
Abstract
Using targeted survey, excavation, and radiocarbon dating, we assess the extent to which human settlement patterns on California’s northern Channel Islands fit predictions arising from the ideal free distribution (IFD): (1) people first established and expanded permanent settlements in the regions ranked high for environmental resource suitability; (2) as population grew, they settled in progressively lower ranked habitats; and (3) changes in the archaeological record associated with high population levels such as increases in faunal diversity and evenness in high-ranked habitats are coincident with the expansion to other areas. On Santa Rosa Island, the early permanent settlements were located in both high- and middle-ranked locations, with the most extensive settlement at the highest ranked locations and only isolated sites elsewhere. Settlement at a low-ranked habitat was confined to the late Holocene (after 3600 cal BP). Drought influenced the relative rank of different locations, which is an example of climate adding a temporal dimension to the model that episodically stimulated population movement and habitat abandonment. Because the IFD includes a wide range of cultural and environmental variables, it has the potential to be a central model for guiding archaeological analysis and targeted field research.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The settlement patterns of mobile hunter-gatherer populations can include ephemeral camps, seasonal residential bases, logistical sites and, in some cases, permanent settlement sites occupied annually or continuously (e.g., Kelly 2013). The location of larger sites with permanent or semipermanent occupation usually takes into account a suite of environmental and socioeconomic factors. The ideal free distribution or IFD (Åström 1994; Fretwell and Lucas 1969; Fretwell 1972; Sutherland 1983, 1996; Treganza 1995) is used in behavioral ecology and increasingly in anthropology and archaeology to understand how such factors affect human settlement and broader patterns of decision-making and culture change (e.g., Kennett 2005; Kennett et al. 2006, 2009; Kennett and Winterhalder 2008; McClure et al. 2009; Winterhalder et al. 2010; Culleton 2012; O’Connell and Allen 2012; Jazwa et al. 2013; Codding and Jones 2013; Giovas and Fitzpatrick 2014; Moritz et al. 2014; Codding and Bird 2015).
The IFD uses habitat suitability measures to generate predictions about regional population movement and distribution. It can be used to understand settlement patterns on both large (e.g., Fitzhugh and Kennett 2010; Allen and O’Connell 2008) and small (e.g., Kennett et al. 2006; Culleton 2012) scales. It is flexible enough to address questions about where people settled as they entered and expanded throughout California (Codding and Jones 2013) and which drainage on a small island people would choose to establish individual sites (Winterhalder et al. 2010).
The model is constructed by calculating the relative suitability of different habitats based on the spatial distribution of available environmental resources and cultural factors that influence their value. People should first settle the habitat with the highest overall base suitability. As population density increases, resource exploitation and interference competition cause a decrease in the effective suitability of that habitat, eventually making it advantageous to expand into and settle lower ranked habitats. A variety of more specific predictions and implications follow from this pattern (see Kennett et al. 2009; Winterhalder et al. 2010; Jazwa et al. 2013). In a variant of the IFD, the ideal despotic distribution (IDD), the inhabitants of a high-ranked habitat defend a disproportionate share of resources and impede in-migration, causing expansion of the overall population to more marginal habitats earlier than predicted by the IFD (Summers 2005; Kennett and Winterhalder 2008; Kennett et al. 2009, 2013; Culleton 2012; Bell and Winterhalder 2014).
We use the IFD and IDD to aid understanding of human settlement on Santa Rosa Island, one of California’s four northern Channel Islands (NCI), located off the coast of Santa Barbara (Fig. 1). The NCI are particularly well suited to employ the IFD. First, there is a comprehensive archaeological record available for the islands that includes an existing database of radiocarbon dates and diagnostic artifacts, allowing a multi-island perspective that would not be feasible in a less-studied area (see Kennett 2005; Rick et al. 2005; Jazwa and Perry 2013). Second, the limited array of settlement options on the NCI allows for a model that is more inclusive of all potential settlement locations. Third, the islands have a relatively limited and thus manageable number of natural resources, facilitating accurate assessment of suitability. Finally, the islands were visited and used for thousands of years before permanent settlement (see Erlandson et al. 2011). When chosen, settlement locations were selected with a great deal of local environmental experience and knowledge, increasing the likelihood that people made economically informed choices.

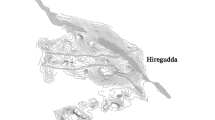
Santa Rosa Island. The 19 major drainages are outlined and coastal sites indicated. Drainages with excavated habitats are highlighted in gray. Sites and drainages mentioned in the text are labeled
One of us (CSJ) surveyed and excavated coastal sites at four differently ranked drainage systems on Santa Rosa Island in 2012, 2013, and 2014: Soledad and Dry Canyons (high-ranked), Cow and Lobos Canyons (middle-ranked, adjacent to high-ranked areas), La Jolla Vieja Canyon (middle-ranked, isolated), and Bee Canyon (low-ranked), using the suitability rankings from Winterhalder et al. (2010). Based on these data, we assess a variety of hypotheses and find that the results largely conform to IFD predictions.
California’s Northern Channel Islands
People visited and seasonally exploited environmental resources available on the NCI by at least 13,000 years ago (Erlandson et al. 2007, 2008, 2011; Johnson et al. 2002; Kennett 2005; Kennett et al. 2008). Sea levels were lower than at present (∼70–75 m at 13,000 years ago) and the four islands were all connected to form one landmass, Santarosae (Kennett et al. 2008; Watts et al. 2011; Reeder-Myers et al. 2015), but they were never connected to the mainland. With eustatic sea level rise, the islands had become separated by the time of the earliest evidence for their permanent settlement at approximately 8000 years ago (Winterhalder et al. 2010). This rise in sea level caused a 65 % decrease in land area and may have submerged evidence of early permanent settlement (Kennett et al. 2008). Thus, it is unclear whether early ephemeral sites on the island were culturally associated with submerged coastal sites or with more permanent sites on the mainland (Kennett 2005). Even if some early settlement sites have been lost, we can use the IFD to assess decisions about settlement location and persistence subsequent to the rise in sea level. Any patterns could then guide a search for submerged sites.
The productive environment of the Santa Barbara Channel region has prompted an emphasis among researchers on marine resources (e.g., Kennett 2005; Rick et al. 2005; Erlandson et al. 2007, 2008, 2011). The earliest known occupants of these islands targeted fish, shellfish, and sea mammals such as pinnipeds and dolphins. During the middle Holocene (7550–3600 cal BP), a distinctive settlement system included large coastal settlement sites at the mouths of major streams, interior residential bases, and special purpose sites for collecting and shucking abalones (Clifford 2001; Kennett and Clifford 2004; Jazwa et al. 2015a). This system was primarily concentrated on the resource-rich north coast of Santa Rosa Island, with the exception of a series of abalone shucking sites on the west coast.
Throughout the Santa Barbara Channel region, there was a general increase in fishing and sea mammal hunting through time, which was accompanied at many sites by a decrease in shellfish collecting. This trend is particularly pronounced on the islands (Arnold 1992a; Braje et al. 2007; Colten 2001; Glassow 1977, 1993; Jazwa et al. 2012, 2013; Jazwa and Perry 2013; Kennett 2005; Kennett and Kennett 2000; Kennett and Conlee 2002; Raab et al. 1995; Rick 2007; Rick et al. 2008). Fish and sea mammals became particularly important during the late Holocene (after 3600 cal BP). Changes in diet like these can result from multiple causes, including resource depression, environmental change, sociocultural change, technological developments, and economic change. For example, a decrease in the size and abundance of high-ranked dietary items, coupled with an increase in dietary diversity, can reflect resource depression; opposite trends would be associated with resource recovery (see Braje et al. 2007; Jazwa et al. 2012 for NCI examples; more generally, see Bettinger 1991; Nagaoka 2002; Codding and Bird 2015; Singh and McKechnie 2015).
The relationship between environmental and sociopolitical change has been an important topic of research on the NCI (Arnold 1992a, 2001a; Arnold and Graesch 2004; Arnold and Tissot 1993; Braje et al. 2007; Erlandson and Jones 2002; Jazwa et al. 2012; Jazwa and Perry 2013; Kennett and Kennett 2000; Kennett et al. 2007, 2008; Raab and Larson 1997; Glassow et al. 2012). In parallel, there have been several attempts to generate high-resolution records for sea surface temperature (Arnold and Tissot 1993; Kennett 2005; Kennett and Kennett 2000; Pisias 1978) and drought (Raab and Larson 1997) in the Santa Barbara Channel region.
During the late Holocene, there were important environmental, technological, economic, and sociopolitical changes on the NCI (see Rick et al. 2005; Glassow et al. 2010). More intensive predation on fish and sea mammal populations during the late Holocene may be related in part to the development of the single-piece fishhook by 2500 cal BP (Strudwick 1986; Rick et al. 2002) and ocean going plank canoes (tomol) after 1500 cal BP. The latter provided a more reliable form of marine transportation than was available previously. It allowed the occupants of the Santa Barbara Channel region to travel more quickly and farther from shore with larger amounts of cargo, and it provided a more stable platform for fishing and sea mammal hunting (Arnold 1992a, 1995, 2001a; Gamble 2002; Fagan 2004). Beginning around 1300 cal BP, the rate of population increase on the NCI accelerated and the number of permanent settlements increased dramatically (Arnold 2001a; Kennett 2005; Kennett and Conlee 2002; Winterhalder et al. 2010). The first signs of institutionalized status differences appeared after this time (e.g., Arnold 1992a, 2001a; Kennett et al. 2009).
The Middle to Late Period Transition (MLT; 800–650 cal BP) has also been associated with important changes (Arnold 1991, 1992a, 1997, 2001b; Arnold and Tissot 1993; Arnold et al. 1997; Jazwa et al. 2012; Jazwa and Perry 2013; Kennett 2005; Kennett and Conlee 2002; Perry and Jazwa 2010; Raab and Larson 1997). For example, the Callianax biplicata (purple olive) shell bead industry grew significantly, providing an important trade good (Arnold 1987, 1990, 1992a, b, 2001a; Arnold and Munns 1994; Munns and Arnold 2002, pp. 132–133; Kennett 2005; King 1990; Rick 2007). Many of the changes of this period can be related to environmental stress (Arnold 1991, 1992a, 1997, 2001b; Arnold and Tissot 1993; Arnold et al. 1997; Colten 2001; Jazwa et al. 2012; Jones et al. 1999; Jones and Schwitalla 2008; Lambert and Walker 1991; Lambert 1993, 1997; Kennett 2005; Kennett and Conlee 2002; Kennett and Kennett 2000; Kennett et al. 2007, 2008; Pisias 1978; Pletka 2001; Raab and Larson 1997; Stine 1994; Yatsko 2000).
The MLT occurred during a period of extreme droughts associated with the Medieval Climatic Anomaly (1150–600 cal BP; Kennett 2005; Jones et al. 1999; Jones and Schwitalla 2008; Raab and Larson 1997; Stine 1994; Yatsko 2000) that were felt throughout the American Southwest (Jones et al. 1999; Jones and Schwitalla 2008; Bocinsky and Kohler 2014). Osteological data from the Santa Barbara Channel region reveal peaks in health problems and violence (Lambert and Walker 1991; Walker 1989; Walker and Lambert 1989; Lambert 1993) associated with increasing sedentism and diminishing supplies of fresh water and terrestrial foods. Kennett (2005; also Jones and Schwitalla 2008) extends the period of drought and violence further back in time to 1500 cal BP. Population aggregation around wetter, more resource-rich areas may have been a stimulus for some of the changes that occurred during this period. However, there are relatively few well-documented and precisely dated sites that have been associated with the MLT (e.g., Arnold 1991, 1992a, b; Raab and Larson 1997; Yatsko 2000; Munns and Arnold 2002; Perry 2003; Kennett 2005; Rick et al. 2005; Rick 2007; Glassow et al. 2010; Jazwa et al. 2012).
The Channel Islands and the IFD
Kennett et al. (2009) generated an initial IFD model for the NCI, ranking suitability of potential coastal settlement locations based on weighted assessments of (1) length of accessible rocky intertidal coastline, (2) length of accessible sandy beach, (3) area of kelp forests within a 2-km radius, and (4) fresh water availability, estimated by watershed size. Watersheds were chosen as a unit of analysis for reasons given in Kennett (2005) and Kennett et al. (2009). Fresh water availability is watershed dependent (Jazwa et al. 2015b; Winterhalder et al. 2010), and large primary NCI settlements are almost all along the coast at the mouths of major drainages (Kennett et al. 2009; Winterhalder et al. 2010; Jazwa et al. 2013). These suitability rankings were then compared with archaeological evidence for age of first permanent settlement and the persistence of use. However, evidence for environmental change, population change within sites, and human impacts on suitability (i.e., cultural and technological changes) was not considered.
In a follow-up analysis that uses settlement order to assess environmental suitability, Winterhalder et al. (2010) found that watershed size, used as a proxy for fresh water availability, was the most important predictor of initial settlement and persistence. Kelp forest area and sandy beach length became more important through time, consistent with the increasing reliance on fish and sea mammals, the development of the tomol, and the expansion of the C. biplicata shell bead industry. In a second follow-up, Jazwa et al. (2013) examined archaeological collections from sites at the mouth of Old Ranch Canyon in eastern Santa Rosa Island to assess consequences for IFD predictions associated with closure of an estuary and the technological developments of the late Holocene. Both of these temporal changes in suitability are reflected in the archaeological assemblage from the site in a manner consistent with IFD predictions.
Here, we further adapt application of the IFD model using a regimen of targeted survey, excavation, and analysis of shell midden collections. We define permanent settlements as having substantial residential middens, cemeteries, or houses (Winterhalder et al. 2010), but do not require that these sites be inhabited year-round or without short hiatuses. Occupation of these sites indicates persistent settlement in a given location. We use watersheds as our unit of analysis, but refine the estimates of fresh water flow and drainage resilience using hydrological simulations (Jazwa et al. 2015b) to assess the assumption by Winterhalder et al. (2010) that watershed size is a good proxy for fresh water availability. We use weights of environmental contributions to suitability similar to those of Kennett et al. (2009). We assign weighted z-scores for drainage size (10 %), water flow (30 %), drainage resilience (10 %; percent water flow loss from a median to a dry century), length of rocky intertidal shoreline (30 %), length of sandy beach (5 %), and area of kelp beds (15 %; Table 1).
Model Expectations
Model predictions are similar to those outlined by Kennett et al. (2009), Winterhalder et al. (2010), and Jazwa et al. (2013): (1) the highest ranked locations should contain the earliest evidence for permanent settlement; (2) over time there should be evidence for resource depression at these locations, including decreases in the abundance and perhaps size of high-ranked resources along with evidence for dietary diversification to include lower ranked resources; (3) dates of settlement for lower ranked locations should coincide with periods of resource depression in nearby higher ranked locations; and (4) settlement at the highest ranked locations should persist even as settlement expands to other sites.
Two further predictions arise from the observation that habitat suitability can be affected by factors not directly related to population density: (5) extrinsic environmental change can depress suitability (Braje et al. 2007; Jazwa et al. 2012) and (6) technological developments, such as the ocean-going plank canoe, can influence habitat suitability and settlement rankings (Jazwa et al. 2013). Prediction (6) may not be entirely density independent if, for instance, economic development is initiated in response to larger populations.
More specifically, we predict that the high-ranked habitat of Dry and Soledad Canyons should be settled first (prediction 1). The faunal record at this habitat should reflect increasing population density over time, increasing shellfish diversity, the incorporation into the diet of lower ranked shellfish species other than Mytilus californianus and Haliotis spp. (see Braje et al. 2007: Table 1 for shellfish rankings and their justification) and evidence of an expansion of fishing and sea mammal hunting (prediction 2). Settlement should expand from the high-ranked habitat progressively to lower ranked Cow/Lobos, La Jolla Vieja, and Bee Canyons (prediction 3). These locations should then experience a similar pattern of resource depression as population density increases. Higher ranked drainages should exhibit evidence of resource depression before lower ranked drainages. Once settled, occupation is persistent unless environmental, cultural, or technological factors alter suitability sufficiently that it falls below the equilibrium level (predictions 4–6).
Methods
Strategic Approach
Previous IFD studies on the NCI compared regional settlement patterns to model predictions (see Kennett et al. 2009; Winterhalder et al. 2010). Here we focus specifically on four locations on Santa Rosa Island: the high-ranked Dry and Soledad Canyons, middle-ranked locations adjacent to high-ranked ones (Cow and Lobos Canyons), and two middle-low-ranked habitats (La Jolla Vieja and Bee Canyons), which invert their relative position depending on the ranking system (compare Table 1 with Kennett et al. 2009; Winterhalder et al. 2010). The topography and geology of Santa Rosa Island combined with resource-rich nearshore kelp forests ensure that even the lowest ranked habitat on Santa Rosa is 28th out of the 46 ranked habitats across all four islands (Winterhalder et al. 2010; Table 1).
We surveyed interior drainages to complement the excavation work done at the drainage mouths where permanent settlements typically are located. During the summers of 2012, 2013, and 2014, CSJ led a survey of the drainage bottoms and walls of Soledad, Dry, Cow, Lobos, Bee, Acapulco, and Tecolote Canyons. La Jolla Vieja Canyon had previously been surveyed as part of a National Park Service cultural resources project in 1995. The team recorded all observed sites in these drainages, covering 100 % of the survey area. Survey crews walked 10-m transect lines in all flat areas (<20° slope) of the drainage bottoms, performing light surface scrapes at 10-m intervals to clear vegetation and look for surface exposures of sites. All erosional banks along stream cuts and all accessible rockshelters along the drainages were inspected. Because of erosion and vegetation, sites in canyon bottoms were most readily discovered on the floors of rockshelters and the walls of cut banks.
Excavation targeted drainage mouths because they feature the most extensive and persistent permanent settlements on the NCI (Kennett 2005; Kennett et al. 2009; Winterhalder et al. 2010). During the summer of 2013, CSJ and his field crew excavated 25-cm × 25-cm column samples from sites within each of the four primary habitats included in this study (Table 2). All were from low coastal terraces, locations protected from the ocean tides and floods in the adjacent canyons but offering easy access to the coast and its marine resources. Nearly all large NCI coastal sites are found on these landforms. The number of samples from each site was set based on site size and number of distinct components. At least one column sample was placed in each locus of larger sites to better represent the range of occupation at each of the four locations. The 25 × 25-cm size of the column samples accommodates the directive of the National Park Service for preservation of cultural and natural resources and allows us to generate faunal assemblages from a broad variety of sites. Samples were excavated from eroding banks to minimize impacts.
From Dry and Soledad Canyons, we excavated three units from CA-SRI-19 and one from CA-SRI-821. The mouths of these two drainages are approximately 300 m apart; the preferred landscape for habitation is entirely encompassed by these two sites. CA-SRI-19 is a very large site (∼56,000 m2) and has evidence for several distinct phases of occupation. From Cow and Lobos Canyons, we excavated two units from CA-SRI-115 and one unit from CA-SRI-541. These two drainages, the mouths of which are approximately 700 m apart, also have a densely occupied cultural landscape connecting them. In 1996, Kennett and Morris excavated one 25 × 25-cm column sample from CA-SRI-116, which we incorporate into this analysis. From La Jolla Vieja Canyon, we excavated two units from CA-SRI-138 and one unit from CA-SRI-131. And from Bee Canyon, we excavated two units from CA-SRI-313 and one unit from CA-SRI-333. In 1996, Kennett and Morris excavated two 25 × 25-cm column samples from CA-SRI-31, which we also include (Kennett 1998). This approach includes samples from both the large, primary settlement sites in each of these drainages and the smaller adjacent sites. Together, they provide a more complete perspective of persistent occupation in each location. Our discussion of settlement focuses on the sites that are the best candidates for permanent habitation at each location (Table 2).
Chronology
We obtained at least two radiocarbon dates from each excavated column sample except for the thin deposit at CA-SRI-31, unit 2. We selected samples from the top and bottom strata with datable material for each unit to represent the beginning and end of occupation of that unit. We later dated intermediate levels from those units with a long occupation history, targeting strata with evidence for change in the faunal record. Column samples were placed strategically to try to encompass the full range of occupation at each site, increasing our confidence that the dates generated approximate initial and terminal occupation of each habitat. Kennett (1998, pp. 456, 458) obtained two standard radiometric dates from CA-SRI-116 and one each from CA-31 units 1 and 2 from Beta Analytic, Inc. From 2012 to 2014, we obtained 32 additional AMS radiocarbon dates to establish the chronology of the excavated sites and another 17 dates from other coastal locations and interior drainage bottoms (Tables 3 and 4; Fig. 2). All samples were tested at the Keck Carbon Cycle AMS Facility at the University of California, Irvine.

We calibrated all of the dates in OxCal 4.1 (Bronk Ramsey 2009) using the most recent marine calibration curve, Marine13 (Reimer et al. 2013). We used a variable ∆R value for the Santa Barbara Channel region for samples with dates of less than 2600 14C BP (Tables 3 and 4; Brendan Culleton, personal communication, 2015; Hendy et al. 2013) and an updated ∆R value for the Santa Barbara Channel region for older samples (261 ± 21 14C year; Brendan Culleton, personal communication 2012; Jazwa et al. 2012). Application of the Bayesian statistical model in OxCal further constrained the error ranges on dates based on the relative stratigraphic position of the radiocarbon samples within each of the units.
Faunal Analysis
We excavated eroding natural exposures from the surface to the base of the deposit in arbitrary 10-cm levels, with materials from different stratigraphic levels kept separate. All excavated materials were size sorted using 1/2-, 1/4-, and 1/8-in. mesh and identified at the Penn State University Zooarchaeology Lab. The lead author (CSJ) and trained undergraduate and graduate students sorted all shellfish, other faunal constituents, and cultural materials (see Jazwa et al. 2012, 2013) for methodology. Meat weights were calculated for dietary constituents using the multipliers compiled and summarized by Rick (2004, p. 79). Data are combined from individual excavation units for intersite comparisons. In units with deep deposits and clear changes in the faunal record, we separated the data into two distinct cultural components between the strata that best exemplified this change. Shell and meat weight data are normalized to a volume of 1 m3 for comparison.
We analyze shellfish and other faunal materials by weight rather than minimum number of individuals because of the fragmentary nature of the midden and the difficulty of assigning meaningful counts to the species present in low densities (see Glassow 2000; Giovas 2009; contra Mason et al. 1998, 2000). We used the Shannon-Weaver function (Shannon and Weaver 1949) to calculate the taxonomic diversity of the assemblages from both shell and meat weights of all dietary shellfish species recovered. We also calculated shellfish species richness and evenness for both shell and meat weights. In all calculations, abalone and turban snail fragments that were indistinguishable to species (i.e., Haliotis spp., Chlorostoma spp.) were apportioned to species by weight using the relative weights of individual species (red and black abalone, brown and black turban snails) for each unit or deposit.
Lastly, we estimated the size of M. californianus shells using the equation relating umbo thickness to shell length presented in Singh and McKechnie (2015; Singh et al. 2015; see also Campbell and Braje 2015):
Singh and McKechnie demonstrate that this method has an R 2 value of 0.80 and average error of 10.2 ± 1.9 mm. The only site for which this method was not employed was CA-SRI-31. These assemblages are not currently available and mussel sizes from this site were estimated using the template from White (1989). We do not include mussels less than 20 mm in length in average mussel size because we assume that they were too small to be targeted for consumption, and were instead inadvertently collected because they were attached to larger individuals. A high proportion of these small individuals could reflect mussel collection through stripping rather than plucking (see Bettinger et al. 1997).
Results
Radiocarbon Chronology
The sites excavated and reported encompass the entire known range of permanent occupation on the NCI from 7545–7415 cal BP (CA-SRI-138, unit 1, 73 cm below surface) through 315–140 cal BP (CA-SRI-333, unit 1, 5 cmbs; Table 3; Fig. 2). The highest ranked drainages along the northwest coast are the location of the earliest intensive permanent occupation on Santa Rosa Island. Once settled, it appears that these habitats were occupied consistently throughout the Holocene. The earliest available evidence for persistent settlement anywhere on the island is at the mouth of Tecolote Canyon, dating to 7890–7520 cal BP (Kennett 1998, p. 456; Winterhalder et al. 2010; Jazwa 2015). During our survey, we recorded a midden lens at CA-SRI-780 that we date to 8410–8285 2σ cal BP on the flat canyon bottom approximately 1 km from the drainage mouth (Table 4). It is difficult to determine whether these deposits could be associated with permanent occupation because they were only visible in a small sinkhole. There is also evidence for early settlement at the high-ranked habitat at the mouth of Old Ranch and Old Ranch House Canyons dating to 8050–7840 cal BP, but this habitat was apparently depopulated following the closure of a productive estuary there (Kennett 1998; Jazwa et al. 2013).
The earliest date from an excavation unit was from unit 1 of CA-SRI-138 (7545–7415 2σ cal BP; 73 cmbs). This site is located at the mouth of La Jolla Vieja Canyon, a middle-low-ranked habitat. One of the deepest levels of CA-SRI-116, in a middle-ranked habitat, Lobos and Cow Canyons, dates to 6380–6020 cal BP. The earliest date from the highest ranked habitat in this study, 5945–5760 cal BP, comes from 77.5 cmbs at unit 1 of CA-SRI-821 at the mouth of Dry Canyon (Fig. 2). The early dates from the other habitats appear not to be consistent with the predictions of the IFD (Table 5), a point that we take up in the discussion.
At the mouth of Dry and Soledad Canyons, there is evidence that the large site of CA-SRI-19 was occupied from 5425–5420 2σ cal BP through at least 1260–1090 2σ cal BP. Between that site and CA-SRI-821, the entire coastal plain within this habitat was occupied throughout much of the middle Holocene, like the nearby high-ranked habitat at the mouth of Tecolote and Arlington Canyons (CA-SRI-3, CA-SRI-4, CA-SRI-5, and CA-SRI-6) (Kennett 1998, pp. 456–457; Winterhalder et al. 2010). The radiocarbon record also shows that the mouths of Cow and Lobos Canyons likely were occupied nearly continuously from their initial occupation at CA-SRI-116 through at least 730–545 2σ cal BP (CA-SRI-541, 8 cmbs). However, there is no evidence of more than one site being inhabited contemporaneously until both CA-SRI-542 (2575–2325 2σ cal BP, 40–50 cmbs) and CA-SRI-115, unit 1 (2545–2315 2σ cal BP, 57 cmbs), were occupied.
La Jolla Vieja Canyon is ranked lower in the IFD and it is on the south coast of the island, isolated from the higher ranked drainages. The initial occupation at this habitat (CA-SRI-138, unit 1) is earlier than expected (7545–7415 2σ cal BP), but there is no evidence of any other coastal occupation of the area until 3585–3405 2σ cal BP (CA-SRI-131, 88 cmbs). It is unclear how continuously CA-SRI-138 was occupied. The latest date from unit 1 is 4845–4655 2σ cal BP, but it is from 49 cmbs. There is a major stratigraphic change immediately above where the sample was collected, but the sample from 9 cmbs (5890–5725 2σ cal BP) is anomalously early and was likely eroded from uphill. Therefore, it is unclear how late in time the upper strata at this unit continued to be occupied, and whether it overlapped with the lowest strata at CA-SRI-131. Beginning with the occupation of CA-SRI-131, there is evidence for continuous occupation at the mouth of La Jolla Vieja Canyon at least until 645–530 2σ cal BP (CA-SRI-138, unit 2, 14 cmbs). The rockshelter/valley bottom site CA-SRI-147 in the interior of La Jolla Vieja Canyon, more than 2 km from the coast, has evidence for intensive occupation from 7300 to 350 cal BP (Braje et al. 2007). This suggests that permanent occupation of this drainage was not isolated to a single site.
The habitat with the latest evidence for initial permanent settlement is Bee Canyon. The earliest dates from permanent settlement sites are from CA-SRI-31, unit 2 (10–25 cmbs; 3240–2790 2σ cal BP), and CA-SRI-309 (65 cmbs; 2825–2695 2σ cal BP). After these dates, the next evidence for occupation of this habitat is not until 1680–1440 2σ cal BP (CA-SRI-31, unit 1, 28–40 cmbs). Beginning around this date, the mouth of Bee Canyon was occupied until historic contact (CA-SRI-333, 5 cmbs). Despite the fact that Bee Canyon does not have evidence of permanent occupation until relatively late, people were there for short periods of time during the middle Holocene. CA-SRI-338 is a special purpose red abalone shucking site dating to ∼5900–5700 cal BP (Jazwa et al. 2015a). Several other similar red abalone middens are located farther to the south near the mouth of Acapulco Canyon. There is also an early date from ephemeral deposits from CA-SRI-331 (8925–8625 2σ cal BP). Rick et al. (2013) noted the presence of several nonpermanent Paleocoastal (before 9000 cal BP) sites along the higher marine terrace on the west coast of the island.
Although middle Holocene sites are present along interior ridgelines (Kennett 1998, 2005; Jazwa et al. 2015a), most evidence from the bottom of drainages suggests recent occupation. There are two notable exceptions. There are thin lenses of red abalone shell in erosional banks in Tecolote Canyon, one of which we have dated to 7385–7225 2σ cal BP (CA-SRI-778). These may be secondary deposits eroded into the drainage. CA-SRI-147, the rockshelter/valley bottom site in La Jolla Vieja Canyon, has evidence for occupation from approximately 7300 to 350 cal BP (Braje et al. 2007). Most other rockshelters have initial dates of occupation during the late Holocene, within the last 1200 years (Table 4). All radiocarbon samples from these sites were taken from the deepest stratum exposed by erosion, in most cases directly overlying sterile soil. The earliest dates are from CA-SRI-786 (1140–915 cal BP) near the head of Tecolote Canyon and CA-SRI-770 (1160–935 cal BP), approximately 2 km from the mouth of Cow Canyon. Other rockshelters from throughout Cow and Lobos Canyon all date to the Late Period (650–168 cal BP), when people were clustered in large coastal villages and rockshelters were largely used as temporary camps to access interior resources (Kennett 2005).
Excavation Results
As discussed earlier, consistent with data from elsewhere on the NCI, shellfish is the dominant component prior to the late Holocene, when fish and sea mammal increased in importance. This is clearly evident in the middle and late Holocene deposits from the mouth of Dry and Soledad Canyons (Table 6; Fig. 3). The middle Holocene faunal record from both CA-SRI-821 and CA-SRI-19, unit 2, are strongly dominated by shellfish, 67 % and 66 %, respectively. Fish (20 %) is the second largest contributor at CA-SRI-821 and sea mammal (13 %) is third. This is reversed at CA-SRI-19, at which sea mammal (30 %) is more prominent than fish (3 %). During the late Holocene, fish and sea mammal are more prominent. At CA-SRI-19, unit 1, fish (47 %) overtakes shellfish (38 %) in importance. In the assemblage from CA-SRI-19, unit 3, both sea mammal (48 %) and fish (33 %) were more prominent contributors to meat weight than shellfish (19 %). This unit also had the greatest amount of meat weight represented by the assemblage, at 270.1 kg/m3. CA-SRI-19, unit 2, is second with 196.2 kg/m3.

a–d Meat weight density of the different faunal components of each excavation unit, separated by habitat
Similar patterns are evident in the other habitats. At both Cow/Lobos and La Jolla Vieja Canyons, shellfish are the dominant meat weight component of all excavation units. Both CA-SRI-116 and CA-SRI-115, unit 2, are more than 80 % shellfish; fish is 10 % and 12 %, respectively. During the late Holocene, fish become slightly more important, at 17 % early in this period (CA-SRI-115, unit 1) and during the MLT and Late Period (CA-SRI-541). Sea mammal also increases in importance through time. At La Jolla Vieja Canyon, the dominance of shellfish during the middle Holocene is even more prominent, accounting for more than 93 % of the meat weight throughout CA-SRI-138, unit 1. Toward the end of the middle Holocene, this drops to 64 % (CA-SRI-131, 60–95 cmbs), with fish increasing to 26 %. During the late Holocene, fish become much more important, making up 45 % of the meat weight from CA-SRI-138, unit 2, 40–77 cmbs (vs. 51 % for shellfish). During the Late Middle Period and MLT (CA-SRI-138, unit 2, 0–40 cmbs), fish (5 %) decreases in importance compared to shellfish (63 %) and sea mammal (32 %). The mouth of Bee Canyon was not settled until the late Holocene, and fish is the dominant contributor to meat weight at all times (between 48 % and 61 %). Shellfish is second (between 36 % and 45 %), and sea mammal only accounts for more than 7 % of the total in one unit (CA-SRI-31, unit 1; 19 %).
Overall, meat weight density from excavation units in all habitats generally tends to increase through time. The exception is Cow/Lobos Canyons, where CA-SRI-116 is the earliest unit and has the highest total meat weight (222 kg/m3). In all other cases, the latest excavation units have the highest meat weight density, perhaps suggesting increased population density in all habitats through time.
The relative proportions of shellfish species in the faunal record from different excavation units are more strongly associated with their particular drainages than with the age of the site (Table 7; Fig. 4). California mussel (M. californianus) is the primary species in all excavation units except CA-SRI-333, but its prominence varies between drainages (Table 8). With the exception of the later deposits at Bee Canyon, the highest shellfish diversity is at the mouth of Dry and Soledad Canyons (Table 9; Fig. 5). It is consistent in these units over time, with a Shannon-Weaver diversity index between 1.7 and 1.9 (Shannon and Weaver 1949). Diversity of meat weight is higher, ranging from 1.9 to 2.1. This is because mussel makes up a high proportion of the meat weight but has a relatively low ratio of meat to shell. Evenness values are also among the highest at Dry and Soledad Canyons (Fig. 6). During the middle Holocene, the second most prominent contributor to the shellfish assemblage is black turban snails (Chlorostoma funebralis), which is 26 % of the shell at CA-SRI-19, unit 2, and 14.5 % of the shell at CA-SRI-821. This is followed by abalone (Haliotis cracherodii and Haliotis rufescens), chiton (Ischnochiton conspicuus and Cryptochiton stelleri), and crab (Cancer spp.).

a–d Shell weight density of the seven most important dietary components, separated by habitat

a–d Shannon-Weaver diversity function for each site examined in this study, divided by habitat

a–d Evenness values for each site examined in this study, separated by habitat
During the late Holocene, chiton and crab are the second and third most prominent shell species. However, the shell meat weight densities at CA-SRI-19, unit 1 (25 kg/m3) and unit 3 (50.1 kg/m3) are substantially less than the middle Holocene unit 2 (128.9 kg/m3). This reflects the increasing importance of fish and sea mammal. From the perspective of the IFD, it makes sense that there is high species diversity at the highest ranked habitat. Population increase there caused a depression of the highest ranked species (California mussel; see Braje et al. 2007), eventually prompting a population expansion to lower ranked habitats. This is also reflected in average mussel size data (Table 8, Fig. 7). There is a slight increase in mussel size throughout the middle Holocene, but there is a clear decrease in size during the late Holocene, suggesting overexploitation. The high average from CA-SRI-19, unit 1, is from only 22 individuals and therefore is not a reliable indicator.

a–d Average mussel size and percent of small (<20 mm) mussels, separated by habitat. Note that the y-axis represents both average mussel length (mm) and percent of small mussels (%)
The middle-ranked habitat at Cow and Lobos Canyons has the lowest species diversity overall, with a Shannon-Weaver diversity index between 0.7 and 1.1 for shell weight and 1.1 and 1.6 for meat weight. Evenness is also lower here overall than at any of the other habitats. This is because California mussel contributes more than 73 % of all shell assemblages. The only other species to make up significant proportions of the assemblage were turban snails (9 % of CA-SRI-116) and sea urchin (Strongylocentrotus purpuratus; 13 % of CA-SRI-541). The highest density shell deposits occur at CA-SRI-116 (187.4 kg/m3), with the second highest at the MLT/Late Period deposits from CA-SRI-541 (157.6 kg/m3). Mussel size is relatively consistent over time, with a slight decrease during the late Holocene. There is a relatively high proportion of small individuals at CA-SRI-115, unit 1, suggesting that people may have increasingly employed stripping as a strategy as the overall availability of large mussels decreased.
The shell deposits from La Jolla Vieja Canyon exhibit a similar pattern. Diversity is low overall, although higher than at Cow and Lobos Canyons, with a Shannon-Weaver index ranging from 0.7 to 1.4 for shell weight and 1.4 to 1.8 for meat weight. California mussel is again the dominant component, at least 54 % of the shell weight in all cultural components of the excavation units. Abalone is more important early in time, particularly in the middle Holocene deposits from CA-SRI-138, unit 1 (8 %, 50–85 cmbs; 6 %, 0–50 cmbs), as are sea urchin (8 %, 50–85 cmbs; 7 %, 0–50 cmbs) and turban snails (6 %, 50–85 cmbs; 8 %, 0–50 cmbs). There is a general trend toward decreasing diversity and evenness through time. The Late Middle Period/MLT deposits from CA-SRI-138, unit 2, 10–40 cmbs, are more than 84 % California mussel.
One possible explanation for this pattern is that it was coincident with the establishment of the nearby site of CA-SRI-130, which has high-density midden deposits and clear house depressions (Kennett 1998). CA-SRI-138 may have become a specialized mussel collection and shucking site; many of the shells there were unbroken. The decreased diversity and increased focus on mussels over time at the expense of other shellfish species in these habitats could be a result of increased interest in collecting fish and sea mammals in the overall diet breadth. Rather than collecting a suite of shellfish, occupants of the region at the mouth of La Jolla Vieja Canyon may have focused only on mussels to complement the fish in their diet. This part of the island has the clearest pattern in mussel size over time, with a gradual decrease in size and proportion of small individuals, suggesting a similar pattern to Cow and Lobos Canyon. CA-SRI-138, unit 2, also has by far the greatest density of mussel MNI, further supporting the interpretation of overpredation of mussels (Table 8).
The shellfish assemblages from Bee Canyon are among the most diverse overall, with a Shannon-Weaver index ranging from 1.6 to 1.9 for shell weight and 1.9 to 2.2 for meat weight. These values increase over time. Values for evenness are similarly high. Overall, Bee Canyon is the drainage in which California mussel is the least important component. Sea urchin is one of the dominant species in all units except CA-SRI-31, unit 2. In the other units, it is between 12 % (CA-SRI-313, unit 2) and 33 % (CA-SRI-333) of the total. Turban snails, chiton, and barnacles (Balanus spp.) are also important contributors to the assemblage. Overall, shellfish meat weight is lower than at either Cow/Lobos or La Jolla Vieja Canyons. Occupation of these sites occurred only during the late Holocene when fishing and sea mammal hunting were more important than earlier. The increased diversity of shellfish over time may be related to an increased human population density. Mussel size is largest at Bee Canyon of all areas that we tested, which may be related to the fact that it was settled for only a short time. CA-SRI-31, unit 1, has unusually small mussels and a high proportion of small individuals, which may reflect overpredation, but could also be because sizes were estimated using a less reliable method than the other units. By the beginning of the MLT, there were permanent settlements at the mouths of all of the major drainages on the island, limiting the possibilities for expansion to new settlement locations. Because opportunities for further expansion would have been limited, it would have been more difficult for population movement to alleviate resource stress.
Discussion
Kennett et al. (2009) and Winterhalder et al. (2010) have shown that the existing radiocarbon chronology for permanent coastal settlement across the four NCI supports IFD predictions of initial occupation and persistence. Jazwa et al. (2013) added a temporal dimension by demonstrating how suitability rises or falls with environmental or technological change independent of population density. Fieldwork reported in this study goes a step further, targeting not only the chronology of initial occupation of different habitats but also the timing of changes in the faunal record as a proxy for population density and changing habitat suitability (Fig. 8).

Idealized summary of settlement patterns at the excavated habitats through time. Settlement intensity is inferred from the number and size of occupied sites. Shellfish diversity is used to determine resource stress. The increase in fishing suggests that population at individual sites is larger
According to the IFD, the earliest permanent settlement should occur at the highest ranked locales. The earliest dates available from permanent settlements on Santa Rosa are from Old Ranch and Tecolote Canyons, both high-ranked habitats. CA-SRI-187, at the mouth of Old Ranch Canyon and Old Ranch House Canyons, has been dated to 8050–7840 cal BP (Kennett 1998; Jazwa et al. 2013). The suitability of this site was increased by the presence of a productive estuary at the mouth of those drainages during the middle Holocene (Jazwa et al. 2013). CA-SRI-3, at the mouth of Tecolote Canyon, dates to 7890–7520 cal BP (Kennett 1998, p. 456; Winterhalder et al. 2010). It is uncertain if the midden lens at nearby (∼1 km up canyon) CA-SRI-780, dated to 8410–8285 2σ cal BP, can push this date further back in time because only a small part of the site is exposed and it is unclear whether it meets the criteria of a permanent settlement. However, permanent early settlements on the island are not confined to the highest ranking habitats, but also can be associated with middle-ranked habitats like Lobos Canyon (CA-SRI-116) and La Jolla Vieja Canyon (CA-SRI-138, unit 2).
Expansion of settlement sites during the middle Holocene is consistent with the predictions of the IFD. High-ranked habitats along the northwest coast of Santa Rosa Island are the first to experience clear evidence of more intensive occupation (prediction 1; Table 10). There is an important difference between these habitats and the others with early settlement (Fig. 2). In the high-ranking habitats, there are multiple locales that were occupied contemporaneously. At Dry and Soledad Canyons, this includes units at CA-SRI-19 and CA-SRI-821. Similarly, CA-SRI-3 and CA-SRI-5 at the mouth of Tecolote and Arlington Canyons were both occupied during the middle Holocene (Kennett 1998; Winterhalder et al. 2010; Jazwa et al. 2015a). Intensive settlement persists at both of these locations through the late Holocene (prediction 4). There is a continuous record of occupation through the middle Holocene at the nearby middle-ranked habitat of Cow and Lobos Canyon, but there is no evidence of contemporaneous occupation of multiple sites in this habitat until the late Holocene. At La Jolla Vieja Canyon, evidence for permanent coastal settlement is sparse and limited to a single locus of CA-SRI-138 until the very end of the middle Holocene (prediction 3).
Dry conditions during the middle Holocene enhanced the suitability of drainages along the northwest coast relative to other locales. These drainages are among the most resilient to drought on the island (prediction 5). Climate records suggest that the middle Holocene, particularly the period from about 6500 to 3800 cal BP, was a dry interval dominated by drought conditions (e.g., Antevs 1948, 1952, 1955; LaMarche 1973, 1974; Lindström 1990; Morgan et al. 1991; Thompson 1992; Cole and Liu 2004; Quade et al. 1998; Hughes and Graumlich 2000; Benson et al. 2002; Erlandson et al. 2005; Kennett 2005; Kennett et al. 2007). To examine the local effects of this climate change, Jazwa et al. (2015b) modeled fresh water flow and drainage resilience at the mouth of the 19 major drainages on Santa Rosa Island (Table 1). We use these results to generate several different sets of drainage rankings by altering the weights given to the different environmental variables (Table 11). Across our weightings, Tecolote and Arlington Canyons are always the top two ranked drainages. Soledad and Dry Canyons fall by 7 and 5 ranking spots, respectively, when water is ignored and gain 2 and 5 spots when taking only water into account. This reflects the important contribution of water availability to habitat suitability along the northwest coast.
At the start of the late Holocene, permanent settlement density increased at locations that had previously been occupied and settlement expanded to new drainages (predictions 3 and 5; Fig. 8). Increasingly wet conditions in California at the end of the middle Holocene (Antevs 1955; Lindström 1990; Davis 1992; Thompson 1992; Quade et al. 1998; Benson et al. 2002; Kennett 2005; Kennett et al. 2007) increased the suitability of multiple drainages, including Bee Canyon. Climate change lowered the premium on drought-resistant drainages. Dry and Soledad Canyons maintained a high settlement density at this time, as did the other high-ranked drainages along the northwest coast of the island like Tecolote and Arlington Canyons (Fig. 2; Kennett 1998). Apparently, continuous occupation persists at Cow and Lobos Canyons through the middle and late Holocene.
There was a clear increase in settlement at La Jolla Vieja Canyon, with evidence for contemporaneous occupation at CA-SRI-131, CA-SRI-138, and CA-SRI-488 during the late Holocene. This matches IFD predictions (4), as increasing population led to an increase in population density at all occupied habitats. The late Holocene is also when lower ranked or more isolated habitats like Bee Canyon first experienced permanent settlement. Jazwa et al. (2013) show that Old Ranch Canyon was reoccupied at the end of the middle Holocene following a gap of at least 1500 years after the closure of the estuary that was present there. This pattern is consistent with the model discussed here. After about 1300 cal BP, the number of settlement sites on the NCI reaches its maximum and there is evidence for permanent settlement at all of the major drainages on Santa Rosa Island (Winterhalder et al. 2010), including all four of the locations investigated here. The population expansion that occurred during this period makes this pattern consistent with the IFD.
There are two periods during which the settlement chronology of Santa Rosa Island does not fully conform to IFD predictions: initial permanent occupation (contra prediction 1) and just before historic contact (contra prediction 4). The available evidence for earliest permanent settlement is not confined to the highest ranked drainages, but is distributed throughout high- and middle-ranked habitats. Excavation units from middle-ranked habitats examined here predate the earliest deposits from high-ranked Dry and Soledad Canyons (CA-SRI-821) by approximately 1000–1500 years (La Jolla Vieja) and 100–500 years (Lobos). We propose several potential explanations. First, at especially low overall population densities, any of the better locations on Santa Rosa Island would have provided adequate resources, allowing sociocultural factors to play a larger role in the population distribution. Occupants of the high-ranked northwest coast may have acted despotically, dissuading nearby settlement and prompting new groups to locate on the south side of the island or along the east coast. Likewise, factors like weather could have drawn small populations to the south coast of the island, which is sunnier and more protected from wind. It also is possible that this anomaly is an artifact of rising sea levels, which may have obscured the evidence of earliest occupation in the highest ranking habitats. If that is the case, the truncated evidence available to us at present will conform to a pattern of contemporaneous settlement across the upper end of the suitability range, similar to what we see (Fig. 9). Unfortunately, we do not currently have evidence to sort out these possibilities.

Potential effect of sea level rise on evidence for the IFD. We show idealized dates for initial settlement as would be predicted by the IFD. Habitats are settled in order of suitability and settlement persists through time. However, if sea level rise submerges evidence for early coastal settlement at the mouths of the highest ranked drainages, all habitats with submerged cultural components will appear to have their initial evidence of occupation at approximately the same time (triangles) and across the upper end range of suitabilities, not only in the highest ranked drainages
In the second apparent exception to the IFD, despotism is more clearly present during the latest occupation on the islands, particularly during and after the MLT. Despite the fact that population continued to increase (Glassow 1999; Kennett 2005) after the MLT, permanent settlement condenses to a smaller number of large villages rather than continuing to expand to new settlement locations. By Spanish contact, residents of Santa Rosa Island occupied only eight large coastal villages (Johnson 1982, 1993, 2001; Kennett 1998, 2005; Glassow et al. 2010). There is no evidence for occupation of the mouth of Dry and Soledad Canyons after 1260–1090 2σ cal BP (CA-SRI-19), La Jolla Vieja Canyon after 645–530 2σ cal BP (CA-SRI-138, unit 2), and Cow and Lobos Canyons after 730–545 2σ cal BP (CA-SRI-541). In fact, the only site examined intensively in this study with clear evidence of occupation until historic contact is CA-SRI-333, which is relatively small and perhaps subsidiary to the large village of Nawani (CA-SRI-97) at the mouth of Acapulco Canyon to the south (Jazwa 2015). The Late Period population aggregated into larger villages partially in response to environmental stress and conflict (e.g., Walker 1989; Walker and Lambert 1989; Lambert and Walker 1991; Lambert 1993, 1997; Raab and Larson 1997; Jones et al. 1999; Kennett and Kennett 2000; Kennett 2005; Kennett et al. 2013). Associated changes included the increase of institutionalized differences in social status (Kennett 2005; Kennett et al. 2009) and more rigid territorial boundaries (Kennett 2005; Jazwa 2015).
The end of the Middle Period and the MLT are coincident with the Medieval Climatic Anomaly, a period of extreme drought conditions throughout the western USA (1150–600 cal BP; Kennett 2005; Jones et al. 1999; Jones and Schwitalla 2008; Raab and Larson 1997; Stine 1994; Yatsko 2000). In the Santa Barbara Channel region, Lambert and Walker (1991; Walker 1989; Walker and Lambert 1989; Lambert 1993, 1997) document declining health and increased violence based on skeletal samples from the MLT, which they associate with increasing sedentism and diminishing supplies of fresh water and terrestrial foods. Resource shortfalls could have prompted people to move to villages at the mouths of larger, more reliable watersheds. These locations were also typically near large, protected sandy beaches where plank canoes could be launched, increasing access to offshore resources like sea mammals and kelp forest fish species. These canoes provided greater connectivity with the mainland, allowing people to more efficiently obtain subsistence and nonsubsistence resources. Sandy beaches are also the habitat of C. biplicata snails, the shells of which were used to create shell beads traded for external resources (prediction 6; e.g., Arnold 1987, 1990, 1992a, b, 2001a; Arnold and Munns 1994; Munns and Arnold 2002; Kennett 2005; King 1990; Rick 2007).
The settlement pattern that emerged during the Late Period is consistent with an IDD at the regional scale. Despotic behavior by groups (i.e., territoriality imposed by violence or the threat of violence) may have emptied locations that were otherwise suitable, high-ranked habitats like Dry and Soledad Canyon. Within these territories, a more centralized, defendable village was established at the highest ranked location, as predicted by the IFD. On Santa Rosa, these locations were often positioned on promontories along the coastline that offered clear views to monitor approaching tomols or potential attackers. Population aggregation is also consistent with the idea that increased conflict led to buffer zones between villages (Kennett 2005).
Implications of the Faunal Record
The faunal record from Santa Rosa Island is also consistent with the IFD and IDD (Fig. 8). The highest diversity of middle Holocene shellfish in our excavation units comes from the highest ranked habitat at Dry and Soledad Canyons (CA-SRI-821 and CA-SRI-19, unit 2). We hypothesize that this represents the incorporation of more low-ranked shellfish species like crab, chiton, and platform mussels (Septifer bifurcatus) into the diet (see Braje et al. 2007). This is a manifestation of resource depression and decrease in the basic habitat suitability that comes with increased population density (prediction 2). Dry and Soledad Canyons are especially attractive because of their relatively abundant and dependable water sources. Water is sufficiently important that it might have held increasing population there despite depressed shellfish resources, delaying expansion into lower ranked habitats.
The middle Holocene occupation at the mouth of Cow/Lobos and La Jolla Vieja Canyons appears to be different from that at Dry/Soledad Canyons. These drainages do not have the same fresh water flow nor are they as resilient to drought as the high-ranking drainages along the northwest coast (Jazwa et al. 2015b). The middle Holocene occupation at Cow/Lobos Canyons is more extensive than that at La Jolla Vieja Canyon, perhaps because of their relative locations. Cow and Lobos Canyons are along the north coast of the island, adjacent (along with Cañada Verde) to high-ranking drainages. This offered benefits including the potential for social or marriage relationships with nearby communities. La Jolla Vieja is more isolated and without such benefits. The shellfish assemblages from the middle Holocene deposits at the middle-ranked habitats are less diverse and perhaps reflective of less stress on shellfish resources than at the high-ranked habitats. Nonetheless, high densities of shellfish at CA-SRI-116 and CA-SRI-138, unit 1, do suggest that the populations at these sites were substantial.
At the end of the middle and beginning of the late Holocene, the number of settlement locations and sites expands and there is a general increase in the intensity of occupation of permanent settlements (Fig. 8). There is also a general trend toward smaller mussels at those locations that continued to be occupied from the middle Holocene, further supporting the interpretation of resource stress. With the exception of the relatively sparse deposits at CA-SRI-19, unit 1, there is an increase in total meat weight between the final middle Holocene deposits and initial late Holocene deposits (Table 6; Fig. 3), including an increase in fish and sea mammal in all cases and an increase in shellfish in all except Dry/Soledad Canyons. This presumably reflects an increase in population density in these habitats and, thus, an impetus for a geographic expansion to lower ranked settlement sites. In addition, there is a clear increase in fishing from the middle to late Holocene (Figs. 3 and 8). Fishing requires more technological investment than collecting shellfish, but it also is more easily intensified and therefore a more flexible strategy for supporting larger population densities and sedentism (Raab 1992; Glassow 1993). The decrease in shellfish diversity and evenness between the middle and late Holocene could also reflect the increased focus on fishing, a shift that continues into the Late Period, at which time fishing, particularly offshore, increases even more at village sites at the expense of shellfish (Jazwa 2015).
CA-SRI-333 and the Late Period rockshelter sites in Cow, Lobos, and Tecolote Canyon (Table 4) suggest that population was not entirely condensed at large villages late in time. A seasonality study at CA-SRI-333 indicates that people were there to some degree in all seasons except winter (Jazwa 2015). Because our investigation of rockshelter sites is limited to a single date for initial occupation and observations of surface deposits and erosional cuts, it is difficult to determine the extent to which those sites were used. Given their location and superficial deposits, it seems likely that they were short-term or seasonal sites, used to get out of the wind or to stay while collecting terrestrial resources from the island interior. Plant resources were likely the primary objective (Timbrook 2007; Gill 2013, 2014), but low quality chalcedony, which outcrops near the head of Cow and Lobos Canyon, may have also been important.
Caveats and Future Directions
Limitations potentially affecting our conclusions include the issue of sample size. We include excavation materials from the mouths of only 6 of the 19 major drainages on Santa Rosa Island. Within sites, we did our best to sample different settlement loci, but we cannot guarantee that we have sampled all periods of settlement in each location. We excavated relatively small 25 × 25-cm column samples to minimize site disruption and because of the time and resources required to sort the high density of midden. The comparative nature of our study design favored sampling a broad range of locations rather than intensively excavating a small number of them. While there is always bias in subsampling a site, the density of shell and fish bone were high in all units, increasing confidence that they are representative of the faunal assemblage, at least in the immediate area (see Kennett 2005; Joslin 2012). Given their much larger size, sea mammal remains are distributed more unevenly, making extrapolations from small column samples less trustworthy than for fish or shellfish.
We trace habitat and settlement on Santa Rosa in much greater detail than previous IFD applications (cf., Kennett et al. 2009; Winterhalder et al. 2010). There is more to settlement mobility than such an island-wide approach can take into account, including movement to smaller seasonal and special purpose sites (see Kennett 2005; Jazwa et al. 2015a). Finally, we note that Santa Rosa Island includes only relatively high-ranked settlements. We predict that the low-ranked settlements along the northwest coast of Santa Cruz Island will have late initial settlement, if any, and have limited evidence for continuous occupation. Further studies will include survey and excavation there.
Conclusion
Field studies in four NCI drainages including targeted survey and excavation of sites differing in basic suitability provide partial support for predictions of the IFD (Table 10). In general, the high-ranked locations were settled early at the beginning of the middle Holocene and were persistently occupied through the late Holocene. However, we observe early permanent settlement distributed across high- and middle-ranked locations. We offer several potential explanations for this pattern, including the possibility that it is the product of early despotic behavior, that low population densities allowed people to value factors not included in the model, or because of submergence of early settlement sites by rising sea levels (Fig. 9).
Dry and Soledad Canyons, our high-ranked habitat, were the first to experience intensive settlement that persisted throughout the middle and late Holocene. The middle-ranked drainages at Cow/Lobos and La Jolla Vieja Canyons were occupied through the middle Holocene, but less intensively. The lowest ranked habitat in this study, Bee Canyon, was not a site of permanent settlement until the late Holocene. All of the four drainages on Santa Rosa examined in detail in this study are in the top 2 quartiles of suitability for drainages on all of the NCI (Winterhalder et al. 2010), and therefore, all were settled before even lower ranked habitats on Santa Cruz and San Miguel Islands.
During the middle Holocene, the assemblages with the highest shellfish diversity were from the high-ranked habitats, which also would have had the highest population densities. During the late Holocene, population expansion was supported in part from an increase in fishing and sea mammal hunting, which could readily be intensified using new technologies. After the MLT, the population of the island condensed to relatively few large coastal villages, following a settlement pattern more consistent with the IDD. Populations likely congregated at villages to secure access to scarce resources and to seek protection from competing factions. Despotism in the form of territoriality expanded as previously occupied drainages were abandoned, a process that may also have been affected by economies of scale offered by the new technologies available in the more aggregated sites (see Winterhalder et al. 2010). Across the occupation of the NCI, we find good support for the IFD model, now applied with much more explicit attention to temporal dynamics of habitat suitability (Jazwa et al. 2013) arising from population-independent environmental change in habitat suitability. These temporal dynamics are related to fresh water availability, diet breadth change, technology, and changes in political economy.
The use of the IFD and IDD is becoming more prominent in anthropology (see Codding and Bird 2015). Perhaps the greatest benefit of these models is their ability to represent in one formulation economic relationships among variables considered critical by human ecologists: population density and growth, migration, settlement, resource selection and exploitation, environmental change, subsistence efficiency and economic returns, hierarchy, and competition. The IFD/IDD subsumes several other well-established archaeological models (see Jazwa 2015). The models adapt well to changing scale, from single sites through time (Jazwa et al. 2013), to detailed data-driven comparisons of multiple habitats (this paper), to regional settlement (Kennett et al. 2009; Winterhalder et al. 2010), to settlement of broader regions such as California (Codding and Jones 2013) or the Pacific (Kennett et al. 2006) and Australia (O’Connell and Allen 2012). Confirmed from several angles using data from the Channel Islands, the IFD/IDD provides an example for understanding human settlement patterns among other hunter-gatherer populations and can be expanded to model human decision-making among larger, more complex populations throughout the world.
References
Allen, J., & O’Connell, J. F. (2008). Getting from Sunda to Sahul. In G. A. Clark, F. Leach, & S. O’Connor (Eds.), Islands of inquiry: colonisation, seafaring and the archaeology of maritime landscapes (pp. 31–46). Canberra: Australian National University.
Antevs, E. (1948). Climatic changes and pre-white man in the Great Basin, with emphasis on glacial times and post-glacial times. University of Utah Bulletin, 38, 168–191.
Antevs, E. (1952). Climatic history and the antiquity of man in California. University of California Archaeological Survey Reports, 16, 23–31.
Antevs, E. (1955). Geologic-climatic dating in the west. American Antiquity, 20, 317–335.
Arnold, J. E. (1987). Craft specialization in the prehistoric Channel Islands, California. Berkeley: University of California Press.
Arnold, J. E. (1990). Lithic resource control and economic change in the Santa Barbara Channel region. Journal of California and Great Basin Anthropology, 12(2), 158–172.
Arnold, J. E. (1991). Transformation of a regional economy: sociopolitical evolution and the production of valuables in southern California. Antiquity, 65, 953–962.
Arnold, J. E. (1992a). Complex hunter-gatherer-fishers of prehistoric California: chiefs, specialists, and maritime adaptations of the Channel Islands. American Antiquity, 57(1), 60–84.
Arnold, J. E. (1992b). Early-stage biface production industries in coastal southern California. In J. E. Arnold (Ed.), Stone tool procurement, production, and distribution in California prehistory. Perspectives in California archaeology, volume 2 (pp. 67–130). Los Angles: Cotsen Institute of Archaeology.
Arnold, J. E. (1995). Transportation innovation and social complexity among maritime hunter-gatherer societies. American Anthropologist, 97(4), 733–747.
Arnold, J. E. (1997). Bigger boats, crowded creekbanks: environmental stresses in perspective. American Antiquity, 62, 337–339.
Arnold, J. E. (2001a). The Chumash in world and regional perspectives. In J. E. Arnold (Ed.), The origins of a Pacific coast chiefdom: the Chumash of the Channel Islands (pp. 1–20). Salt Lake City: University of Utah Press.
Arnold, J. E. (2001b). Social evolution and the political economy in the northern Channel Islands. In J. E. Arnold (Ed.), The origins of a Pacific coast chiefdom: the Chumash of the Channel Islands (pp. 287–296). Salt Lake City: University of Utah Press.
Arnold, J. E., & Graesch, A. P. (2004). The later evolution of the Island Chumash. In J. E. Arnold (Ed.), Foundations of Chumash complexity (pp. 1–16). Los Angeles: Cotsen Institute of Archaeology.
Arnold, J. E., & Munns, A. (1994). Independent or attached specialization: the organization of shell bead production in California. Journal of Field Archaeology, 21(4), 473–489.
Arnold, J. E., & Tissot, B. N. (1993). Measurement of significant paleotemperature variation using black abalone shells from prehistoric middens. Quaternary Research, 39, 390–394.
Arnold, J. E., Colten, R. H., & Pletka, S. (1997). Contexts of cultural change in insular California. American Antiquity, 62(2), 300–318.
Åström, M. (1994). Travel cost and the ideal free distribution. Oikos, 69(3), 516–519.
Bell, A. V., & Winterhalder, B. (2014). The population ecology of despotism: concessions and migration under the Ideal Despotic Distribution. Human Nature, 25, 121–135.
Benson, L., Kashgarian, M., Rye, R., Lund, S., Paillet, F., Smoot, J., Kester, C., Mensing, S., Meko, D., & Lindström, S. (2002). Holocene multidecadal and multicentennial droughts affecting northern California and Nevada. Quaternary Science Reviews, 21, 659–682.
Bettinger, R. L. (1991). Hunter-gatherers: archaeological and evolutionary theory. New York: Plenum.
Bettinger, R. L., Malhi, R., & McCarthy, H. (1997). Central place models of acorn and mussel processing. Journal of Archaeological Science, 24, 887–899.
Bocinsky, R. K., & Kohler, T. A. (2014). A 2,000-year reconstruction of the rain-fed maize agricultural niche in the US southwest. Nature Communications, 5, 5618. doi:10.1038/ncomms6618.
Braje, T. J., Kennett, D. J., Erlandson, J. M., & Culleton, B. J. (2007). Human impacts on nearshore shellfish taxa: a 7,000 year record from Santa Rosa Island, California. American Antiquity, 72(4), 735–756.
Bronk Ramsey, C. (2009). Bayesian analysis of radiocarbon dates. Radiocarbon, 51(1), 337–360.
Campbell, B., & Braje, T. J. (2015). Estimating California mussel (Mytilus californianus) size from hinge fragments: a methodological application in historical ecology. Journal of Archaeological Science, 58, 167–174.
Clifford, R.A. (2001). Middle Holocene hilltop and ridgeline settlement on the northern Channel Islands of California: a study of evolutionary stability. M.A. thesis, Department of Anthropology, California State University, Long Beach.
Codding, B. F., & Bird, D. W. (2015). Behavioral ecology and the future of archaeological science. Journal of Archaeological Science, 56, 9–20.
Codding, B. F., & Jones, T. L. (2013). Environmental productivity predicts migration, demographic, and linguistic patterns in prehistoric California. Proceedings of the National Academy of Science, 110(36), 14569–14573.
Cole, K. L., & Liu, G. (2004). Holocene paleoecology of an estuary on Santa Rosa Island, California. Quaternary Research, 41, 326–335.
Colten, R. H. (2001). Ecological and economic analysis of faunal remains from Santa Cruz Island. In J. E. Arnold (Ed.), The origins of a Pacific coast chiefdom: the Chumash of the Channel Islands (pp. 199–210). Salt Lake City: University of Utah Press.
Culleton, B.J. (2012). Human ecology, agricultural intensification and landscape transformation at the ancient Maya polity of Uxbenká, southern Belize. Ph.D. dissertation, Department of Anthropology, University of Oregon, Eugene.
Davis, O. K. (1992). Temporal and spatial patterns of distribution and recovery in a kelp forest community. Ecological Monographs, 62, 421–445.
Erlandson, J. M., & Jones, T. L. (Eds.). (2002). Catalysts to complexity: Late Holocene societies of the California coast. Los Angles: Cotsen Institute of Archaeology.
Erlandson, J. M., Rick, T. C., & Peterson, C. (2005). A geoarchaeological chronology of Holocene dune building on San Miguel Island, California. The Holocene, 15, 1227–1235.
Erlandson, J. M., Rick, T. C., Jones, T. L., & Porcasi, J. F. (2007). One if by land, two if by sea: who were the first Californians? In T. L. Jones & K. A. Klar (Eds.), California prehistory: colonization, culture and complexity (pp. 53–62). Landam: Altamira.
Erlandson, J. M., Rick, T. C., Braje, T. J., Steinberg, A., & Vellanoweth, R. L. (2008). Human impacts on ancient shellfish: a 10,000 year record from San Miguel, California. Journal of Archaeological Science, 35, 2144–2152.
Erlandson, J. M., Rick, T. C., Braje, T. J., Casperson, M., Culleton, B., Fulfrost, B., Garcia, T., Guthrie, D., Jew, N., Kennett, D., Moss, M. L., Reeder, L., Skinner, C., Watts, J., & Willis, L. (2011). Paleoindian seafaring, maritime technologies, and coastal foraging on California’s Channel Islands. Science, 441, 1181–1185.
Fagan, B. (2004). The house on the sea: an essay on the antiquity of planked canoes in southern California. American Antiquity, 69(1), 7–16.
Fitzhugh, B., & Kennett, D. J. (2010). Seafaring intensity and island-mainland interaction along the Pacific coast of North America. In A. Anderson, J. Barrett, & K. Boyle (Eds.), The global origins and development of seafaring (pp. 69–80). Cambridge: McDonald Institute for Archaeological Research.
Fretwell, S. D. (1972). Population in a seasonal environment. Princeton: Princeton University Press.
Fretwell, S. D., & Lucas, H. L., Jr. (1969). On territorial behavior and other factors influencing habitat distribution in birds. I. Theoretical development. Acta Biotheoretica, 19, 16–36.
Gamble, L. H. (2002). Archaeological evidence for the origin of the plank canoe in North America. American Antiquity, 67(2), 301–315.
Gill, K. M. (2013). Paleoethnobotanical investigations on the Channel Islands: current directions and theoretical considerations. In C. S. Jazwa & J. E. Perry (Eds.), California’s Channel Islands: the archaeology of human-environment interactions (pp. 113–136). Salt Lake City: University of Utah Press.
Gill, K. M. (2014). Seasons of change: using seasonal morphological changes in Brodiaea corms to determine season of harvest from archaeobotanical remains. American Antiquity, 79(4), 638–654.
Giovas, C. M. (2009). The shell game: analytic problems in archaeological mollusc quantification. Journal of Archaeological Science, 36, 1557–1564.
Giovas, C. M., & Fitzpatrick, S. M. (2014). Prehistoric migration in the Caribbean: past perspectives, new models and the ideal free distribution in West Indian colonization. World Archaeology, 46(4), 569–589.
Glassow, M. A. (1977). An archaeological overview of the northern Channel Islands, California, including Santa Barbara Island. Tucson: National Park Service.
Glassow, M.A. (1993). Changes in subsistence on marine resources through 7,000 years of prehistory on Santa Cruz Island. In Glassow, M.S. (Ed.), Archaeology on the northern Channel Islands of California (pp. 75–94). Coyote Press Archives of California Prehistory, vol. 34. Salinas: Coyote Press.
Glassow, M. A. (1999). Measurement of population growth and decline during California prehistory. Journal of California and Great Basin Anthropology, 21(1), 45–66.
Glassow, M. A. (2000). Weighing vs. counting shellfish remains: a comment on Mason, Peterson, and Tiffany. American Antiquity, 65(2), 407–414.
Glassow, M. A., Braje, T. J., Costello, J. G., Erlandson, J. M., Johnson, J. R., Morris, D. P., Perry, J. E., & Rick, T. C. (2010). Channel Islands National Park archaeological overview and assessment. Ventura: Cultural Resources Division, Channel Islands National Park.
Glassow, M. A., Thakar, H. B., & Kennett, D. J. (2012). Red abalone collecting and marine water temperature during the middle Holocene occupation of Santa Cruz Island, California. Journal of Archaeological Science, 39, 2574–2582.
Hendy, I. L., Dunn, L., Schimmelmann, A., & Pak, D. K. (2013). Resolving varve and radiocarbon chronology differences during the last 2000 years in the Santa Barbara sedimentary record, California. Quaternary International, 310, 155–168.
Hughes, M. K., & Graumlich, L. J. (2000). Multi-millennial Nevada precipitation reconstruction. International tree-ring data bank. IGBP PAGES/World Data Center-A for Paleoclimatology Data Contribution Series #2000-049. Boulder: NOAA/NGDC Paleoclimatology Program.
Jazwa, C.S. (2015). A dynamic ecological model for human settlement on California’s northern Channel Islands. Ph.D. dissertation, Department of Anthropology, Pennsylvania State University, University Park.
Jazwa, C. S., & Perry, J. E. (2013). The ecological, environmental, and cultural contexts for island archaeology. In C. S. Jazwa & J. E. Perry (Eds.), California’s Channel Islands: the archaeology of human-environment interactions (pp. 5–25). Salt Lake City: University of Utah Press.
Jazwa, C. S., Kennett, D. J., & Hanson, D. (2012). Late Holocene subsistence change and marine productivity on western Santa Rosa Island, California. California Archaeology, 4(1), 69–97.
Jazwa, C. S., Kennett, D. J., & Winterhalder, B. (2013). The ideal free distribution and settlement history at Old Ranch Canyon, Santa Rosa Island. In C. S. Jazwa & J. E. Perry (Eds.), California’s Channel Islands: the archaeology of human-environment interactions (pp. 75–96). Salt Lake City: University of Utah Press.
Jazwa, C. S., Braje, T. J., Erlandson, J. M., & Kennett, D. J. (2015a). Central place foraging and shellfish processing on California’s northern Channel Islands. Journal of Anthropological Archaeology. doi:10.1016/j/jaa.2015.05.005.
Jazwa, C.S., Duffy, C.J., Leonard, L., & Kennett, D.J. (2015b). Hydrological modeling and prehistoric settlement on Santa Rosa Island, California. Geoarchaeology: An International Journal. doi:10.1002/gea.21532.
Johnson, J.R. (1982). An ethnographic study of the island Chumash. M.A. thesis, Department of Anthropology, University of California, Santa Barbara.
Johnson, J. R. (1993). Cruzeño Chumash social geography. In M. A. Glassow (Ed.), Archaeology on the northern Channel Islands of California (pp. 19–46). Salinas: Coyote.
Johnson, J. R. (2001). Ethnohistoric reflections of Cruzeño Chumash society. In J. E. Arnold (Ed.), The origins of a Pacific coast chiefdom: the Chumash of the Channel Islands (pp. 21–52). Salt Lake City: University of Utah Press.
Johnson, J. R., Stafford, T. W., Jr., Ajie, H. O., & Morris, D. P. (2002). Arlington Springs revisited. In D. R. Brown, K. C. Mitchell, & H. W. Chaney (Eds.), The fifth California islands symposium (pp. 541–545). Santa Barbara: Santa Barbara Museum of Natural History.
Jones, T. L., & Schwitalla, A. (2008). Archaeological perspectives on the effects of Medieval drought in prehistoric California. Quaternary International, 188, 41–58.
Jones, T. L., Brown, G. M., Raab, L. M., McVicar, J. L., Spaulding, W. G., Kennett, D. J., York, A., & Walker, P. L. (1999). Environmental imperatives reconsidered: demographic crises in western North America during the Medieval Climatic Anomaly. Current Anthropology, 40(2), 137–170.
Joslin, T. L. (2012). Analytical sampling strategies for marine fish remains: measuring taxonomic diversity and abundance in central California middens. In M. A. Glassow & T. L. Joslin (Eds.), Exploring methods of faunal analysis: insights from California archaeology (pp. 135–147). Los Angeles: Cotsen Institute of Archaeology.
Kelly, R. L. (2013). The lifeways of hunter-gatherers: the foraging spectrum. Cambridge: Cambridge University Press.
Kennett, D.J. (1998). Behavioral ecology and the evolution of hunter-gatherer societies on the northern Channel Islands, California. Ph.D. dissertation. Department of Anthropology, University of California, Santa Barbara.
Kennett, D. J. (2005). The island Chumash: behavioral ecology of a maritime society. Berkeley: University of California Press.
Kennett, D. J., & Clifford, R. A. (2004). Flexible strategies for resource defense on the northern Channel Islands of California: an agent-based model. In S. M. Fitzpatrick (Ed.), Voyages of discovery: the archaeology of islands (pp. 21–50). Westport: Praeger.
Kennett, D. J., & Conlee, C. A. (2002). Emergence of late Holocene sociopolitical complexity on Santa Rosa and San Miguel Islands. In J. M. Erlandson & T. L. Jones (Eds.), Catalysts to complexity: Late Holocene societies of the California coast (pp. 147–165). Los Angeles: Cotsen Institute of Archaeology.
Kennett, D. J., & Kennett, J. P. (2000). Competitive and cooperative responses to climatic instability in coastal southern California. American Antiquity, 65(2), 379–395.
Kennett, D. J., & Winterhalder, B. (2008). Demographic expansion, despotism and the colonisation of east and south Polynesia. In G. Clark, F. Leach, & S. O’Connor (Eds.), Islands of inquiry: colonisation, seafaring and the archaeology of maritime landscapes (Terra Australis, 29) (pp. 87–96). Canberra: ANU E Press.
Kennett, D. J., Anderson, A. J., & Winterhalder, B. (2006). The ideal free distribution, food production, and the colonization of Oceania. In D. J. Kennett & B. Winterhalder (Eds.), Human behavioral ecology and the origins of agriculture (pp. 265–288). Berkeley: University of California Press.
Kennett, D. J., Kennett, J. P., Erlandson, J. M., & Cannariato, K. G. (2007). Human responses to middle Holocene climate change on California’s Channel Islands. Quaternary Science Reviews, 26, 351–367.
Kennett, D. J., Kennett, J. P., West, G. J., Erlandson, J. M., Johnson, J. R., Hendy, I. L., West, A., Culleton, B. J., Jones, T. L., & Stafford, T. W., Jr. (2008). Wildfire and abrupt ecosystem disruption on California’s northern Channel Islands at the Ållerød-Younger Dryas boundary (13.0–12.9 ka). Quaternary Science Reviews, 27, 2528–2543.
Kennett, D. J., Winterhalder, B., Bartruff, J., & Erlandson, J. M. (2009). An ecological model for the emergence of institutionalized social hierarchies on California’s northern Channel Islands. In S. Shennan (Ed.), Pattern and process in cultural evolution (pp. 297–314). Berkeley: University of California Press.
Kennett, D. J., Lambert, P. M., Johnson, J. R., & Culleton, B. J. (2013). Sociopolitical effects of bow and arrow technology in prehistoric Coastal California. Evolutionary Anthropology, 22, 124–132.
King, C. D. (1990). Evolution of Chumash society: a comparative study of artifacts used for social system maintenance in the Santa Barbara Channel region before A.D. 1804. New York: Garland.
LaMarche, V. C. (1973). Holocene climatic variations inferred from treeline fluctuations in the White Mountains, California. Quaternary Research, 3, 632–660.
LaMarche, V. C. (1974). Paleoclimatic inferences from long tree-ring records. Science, 183, 1043–1048.
Lambert, P. M. (1993). Health in prehistoric populations of the Santa Barbara Channel Islands. American Antiquity, 58, 509–522.
Lambert, P. M. (1997). Patterns of violence in prehistoric hunter-gatherer societies of coastal Southern California. In D. L. Martin & D. W. Frayer (Eds.), War and society, vol. 6, troubled times: violence and warfare in the past (pp. 77–109). Amsterdam: Gordon and Beach.
Lambert, P. M., & Walker, P. L. (1991). Physical anthropological evidence for the evolution of social complexity in coastal southern California. Antiquity, 65, 963–973.
Lindström, S. (1990). Submerged tree stumps as indicators of mid-Holocene aridity in the Lake Tahoe basin. Journal of California and Great Basin Anthropology, 12, 146–157.
Mason, R. D., Peterson, M. L., & Tiffany, J. A. (1998). Weighing vs. counting: measurement reliability and the California school of midden analysis. American Antiquity, 63(2), 303–324.
Mason, R. D., Peterson, M. L., & Tiffany, J. A. (2000). Weighing and counting shell: a response to Glassow and Claasen. American Antiquity, 65(4), 757–761.
McClure, S. B., Barton, C. M., & Jochim, M. A. (2009). Human behavioral ecology and climate change during the transition to agriculture in Valencia, Eastern Spain. Journal of Anthropological Research, 65(2), 253–269.
Morgan, A., Cummings, L. S., & Rudolph, J. L. (1991). Paleoenvironmental change. In C. F. J. L. Woodman & T. P. Rudolph (Eds.), Resource use and settlement in the Santa Ynez River Valley (pp. 65–101). Santa Barbara: Science Applications International Corporation.
Moritz, M., Hamilton, I. M., Chen, Y., & Scholte, P. (2014). Mobile pastoralists in the Logone floodplain distribute themselves in an ideal free distribution. Current Anthropology, 55(1), 115–122.
Munns, A. M., & Arnold, J. E. (2002). Late Holocene Santa Cruz Island: patterns of continuity and change. In J. M. Erlandson & T. L. Jones (Eds.), Catalysts to complexity: Late Holocene societies of the California coast (pp. 127–146). Los Angeles: Cotsen Institute of Archaeology.
Nagaoka, L. (2002). The effects of resource depression on foraging efficiency, diet breadth, and patch use in southern New Zealand. Journal of Anthropological Archaeology, 21, 419–442.
O’Connell, J. F., & Allen, F. (2012). The restaurant at the end of the universe: modelling the colonisation of Sahul. Australian Archaeology, 74, 5–17.
Perry, J.E. (2003). Changes in prehistoric land and resource use among complex hunter-gatherer-fishers on eastern Santa Cruz Island, California. PhD dissertation. University of California, Santa Barbara.
Perry, J. E., & Jazwa, C. S. (2010). Spatial and temporal patterning in chert exploitation on eastern Santa Cruz Island, California. American Antiquity, 75(1), 177–198.
Pisias, N. G. (1978). Paleoceanography of the Santa Barbara Basin during the last 8,000 years. Quaternary Research, 10, 366–384.
Pletka, S. (2001). Bifaces and the institutionalization of exchange relationships in the Chumash sphere. In J. E. Arnold (Ed.), The origins of a Pacific coast chiefdom: the Chumash of the Channel Islands (pp. 133–150). Salt Lake City: University of Utah Press.
Quade, J., Forester, R. M., Pratt, W. L., & Carter, C. (1998). Black mats, spring-fed streams, and late-glacial-age recharge in the southern Great Basin. Quaternary Research, 49, 129–148.
Raab, L. M. (1992). An optimal foraging analysis of prehistoric shellfish collecting on San Clemente Island, California. Journal of Ethnobiology, 12(1), 63–80.
Raab, L. M., & Larson, D. O. (1997). Medieval Climatic Anomaly and punctuated cultural evolution in coastal southern California. American Antiquity, 62(2), 319–336.
Raab, L. M., Bradford, K., Porcasi, J. F., & Howard, W. J. (1995). Return to Little Harbor, Santa Catalina Island, California: a critique of the marine paleoenvironmental model. American Antiquity, 60(2), 287–308.
Reeder-Myers, L., Erlandson, J. M., Muhs, D. R., & Rick, T. C. (2015). Sea level, paleogeography, and archaeology on California’s northern Channel Islands. Quaternary Research, 83, 263–272.
Reimer, P. J., Bard, E., Bayliss, A., Beck, J. W., Blackwell, P. G., Bronk Ramsey, C., et al. (2013). IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon, 55(4), 1869–1887.
Rick, T.C. (2004). Daily activities, community dynamics, and historical ecology on California’s northern Channel Islands. PhD dissertation. University of Oregon, Eugene.
Rick, T. C. (2007). The archaeology and historical ecology of late Holocene San Miguel Island, California. Los Angeles: Cotsen Institute of Archaeology.
Rick, T. C., Vellanoweth, R. L., Erlandson, J. M., & Kennett, D. J. (2002). On the antiquity of a single-piece shell fishhook: AMS radiocarbon evidence from the southern California coast. Journal of Archaeological Science, 29, 933–942.
Rick, T. C., Erlandson, J. M., Vellanoweth, R. L., & Braje, T. J. (2005). From Pleistocene mariners to complex hunter-gatherers: the archaeology of the California Channel Islands. Journal of World Prehistory, 19, 169–228.
Rick, T. C., Erlandson, J. M., Braje, T. J., Estes, J. A., Graham, M. H., & Vellanoweth, R. L. (2008). Historical ecology and human impacts on coastal ecosystems of the Santa Barbara Channel region, California. In T. C. Rick & J. M. Erlandson (Eds.), Human impacts on ancient marine ecosystems (pp. 77–101). Berkeley: University of California Press.
Rick, T. C., Erlandson, J. M., Jew, N. P., & Reeder-Myers, L. A. (2013). Archaeological survey, paleogeography, and the search for late Pleistocene paleocoastal peoples of Santa Rosa Island, California. Journal of Field Archaeology, 38(4), 321–328.
Shannon, C. E., & Weaver, W. (1949). The mathematical theory of communication. Urbana: University of Illinois Press.
Singh, G. G., & McKechnie, I. (2015). Making the most of fragments: a method for estimating shell length from fragmentary mussels (Mytilus californianus and Mytilus trossulus) on the Pacific coast of North America. Journal of Archaeological Science, 58, 175–183.
Singh, G. G., McKechnie, I., Braje, T. J., & Campbell, B. (2015). “All models are wrong but some are useful”: a response to Campbell’s comment on estimating Mytilus californianus shell size. Journal of Archaeological Science. doi:10.1016/j.jas.2015.08.021.
Stine, S. (1994). Extreme and persistent drought in California and Patagonia during Mediaeval time. Nature, 369, 546–549.
Strudwick, I.H., 1986. Temporal and areal considerations regarding the prehistoric circular fishhook of coastal California. M.A. thesis, Department of Anthropology, California State University, Long Beach.
Summers, K. (2005). The evolutionary ecology of despotism. Evolution and Human Behavior, 26, 106–135.
Sutherland, W. J. (1983). Aggregation and the ‘Ideal Free’ Distribution. Journal of Animal Ecology, 52(3), 821–828.
Sutherland, W. J. (1996). From individual behaviour to population ecology. Oxford: Oxford University Press.
Thompson, R. S. (1992). Late Quaternary environments in Ruby Valley, Nevada. Quaternary Research, 37, 1–15.
Timbrook, J. (2007). Chumash ethnobotany: plant knowledge among the Chumash people of southern California. Santa Barbara: Santa Barbara Museum of Natural History.
Treganza, T. (1995). Building on the ideal free distribution. Advances in Ecological Research, 26, 253–307.
Walker, P. L. (1989). Cranial injuries as evidence of violence in prehistoric southern California. American Journal of Physical Anthropology, 80, 313–323.
Walker, P.L., & Lambert, P.M. (1989). Skeletal evidence for stress during a period of cultural change in prehistoric California. Advances in Paleopathology, Journal of Paleopathology: Monographic Publication No. 1.
Watts, J., Fulfrost, B., & Erlandson, J. M. (2011). Searching for Santarosae: surveying submerged landscapes for evidence of Paleocoastal habitation off California’s northern Channel Islands. In B. Ford (Ed.), The archaeology of maritime landscapes (pp. 11–26). New York: Springer.
White, G.G. (1989). A report of archaeological investigations at eleven Native American coastal sites, MacKerricher Stae Park, Mendocino County, California. Sacramento: On file with California State Parks.
Winterhalder, B., Kennett, D. J., Grote, M. N., & Bartruff, J. (2010). Ideal free settlement on California’s northern Channel Islands. Journal of Anthropological Archaeology, 29, 469–490.
Yatsko, A. (2000). From sheepherders to cruise missiles: a short history of archaeological research at San Clemente Island. Pacific Coast Archaeological Society Quarterly, 36(1), 18–24.
Acknowledgments
We would like to thank Channel Islands National Park, including Kelly Minas, Don Morris, and Ann Huston, for the help with this project. This project was supported by Channel Islands National Park (135414, P11AC30805, Jazwa; 1443CA8120-96-003, Kennett), the National Science Foundation (BCS-1338350, Jazwa; BCS-1460369, SBR-9521974, Kennett), University of California, Santa Barbara, California State University, Long Beach, the University of Oregon, and Pennsylvania State University. Brendan Culleton provided insight about the IFD and Claire Ebert helped with some of the GIS work. Don Morris, Gil Unzueta, Henry Chodsky, Stephen Hennek, Terry Joslin, and Reilly Murphy assisted with excavations. Sheryl Gerety assisted with editing and proofing of this document. We would also like to thank Catherine Cameron and James Skibo, along with two anonymous reviewers, for their comments on this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Jazwa, C.S., Kennett, D.J. & Winterhalder, B. A Test of Ideal Free Distribution Predictions Using Targeted Survey and Excavation on California’s Northern Channel Islands. J Archaeol Method Theory 23, 1242–1284 (2016). https://doi.org/10.1007/s10816-015-9267-6
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10816-015-9267-6