination of invasive plants in particular sections within subtropical dry forests at Elliott and Adams Keys (Whelan 2011; Whelan and Atkinson 2015); over 3000 host plants for the H. a. ponceanus, mostly A. elemifera and Zanthoxylum fagara (Rutaceae), were planted and nurtured in restoration sites at Elliott and Adams Keys. Despite a bottom-up approach (increased food source) to bolster H. a. ponceanus populations (Jameson 2002; Dennis et al. 2004; Mathew and Anto 2007), investigations of top-down effects on a rare butterfly concentrated in BNP and NKL are nonexistent (Didham et al. 2007; Salvato 2008; United States Fish and Wildlife Service 2008). Predation by invertebrates, such as ants, can thwart the recovery process (Schoener et al. 2001; Wagner and Van Driesche 2010; Lach et al. 2016). Specifically, most ants are generalist and opportunistic foragers, numerically dominant in subtropical and tropical ecosystems, and active day and night foragers (Carroll and Janzen 1973; Jeanne 1979; Brown 2000; Floren et al. 2002). The ecological impacts of exotic ants inhabiting the islands are unknown, despite their ubiquitous presence in the forests (United States Fish and Wildlife Service 2008; Moreau et al. 2014).

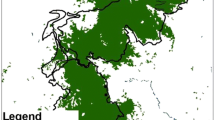
Geographic range of the federally endangered Heraclides aristodemus ponceanus in south Florida. The historic ranged is outlined in yellow, the current range is outlined in red, and the boundary of Biscayne National Park is outlined in blue. Map generated by Lydia Cuni
Exotic ants can negatively affect other organisms through predation, competition, and alteration of local habitats (Williams 1994; Vitousek et al. 1996; Mack and D’Antonio 1998; Holway et al. 2002; Lach and Hooper-Bùi 2010; Lach et al. 2016). Prolific exotic ants can become invasive in new territories with novel food resources, reduced interspecific and intraspecific competition, release from natural enemies, and enhanced physiologic abilities to thrive in disturbed environments (Holway et al. 2002; Krushelnycky et al. 2010; Lach et al. 2016; Calcaterra et al. 2016). Approximately 37 exotic ant species have been identified in the Florida Keys, including three species on the top 100 world’s worst invasive alien species list: the big-headed ant (Pheidole megacephala), the little fire ant (Wasmannia auropunctata), and the red imported fire ant (Solenopsis invicta) (Lowe et al. 2000; Moreau et al. 2014). Few studies have investigated direct and indirect impacts of exotic ants on native flora and fauna in the Florida Keys, with the exception of red imported fire ants; a laboratory study confirmed that red imported fire ants are highly efficient predators of all immature life stages of the giant swallowtail butterfly (Heraclides cresphontes), which were used as a surrogate species for H. a. ponceanus (Forys et al. 2001).
Solenopsis invicta has been considered a potential threat to H. a. ponceanus because of its aggressive nature (Forys et al. 2001). Solenopsis invicta displace native species in disturbed environments; they are efficient predators and scavengers, consuming vulnerable organisms including ticks, caterpillars, beetle grubs, and even mosquito larvae in moist substrate (Porter and Savignano 1990; Forys et al. 2002; Zettler et al. 2004; Allen et al. 2004; Beuzelin et al. 2009). Solenopsis invicta form large colonies that can monopolize disturbed sites adjacent to subtropical dry forests (Forys et al. 2001, 2002; Tschinkel 2006). In south Florida, they are unlikely to colonize interior parts of subtropical dry forests because they are mainly habitat-restricted to human-modified, disturbed sites (Tschinkel 1988, 2006; King and Tschinkel 2006).
The exotic graceful twig ant (Pseudomyrmex gracilis), is arboreal and also poses potential risk to rare butterflies as the ants search for prey on shrubs and trees (Saarinen and Daniels 2006; Wetterer 2010). An opportunistic, solitary forager, P. gracilis mainly scavenges for food in its natural range (Mexico, Central, and South America; Whitcomb et al. 1972). In south Florida, its predatory behavior is deleterious to rare and vulnerable invertebrate species because these ants are physically larger than the other native Pseudomyrmex ant species in south Florida (Wetterer 2010; Deyrup 2016); years earlier, Deyrup et al. (2000) warned that the graceful twig was a “possible ecological villain” in Florida. Pseudomyrmex gracilis is a common pest on shrubs and trees in both urban and natural environments (MacGown and Hill 2010).
The exotic little fire ant (Wasmannia auropunctata) is a small stinging ant that can overwhelm invertebrate prey through aggressive recruitment (Wetterer and Porter 2003). In BNP, W. auropunctata has been collected on both trees and ground. The little fire ant inhabits both disturbed and intact forests in BNP. Wasmannia auropunctata and P. gracilis are native to Mexico and Central America (Wetterer and Porter 2003; Wetterer 2010). All three exotic ant species are native to South America (Buren et al. 1974; McGlynn 1999).
The limited range of Heraclides aristodemus ponceanus, reduced habitat, and exposure to natural extreme weather events may not allow its immature stages to withstand exotic and native ant predation over time (Schoener et al. 2001; United States Fish and Wildlife Service 2008), limiting the number of adult butterflies of this endangered species. The current government management protocol for H. a. ponceanus reported exotic ants as major threats to the eggs and larvae (United States Fish and Wildlife Service 2008). Here we address potential impacts ants may have on caterpillars that use A. elemifera and Z. fagara as larval host plants. We want to know: (1) What ant species forage in the canopies of A. elemifera and Z. fagara in BNP?; (2) Are the most common ant species native or exotic?; (3) Are crawling or non-crawling predators greater threats to Heraclides caterpillar species?; and (4) How do interactions between the most common ant species and Heraclides caterpillar species compare?
Methods
Site description
Biscayne National Park (BNP) became a national park in 1980, with most of its area (95%) comprised of marine environments. The terrestrial area consists of a narrow strip of land on the mainland and 42 islands, most of which is dominated by mangrove forest (2400 ha). Coastal subtropical dry forests, critical habitat for H. a. ponceanus, make up only 723 ha (23%) of the terrestrial lands (Whelan et al. 2013). Elliott Key (25°27′12.39″N, 80°11′39.05″W), the largest island, is roughly 7 miles long, and nearly a mile wide, with subtropical dry forests covering 68 percent of the area (Whelan et al. 2013). On Adams Key (25°23′52.34″N, 80°14′05.67″W), subtropical dry forests account for 43% of the area.
South Florida has a distinct wet-dry season: the rainy season typically starts early June and continues through October; however, dry spells can occur in July and part of August (Snyder et al. 1990). The univoltine H. a. ponceanus emerges from its chrysalis typically in May with a flight season lasting until mid-June; occasionally, a second emergence happens mid-August and September depending on the amount of rain (Loftus and Kushlan 1984; Emmel et al. 1988; Minno and Emmel 1993). Heraclides aristodemus ponceanus have been reported flying as early as April. Currently, the largest population of H. a. ponceanus inhabits BNP, particularly Elliott and Adams Keys (Minno 2015).
Ant survey
Ant surveys were conducted for 1 year (December 2013–November 2014) in the subtropical dry forest at Elliott and Adams Keys. Ants were collected using pitfall traps to characterize relative species abundance and diversity (Wang et al. 2001; Andersen et al. 2002); each trap was a 50 mL plastic test tube filled with 20 mL of lemon-scented, soapy water solution, with one hole (1.5 cm diameter) drilled into each screw-on cap to protect against adverse weather conditions (Wang et al. 2001; Ribas et al. 2003; Lubertazzi and Tschinkel 2003). Each sampled area was selected based on the presence of A. elemifera and Z. fagara, along a North–South gradient at Elliott Key, and an East–West gradient at Adams Key, differing because of the islands’ overall shapes. Non-host trees in the forest, 10–15 m away from each A. elemifera and Z. fagara sampled, were also sampled. Four pitfall traps were placed on or near each tree: one at the base, one on the trunk (2 m above the ground), and two in the canopy. Pitfall traps were dispersed throughout the sampled areas every month, collecting crawling insects for 5 days. Afterwards, collected ants were preserved for subsequent determination. A total of 296 pitfall traps was deployed on 74 trees at Elliott and Adams Keys. One hundred forty-eight traps were placed in tree canopies, 74 were placed on the trunk, and 74 were partially buried in the ground adjacent to the base of each tree. Thirty A. elemifera, 14 Z. fagara, and 30 non-host trees were sampled during the study.
Caterpillar predator exclusion experiment
A 2 × 6 factorial design with six treatments was employed to compare the effects of crawling and non-crawling predators against caterpillars of two different sizes (modified from Koptur et al. 1998; Cuautle and Rico-Gray 2003; Mestre et al. 2016). This experiment was conducted from March 2014 to November 2014. Using soft forceps, early (<1.5 cm) and late (>3 cm) instar H. cresphontes caterpillars (used as a surrogate for H. a. ponceanus caterpillar) were gently placed on Z. fagara in the forest at Elliott Key only; logistical constraints did not allow this experiment to be adequately conducted at Adams Key. Despite H. a. ponceanus preference for A. elemifera, they also utilize Z. fagara as a secondary host plant (Jameson 2002; United States Fish and Wildlife Service 2008). The majority of H. cresphontes caterpillars, raised in an outdoor nursery at Florida International University for both the caterpillar predator exclusion experiment and ant-caterpillar interaction study, were fed Z. fagara leaves, due to both logistical and financial constraints of maintaining an adequate supply of A. elemifera.
Treatments applied to caterpillars were: (1) no barriers (no tanglefoot/no cage), (2) tanglefoot present/no cage, (3) no tanglefoot/cage with holes present, (4) tanglefoot present/cage with holes present, (5) no tanglefoot/cage present, and (6) tanglefoot present/cage present. Tanglefoot® is a non-toxic, non-drying, sticky resin that creates a barrier and deters crawling insects from crossing it. Cages were made from recycled 32 oz sports drink bottles that were cut open, shaped into a frame, and placed over a wild lime branch. Ultra-fine cotton cheesecloth was placed over the sports drink bottle frame and sealed with staples and white zip ties. One caterpillar was placed on a branch inside the cage or without a cage; for early instar caterpillars, each treatment had 16 replicates; for late instar caterpillars, each treatment had 15 replicates. Some treatments required holes in the mesh cage; therefore, six holes (r = 1.5 cm) were cut into the mesh. Each individual Z. fagara tree received a randomly assigned assortment of 6–8 treatments spread across different branches. Each Z. fagara tree was thoroughly scrutinized for caterpillar presence or absence after 5 days and recorded to determine survivorship for each treatment. No Z. fagara tree was used more than once during the experiment.
Ant-caterpillar interaction study
Ant interactions with both early (<1.5 cm in length) and late (>3 cm) instar caterpillars were conducted in the forests at Elliott and Adams Keys from April 2014 to October 2014. H. cresphontes caterpillars were again used as surrogate species, as they also use A. elemifera and Z. fagara as host plants in BNP, and interact with the same ant species. Foraging ant locations were selected based on relative abundance (determined from previous ant surveys) in the canopy of A. elemifera and Z. fagara. An individual caterpillar was placed on a leaf (A. elemifera and Z. fagara) and allowed to settle before interacting with ants. Observations ended after 10 min upon first contact, unless the caterpillar was removed before that time (Oliveira et al. 1987; Bächtold et al. 2012; Sendoya and Oliveira 2015). Observers timed each interaction trial from the first contact between ant and caterpillar. The caterpillar’s fate was reported after each trial. Caterpillar mortality was defined as removal by ant species or caterpillar leaf abandonment (dropping from the leaf to avoid ant harassment). No caterpillars or ants were used more than once during this study; for early instar caterpillars, 16 trials each were conducted with Camponotus floridanus and Camponotus planatus, 15 trials with P. gracilis, and 10 trials with Cremastogaster ashmeadi; for late instar caterpillars, 15 trials each were conducted with C. floridanus, C. planatus, and P. gracilis, and 11 trials with C. ashmeadi.
Data analysis
A generalized linear model (negative binomial with log link to account for overdispersion) was used to model the frequency of P. gracilis on trees. The negative binomial model was selected as the best-fit model to account for overdispersion and true zeros. Model comparisons (poisson and negative binomial) were analyzed using the program IBM® SPSS® Statistics 22 (2013).
A contingency table analysis was performed on data collected from the caterpillar predator exclusion experiment. Adjusted standardized residuals (Z-scores) calculated from the contingency table were transformed by squaring them to obtain Chi square values. Afterwards, the Chi square values were used to calculate exact p values with the significance function group in SPSS. Exact p values were compared to the adjusted α-value to determine significance. Adjusted α-values were calculated by dividing the accepted α-value of 0.05 by the total number of factors (12) which equaled 0.004167. Exact p values equal to or less than the adjusted α-value were considered significant.
A Shapiro–Wilk test for normality was performed on the discovery time for the ant-caterpillar interaction study. A one-way ANOVA was performed followed by the Fisher’s Least Significant Difference post-hoc test. All data analyses reported above were conducted using IBM® SPSS® Statistics 22 (2013).
An Ant Danger Index (ADI) was generated to rank the predatory abilities of ants against caterpillars. Each ant species was given an ADI score based on relative abundance in tree canopies (A. elemifera and Z. fagara), average time to first interaction with caterpillar, rate of ant recruitment, and ant-caterpillar interactions leading to caterpillar mortality. The total score for each ant species was divided by 25 (the maximum score an ant species could receive) and then multiplied by 100 (percentage of total possible score) for their final ADI score, which ranged from 0 to 100, allowing the species to be ranked (Possley et al. 2016).
Results
Ants on and around plants
Twenty-five ant species were collected in pitfall traps on trees at Elliott and Adams Keys in BNP (Table 1; Fig. 2), a total of 1418 individual ants. Taxa were determined with help from Mark Deyrup (Archbold Biological Station, Venus, Florida). Twelve ant species (243 total ants) were collected in canopies of torchwood and wild lime (Table 2; Fig. 3); the four most common of these were P. gracilis, C. planatus, C. ashmeadi, and C. floridanus. The most abundant ant collected in canopies of A. elemifera and Z. fagara was P. gracilis, representing 35% of the total ants collected; C. planatus was second at 21%; C. ashmeadi was third at 18%; and C. floridanus was fourth at 13% (Fig. 3). These four ant species represented 87% of collected ants in canopies of A. elemifera and Z. fagara. Both P. gracilis and C. planatus are exotic ant species in south Florida (Moreau et al. 2014; Deyrup 2016).

Percentage of ant species captured in pitfall traps at Elliott and Adams Keys (tree canopy, trunk, and base). Overall, 1418 total ants comprising 25 ant species were captured and identified from pitfall traps

Percentage of ant species captured in pitfall traps on Amyris elemifera and Zanthoxylum fagara. Overall, 243 total ants comprising 12 ant species were captured and identified from pitfall traps used in the canopy. The four most common ants collected were P. gracilis, C. planatus, Crematogaster ashmeadi, and C. floridanus. They represented 87% of all collected ants on A. elemifera and Z. fagara. Pseudomyrmex gracilis and C. planatus are exotic species and represented 56% of the total sampled ants
The frequency of P. gracilis individuals collected on three different groups of trees was compared using a generalized linear model (negative binomial with log link). This ant species was most frequent on A. elemifera (mean = 1.590, SD = 2.080), followed by Z. fagara (mean = 1.450, SD = 1.927), and then non-host trees (mean = 0.990, SD = 1.421). The omnibus test (compares the fitted model against the intercept only model) for the negative binomial model was not significant (χ2 = 4.291, df = 2, p value = 0.117; Table 3). However, individual parameters (tree groups) were investigated to determine significance using non-host trees as the baseline group to compare the frequency of P. gracilis collected for each tree group. Only P. gracilis collected on A. elemifera (the primary host tree for H. a. ponceanus) was significantly higher than non-host trees (χ2 = 4.119, df = 1, p value = 0.042; Table 3; Fig. 4).

Graphical representation of Pseudomyrmex gracilis collected on three different group of trees at Elliott and Adams Keys in Biscayne National Park, Florida. The bars represent averages of count data with standard error bars. Lowercase letters reflect statistical significance. Overall, P. gracilis was only significantly more frequent on Amyris elemifera than non-host trees. Amyris elemifera is the primary host tree for the Heraclides aristodemus ponceanus
Caterpillar predator exclusion experiment
Exclusion experiments revealed that early instar caterpillars were vulnerable to both crawling and non-crawling predators. Contingency table analysis revealed that survivorship, defined as caterpillar present on leaf, was significantly lower for early instar caterpillars in treatments with no barriers (18.8%; exact p value = 0.000003 < adjusted α-value = 0.004160), with tanglefoot only (31.3%; exact p value = 0.000465 < adjusted α-value = 0.004160), and for cages with holes and tanglefoot absent (37.5%; exact p value = 0.002700 < adjusted α-value = 0.004160), compared to the other barrier treatments (Table 4; Fig. 5). For the larger caterpillars, survivorship was only significantly lower when there were no barriers at all (33.3%; exact p value = 0.001374 < adjusted α-value = 0.004160; Table 4; Fig. 5).

The predator exclusion experiment involved 6 treatments, with physical barriers (tanglefoot and mesh cages) used in different combinations on Zanthoxylum fagara to compare caterpillar (early and late instar) survivorship against crawling and non-crawling predators. Survivorship was defined as persistence on leaf for 5 days. A contingency table analysis was performed. Asterisks denote significance in survivorship for the various treatments. Each treatment using early instar caterpillars had 16 trials, each treatment using late instar caterpillars had 15 trials. Refer to Table 4 for more details
Ant-caterpillar interactions
Pseudomyrmex gracilis (n = 30 trials; mean = 56.533, SD = 32.324, p < 0.001) discovered caterpillars significantly faster than did C. planatus (n = 31 trials; mean = 115.806, SD = 58.579), C. ashmeadi (n = 21 trials; mean = 123.524, SD = 64.697), and C. floridanus (n = 29 trials; mean = 122.690, SD = 87.015; Fig. 6). C. ashmeadi (n = 21 trials; mean = 0.429, SD = 0.148) and C. floridanus (n = 29 trials; mean = 0.345, SD = 0.614) occasionally recruited workers to combat caterpillars; P. gracilis and C. planatus did not recruit workers when interacting with caterpillars (Fig. 7). Overall, early instar caterpillars had higher rates of mortality compared to late instar caterpillars. Early instar caterpillars suffered the most damage when interacting with P. gracilis (n = 15 trials, 86.7% mortality); late instar caterpillars successfully foiled P. gracilis advances (n = 15 trials, 0% mortality). Early instar caterpillar mortality was moderately high with C. floridanus (n = 16 trials, 56.3% mortality); these were the only ant species tough enough to remove or force some late instar caterpillars to completely abandon the leaf (n = 15 trials, 40% mortality). Crematogaster ashmeadi attacked only a few early instar caterpillars during the trials (n = 10 trials, 20% mortality) and were ineffective in harming late instar caterpillars (n = 11 trials, 0% mortality). Finally, C. planatus demonstrated no aggressive encounters with either early (n = 16 trials, 0% mortality) or late instar (n = 15 trials, 0% mortality) caterpillars (Table 5).

The average time (±SE) the four most abundant ant species took to discover Heraclides cresphontes caterpillars placed on Amyris elemifera and Zanthoxylum fagara at Elliott Key, Biscayne National Park, Florida. A one-way ANOVA was performed followed by the Fisher’s Least Significant Difference post-hoc test. Asterisk denotes significance

Ant recruitment to Heraclides cresphontes caterpillars placed on Amyris elemifera and Zanthoxylum fagara by the four most abundant ant species at Elliott Key, Biscayne National Park, Florida. Recruitment is displayed as the average number of ants attracted to caterpillars (±SE). Ant recruitment allows ants to overwhelm larger prey such as late instar caterpillars
The Ant Danger Index ranked the predatory abilities of C. ashmeadi, C. planatus, C. floridanus, and P. gracilis against caterpillars that utilized A. elemifera and Z. fagara as host plants; since all four ant species represented 87% of the total sampled ants collected in canopies of A. elemifera and Z. fagara, only these ants were compared. Pseudomyrmex gracilis received the highest final score at 52, and C. floridanus had the next highest score at 40; both C. ashmeadi and C. planatus received lower scores of 24 (Table 6).
Discussion
Ants are important predators in tropical forest ecosystems (Jeanne 1979; Smiley 1985; Tobin 1995; Floren et al. 2002). Their abundance and active foraging day and night presents chronic passive and aggressive encounters with caterpillars (Lopez and Potter 2000; Seifert et al. 2016). Many factors, however, determine whether ant species will have beneficial, neutral, or negative effects toward caterpillars (Buckley 1987; Kaminski et al. 2010; Mestre et al. 2016). With the exception of those Lepidoptera whose caterpillars have mutualistic relationships with ants, early instar and small caterpillars are negatively associated with invertebrate predators, especially in subtropical and tropical ecosystems (Jeanne 1979; Montllor and Bernays 1993; Reavey 1993; Floren et al. 2002; Sam et al. 2015). Late instar and large caterpillars are negatively associated with birds especially in temperate ecosystems (Heinrich 1993; Reavey 1993; Remmel et al. 2009). Generalist and specialist parasitoids typically attack eggs, caterpillars and even the pupal stage (Weseloh 1993; Gentry and Dyer 2002; Stireman et al. 2009). We did not investigate either bird or parasitoid impacts during this study, which are other areas of research interest for future studies (Gentry and Dyer 2002; Timms et al. 2016).
Results from the predator exclusion experiment demonstrated that early instar caterpillars were less likely to survive when exposed to crawling and non-crawling predators (Fig. 5), but this conclusion was based on several assumptions. For example, caterpillar absence assumed caterpillar mortality, though caterpillars can drop off the leaf or migrate to another area if conditions are less than desirable (Castellanos et al. 2015). We accounted for this by scanning the entire tree after each trial to see if the caterpillar relocated to a different section on the tree. Early instar caterpillars experienced reduced survivorship in treatments when crawling insects gained access to them (no tanglefoot and holes in the cage), with the exception of tanglefoot present and no cage to protect the caterpillar. This outcome suggests that crawling predators such as ants, beetles, true bugs, spiders, and even lizards play a significant predator role. Late instar caterpillars had high survivorship for all treatments, except the treatment without barriers. As mentioned above, late instar caterpillar absence assumed caterpillar mortality, despite caveats including migration to pupate, foraging for more food resources, or leaf abandonment. Overall, results were consistent with past studies that concluded early instar (small) caterpillars suffered higher mortality than late instar (bigger) caterpillars (Tilman 1978; Watanabe 1981; Sam et al. 2015). The predator exclusion experiment elucidated coarse predator impacts (caterpillars present or absent) without defining specific causes (what agents were the cause of caterpillar mortality or disappearance). Since ants are ubiquitous in the forest in BNP, we assumed they would interact with caterpillars more frequently than other predators, though certainly other invertebrates (wasps, parasitoids, spiders), birds, and other vertebrates (lizards, treefrogs) may also play a role (Jeanne 1979; Sam et al. 2015; Seifert et al. 2016).
Ants, particularly P. gracilis, may pose a significant threat to butterfly eggs and larvae, but butterflies have developed ways to cope with such predators (Morais et al. 1999). Heraclides aristodemus ponceanus and other closely related swallowtails physically resemble lizard and bird droppings, an adaptation that may thwart vertebrate predation for predators that rely upon visual detection (Minno and Emmel 1992). Accordingly, invertebrate predators are more likely deterred by defensive tactics such as offensive chemicals stored in the caterpillars’ osmeteria (defensive organs that discharge a noxious odor), as well as physical head butts by the caterpillar to its attacker(s) (Fig. 8), and the most extreme response, leaf abandonment, where some caterpillars silk off (with a strand of silk that enables them to climb back up once the threat is gone) or directly drop off the leaf to escape predators (Honda 1983; Nafus 1993; Salazar and Whitman 2001; Gentry and Dyer 2002; Frankfater et al. 2009). All these defensive strategies may be less effective against aggressive, exotic ants (Forys et al. 2001; Lach et al. 2016). Introduced predators can overwhelm and eat their prey to extinction when the prey population reaches dangerously low numbers; predation effects are exacerbated when prey populations are constrained by other environmental factors such as habitat loss, inclement weather, climate change, and local anthropogenic impacts (Schoener et al. 2001; Lach et al. 2016). The most common ant on A. elemifera and Z. fagara in BNP was P. gracilis, which aggressively attacked and removed early instar caterpillars (86.7%; Table 5). Field observations revealed that sheer caterpillar size was a fair defense against these ants; in fact, head butting and everting osmeteria were also effective against more aggressive individual ants. As these experiments employed a surrogate species of caterpillars, H. cresphontes, we do not know how often ants of this species encounter H. a. ponceanus caterpillars in the field. Continued research should take steps to quantify those encounters.

Early instar Heraclides aristodemus ponceanus caterpillar fends off an approach by Camponotus planatus. This approach is not aggressive; however, the H. a. ponceanus caterpillar does not tolerate contact by other organisms
Camponotus floridanus ants did attack some late instar caterpillars (40%), but the attacks did not result in direct mortality. Harassed late instar caterpillars initially defended themselves, until more ants were recruited to attack. Eventually, the late instar caterpillar abandoned its leaf, falling to lower leaves or to the ground. Conversely, C. floridanus ants attacked and killed early instar caterpillars by simply carrying them away. Overall, late instar caterpillars are more equipped to defend themselves and avoid direct predation from these ants, the most common native ant species foraging on A. elemifera and Z. fagara.
Amyris elemifera should be inspected for the presence of extrafloral nectaries (EFNs) since P. gracilis was significantly more frequent on them compared to Z. fagara and non-host trees (Fig. 4). In the same plant family, Z. fagara does have minute EFNs (Koptur 1992), which might explain the higher frequency of P. gracilis on both A. elemifera and Z. fagara. Many non-host trees sampled at Elliott and Adams Keys also have EFNs, including blackbead (Pithecellobium keyense), catclaw blackbead (P. unguis-cati), and soldierwood (Colubrina elliptica), but presence of EFNs on trees does not solely determine whether ants will actively forage on them. The presence of exudate-producing insects can also encourage ants to occupy and forage on plants (Sendoya et al. 2016). This project did not record and quantify the presence and abundance of exudate producing insects, but future investigations should incorporate plant attractiveness to ants and other invertebrate predators due to intrinsic and extrinsic plant characteristics. Numerous studies conducted on ant attracting plants have demonstrated some caterpillar species are well-equipped to thwart ant attacks using behavioral, chemical, and physical means (Oliveira and Freitas 2004; Bächtold et al. 2012; Sendoya and Oliveira 2015).
Biscayne National Park and North Key Largo are the last remaining sanctuaries for the federally endangered H. a. ponceanus (Daniels 2014; Minno 2015); Biscayne National Park supports the largest population of H. a. ponceanus (Minno 2015). The Schaus swallowtail habitat enhancement project was initiated to bolster H. a. ponceanus populations on Elliott and Adams Key through planting thousands of native trees interspersed with nectar plants in the restored sites. Additional host plants, supplemented with captive-bred H. a. ponceanus reintroductions being conducted by scientists and staff at the University of Florida, present options for a better future for H. a. ponceanus in BNP (Daniels 2014).
Resource managers and scientists should continue to monitor H. a. ponceanus populations over time (Minno 2015). Long-term quantitative research (including video surveillance) may be necessary to assess potential risks imposed by exotic ants. High definition video surveillance cameras can capture predation events in natural settings day and night over an extended period of time to more accurately assess the effects of predator (particularly ants) interactions with all immature stages of H. a. ponceanus (Grieshop et al. 2012). In BNP, P. gracilis is the most potentially destructive force against Heraclides species on A. elemifera and Z. fagara. These negative impacts probably extend to other caterpillar species, since this exotic ant is omnipresent and an opportunistic forager (Wetterer 2010).
Different predators may be more or less deleterious at different life-history stages of butterflies (Nafus 1993; Seifert et al. 2015; Sam et al. 2015). Results obtained from these studies can guide future efforts to re-establish rare butterflies in areas they were once known to exist, by utilizing different techniques for protecting eggs and larvae of endangered butterflies at different life stages in reintroduction programs. A better understanding of which predators interact with butterfly eggs and larvae in natural habitats can help scientists determine suitable habitats for captive-reared butterflies before release. If exotic ants are found to be significantly deleterious to butterflies (Lach et al. 2016), then potential sites should be surveyed for exotic ants before release. Ant control measures could be taken, but in the past, chemical control of ants did more damage to the environment than to the targeted ant species (Summerlin et al. 1977; Williams et al. 2001; Oi et al. 2004; Tschinkel 2006; Plentovich et al. 2010). Future investigations will continue to document winners and losers as exotic and native species co-mingle in positive, neutral, and negative ways (Schoener et al. 2001; Allen et al. 2004; King and Tschinkel 2006; Didham et al. 2007; Moreau et al. 2014; Lach et al. 2016). Protecting pristine habitats and minimizing loss and degradation should be top priority to maintain rare, specialist organisms sensitive to anthropogenic-mediated actions.
References
Allen CR, Epperson DM, Garmestani AS (2004) Red imported fire ant impacts on wildlife: a decade of research. Am Midl Nat 152:88–103. doi:10.1674/0003-0031(2004)152[0088:RIFAIO]2.0.CO;2
Andersen AN, Hoffmann BD, Muller WJ, Griffiths AD (2002) Using ants as bioindicators in land management: simplifying assessment of ant community responses. J Appl Ecol 39:8–17. doi:10.1046/j.1365-2664.2002.00704.x
Bächtold A, Del-Claro K, Kaminski LA, Freitas AVL, Oliveira PS (2012) Natural history of an ant-plant-butterfly interaction in a Neotropical savanna. J Nat Hist 46:943–954. doi:10.1080/00222933.2011.651649
Beuzelin JM, Reagan TE, Akbar W, Cormier HJ, Flanagan JW, Blouin DC (2009) Impact of Hurricane Rita storm surge on sugarcane borer (Lepidoptera: Crambidae) management in Louisiana. J Econ Entomol 102:1054–1061
Brown WL (2000) Diversity of ants. In: Agosti D, Majer JD, Alonso LE, Schultz TR (eds) Ants: standard methods for measuring and monitoring biodiversity. Smithsonian Institution Press, Washington, DC, pp 45–79
Buckley RC (1987) Interactions involving plants, Homoptera, and ants. Annu Rev Ecol Syst 18:111–135. doi:10.1146/annurev.ecolsys.18.1.111
Buren WF, Allen GE, Whitcomb WH, Lennartz FE, Williams RN (1974) Zoogeography of the imported fire ants. J N Y Entomol Soc 82:113–124
Calcaterra L, Cabrera S, Briano J (2016) Local co-occurrence of several highly invasive ants in their native range: are they all ecologically dominate species? Insectes Soc 63:407–419. doi:10.1007/s00040-016-0481-3
Carroll CR, Janzen DH (1973) Ecology of foraging by ants. Annu Rev Ecol Syst 4:231–257. doi:10.1146/annurev.es.04.110173.001311
Castellanos I, Barbosa P, Zuria I, Caldas A (2015) Quantifying insect predation with predator exclusion cages: the role of prey antipredator behavior as a source of bias. Entomol Exp Appl 157:360–364. doi:10.1111/eea.12375
Cuautle M, Rico-Gray V (2003) The effect of wasps and ants on the reproductive success of the extrafloral nectaried plant Turnera ulmifolia (Turneraceae). Funct Ecol 17:417–423. doi:10.1046/j.1365-2435.2003.00732.x
Daniels JC (2014) Conservation matters: status and conservation of the federally endangered Schaus swallowtail butterfly. News of the Lepidopterists’. Society 56:138–139
Dennis RL, Hodgson JG, Grenyer R, Shreeve TG, Roy DB (2004) Host plants and butterfly biology. Do host-plant strategies drive butterfly status? Ecol Entomol 29:12–26. doi:10.1111/j.1365-2311.2004.00572.x
Deyrup M (2016) Ants of Florida: identification and natural history. CRC Press, Boca Raton
Deyrup M, Davis L, Cover S (2000) Exotic ants in Florida. Trans Am Entomol Soc 126:293–326
Didham RK, Tylianakis JM, Gemmell NJ, Rand TA, Ewers RM (2007) Interactive effects of habitat modification and species invasion on native species decline. Trends Ecol Evol 22:489–496. doi:10.1016/j.tree.2007.07.001
Emmel TC, Minno MC, Nation JL Jr, Eliazar PJ, Cotter AJ, Bustamante EG, Ritland DB, Goode MR (1988) Habitat requirements and status of the endemic Schaus’ swallowtail in the Florida Keys. Final Project Report, Project Number GFC-86-023. Submitted to Florida Game and Fresh Water Fish Commission Division of Wildlife Nongame Wildlife Section, Tallahassee, Florida
Floren A, Biun A, Linsenmair KE (2002) Arboreal ants as key predators in tropical lowland rainforest trees. Oecologia 131:137–144. doi:10.1007/s00442-002-0874-z
Forys EA, Quistorff A, Allen CR (2001) Potential fire ant (Hymenoptera: Formicidae) impact on the endangered Schaus swallowtail (Lepidoptera: Papilionidae). Fla Entomol 84:254–258. doi:10.2307/3496176
Forys EA, Allen CR, Wojcik DP (2002) Influence of the proximity and amount of human development and roads on the occurrence of the red imported fire ant in the lower Florida Keys. Biol Conserv 108:27–33. doi:10.1016/S0006-3207(02)00086-1
Frankfater C, Tellez MR, Slattery M (2009) The scent of alarm: ontogenetic and genetic variation in the osmeterial gland chemistry of Papilio glaucus (Papilionidae) caterpillars. Chemoecology 19:81–96. doi:10.1007/s00049-009-0013-y
Gentry GL, Dyer LA (2002) On the conditional, nature of neotropical caterpillar defenses against their natural enemies. Ecology 83:3108–3119
Grieshop MJ, Werling B, Buehrer K, Perrone J, Isaacs R, Landis D (2012) Big brother is watching: studying insect predation in the age of digital surveillance. Am Entomol 58:172–182
Heinrich B (1993) How avian predators constrain caterpillar foraging. In: Stamp NE, Casey TM (eds) Caterpillars ecological and evolutionary constraints on foraging. Chapman and Hall, Inc., New York, pp 224–247
Holway DA, Lach L, Suarez AV, Tsutsui ND, Case TJ (2002) The causes and consequences of ant invasions. Annu Rev Ecol Syst 33:181–233. doi:10.1146/annurev.ecolysis.33.010802.150444
Honda K (1983) Defensive potential of components of the larval osmeterial secretion of papilionid butterflies against ants. Physiol Entomol 8:173–179. doi:10.1111/j.1365-3032.1983.tb00346.x
IBM SPSS Statistics 22 (2013) Statistical Package for the social sciences version 22. http://www.ibm.com/analytics/us/en/technology/spss/.
Jameson A (2002) Host plants and habitats of the Schaus swallowtail butterfly (Papilio aristodemus ponceanus). MSc Thesis. University of Miami
Jeanne RL (1979) A latitudinal gradient in rates of ant predation. Ecology 60:1211–1224. doi:10.2307/1936968
Kaminski LA, Freitas AVL, Oliveira PS (2010) Interaction between mutualisms: ant-tended butterflies exploit enemy-free space provided by ant-treehopper associations. Am Nat 176:322–334. doi:10.1086/655427
King JR, Tschinkel WR (2006) Experimental evidence that the introduced fire ant, Solenopsis invicta, does not competitively suppress co-occurring ants in a disturbed habitat. J Anim Ecol 75:1370–1378. doi:10.1111/j.1365-2656.2006.01161.x
Koptur S (1992) Plants with extrafloral nectaries and ants in Everglades habitats. Fla Entomol 75:38–50. doi:10.2307/3495479
Koptur S, Rico-Gray V, Palacios-Rios M (1998) Ant protection of the nectaried fern Polypodium plebeium in central Mexico. Am J Bot 85:736–739. doi:10.2307/2446544
Krushelnycky PD, Holway DA, Lebrun EG (2010) Invasion processes and causes of success. In: Lach L, Parr CL, Abbott KL (eds) Ant ecology. Oxford University Press, Oxford, pp 245–260
Lach L, Hooper-Bùi LM (2010) Consequences of ant invasions. In: Lach L, Parr CL, Abbott KL (eds) Ant ecology. Oxford University Press, New York, pp 261–286
Lach L, Volp TM, Greenwood TA, Rose A (2016) High invasive ant activity drives predation of a native butterfly larva. Biotropica 48:146–149. doi:10.1111/btp.12284
Loftus W, Kushlan J (1984) Population fluctuations of the Schaus swallowtail (Lepidoptera: Papilionidae) on the islands of Biscayne Bay, Florida, with comments on the Bahaman swallowtail. Fla Entomol 67:277–287. doi:10.2307/3493950
Lopez R, Potter DA (2000) Ant predation on eggs and larvae of the black cutworm (Lepidoptera: Noctuidae) and Japanese beetle (Coleoptera: Scarabaeidae) in turfgrass. Environ Entomol 29:116–125. doi:10.1603/0046-225X-29.1.116
Lowe S, Browne M, Boudjelas S, De Poorter M (2000) 100 of the world’s worst invasive alien species: a selection from the global invasive species database. Invasive Species Specialist Group, Auckland
Lubertazzi D, Tschinkel WR (2003) Ant community change across a ground vegetation gradient in north Florida’s longleaf pine flatwoods. J Insect Sci 3:1–17. doi:10.1093/jis/3.1.21
MacGowan JC, Hill JG (2010) Two new exotic pest ants, Pseudomyrmex gracilis and Monomorium floricola (Hymenoptera: Formicidae) collected in Mississippi. Midsouth Entomol 3:106–109
Mack MC, D’Antonio CM (1998) Impacts of biological invasions on disturbance regimes. Trends Ecol Evol 13:195–198. doi:10.1016/S0169-5347(97)01286-X
Mathew G, Anto M (2007) In situ conservation of butterflies through establishment of butterfly gardens: a case study at Peechi, Kerala, India. Curr Sci 93:337–347
McGlynn TP (1999) The worldwide transport of ants: geographic distribution and ecological invasions. J Biogeogr 26:535–548. doi:10.1046/j.1365-2699.1999.00310.x
Mestre L, Piñol J, Barrientos JA, Espadaler X (2016) Differential ant exclusion from canopies shows contrasting top-down effects on community structure. Oecologia 180:190–203. doi:10.1007/s00442-015-3442-z
Minno MC (2015) 2015 Schaus’ swallowtail (Heraclides aristodemus ponceanus) monitoring on Key Largo, Florida. South Lepidopterists News 37:106–107
Minno MC, Emmel TC (1992) Larval protective coloration in swallowtails from the Florida Keys. Trop Lepidoptera 3:47–49
Minno MC, Emmel TC (1993) Butterflies of the Florida Keys. Scientific Publishers, Gainesville
Montllor CB, Bernays EA (1993) Invertebrate predators and caterpillar foraging. In: Stamp NE, Casey TM (eds) Caterpillars ecological and evolutionary constraints on foraging. Chapman and Hall, Inc., New York, pp 170–202
Morais HR, Diniz IR, Silva D (1999) Caterpillar seasonality in a central Brazilian Cerrado. Rev Biol Trop 47:1025–1033
Moreau CS, Deyrup MA, Davis LR (2014) Ants of the Florida Keys: species accounts, biogeography, and conservation (Hymenoptera: Formicidae). J Insect Sci 14:295. doi:10.1093/jisesa/ieu157
Nafus DM (1993) Movement of introduced biological-control agents onto nontarget butterflies, Hypolimnas spp. (Lepidoptera, Nymphalidae). Environ Entomol 22:265–272
Oi DH, Watson CA, Williams DF (2004) Monitoring and management of red imported fire ants in a tropical fish farm. Fla Entomol 87:522–527. doi:10.1653/0015-4040(2004)087[0522:MAMORI]2.0.CO;2
Oliveira PS, Freitas AVL (2004) Ant-plant-herbivore interactions in the neotropical cerrado savanna. Naturwissenschaften 91:557–570. doi:10.1007/s00114-004-0585-x
Oliveira PS, Oliveira-Filho AT, Cintra R (1987) Ant foraging on ant-inhabited Triplaris (Polygonaceae) in western Brazil: a field experiment using live termite-baits. J Trop Ecol 3:193–200
Plentovich S, Swenson C, Reimer NJ, Richardson M, Garon N (2010) The effects hydramethylnon on the tropical fire ants (Solenopsis geminata) and non-target arthropods on Spit Island, Midway Atoll. J Insect Conserv 14:459–465. doi:10.1007/s10841-010-9274-6
Porter SD, Savignano DA (1990) Invasion of polygyne fire ants decimates native ants and disrupts arthropod community. Ecology 71:2095–2106. doi:10.2307/1938623
Possley J, Hodges S, Magnaghi E, Maschinski J (2016) Distribution of Croton linearis in Miami-Dade County preserves with potential for supporting the federally endangered butterflies Strymon acis bartrami and Anaea troglodyta floridalis. Nat Area J 36:81–87
Reavey D (1993) Why body size matters to caterpillars. In: Stamp NE, Casey TM (eds) Caterpillars ecological and evolutionary constraints on foraging. Chapman and Hall, Inc., New York, pp 248–278
Remmel T, Tammaru T, Magi M (2009) Seasonal mortality trends in tree-feeding insects: a field experiment. Ecol Entomol 34:98–106. doi:10.1111/j.1365-2311.2008.01044.x
Ribas CR, Schoereder JH, Pic M, Soares SM (2003) Tree heterogeneity, resource availability, and larger scale processes regulating arboreal ant species richness. Aust Ecol 28:305–314. doi:10.1046/j.1442-9993.2003.01290.x
Saarinen EV, Daniels JC (2006) Miami blue butterfly larvae (Lepidoptera: Lycaenidae) and ants (Hymenoptera: Formicidae): new information on the symbionts of an endangered taxon. Fla Entomol 89:69–74. doi:10.1653/0015-4040(2006)89[69:MBBLLL]2.0.CO;2
Salazar JC, Whitman DC (2001) Defensive tactics of caterpillars against predators and parasitoids. In: Ananthakrishnan TN (ed) Insects and plant defence dynamics. Science Publishers, Inc., Plymouth, pp 161–207
Salvato M (2008) Investigator’s annual report to Biscayne National Park: Annual butterfly survey on Elliott Key for the North American Butterfly Association. Fish and Wildlife Service, Vero Beach, FL.
Sam K, Remmel T, Molleman F (2015) Material affects attack rates on dummy caterpillars in tropical forest where arthropod predators dominate: an experiment using clay and dough dummies with green colourants on various plant species. Entomol Exp Appl 157:317–324. doi:10.1111/eea.12367
Schoener TW, Spiller DA, Losos JB (2001) Predators increase the risk of catastrophic extinction of prey populations. Nature 412:183–186. doi:10.1038/35084071
Seifert CL, Lehner L, Adams M, Fiedler K (2015) Predation on artificial caterpillars is higher in countryside than near-natural forest habitat in lowland south-western Costa Rica. J Trop Ecol 31:281–284. doi:10.1017/S0266467415000012
Seifert CL, Schulze CH, Dreschke CT, Frötscher H, Fiedler K (2016) Day vs. night predation on artificial caterpillars in primary rainforest habitats—an experimental approach. Entomol Exp Appl 158:54–59. doi:10.1111/eea.12379
Sendoya SF, Oliveira PS (2015) Ant–caterpillar antagonism at the community level: Interhabitat variation of tritrophic interactions in a neotropical savanna. J Anim Ecol 84:442–452. doi:10.1111/1365-2656.12286
Sendoya SF, Blüthgen N, Tamashiro JY, Fernandez F, Oliveira PS (2016) Foliage-dwelling ants in a neotropical savanna: effects of plant and insect exudates on ant communities. Arthropod-Plant Interact 10:183–195. doi:10.1007/s11829-016-9423-2
Smiley JT (1985) Heliconius caterpillar mortality during establishment on plants with and without attending ants. Ecology 66:845–849. doi:10.2307/1940546
Snyder JR, Herndon A, Robertson WBJ (1990) South Florida Rockland. In: Myers RL, Ewel JJ (eds) Ecosystems of Florida. University Press of Florida, Gainesville, pp 230–277
Stireman JO, Greeney HF, Dyer LA (2009) Species richness and host associations of Lepidoptera-attacking Tachinidae in the northeast Ecuadorian Andes. J Insect Sci 9:1–19
Summerlin JC, Hung ACF, Vinson SB (1977) Residues in nontarget ants, species simplification and recovery of populations following aerial applications of mirex. Environ Entomol 6:193–197
Tilman D (1978) Cherries, ants and tent caterpillars: timing of nectar production in relation to susceptibility of caterpillars to ant predation. Ecology 59:686–692. doi:10.2307/1938771
Timms LL, Schwarzfeld M, Saaksjarvi IE (2016) Extending understanding of latitudinal patterns in parasitoid wasp diversity. Insect Conserv Divers 9:74–86. doi:10.1111/icad.12144
Tobin JE (1995) Ecology and diversity of tropical forest canopy ants. In: Lowman MD, Nadkarni NM (eds) Forest canopies. Academic Press, New York, pp 129–147
Tschinkel WR (1988) Distribution of the fire ants Solenopsis invicta and S. geminata (Hymenoptera: Formicidae) in northern Florida in relation to habitat and disturbance. Ann Entomol Soc Am 81:76–81
Tschinkel WR (2006) The fire ants. The Belknap Press of Harvard University Press, Massachusetts
United States Fish and Wildlife Service (FWS) (2008) Schaus swallowtail butterfly (Heraclides aristodemus ponceanus) 5-Year review: Summary and evaluation. http://ecos.fws.gov/docs/five_year_review/doc1983.pdf. Accessed 11 Nov 2016.
Vitousek PM, D’Antonio CM, Loope LL, Westbrooks R (1996) Biological invasions as global environmental change. Am Sci 84:468–478
Wagner DL, Van Driesche RG (2010) Threats posed to rare and endangered insects by invasions of nonnative species. Annu Rev Entomol 55:547–568. doi:10.1146/annurev-ento-112408-085516
Wang CL, Strazanac J, Butler L (2001) A comparison of pitfall traps for studying leaf litter ant communities. J Econ Entomol 94:761–765. doi:10.1603/0022-0493-94.3.761
Watanabe M (1981) Population dynamics of the swallowtail butterfly, Papilio xuthus L., in a deforested area. Res. Popul Ecol 23:74–93. doi:10.1007/BF02514094
Weseloh RM (1993) Potential effects of parasitoids on the evolution of caterpillar foraging behavior. In: Stamp NE, Casey TM (eds) Caterpillars ecological and evolutionary constraints on foraging. Chapman and Hall, Inc., New York, pp 203–223
Wetterer JK (2010) Worldwide spread of the graceful twig ant, Pseudomyrmex gracilis (Hymenoptera: Formicidae). Fla Entomol 93:535–540. doi:10.1653/024.093.0410
Wetterer JK, Porter SD (2003) The little fire ant, Wasmannia auropunctata: distribution, impact, and control. Sociobiology 42:1–41
Whelan KRT (2011) Schaus swallowtail butterfly (Heraclides aristodemus ponceanus) habitat enhancement in Biscayne National Park. Project Report. Funded by FWS South Florida Coastal Program. National Park Service, Palmetto Bay, Florida
Whelan KRT, Atkinson A (2015) Schaus Swallowtail Butterfly (Heraclides aristodemus ponceanus) Habitat Enhancement in Biscayne National Park. South Florida Coastal Program Project Progress Report. NPS/SFCN. National Park Service, Palmetto Bay, Florida
Whelan KRT, Ruiz PL, Shamblin RB, Houle PA, Ross MS, Atkinson AJ, Patterson JM, Alonso J (2013) Biscayne National Park Vegetation Map Project. Natural Resource Technical Report. NPS/SFCN/NRTR-2013/774. National Park Service, Fort Collins, Colorado
Whitcomb WH, Denmark HA, Buren WF, Carroll JF (1972) Habits and present distribution in Florida of the exotic ant, Pseudomyrmex mexicanus (Hymenoptera: Formicidae). Fla Entomol 55:31–33. doi:10.2307/3493638
Williams DF (1994) Biology, impact, and control of introduced species. Westview Press, Boulder
Williams DF, Collins HL, Oi DH (2001) The red imported fire ant (Hymenoptera: Formicidae): an historical perspective of treatment programs and the development of chemical baits for control. Am Entomol 47:146–159
Zettler JA, Taylor MD, Allen CR, Spira TP (2004) Consequences of forest clear-cuts for native and non-indigenous ants (Hymenoptera: Formicidae). Ann Entomol Soc Am 97:513–518. doi:10.1603/0013-8746(2004)097[0513:COFCFN]2.0.CO;2
Acknowledgements
We thank Vicente Martinez, Shane Harrington, and Steve Newman for their support and transportation to the islands. The support and cooperation of Biscayne National Park and the National Park Service staff were essential in project development to completion. We greatly appreciate field assistance from many, including Leslie Bayas, Lydia Cuni, Darlene Darrican, Michael Foguer, Tanjim Hossain, Eduardo Ibarra, Craig Perry, Brenda Riveria, Andrea Salas, Adriana Samani, and Yadira Reynaldo. Identification of difficult ant specimens was provided by Mark Deyrup. Paulo Oliveira provided invaluable information and guidance on the ant-caterpillar interaction study. Constructive comments on the manuscript were made by Ian Jones and Scott Zona. Permission to work in Biscayne National Park was granted through the scientific research and collecting permit # BISC – 2014 – SCI – 0031. This is contribution #344 to the Tropical Biology Program at Florida International University.
Funding
This study was financially supported through the Zoo Miami Conservation and Research Grant and the Kelly Foundation Botany Scholarship.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Data Availability
The data are in the process of being archived in Dryad Digital Respository.
Rights and permissions
About this article

Cite this article
Clayborn, J., Koptur, S. Mortal combat between ants and caterpillars: an ominous threat to the endangered Schaus swallowtail butterfly (Heraclides aristodemus ponceanus) in the Florida Keys, USA. J Insect Conserv 21, 689–702 (2017). https://doi.org/10.1007/s10841-017-0012-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-017-0012-1