
Abstract
Purpose
The aim of this retrospective study was to compare the competence of oocytes obtained from preovulatory and antral follicles.
Methods
Mature oocytes from preovulatory follicles were retrieved from women selected for standard IVF treatment (Group A). Mature oocytes from antral follicles were recovered from women undergoing hCG-primed in vitro maturation (IVM) treatment (Group B). Patients groups were matched for age, BMI, FSH, AMH and antral follicle count (AFC) values. In vivo matured oocytes from both groups were microinjected and resulting embryos were culture and selected on day 3 for embryo transfer.
Results
Oocyte pick-ups (OPU) were 315 and 204 in Groups A and B, respectively. Fertilization rates were comparable (72.8 and 75.9 %, respectively; P = 0.137). In Group A, in which the average number of embryos transferred was higher, clinical pregnancy rates per OPU (37.5 %) and embryo transfer (38.4 %) were superior in comparison to Group B (27.0 %, P = 0.013; 29.4 %, P = 0.041; respectively). On the other hand, implantation rates (Group A, 23.7 %; Group B, 20.8 %) and proportions of babies born per transferred embryo (Group A, 19.5 %; Group B, 16.9 %) were similar (P = 0.528 and 0.332, respectively).
Conclusions
Overall, this suggests that oocyte competence is already achieved at the antral stage of follicle development.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
It has long been recognized that oocyte growth is mutually interdependent with development and differentiation of the follicle. This close relationship is believed to occur uninterruptedly, from the awakening of the primordial follicle until ovulation [1]. This is reflected in the fact that, during the primary, secondary and incipient antral stages of follicle development, the trajectories of oocyte and follicle growth run parallel to each other. Then, as soon as a well-defined antral cavity forms and the somatic compartment generates the two distinct populations of mural and cumulus cells, the size of oocyte stabilizes whilst, until ovulation, the follicle structure experiences a further significant increase in size [2]. Such divergent growth paths, that encompass most of the antrum expansion phase, poses several questions about the significance of the final stages of follicle development and, above all, allude to the possibility that oocytes achieve full developmental competence well before the follicle reaches the final preovulatory stage.
In an assisted reproduction scenario, previous studies on humans attempted to verify this hypothesis by comparing the competence of oocytes recovered from follicles of different sizes, developed after IVF combined with controlled ovarian stimulation (IVF-COS). Collectively, it was found that oocytes from smaller follicles are less efficient [3–5]. Nevertheless, the conclusions that may be drawn from this kind analyses should be considered with caution. In fact, it is possible that poor oocyte quality is the cause, rather than the effect, of scarce follicle growth in response to exogenous FSH administration. Hence, these studies are unlikely to answer questions on the relationship between oocyte competence and follicle size.
The assisted reproduction approach founded on oocyte in vitro maturation (IVM) offers a unique model to study oocyte competence and follicle growth. The original strategy of IVM involves the collection of oocytes from antral follicles sized 8–12 mm, in the absence of exogenous gonadotropins. The recovered oocytes, still arrested at the immature germinal vesicle (GV) stage, are subsequently matured in vitro to the metaphase II (MII) stage and used to generate embryos suitable for transfer [6]. From a clinical standpoint, the concept of IVM has been further elaborated by favouring the initiation of oocyte maturation in vivo through the administration of hCG before the leading follicle exceeds 12 mm in size [7] that is to say, before dominance is established and atresia is induced in smaller follicles. Interestingly, a number of oocytes collected from such antral follicles exposed to hCG not only initiate but also complete the maturation process in vivo [8, 9]. This suggests that such oocytes acquire the competence to mature meiotically in vivo several days before their follicle achieves the preovulatory stage. Yet, it is not definitely established whether their developmental competence is comparable or equivalent to that of oocytes derived from fully mature follicles obtained in conventional IVF cycles.
Motivated by the experience gained over years of clinical practice in IVM [10–13], in the present study we aimed to compare not IVM with standard IVF-COS as clinical approaches, but rather to evaluate the relative competence of oocytes matured in vivo recovered from follicles of different developmental stages. Pursuing this objective, oocytes matured in vivo retrieved from antral follicles in IVM cycles were analyzed together with oocytes normally recovered from preovulatory follicles following conventional IVF-COS in order to assess comparatively their ability of establishing a viable pregnancy. Overall, this study raises interesting considerations on oocyte and follicle physiology.
Materials and methods
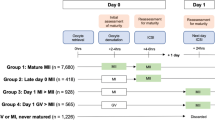
During the period June 2011 to December 2012, a consecutive cohort of 454 infertile women requiring IVM or standard IVF treatment at Biogenesi Reproductive Medicine Centre, Monza – Italy, was included in the study. A total of 519 cycles of IVM and standard IVF were carried out, of which: 315 standard IVF cycles out of 285 women and 204 IVM cycles out of 169 women were investigated and retrospectively analysed. The study was approved by the competent ethical committee. Written informed consent was obtained from all couples before starting treatment.
Inclusion and exclusion criteria
Couples included in the study were eligible for an IVF procedure because of infertility due to male factor, tubal factor and polycystic ovary with or without chronic anovulation (PCO, PCOS). IVM cases were designated according to the selective criteria previously described [11, 12, 14]. Concerning this, it should be noted that IVM was initially conceived to reduce in certain cases the risk of OHSS and at the same time exploit the potential of numerous small antral follicles, factors that characterize women with a large ovarian reserve (PCO/PCOS subjects). However, over the years the field of application of IVM has been extended to other cases. In particular, IVM has been proposed as an alternative treatment for women who are not comfortable with being exposed to relatively larger doses of hormones or undergoing more demanding treatment schedules. For these reasons, at Biogenesi IVM has been developed over the last decade as a treatment also for carefully selected non-PCO women.
The standard IVF stimulated cases were attentively matched for age, BMI and parameters of ovarian reserve (Table 1). Women with poor ovarian reserve or endometriosis as cause of infertility were excluded, as well as those couples with very severe male infertility (less than 0.1 spermatozoa/ml after semen preparation) or with surgically retrieved spermatozoa due to azoospermia. Women were at their first or second ART procedure.
Since the study design was to compare MII stage oocytes derived from antral follicles with those originated from preovulatory follicles, only cycles in which MII oocytes were available at the day of oocyte pick-up (OPU) were considered in the analysis.
In the IVM procedures expanded cumulus complexes enclosing MII oocytes (EC-MII) were identified and selected [15], and only cases in which it was possible to transfer embryos exclusively derived from EC-MII were included in the study.
Cycle preparation
In standard IVF stimulated cycles, pituitary down-regulation was achieved by a gonadotropin-releasing hormone agonist daily long protocol (Ferring, Italy). Ovarian stimulation was carried out with rFSH (Merck Sharp & Dohme, Italy or Merck Serono, Italy), tailoring doses and duration of stimulation according to the patient’s characteristics and needs [16]. Final oocyte maturation was triggered with 10,000 IU human hCG (Merck Sharp & Dohme, Italy) when at least three follicles with diameter ≥18 mm were visible at ultrasound scan monitoring. Oocyte retrieval was performed 36–38 h after hCG administration. Cycles were supplemented with intravaginal progesterone (Rottapharm, Italy) 600 mg/day starting from the day of OPU.
IVM cycles were carried out with gonadotropin priming rFSH 150 IU (Merck Sharp & Dohme, Italy or Merck Serono, Italy) for 3 days from the 3rd day of the cycle as previously described [9]. Before starting priming, a transvaginal ultrasonography was performed on day three of menstrual cycle so as to exclude the presence of ovarian cysts, evaluate endometrial thickness and the antral follicle count. Subsequent ultrasound scans were scheduled after a three-day FSH administration until a leading antral follicle of 10–12 mm and an endometrial thickness > 6 mm were assessed. Under those conditions a trigger with hCG 10,000 IU (Merck Sharp & Dohme, Italy) was administered and oocyte retrieval was scheduled 38 h later [11, 14].
COCs were examined and classified according to cumulus oophorus morphology and stage of oocyte maturation [15]. COCs were transferred in petri dishes, according to cumulus morphology, and cultured in IVM medium (Origio, Denmark) supplemented with rFSH 0.075 IU/ml (Merck Sharp & Dohme, Italy), hCG 0.10 IU/ml (Merck Sharp & Dohme, Italy), and 10 % serum substitute supplement (Irvine, USA), at 37 °C in a 6 % CO2 humidified atmosphere. The expanded cumulus COCs (EC), were cultured for 6 h and then treated with 80 IU/ml Cumulase solution (Origio, Denmark) to remove cumulus cells.
A luteal phase supplementation was provided in all women by 6 mg/day of oral oestradiol valerate (Bayer, Italy) plus 600 mg/day intravaginal progesterone (Rottapharm, Italy) starting on the day of oocyte retrieval.
Semen preparation, fertilization and embryo culture
Semen preparation, fertilization and embryo culture were carried out using the same procedure in the two groups. The semen samples were prepared by discontinuous gradients (47.5 and 90 %) of Sil-Select (Ferti-Pro, Belgium) and the spermatozoa were washed and re-suspended in IVF Medium (Origio, Denmark) and stored in an incubator at 37 °C and 6 % of CO2 humidified atmosphere until use. Since in IVM cycles ICSI is the standard insemination technique, in order to better compare the two groups all the included standard IVF cases were fertilised by ICSI. Fertilization was assessed 16–18 h after microinjection and confirmed by the presence of two pronuclei and two polar bodies. Pre-zygotes, zygotes and embryos up to 48 h were cultured in microdrops of ISM1 under oil (Origio, Denmark). After 48 h post-insemination, embryos were cultured in microdrops of Blast Assist under oil (Origio, Denmark) until embryo replacement.
Embryo development was followed and classified daily by observing the number of blastomeres and percentage of anucleate cytoplasmic fragments. High quality embryos were defined as having 8 blastomeres and less than 20 % of fragmentation on day 3. All embryo transfers were carried out 72 h after fertilization.
Pregnancy was tested 12–13 days after transfer by quantitative definition of serum β-hCG. If pregnancy was confirmed, oestrogen and progesterone supplementation was continued until the 12th week of gestation. The clinical pregnancy was defined by the presence of a gestational sac, with or without foetal heartbeat.
Statistical analysis
Analysis of demographic characteristics and biological and clinical outcomes in the two groups was performed by Student’s t-test for continuous variables and by chi-squared test for categorical or dichotomic variables. A logistic multivariate model was also studied to identify the variables independently associated with clinical pregnancy. Stata software 9.0 (Stata Corporation, Texas, USA) was used for performing the statistical analysis. A level of P < 0.05 was adopted for significance.
Results
Out of 519 analyzed cycles, 315 were IVF-COS cycles in which MII oocytes were collected from pre-ovulatory follicles (Group A) and 204 were IVM cycles, in which MII oocytes were collected from antral follicles (Group B). The proportion of PCO/PCOS patients was similar in the IVM and standard IVF groups (15/204, 12.2 % and 38/315 n = 12.1 %, respectively. P = NS).
Mean ± SD female age was 33.7 ± 2.7 years (range: 24–38) in group A and 33.3 ± 2.9 years (range: 27–38) in group B. As far as the other demographic characteristics are concerned, there were no statistical differences between the two groups, as shown in Table 1.
In Table 2 all the laboratory and clinical results related to both groups are summarized in detail as explained below.
Laboratory outcome
In group A, at oocyte recovery at least three follicles with diameter >18 mm were visible at ultrasound scan monitoring. A total of 3298 oocytes were collected, with an average number of recovered oocytes per cycle of 10.5 ± 5.0. One thousand nine hundred seven oocytes were inseminated (6.0 ± 2.3 per cycle), resulting in 1388 zygotes, with a fertilization rate of 72.8 %.
In group B, the average leading follicle diameter measured on the day of the OPU was 13.4 ± 1.4 mm. A total of 1377 oocytes were aspirated with an average number of recovered oocytes per cycle of 6.8 ± 4.2. As previously reported [9, 15], in IVM cycles carried out with FSH priming and hCG triggering, up to 20.3 % of recovered oocytes are classified as EC-MII, the remaining oocytes being immature and requiring IVM. EC-MII oocytes at retrieval were 618 (3.0 ± 2.2 per cycle) of which 561 (2.8 ± 1.7 per cycle) were inseminated resulting in 426 zygotes, with a fertilization rate of 75.9 %. No significant difference was found in fertilization rates between the two groups. Data relevant to in vitro maturation of oocytes collected at the GV or metaphase I (MI) stage were not described since the aim of the present study was not to analyse the outcome of such oocytes.
A total of 892 high quality embryos were obtained in Group A (64.3 %) and 272 in Group B (63.8 %) without any significant difference between the two groups.
Three hundred seven embryo transfers were performed in Group A and 187 in Group B. The number of transferred embryos was 620 and 308 respectively in the two groups. In group A, a significantly higher number of embryos was transferred: 2.0 ± 0.5 in Group A versus 1.5 ± 0.7 in Group B (p < 0.0001). In Group A, 7.2 % of transfers were carried out with one embryo, 83.7 % with two embryos and 9.1 % with three embryos. In group B, 37.4 % of transfers were performed with one embryo, 60.4 % with two embryos and 2.2 % with three embryos. As expected such distributions were significantly different between the two groups (p = <0.0001).
Clinical outcome
In group A, 118 clinical pregnancies, of which 29 resulting in twins, were observed. Therefore the clinical pregnancy rates per OPU and per embryo transfer (ET) in this group were 37.5 and 38.4 %, respectively. The implantation rate was of 23.7 %, with a miscarriage rate of 16.1 %. In group B, 55 clinical pregnancies, of which 9 resulting in twins, were established. The clinical pregnancy rate per OPU was 27.0 % and the clinical pregnancy rate per ET was 29.4 %, with an implantation rate of 20.8 % and a miscarriage rate of 20.0 %. Both clinical pregnancy rates per OPU and per ET were significantly higher in group A (P = 0.013 and P = 0.041 respectively). On the other hand, implantation and miscarriage rates were comparable between the two groups. In addition, the rate of babies born per transferred embryos was 19.5 % in Group A and 16.9 % in Group B, showing no significant difference between the two groups. Sub-analysis of double embryo transfer (DET) cycles was also indicative of a comparable clinical outcome. In fact, in DET cycles of group A and B, differences respectively in pregnancy rate per oocyte collection (37.5 vs. 27.0), implantation rate (24.9 vs. 19.5) and rate of babies born (20.2 vs. 15.9) were not statistically different.
Logistic regression analysis
A logistic model was elaborated to analyse the association between clinical pregnancy and different parameters. Female age, number of inseminated and fertilized oocytes, number of embryos transferred and the type of MII oocyte, derived from ovulatory follicles (Group A) or from antral follicles (Group B) were included in the model. As shown in Table 3, among the parameters evaluated the only independent factor associated with a higher probability of pregnancy was the number of embryos transferred. As expected, the number of embryos transferred was significantly higher in group A, suggesting that the higher pregnancy rate observed in group A was mainly related to the number of the embryos transferred. Separate logistic regression analysis of DET cycles did not reveal an association between female age, number of inseminated and fertilized oocytes or the type of MII oocyte and pregnancy rate (data not shown).
Discussion
In humans, histological [17] and ultrasonographic [18] investigations indicate that ovulation occurs when the dominant follicle attains a diameter of approximately 20 mm. It is therefore assumed that maximal developmental competence of the oocyte is gained concomitantly with the accomplishment of the preovulatory stage.
However, several lines of evidence in animal models challenge this belief, suggesting that completion of oogenesis precedes the follicular preovulatory stage. It was shown that, across a spectrum including different species such as hamsters, mice and pigs, full oocyte growth is almost or entirely reached by the time that follicle development has only progressed to the early antral stage [2]. Furthermore, in the cow it is well-established that oocytes are fully competent in completing maturation and supporting development as soon as the follicle reaches the size of 6 mm [19], well before the achievement of the preovulatory stage, corresponding to a diameter of 20 mm. Gene transcription studies also imply that oocyte developmental competence is gained long before ovulation. In particular in the cow, it emerged that maternal-effect genes, essential for early embryo development, such as Mater and Zar-1, are expressed at initial stages of follicular growth, considerably earlier than the final follicle expansion [20].
In humans, several studies attempted to shed light on this question. Ectors and colleagues [3] reported that rates of fertilization achieved by ICSI and embryo quality were positively associated to follicle volumes. In another analysis [4], it was found that oocytes recovered from follicles of different volume had a similar ability to undergo fertilization and generate pregnancies in ICSI, but not in IVF, cycles. Salha and colleagues [5] also described that oocytes derived from follicles of smaller volume had a reduced fertilization rate, although pre- and post-implantation development did not appear to be affected. Nogueira and collaborators [21] added further emphasis to the assumption that oocytes derived from smaller follicles had a limited competence, as shown by a higher incidence of embryo developmental delay observed on day 3 after ICSI. Altogether, these experiences suggest that acquisition of competence is not fully achieved in oocytes of smaller follicles. Although such study models are clinically relevant to a COS scenario, they are inappropriate to describe the competence of MII oocytes derived from follicles with different sizes. In fact, follicles displaying different sizes at the time of oocyte recovery in conventional IVF cycles, not only differ in developmental stage, but are also likely to be inherently diverse, as suggested by their different growth response to the same regimen of exogenous FSH. An ideal study, instead, should involve the opportunity to retrieve and assess mature oocytes from small follicles at a time when other follicles have not yet progressed to the preovulatory stage. Such model is offered by hCG-primed IVM cycles, in which fully mature oocytes can be retrieved even though maximal follicle diameter does not exceed a size corresponding to the antral stage. Oocytes derived from these healthy antral follicles of relatively smaller diameter, unlike those of equivalent size found in the cohorts of conventional IVF-COS cycles, are not expected to be affected by degenerative processes, such as atresia. The IVM model has been to some extent investigated by Son and colleagues. In particular, they examined hCG-primed cycles with and without MII-stage oocytes on the day of retrieval. They concluded that the cycles with in vivo matured oocytes result in a good clinical outcome. However, this study did not involve a comparison between in-vivo matured oocytes derived from antral (<13 mm) and preovulatory follicles (Son et al., 2008). In a subsequent analysis, the same authors compared sibling mature oocytes collected from different follicle sizes (<10 and 10–14 mm) on day of retrieval in in vitro maturation (IVM) cycles. Therefore, again, they did not extend their assessment to oocytes obtained from preovulatory follicles and did not report separate clinical outcomes of such groups of oocytes, limiting their observations solely to oocyte size, fertilization rate and embryo quality (Son et al., 2011). Guzman and colleagues (2013) reported fertilization, embryo quality and pregnancy rates generated from in-vitro matured oocytes derived from follicles <6 mm in diameter. No comparison was made with any type of in vivo matured oocytes.
On the basis of this premise and taking advantage of the unique human model represented by hCG-primed IVM cycles, the present study focused on the relationship between oocyte competence and stages of follicle development. Overall, the findings of the study suggest that the oocytes matured in vivo, derived from antral follicles of diameter not exceeding 12 mm at the time of hCG trigger, are comparable to those obtained from follicles having a size corresponding to or approaching the preovulatory stage, underpinning the view that acquisition of full competence is realized at the antral stage. This can be inferred from the fact that in the two study groups the rates of fertilization, proportion of high quality embryos and implantation rates were statistically comparable. We expected the pregnancy rates per oocyte recovery and embryo transfer to be higher in the IVF-COS group considering that, on average more embryos were transferred in such cycles. Likewise, the multivariate analysis indicated the number of transferred embryos as the sole parameter able to influence the pregnancy rate. In addition, extending the analysis beyond the implantation event, both the miscarriage rate and the proportion of babies born per transferred embryos were similar. Consistent with this, a comparable clinical outcome was also observed in DET cycles of IVM and IVF-COS groups. Clearly, the recovery of competent in vivo matured oocytes in IVM cycles primed with hCG greatly improves the clinical outcome of the IVM approach, as indicated by the fact that that implantation rate of embryos derived only by in vitro matured oocytes in hCG-primed cycles is disappointingly low, barely exceeding 6 % [15]. The hypothesis that a medium-sized antral follicle encloses an oocyte that has already achieved its developmental competence poses important considerations on the physiological significance of the later stages of follicle growth. In monovular species, perhaps the most obvious teleological explanation of the final phases of follicle development is the requirement to induce atresia in the less developed or compromised follicles so as to favour the shedding of a single oocyte from the leading follicle at ovulation [22, 23]. Moreover, the huge increase in oestradiol secretion, that characterizes the late follicular expansion, plays a crucial role in triggering the LH surge, crucial for ovulation, and promoting endometrial receptivity [24]. The endometrium has been suggested to be a limiting factor in IVM cycles performed in the absence of hCG priming, as indicated by the fact that cryopreserved embryos display a higher implantation ability in comparison to fresh embryos [25]. However, such element does not not seem relevant to the present study, because our analysis included only IVM cycles primed with FSH and hGC.
In conclusion, this study suggests that mature oocytes recovered from follicles of different size and development retain a comparable ability to give rise to a viable pregnancy. The data are consistent with the hypothesis that full acquisition of oocyte developmental ability is completed at the antral stage, preceding final preovulatory growth. As a consequence, this would also suggest that further follicle growth play a modest role in the process of oogenesis. However, being generated by a retrospective analysis our data cannot be considered conclusive for various reasons. For example, although severe and very severe male factor cases were not included in our analysis, the possible impact of paternal influences should be investigated in detail. Also, it should be also taken into account that in the present study the oocytes recovered from antral follicles were exposed to FSH for 3 days, a factor that may have had an impact on oocyte competence irrespective of the growth phase of the follicle. Additional studies are needed to test more extensively, also at the molecular and cellular level, similarities and difference between oocytes and follicles of different developmental stage. Consistent with this, we are currently investigating and comparing the transcriptomic profile of cumulus cells associated to the oocyte classes analysed in the present study.
Finally, in everyday practice, the study findings lend credit to the attitude towards priming IVM cycles with hCG and also add value to the approach of rescuing a standard IVF treatment in occurrence of an actual risk of ovarian hyperstimulation syndrome, by prematurely truncating the cycles through an hCG trigger exposure long before the leading follicle reaches the preovulatory size.
References
Edson MA, Nagaraja AK, Matzuk MM. The mammalian ovary from genesis to revelation. Endocr Rev. 2009;30(6):624–712.
Griffin J, Emery BR, Huang I, Peterson CM, Carrell DT. Comparative analysis of follicle morphology and oocyte diameter in four mammalian species (mouse, hamster, pig, and human). J Exp Clin Assist Reprod. 2006;3:2.
Ectors FJ, Vanderzwalmen P, Van Hoeck J, Nijs M, Verhaegen G, Delvigne A, et al. Relationship of human follicular diameter with oocyte fertilization and development after in-vitro fertilization or intracytoplasmic sperm injection. Hum Reprod. 1997;12(9):2002–5.
Bergh C, Broden H, Lundin K, Hamberger L. Comparison of fertilization, cleavage and pregnancy rates of oocytes from large and small follicles. Hum Reprod. 1998;13(7):1912–5.
Salha O, Nugent D, Dada T, Kaufmann S, Levett S, Jenner L, et al. The relationship between follicular fluid aspirate volume and oocyte maturity in in-vitro fertilization cycles. Hum Reprod. 1998;13(7):1901–6.
Trounson A, Wood C, Kausche A. In vitro maturation and the fertilization and developmental competence of oocytes recovered from untreated polycystic ovarian patients. Fertil Steril. 1994;62(2):353–62.
Chian RC, Buckett WM, Tulandi T, Tan SL. Prospective randomized study of human chorionic gonadotrophin priming before immature oocyte retrieval from unstimulated women with polycystic ovarian syndrome. Hum Reprod. 2000;15(1):165–70.
Son WY, Tan SL. Laboratory and embryological aspects of hCG-primed in vitro maturation cycles for patients with polycystic ovaries. Hum Reprod Update. 2010;16(6):675–89.
Fadini R, Dal Canto MB, Mignini Renzini M, Brambillasca F, Comi R, Fumagalli D, et al. Effect of different gonadotrophin priming on IVM of oocytes from women with normal ovaries: a prospective randomized study. Reprod BioMed Online. 2009;19(3):343–51.
Dal Canto MB, Mignini Renzini M, Brambillasca F, Cepparo H, Comi R, Villa A, et al. IVM–the first choice for IVF in Italy. Reprod BioMed Online. 2006;13(2):159–65.
Fadini R, Dal Canto MB, Renzini MM, Brambillasca F, Comi R, Fumagalli D, et al. Predictive factors in in-vitro maturation in unstimulated women with normal ovaries. Reprod BioMed Online. 2009;18(2):251–61.
Fadini R, Comi R, Mignini Renzini M, Coticchio G, Crippa M, De Ponti E, et al. Anti-mullerian hormone as a predictive marker for the selection of women for oocyte in vitro maturation treatment. J Assist Reprod Genet. 2011;28(6):501–8.
Fadini R, Mignini Renzini M, Guarnieri T, Dal Canto M, De Ponti E, Sutcliffe A, et al. Comparison of the obstetric and perinatal outcomes of children conceived from in vitro or in vivo matured oocytes in in vitro maturation treatments with births from conventional ICSI cycles. Hum Reprod. 2012;27(12):3601–8.
Fadini R, Mignini Renzini M, Dal Canto M, Epis A, Crippa M, Caliari I, et al. Oocyte in vitro maturation in normo-ovulatory women. Fertil Steril. 2013;99(5):1162–9.
Dal Canto M, Brambillasca F, Mignini Renzini M, Coticchio G, Merola M, Lain M, et al. Cumulus cell-oocyte complexes retrieved from antral follicles in IVM cycles: relationship between COCs morphology, gonadotropin priming and clinical outcome. J Assist Reprod Genet. 2012;29(6):513–9.
Fadini R, Brambillasca F, Renzini MM, Merola M, Comi R, De Ponti E, et al. Human oocyte cryopreservation: comparison between slow and ultrarapid methods. Reprod BioMed Online. 2009;19(2):171–80.
Gougeon A. Dynamics of follicular growth in the human: a model from preliminary results. Hum Reprod. 1986;1(2):81–7.
Hanna MD, Chizen DR, Pierson RA. Characteristics of follicular evacuation during human ovulation. Ultrasound Obstet Gynecol. 1994;4(6):488–93.
Luciano AM, Franciosi F, Modina SC, Lodde V. Gap junction-mediated communications regulate chromatin remodeling during bovine oocyte growth and differentiation through cAMP-dependent mechanism(s). Biol Reprod. 2011;85(6):1252–9.
Pennetier S, Uzbekova S, Perreau C, Papillier P, Mermillod P, Dalbies-Tran R. Spatio-temporal expression of the germ cell marker genes MATER, ZAR1, GDF9, BMP15, andVASA in adult bovine tissues, oocytes, and preimplantation embryos. Biol Reprod. 2004;71(4):1359–66.
Nogueira D, Friedler S, Schachter M, Raziel A, Ron-El R, Smitz J. Oocyte maturity and preimplantation development in relation to follicle diameter in gonadotropin-releasing hormone agonist or antagonist treatments. Fertil Steril. 2006;85(3):578–83.
Hillier SG. Gonadotropic control of ovarian follicular growth and development. Mol Cell Endocrinol. 2001;179(1–2):39–46.
Sirard MA, Desrosier S, Assidi M. In vivo and in vitro effects of FSH on oocyte maturation and developmental competence. Theriogenology. 2007;68:S71–6.
Chabbert Buffet N, Djakoure C, Maitre SC, Bouchard P. Regulation of the human menstrual cycle. Front Neuroendocrinol. 1998;19(3):151–86.
De Vos M, Ortega-Hrepich C, Albuz FK, Guzman L, Polyzos NP, Smitz J, et al. Clinical outcome of non-hCG-primed oocyte in vitro maturation treatment in patients with polycystic ovaries and polycystic ovary syndrome. Fertil Steril. 2011;96(4):860–4.
Author information
Authors and Affiliations
Corresponding author
Additional information
Capsule
Mature oocytes from antral and preovulatory follicles develop into embryos that implant with similar efficiency. This suggests that oocyte competence is already achieved at the antral stage of follicle development.
Rights and permissions
About this article

Cite this article
Fadini, R., Coticchio, G., Brambillasca, F. et al. Clinical outcomes from mature oocytes derived from preovulatory and antral follicles: reflections on follicle physiology and oocyte competence. J Assist Reprod Genet 32, 255–261 (2015). https://doi.org/10.1007/s10815-014-0386-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10815-014-0386-x