
Abstract
Cyanobacteria exhibit a variety of adaptive strategies that allow them to thrive in ever-changing aquatic environments. Here, successive steady states of continuous cultures were used to investigate the effects of quantified turbulence on the biochemical compounds and physiological processes of Anabaena flos-aquae in a photobioreactor under different dilution rates. A rapid increase in cell density was clearly observed following an increase in the turbulent dissipation rate at all growth rates of A. flos-aquae. The photosynthetic response to irradiance curves showed that the turbulence-treated strains exhibited lower photosynthetic oxygen evolution and saturating irradiance as well as higher respiration in rapidly growing young cells, indicating that they might not be very adaptable to high turbulent dissipation rates. Additionally, there was an increase in the protein levels of A. flos-aquae with increasing turbulence at all growth rates, whereas carbohydrate formation and lipid accumulation demonstrated the opposite trends. At a high growth rate, the level of carbohydrates decreased whereas that of lipids increased, which was interpreted as reflecting an adaptation to the turbulent environment. These findings suggest that turbulence sensitivity is shear regimen- and growth rate-dependent in A. flos-aquae. The high respiration capacity, low saturating irradiance, and conversion of carbohydrates to lipids represent effective measures for revealing the adaptive strategies of rapidly growing young cells under hydrodynamic regimes. The results regarding optimum lipid accumulation with specific growth traits have important implications for the design of cultivation methods of cyanobacteria resource utilization with respect to regulating turbulence.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Cyanobacteria receive attention and study worldwide due to their ability to proliferate extensively and form massive blooms in the upper water layer. Frequent outbreaks of cyanobacterial blooms have adverse effects on aquatic ecosystems and human health (Merel et al. 2013). Cyanobacteria have been recognized as a unique group of microorganisms because they exhibit characteristics of both algae and bacteria. Similar to eukaryotic microalgae, cyanobacteria perform oxygenic photosynthesis and show rapid growth and high biomass productivity. To develop cyanobacteria as a resource some researchers have attempted to develop cultivation technologies to apply cyanobacteria as precursors for fuels and other products (Grondahl 2009; Steinhoff et al. 2014).
The synthesis of valuable compounds in high amounts in microalgae can be amplified under stress or stimulation conditions (Suali and Sarbatly 2012; Han et al. 2015). However, stress conditions can also have negative effects on microalgal growth (Markou and Nerantzis 2013). Cyanobacteria exhibit a variety of adaptive strategies that allow them to perform well under environmental stress conditions, including low or high light intensity, high ultraviolet light intensity, high temperature, turbulent mixing, grazing pressure, and scarce or overabundant nutrients (Oliver et al. 2012; Paerl and Otten 2013). Several studies have proposed various hypotheses regarding how cyanobacterial species achieve this versatility by adjusting their buoyancy, using atmospheric nitrogen, utilizing phosphorus and inorganic carbon efficiently, optimizing cellular nutrient storage, and transforming their phenotype (Litchman et al. 2010; Carey et al. 2012). Cyanobacteria can accumulate intracellular compounds in response to harsh environmental conditions; therefore, they have the potential to be transformed into valuable fuels (Quintana et al. 2011; Kumar et al. 2017). However, cyanobacteria as a potent candidate for efficient production have not been given much attention.
Studies have shown that changes in component synthesis, including lipids, carbohydrates, proteins, and chlorophyll, may be induced by extreme temperature and light fluctuations, physical disturbances, pH, nutrient depletion, and predators (Tedesco and Duerr 1989; Tang et al. 2004; Han et al. 2015; Çelekli et al. 2016). Among several environmental parameters, turbulent mixing has been widely accepted as a source of external energy in the planktonic environment. Turbulence has a variety of effects on phytoplankton, such as affecting the light/dark frequencies in a well-documented phenomenon known as the “flashing light effect” (Grobbelaar 1994; Rubio et al. 2003), changing the diffusive sublayers and regulating the nutrient fluxes of cells (Guasto et al. 2012), causing mechanical damage (Chisti 2010; Michels et al. 2016) and altering behavior (Prairie et al. 2012; Sengupta et al. 2017). The impacts of turbulence on phytoplankton are complex and can influence a wide range of physiological aspects of organisms (Hondzo and Wüest 2009). In-depth researches of photosynthesis and nutrient strategies of phytoplankton in response to turbulence have been obtained from a few key genera, including Phaeodactylum, Porphyridium, Cyclotella, Fragilaria, Chlorella, Chlamydomonas, Alexandrium, Lingulodinium, Nodularia, and Aphanizomenon. (Moisander et al. 2002; Sullivan et al. 2003; Sobczuk et al. 2006; Wang et al. 2012; Leupold et al. 2013). Recently, the importance of turbulent mixing in shaping algal nutrient stoichiometry and its consequences for biochemical compounds has received attention. Hansen et al. (2013) suggested that mixing had an effect on the swimming speed of Chlamydomonas reinhardtii by affecting the biochemical composition, for example, through fatty acid accumulation. Chengala et al. (2013) reported that certain levels of moving fluid flow in a bioreactor enhanced the growth, protein, and fatty acid accumulation of Dunaliella primolecta. Our previous study found that cyanobacteria can adjust their growth rates in response to turbulence levels via the asynchronous cellular stoichiometry of C, N, and P (Xiao et al. 2016). Although the effects of turbulence on the cellular responses of phytoplankton have been extensively studied, such studies are lacking for the great majority of cyanobacteria. Furthermore, studies have demonstrated that carbon partitioning between carbohydrates and lipids in several species can be improved by applying stress, and carbon storage can differ greatly among the different adaptive strategies of algae (Michel et al. 2010; Pandit et al. 2017). However, few studies have focused on the turbulence conditions and growth traits that are associated with the accumulation of compounds that could serve as substrates for biofuel production. The partitioning of carbon into different cyanobacterial storage products in response to turbulence has not yet been fully evaluated.
In this work, we studied the filamentous cyanobacterium Anabaena flos-aquae which can be easily observed in freshwater systems and has specific responses for adapting to environmental changes (Singh et al. 2017). Few studies have investigated the potentially beneficial aspects of turbulence as a stress cultivation condition in promoting Anabaena growth. Compared to batch systems continuous cultivation by dilution with fresh media has many benefits that enable the culture environment to remain at the steady state (Tang et al. 2012). In this context, continuous cultivation under different turbulence levels and dilution rates was adopted in the current study to investigate the photosynthetic performance of A. flos-aquae and its partitioning of carbon into different storage products. The results contribute to the understanding of the specific property depending on species/strains growth under turbulence regimes and can be used to increase the production of valuable metabolites in Anabaena.
Materials and methods
Strain and culture conditions
The axenic freshwater filamentous cyanobacterium Anabaena flos-aquae FACHB-245 was kindly provided by the Freshwater Algae Culture Collection of the Institute of Hydrobiology, Chinese Academy of Sciences (FACHB-Collection; Wuhan, China). The strain was maintained in sterile BG11 medium (Rippka et al. 1979) under a cool-white fluorescent light intensity of 50 μmol photons m−2 s−1 with a 12-h:12-h light:dark cycle at a temperature of 25 ± 1 °C. Cells in the logarithmic growth phase in batch mode were incubated in a turbulence-controlled (0, 150, 200, and 50 rpm) continuous photobioreactor under three different dilution rates and then transitioned to continuous mode.
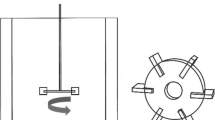
Following the methodology described in Xiao et al. (2016), a two-phase flow Computational Fluid Dynamics (CFD) model of the continuous flow reactor was applied to simulate the hydrodynamic conditions within the reactor. The computational domain of the model was the whole continuous flow reactor, which had slight differences from the batch reactor described in Xiao et al. (2016) (Fig. 1). The domain was discretized using unstructured grids, with finer grids applied in the impeller and shaft regions (maximum size = 1 mm). Approximately 1,200,000 total computational grids (maximum size = 2 mm) were created using the tetra mesh option of ANSYS ICEM CFD (ANSYS Inc.) to obtain the grid-independent solution for the flow. The simulation of the process was considered as a steady state and was numerically solved by using the commercial software package CFX 12.0 (ANSYS Inc.). The calibration and validation approach of the model was as discussed in Xiao et al. (2016). The turbulent mixing rate and Kolmogorov microscale at different levels of rotation speed were estimated at steady state. The integrated results are shown in Table 1.

The continuous photobioreactor system used to culture Anabaena flos-aquae (unit: mm)
Photobioreactor setup and determination of dilution rate
Continuous cultivation of A. flos-aquae was conducted in a modified photobioreactor system (Fig. 1), which was made of a double glass vessel with a width, height, and thickness of 120, 200, and 10 mm, respectively. The working volume (V0) was 1 L with an environmentally controlled growth chamber under an 80- μmol photons m−2 s−1 light intensity at 25 °C with a water jacket connected to a water bath. Sufficient mixing was achieved through axial flow impellers; thus, the system was assumed to be a continuous stirred-tank reactor with different turbulent mixing rates (defined above). Axenic fresh culture medium was pumped into the photobioreactor through silicone rubber tubing with flow rates (F) of 0.1, 0.3, and 0.6 L day−1, corresponding to dilution rates D = F/V0 of 0.1, 0.3, and 0.6 day−1, respectively. The culture medium was withdrawn at the same flow rate from the photobioreactor. In continuous cultivation at steady state, all culture parameters remain constant (culture volume, dissolved oxygen concentration, nutrient and product concentrations, pH, cell density, etc.). Growth of microorganisms occurs at a constant specific growth rate (μ), which is numerically equals to the dilution rate (D) (Tang et al. 2012). The test was performed in triplicate under the conditions described above.
Measurement of cell density and chlorophyll a
The cell density was determined using a hemocytometer under an Olympus CX43 microscope (Olympus, Japan). Prior to cell counting, A. flos-aquae filaments were ultrasonicated for 30 or 60 s at 20 kHz and 100 W (JY92-2D, SCIENTZ, China) to obtain single cells. Chlorophyll a was extracted with 90% acetone and measured spectrophotometrically according to Nusch (1980).
Photosynthetic oxygen evolution analysis
The photosynthetic oxygen evolution was measured by a Clark-type oxygen electrode (Chlorolab 2, Hansatech, UK). The temperature of the chamber was controlled by a thermostatic circulator at 25 °C. Illumination was set from 25 to 1000 μmol photons m−2 s−1, with oxygen evolution measured for at least 10 min at each irradiance. The parameters for the photosynthetic response to irradiance curves (P–E curves) were obtained according to Walsby (1997): P = Pm[1–exp(−αE/Pm)] + Rd + βE, where E is irradiance, P is the photosynthetic rate at irradiance E, Pm is the maximum photosynthesis rate, α is the initial slope of P–E curve, Rd is the dark respiration rate, β is the slope at high irradiance, and Ek is the saturating irradiance for photosynthesis (Ek = Pm/α). The photosynthetic rates were normalized to chlorophyll a, and nonlinear curve fitting of the data was performed with Origin 9.0 (OriginLab, USA).
Carbon contents and carbonic anhydrase activity
Samples were harvested by centrifugation (6300×g, 15 min, 25 °C), washed twice, and homogenized in liquid nitrogen with cold Veronal Buffer (12 mM, pH 8.3). The carbonic anhydrase (CA) activity was measured by the method of Wilbur and Anderson (1948). The time required for the pH to change from 8.3 to 7.3 was measured at 0–2 °C after the addition of 3 mL of CO2-saturated water to 3 mL of buffer samples. CA activity was calculated as an enzyme unit, EU = (T0/T) − 1, where T0 is the time for the uncatalyzed reaction and T is the time for the enzyme-catalyzed reaction. CA activity was normalized to Chl a.
To measure the cellular carbon content of A. flos-aquae, 30 mL culture suspensions were collected from each sample, filtered onto precombusted GF/F filters (Whatman, UK) and dried at 65 °C for 24 h. The dried suspensions were then removed and wrapped in weighted tinfoil and then weighed again to obtain the mass of samples. The carbon concentrations were analyzed by using a Vario El cube elemental analyzer (Elementar, Germany).
Biochemical composition determination
Anabaena flos-aquae culture suspensions were collected from each sample and centrifuged at 6300×g for 15 min (4 °C). Then the pellets were washed twice with distilled water and used to determine biochemical composition, including total carbohydrates, proteins, and lipids. The carbohydrate content was quantified spectrophotometrically by the phenol-sulfuric acid method using glucose as the standard (Dubois et al. 1956). The protein content was determined with Coomassie Blue G-250 reagent using bovine serum albumin as the standard (Bradford 1976). All contents were measured under steady-state conditions and normalized to cell counts. Lipid extraction was based on Bligh and Dyer (1959). Pellets were frozen at − 80 °C for 24 h and then dried in a freeze dryer for up to 4 days. Dried cells were ground into powder and treated with chloroform/methanol (2:1, v/v). The extracted lipid was centrifuged, and the chloroform layer, including lipids, was collected. The residues were subjected to repeated extraction five times. All of the extracts obtained were then combined and the lipid content was evaluated by its weight relative to the weight of dry organism biomass.
Statistical analysis
The data are presented as the mean ± standard deviation (SD). Significant differences between controls and treated samples were determined by ANOVA. All statistical analyses were carried out with Origin 9.0 (OriginLab, USA). Differences were considered significant at P < 0.05.
Results
Cell density of A. flos-aquae at steady-state conditions
Anabaena flos-aquae continuous cultivation in a 1 L photobioreactor was performed under different turbulent mixing levels. Each continuous culture in the steady-state condition was conducted for 20 days. Figure 2 shows the steady-state condition of cell density at various dilution rates (from 0.1 to 0.6 day−1). Changes in the dilution rate of the continuous culture had significant impacts on the steady-state cell density. As expected, reducing the dilution rate increased the steady-state cell density, which ranged from 2.18 × 106 to 3.92 × 107 cells mL−1. Under the same dilution rate, the steady-state cell density of A. flos-aquae increased with increasing turbulence intensity. For instance, the value of the steady-state cell density under a turbulent dissipation rate of 0.03 m2 s−3 was approximately 1.4×, 1.7× and 2.2× higher than that under the stagnation condition at a dilution rate of 0.1, 0.3, and 0.6 day−1, respectively (ANOVA, P < 0.05).

Cell density of Anabaena flos-aquae under steady-state conditions at different dilution rates and turbulent mixing levels. Data are the means ± SD (n = 3)
Photosynthetic characteristics of A. flos-aquae
The photosynthesis–irradiance (P–E) curves and parameters of cultured A. flos-aquae displayed marked differences among dilution rates and turbulent mixing levels (Fig. 3 and Table 2). The maximum photosynthesis rate (Pm) increased gradually with increasing turbulence intensity at a dilution rate of 0.1 day−1 but decreased gradually at a dilution rate of 0.6 day−1. At a dilution rate of 0.3 day−1, Pm and photosynthetic efficiency (α) were increased under the turbulence treatments, with maximum values observed at the dissipation rate of 0.007 m2 s−3. Moreover, α increased with turbulence intensity at the dilution rate of 0.6 day−1. Concomitantly, the value of the dark respiration rate (Rd) and saturating irradiance for photosynthesis (Ek) increased significantly (ANOVA, P < 0.05) under the turbulence treatments of A. flos-aquae at a dilution rate of 0.1 day−1. In contrast, the Rd value dramatically decreased with increasing turbulent dissipation rate at dilution rates of 0.3 and 0.6 day−1 (ANOVA, P < 0.05), as indicated by the higher oxygen consumption rates per cell. The lowest Rd value was − 147.98 ± 6.46 μmol O2 mg−1 Chl a h−1, which occurred at a turbulent dissipation rate of 0.03 m2 s−3 and dilution rate of 0.6 day−1. Relative to Ek under the stagnation condition, Ek was lower in the 0.007, 0.015, and 0.03 m2 s−3 treatments by 0.5, 0.4, and 0.3 times, respectively.

Photosynthetic oxygen evolution as a function of the incident photon flux density (PFD) in Anabaena flos-aquae grown at different dilution rates and turbulent mixing levels. (a) Dilution rate of 0.1 day−1. (b) Dilution rate of 0.3 day−1. (c) Dilution rate of 0.6 day−1. Data are the means ± SD (n = 3)
Changes in cellular carbon with the chlorophyll a ratios of A. flos-aquae
Carbon is used for maintenance, growth, and respiration related to growth. The C/Chl a ratio is a key physiological variable that can be used to evaluate the normalizing photosynthetic rate and carbon fixation efficiency. The results (Fig. 4) demonstrated that under a higher dilution rate, A. flos-aquae had a higher ratio of C/Chl a, while the C/Chl a ratio was decreased in the turbulent treatments relative to the stagnation condition except at low dilution rates. At a dilution rate of 0.1 day−1, the C/Chl a ratio slightly increased with increasing turbulent dissipation rate, although not significantly (ANOVA, P > 0.05). However, the ratios were significantly lower at a higher turbulence intensity (0.03 m2 s−3) than they were under the control treatments at dilution rates of 0.3 and 0.6 day−1 (ANOVA, P < 0.05).

Changes in the cellular carbon to chlorophyll a ratio of Anabaena flos-aquae grown at different dilution rates and turbulent mixing levels. Data are the means ± SD (n = 3)
Carbonic anhydrase activity of A. flos-aquae
To further investigate the effects of turbulence intensity and dilution rate on carbon fixation efficiency, the CA activity of cultured A. flos-aquae was measured under different turbulence intensities and at various dilution rates. The results showed that increasing the turbulence intensity had no significant effect on CA activity at the dilution rates of 0.1 and 0.6 day−1 (ANOVA, P > 0.05). At a dilution rate of 0.1 day−1, CA activity remained approximately stable, and at a dilution rate of 0.6 day−1, it showed a slight increase. However, there was a significant decrease at higher turbulence intensity at the dilution rate of 0.3 day−1 (ANOVA, P < 0.05). In addition, higher CA activity was observed at the higher dilution rate. The CA activity of A. flos-aquae cultured at a dilution rate of 0.6 day−1 was almost two times higher than that observed at a dilution rate of 0.3 day−1 in each case (Fig. 5).

Carbonic anhydrase activity of Anabaena flos-aquae grown at different dilution rates and turbulent mixing levels. Data are the means ± SD (n = 3)
Variation in the biochemical composition of A. flos-aquae
The composition varied among the different turbulence intensity and dilution rate treatments (Fig 6). Slight increases in the carbohydrate content per cell were generally observed in the turbulent conditions relative to the stagnation condition at the lowest dilution rate (0.1 day−1). However, a significant decline in carbohydrate synthesis was observed with increasing turbulent dissipation rate after exposure to the higher dilution rates (ANOVA, P < 0.05). The carbohydrate content per cell in the 0.03 m2 s−3 group was 40.3 and 72.2% lower than that in the control group at dilution rates of 0.3 and 0.6 day−1, respectively. The protein content per cell increased over all of the culture treatments relative to the control treatments. The turbulent samples exhibited a higher protein content per cell than did the control samples at all dilution rates (ANOVA, P < 0.05). The maximum protein content per cell was 2.86, 7.89, and 17.37 pg cell−1 at dilution rates of 0.1, 0.3, and 0.6 day−1, respectively. However, lipid content demonstrated the opposite trend. Total lipid accumulation decreased gradually with increasing turbulence intensity at a dilution rate of 0.1 day−1, while compared to the control samples, the turbulent samples exhibited markedly increased lipid proportions at dilution rates of 0.3 and 0.6 day−1 (ANOVA, P < 0.05). The lipid content peaked (15.8 and 18.7%) under turbulent dissipation rates of 0.007 and 0.03 m2 s−3 and dilution rates of 0.3 and 0.6 day−1, respectively.

Comparison of the biochemical composition of Anabaena flos-aquae grown at different dilution rates and turbulent mixing levels. Data are the means ± SD (n = 3)
Discussion
Cyanobacteria have different degrees of sensitivity to turbulence, which may vary among mixing intensities and species (Peters and Marrasé 2000). This work used continuous cultures to study photosynthetic performance and carbon partitioning in relation to growth traits and turbulence intensity in A. flos-aquae. The turbulent dissipation levels in the photobioreactor covered the range of intensities experienced by cyanobacteria in the field, and the dilution rate, which refers to the growth rate at the steady state in continuous cultivation, was below the maximum specific growth rate of A. flos-aquae (0.78 day−1) reported in the literature (Reynolds 2006).
Continuous cultivation at the laboratory scale has been recognized as useful for holding other relevant parameters constant, thus enabling the optimal levels of factors of interest to be determined (Czitrom 1999; Guedes et al. 2014). In the present study, the impact of turbulence on cells was first evaluated by direct measurement of cell density. A rapid increase in cell density was clearly observed following an increase in the turbulent dissipation rate at all dilution rates of A. flos-aquae under the steady-state conditions (Fig. 2). The dilution rate determines the mean residence time of the population and, thus, the average age of the cells in the reactor (Mirón et al. 2003). At high dilution rates, rapidly growing young cells are more sensitive to an increase in turbulence mixing than are cells cultured at lower dilution rates, exhibiting greater increases in cell density compared to control cells. However, previous studies have proposed that turbulent forces that do not kill a cell may still produce stress responses, such as decreased photosynthetic efficiency (Thomas et al. 1995).
The carbon and energy requirements for phytoplankton metabolism are all derived from the photosynthetic process (Wilhelm and Jakob 2011). The photosynthetic rate, carbohydrate accumulation, and utilization of stored photosynthates may vary among cells with different activities (Olesen and Ganf 1986). Defining the functional relationships between turbulence and the photosynthetic performance of A. flos-aquae in the laboratory is perceived as a first step towards determining the partitioning of carbon into storage under turbulence effects. In this study, the responses of A. flos-aquae to turbulence intensities at varied growth rates varied widely based on the P-E curves and parameter measurements. The maximum oxygen production rates in the turbulence cultures were higher than those of the control culture at low and medium growth rates, whereas the opposite trend was observed at the high growth rate of 0.6 day−1. It appears that the rapidly growing, younger cells were less adaptable to the high dissipation rate than were the older cells. Regarding photosynthesis, the slope of the P–E curve, α, increased with turbulence level at all growth rates of A. flos-aquae (Table 2). This result is consistent with the findings of Chengala et al. (2013) who concluded that turbulent conditions facilitated algal physiology, particularly in photosynthesis, as indicated by the higher O2 levels in the bioreactor. Wang et al. (2012) reported that turbulence imparts a positive effect on the structure of PS II in diatom species. However, in a study of the dinoflagellate Gonyaulax polyedra, photosynthesis was not as sensitive to turbulence as was population growth (Thomas et al. 1995). We speculate that these discrepancies might be due to study variation in the turbulent conditions employed and differences among species in response pattern. The present study showed that respiration was increased with the turbulent dissipation rate at a growth rate of 0.6 day−1 relative to the stagnant condition. Such negative effects could cause reductions in the oxygen production rate. Previous studies of the impacts of turbulence on photosynthesis in phytoplankton have primarily attributed the impacts to the “flashing light effect”, which refers to the changes in the light received by organisms due to the changes in the frequency of light-dark cycling (Grobbelaar 1994; Rubio et al. 2003). Nevertheless, in the case of A. flos-aquae, turbulence significantly decreased the saturating irradiance for photosynthesis at growth rates of 0.3 and 0.6 day−1. The Ek value under a high dissipation rate at a growth rate of 0.6 day−1 was even lower than the experimental light intensity supplied. When planktonic organisms are cultured under light conditions that exceed their light saturation threshold, mixing can improve the light exposure to individual cells (Sobczuk et al. 2006). Therefore, it is likely that the rapidly growing, younger cells of A. flos-aquae can optimize their saturating irradiance to adapt to turbulence. The range of Ek values in our experiment reflects the plasticity of A. flos-aquae, exhibiting different growth traits in response to differences in turbulence. Such plasticity might be a significant contributor to this cyanobacterium’s environmental adaptability.
Turbulence consistently affected the carbon to chlorophyll a ratio in A. flos-aquae, but there was considerable variability in the response among the different growth rates. This variability may arise from at least three sources: (1) Photosynthesis control pathways from light requirement to the capacity for carbon dioxide fixation. Dark respiration, which can promote the consumption of cellular carbon, increased with increasing turbulent dissipation rate in the rapidly growing, younger cells of A. flos-aquae. (2) Turbulence weakened the Calvin Cycle in A. flos-aquae at the growth rate of 0.3 day−1, as shown by the changes in CA activity, and reduced cellular carbon, as the maximum rate of carbon dioxide fixation is related to the amounts of CA or RuBisCo (MacIntyre et al. 2000). (3) At a given growth rate, cells under continuous cultivation were in different physiological states (with younger cells at the high dilution rate and older cells at the low dilution rate). The shear sensitivity of filamentous cyanobacteria may vary depending on the growth stage, as demonstrated in Nodularia (Moisander et al. 2002).
It is believed that microorganisms alter their biosynthetic pathways in an attempt to maintain their growth rates or increase survival under unfavorable environmental conditions (Li et al. 2011). In general, the biochemical contents measured in the present study were all higher at the high growth rates than at the low growth rates. However, the partitioning of photosynthetic carbon into biochemical components in response to turbulence varied among the different growth rates in our experiment. At all growth rates, A. flos-aquae contained increasing amounts of protein as the turbulent dissipation rate increased, and the rapidly growing young cells contained more protein than older cells. Proteins facilitate the growth of cells and participate in the metabolic and structural functions of the cell. The results of our study are in agreement with Chengala et al. (2013), who reported that turbulence favored the accumulation of protein in Dunaliella primolecta. The proportion of protein in the cells increased partly due to the synthesis of new proteins in the rapidly growing young cells and partly due to the consumption of some storage components, mainly carbohydrates (Mirón et al. 2003). It has been reported that carbohydrates share the same precursors with triacylglycerols, which are largely produced under stimulating growth conditions (Klok et al. 2014). However, to date, the relationships between carbohydrate and lipid synthesis remain unclear (Li et al. 2010). The present study showed that the processes of carbohydrate formation and lipid accumulation are asynchronous, and they demonstrated opposing trends at a given growth rate. As shown in Fig. 6, at the low growth rate, the carbohydrate content increased with increasing turbulent dissipation rate, whereas the lipid content decreased. At the high growth rate, the level of carbohydrates decreased with increasing turbulent dissipation rate, whereas that of lipids increased. These trend differences might be related to the differences in the specific shear sensitivity under different growth rates in A. flos-aquae, which indicated that the rapidly growing cells were more sensitive than were the older cells to turbulence. In the rapidly growing young cells, the respiration level increased and saturating irradiance decreased in response to a high turbulent dissipation rate, which can partially explain the negative effects on carbohydrate level. Under stress, some algal species can inhibit the carbohydrate biosynthetic pathway and redirect photosynthetic carbon flux towards lipid biosynthesis (Li et al. 2011). Lipid biosynthesis and accumulation occur in different growth phases (Yu et al. 2017). At high dilution rates, young cells contain lower amounts of storage lipids than do cells cultured at lower dilution rates (Reis et al. 1996). Our results demonstrated that the optimum level of lipid accumulation in response to turbulence was exhibited by the younger cells of A. flos-aquae, indicating that carbon partitioning by turbulence, which underlies the improvements in lipid accumulation, is sensitive to growth rate. Turbulent mixing has been shown to regulate the nutrient fluxes of cells (Karp-Boss et al. 1996). Increments of nitrogen and phosphorus fluxes in the rapidly growing young cells were also observed under the high turbulent dissipation rate (data not shown), resulting in the acceleration of lipid synthesis. Moreover, cyanobacterial lipids play important roles in thylakoid membrane structural components (Chen et al. 2013); therefore, a high lipid content is vital for photosynthetic processes. Taken together, the results suggest that altering the allocation of biochemical compounds might be a strategy for adapting to unfavorable conditions at high growth rates in A. flos-aquae. The results may serve as a theoretical guide for the enhancement of lipid productivity by regulating the turbulence supply. For future research, we suggest that fatty acid composition analysis be conducted and that multiple cyanobacterial species be studied to better understand their implications for the design of cultivation strategies for cyanobacteria resource utilization.
Conclusion
This study revealed shear regimen- and growth rate-dependent sensitivity to turbulence in a strain of Anabaena. Under the continuous culture conditions, a high respiration capacity, low saturating irradiance, and the conversion of carbohydrates to lipids were observed in the rapidly growing young stage of A. flos-aquae. These responses might reflect adaptive strategies employed to promote survival under high turbulence intensity. From a biotechnological perspective, such strategies represent safe, natural approaches for promoting lipid accumulation in rapidly growing young cells.
References
Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction and purification. Can J Biochem Physiol 37:911–917
Bradford M (1976) A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Carey CC, Ibelings BW, Hoffmann EP, Hamilton DP, Brookes JD (2012) Eco-physiological adaptations that favour freshwater cyanobacteria in a changing climate. Water Res 46:1394–1407
Çelekli A, Arslanargun H, Soysal Ç, Gültekin E, Bozkurt H (2016) Biochemical responses of filamentous algae in different aquatic ecosystems in South East Turkey and associated water quality parameters. Ecotoxicol Environ Saf 133:403–412
Chen D, Yan X, Xu J, Su X, Li L (2013) Lipidomic profiling and discovery of lipid biomarkers in Stephanodiscus sp. under cold stress. Metabolomics 9:949–959
Chengala A, Hondzo M, Mashek DG (2013) Fluid motion mediates biochemical composition and physiological aspects in the green alga Dunaliella primolecta Butcher. Limnol Oceanogr Fluids Environ 3:74–88
Chisti Y (2010) Shear sensitivity. In: Flickinger MC (ed) Encyclopedia of industrial biotechnology: bioprocess, bioseparation, and cell technology. Wiley, New York, pp 4360–4398
Czitrom V (1999) One-factor-at-a-time versus designed experiments. Am Stat 53:126–131
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F (1956) Colorimetric method for determination of sugars and related substances. Anal Chem 28:350–356
Grobbelaar JU (1994) Turbulence in algal mass cultures and the role of light/dark fluctuations. J Appl Phycol 6:331–335
Grondahl F (2009) Removal of surface blooms of the cyanobacteria Nodularia spumigena: a pilot project conducted in the Baltic Sea. Ambio 38:79–84
Guasto JS, Rusconi R, Stocker R (2012) Fluid mechanics of planktonic microorganisms. Annu Rev Fluid Mech 44:373–400
Guedes AC, Katkam NG, Varela J, Malcata FX (2014) Photobioreactors for cyanobacterial culturing. In: Sharma NK, Rai AK, Stal LJ (eds) Cyanobacteria: an economic perspective. Wiley, New York, pp 270–292
Han F, Pei H, Hu W, Song MM, Ma GX, Pei RT (2015) Optimization and lipid production enhancement of microalgae culture by efficiently changing the conditions along with the growth-state. Energy Convers Manag 90:315–322
Hansen TJ, Hondzo M, Mashek MT, Mashek DG, Lefebvre PA (2013) Algal swimming velocities signal fatty acid accumulation. Biotechnol Bioeng 110:143–152
Hondzo M, Wüest A (2009) Do microscopic organisms feel turbulent flows? Environ Sci Technol 43:764–768
Karp-Boss L, Boss E, Jumars PA (1996) Nutrients fluxes to planktonic osmotrophs in the presence of fluid motion. Oceanogr Mar Biol 34:71–107
Klok AJ, Lamers PP, Martens DE, Draaisma RB, Wijffels RH (2014) Edible oils from microalgae: insights in TAG accumulation. Trends Biotechnol 32:521–582
Kumar R, Biswas K, Singh PK, Singh PK, Elumalai S, Shukla P, Pabbi S (2017) Lipid production and molecular dynamics simulation for regulation of accD gene in cyanobacteria under different N and P regimes. Biotechnol Biofuels 10:94
Leupold M, Hindersin S, Gust G, Kerner M, Hanelt D (2013) Influence of mixing and shear stress on Chlorella vulgaris, Scenedesmus obliquus, and Chlamydomonas reinhardtii. J Appl Phycol 25:485–495
Li YT, Han DX, Hu GR, Sommerfeld M, Hu Q (2010) Inhibition of starch synthesis results in overproduction of lipids in Chlamydomonas reinhardtii. Biotechnol Bioeng 107:258–268
Li YT, Han DX, Sommerfeld M, Hu Q (2011) Photosynthetic carbon partitioning and lipid production in the oleaginous microalga Pseudochlorococcum sp. (Chlorophyceae) under nitrogen-limited conditions. Bioresour Technol 102:123–129
Litchman E, de Tezanos Pinto P, Klausmeier CA, Thomas MK, Yoshiyama K (2010) Linking traits to species diversity and community structure in phytoplankton. Hydrobiologia 653:15–28
MacIntyre HL, Kana TM, Geider RJ (2000) The effect of water motion on short-term rates of photosynthesis by marine phytoplankton. Trends Plant Sci 5:12–17
Markou G, Nerantzis E (2013) Microalgae for high-value compounds and biofuels production: a review with focus on cultivation under stress conditions. Biotechnol Adv 31:1532–1542
Merel S, Walker D, Chicana R, Snyder S, Baurès E, Thomas O (2013) State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ Int 59:303–327
Michel G, Tonon T, Scornet D, Cock JM, Kloareg B (2010) Central and storage carbon metabolism of the brown alga Ectocarpus siliculosus: insights into the origin and evolution of storage carbohydrates in eukaryotes. New Phytol 188:67–81
Michels MHA, van der Goot AJ, Vermuë MH, Wijffels RH (2016) Cultivation of shear stress sensitive and tolerant microalgal species in a tubular photobioreactor equipped with a centrifugal pump. J Appl Phycol 28:53–62
Mirón AS, Cerón García MC, Contreras Gómez A, Camacho FG, Grima EM, Chisti Y (2003) Shear stress tolerance and biochemical characterization of Phaeodactylum tricornutum in quasi steady-state continuous culture in outdoor photobioreactors. Biochem Eng J 16:287–297
Moisander PH, Hench JL, Kononen K, Paerl HW (2002) Small-scale shear effects on heterocystous cyanobacteria. Limnol Oceanogr 47:108–119
Nusch EA (1980) Comparison of different methods for chlorophyll and phaeopigment determination. Arch Hydrobiol Beih Ergebn Limnol 14:14–36
Olesen TD, Ganf GG (1986) Photosynthate partitioning: a labile, adaptive phenomenon in Microcystis aeruginosa. Arch Hydrobiol 108:55–76
Oliver RL, Hamilton DP, Brookes JD, Ganf GG (2012) Physiology, blooms and prediction of planktonic cyanobacteria. In: Whitton BA (ed) Ecology of cyanobacteria II. Springer, Dordrecht, pp 155–194
Paerl H, Otten T (2013) Harmful cyanobacterial blooms: causes, consequences, and controls. Microb Ecol 65:995–1010
Pandit PR, Fulekar MH, Karuna MS (2017) Effect of salinity stress on growth, lipid productivity, fatty acid composition, and biodiesel properties in Acutodesmus obliquus and Chlorella vulgaris. Environ Sci Pollut Res Int 24:13437–13451
Peters F, Marrasé C (2000) Effects of turbulence on plankton: an overview of experimental evidence and some theoretical considerations. Mar Ecol Prog Ser 205:291–306
Prairie JC, Sutherland KR, Nickols KJ, Kaltenberg AM (2012) Biophysical interactions in the plankton: a cross-scale review. Limnol Oceanogr Fluids Environ 2:121–145
Quintana N, Van der Kooy F, Van de Rhee MD, Voshol GP, Verpoorte R (2011) Renewable energy from Cyanobacteria: energy production optimization by metabolic pathway engineering. Appl Microbiol Biotechnol 91:471–490
Reis A, Gouveia L, Veloso V, Fernández HL, Empis JA, Novais JM (1996) Eicosapentaenoic acid-rich biomass production by the microalga Phaeodactylum tricornutum in a continuous-flow reactor. Bioresour Technol 55:83–88
Reynolds CS (2006) The ecology of phytoplankton. Cambridge University Press, New York
Rippka R, Deruelles J, Waterbury JB, Herdman M, Stanier RY (1979) Generic assignments, strains histories and properties of pure cultures of cyanobacteria. J Gen Microbiol 111:1–61
Rubio FC, Camacho FG, Fernández Sevilla JM, Chisti Y, Grima EM (2003) A mechanistic model of photosynthesis in microalgae. Biotechnol Bioeng 81:459–473
Sengupta A, Carrara F, Stocker R (2017) Phytoplankton can actively diversify their migration strategy in response to turbulent cues. Nature 543:555–558
Singh S, Verma E, Tiwari B, Niveshika, Mishra AK (2017) Modulation of fatty acids and hydrocarbons in Anabaena 7120 and its ntcA mutant under calcium. J Basic Microbiol 57:171–183
Sobczuk T, Camacho F, Grima E, Chisti Y (2006) Effects of agitation on the microalgae Phaeodactylum tricornutum and Porphyridium cruentum. Bioprocess Biosyst Eng 28:243–250
Steinhoff FS, Karlberg M, Graeve M, Wulff A (2014) Cyanobacteria in Scandinavian coastal waters—a potential source for biofuels and fatty acids? Algal Res 5:42–51
Suali E, Sarbatly R (2012) Conversion of microalgae to biofuel. Renew Sust Energ Rev 16(6):4316–4342
Sullivan JM, Swift E, Donaghay PL, Rines JE (2003) Small-scale turbulence affects the division rate and morphology of two red-tide dinoflagellates. Harmful Algae 2:183–199
Tang JW, Wu QY, Hao HW, Chen Y, Wu M (2004) Effect of 1.7 MHz ultrasound on a gas-vacuolate cyanobacterium and a gas-vacuole negative cyanobacterium. Colloids Surf B 36:115–121
Tang HY, Chen M, Ng KYS, Salley SO (2012) Continuous microalgae cultivation in a photobioreactor. Biotechnol Bioeng 109:2468–2474
Tedesco MA, Duerr EO (1989) Light, temperature and nitrogen starvation effects on the total lipid and fatty acid content and composition of Spirulina platensis UTEX 1928. J Appl Phycol 1:201–209
Thomas WH, Vernet M, Gibson CH (1995) Effects of small-scale turbulence on photosynthesis, pigmentation, cell division, and cell size in the marine dinoflagellate Gomaulax polyedra (Dinophyceae). J Phycol 31:50–59
Walsby AE (1997) Numerical integration of phytoplankton photosynthesis through time and depth in a water column. New Phytol 136:189–209
Wang PL, Shen H, Xie P (2012) Can hydrodynamics change phosphorus strategies of diatoms?—nutrient levels and diatom blooms in lotic and lentic ecosystems. Microb Ecol 63:369–382
Wilbur KM, Anderson NG (1948) Electrometric and colorimetric determination of carbonic anhydrase. J Biol Chem 1715:147–154
Wilhelm C, Jakob T (2011) From photons to biomass and biofuels: evaluation of different strategies for the improvement of algal biotechnology based on comparative energy balances. Appl Microbiol Biotechnol 92:909–919
Xiao Y, Li Z, Li C, Zhang Z, Guo JS (2016) Effect of small-scale turbulence on the physiology and morphology of two bloom-forming cyanobacteria. PLoS One 11(12):e0168925
Yu Z, Song MM, Pei HY, Jiang LQ, Hou QJ, Nie CL, Zhang LJ (2017) The effects of combined agricultural phytohormones on the growth, carbon partitioning and cell morphology of two screened algae. Bioresour Technol 239:87–96
Funding
This work was supported by grants from the National Natural Science Foundation (No. 51779240, 51679226, 51509233), the Chongqing Natural Science Program (cstc2015jcyjBX0006), and Open Foundation by Key Laboratory of Sediments in Yellow River, Ministry of Water Resources. Dr. Yan Xiao is supported by the “Youth Innovation Promotion Association” Program funded by the Chinese Academy of Sciences.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Rights and permissions
About this article

Cite this article
Xiao, Y., Zhang, S., Li, Z. et al. Turbulent mixing mediates the photosynthetic activities and biochemical composition of Anabaena: implications for bioengineering. J Appl Phycol 30, 2227–2236 (2018). https://doi.org/10.1007/s10811-018-1465-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-018-1465-7