
Abstract
The ability of two widely used imidazolium-based ionic liquids (ILs) to affect the physiological behavior of the green alga Dunaliella tertiolecta treated in culture media with different salinities (30 and 35 ‰) was investigated. Algae species were exposed to 1-butyl-3-methylimidazolium tetrafluoroborate ([bmim][BF4]), 1-octyl-3-methylimidazolium tetrafluoroborate ([omim][BF4]), and their binary mixture [bmim][BF4]–[omim][BF4] (ratio 1:1) for 96 h, in f/2 medium with different salinities (30 and 35 ‰). Every 24 h, the growth rate (μ) and the percent inhibition (% I) as well as the concentrations of carotenoids (in terms of carotene content) and chlorophyll a (Chl a) were determined. According to the results, the culture media salinity plays a key role in IL-mediated inhibitory effects on algae. In both salinities, ILs can act against algal growth rate rather than Chl a synthesis, merely due to the existence of a cross-linking with carotenoids, whose enhancement prior or in parallel with Chl a synthesis could play an important role against their growth restriction.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In recent years, increasing awareness of the adverse effects of ionic liquids (ILs) has been observed (Cvjetko Bubalo et al. 2014). In fact, the growing interest for their use in a wide range of applications, such as alternative solvents, organic synthesis, catalytic processes, polymerization reactions, low temperature extractions, electrochemical processes, and devices (Grishina et al. 2013; Wasserscheid and Keim 2000), has raised great concerns about their potential environmental hazards.
ILs are characterized by unique properties, such as high solvency ability for both organic and inorganic compounds, high thermal stability and density, and large electrical conductivity (Freire et al. 2007; Grishina et al. 2013). However, despite the fact that ILs have low vapor pressure at room temperature and were believed to be less toxic than conventional solvents, recently published studies have mentioned that certain ILs are able to cause significant adverse environmental effects, posing thus toxicological risk to nontarget organisms, such as algae (Kulacki and Lamberti 2008; Latała et al. 2009; Ma et al. 2010; Matzke et al. 2007; Pham et al. 2008). Although no indications are available in specialized journals for IL persistence in the environment and their toxicity toward simple living organisms, accidental spills, effluent discharges, etc. could trigger a series of undesirable effects in algal health status as well as to their aquatic ecosystems (Campanella et al. 2000; Ma et al. 2010; Wong 2000).
Algae have a significant role in aquatic ecosystems, because of their key position in the trophic chain, their ability to produce high amounts of oxygen, and their participation in nutrient cycles (DeLorenzo 2009; Ma et al. 2010; Perreault et al. 2012). For this reason, algae are frequently used in environmental studies for assessing the relative toxicity of various chemicals and/or waste discharges (APHA 1989; ASTM 1996; OECD 2011; US EPA 1974). In addition, algal biotests are simple and preferable for ethical and economic reasons, thereby avoiding the need to use experimental animals (Bae and Park 2014). Specifically, a number of ecotoxicological studies, concerning the toxic effects of ILs on algae species, were performed during the last decade, with the use of Chlorella vulgaris, Oocystis submarina, Scenedesmus sp., Chlamydomonas reinhardrii, Chlorella ellipsoidea, and Selenastrum capricornutum (Cho et al. 2007; Kulacki and Lamberti 2008; Latała et al. 2005; 2009; Latała et al. 2010; Ma et al. 2010; Matzke et al. 2007; Ventura et al. 2010; 2013). However, knowledge of the ability of ILs to induce toxic effects to algae is still incomplete, due to the great variety of ILs whose toxic ability may differ depending on the ions of which they are produced, as well as the different algal species sensitivity (Boyle 1984; Perreault et al. 2012). In addition, the knowledge about potential effects on algal pigment production and/or the induction of pre-pathological alterations in IL-exposed cells is still limited.
The green microalga, Dunaliella tertiolecta, a common representative marine and estuarine unicellular flagellate species (Class: Chlorophyceae), fulfills most of the criteria for a bioassay organism (in terms of its cultivation in the laboratory, its rapid growth, and its response to environmental contaminants) and has been proposed as a standard organism for ecotoxicological tests (APHA 1989; ASTM 1996; OECD 2011; US EPA 1974). In fact, species of the genus Dunaliella, such as D. tertiolecta, have been widely used for the investigation of osmoregulation, carotenoid production, and photosynthesis under extreme conditions (reviewed from Oren 2005). A battery of endpoints such as cell density, growth rate, and chlorophyll a could be used for determining functional and structural effects, thus playing an important role in environmental risk assessment and management, especially when other parameters/endpoints are estimated (DeLorenzo 2009). For instance, its ability to induce carotenoid production under stress conditions has made this species a potentially useful model, in order to understand the important role of their formation during algal growth and survival (Nikookar et al. 2005; Oren 2005; Tsiaka et al. 2013). However, no data regarding the role of carotenoids in algae subjected to ILs are available.
Since Latala et al. (2005) reported that IL-mediated effects on algae could be affected by abiotic factors, such as salinity, it was of great interest to investigate the impact of two commonly used imidazolium ILs, such as 1-butyl-3-methylimidazolium tetrafluoroborate ([bmim][BF4]) and/or 1-octyl-3-methylimidazolium tetrafluoroborate ([omim][BF4]) on the current algal species. In order to meet the challenge of making realistic predictions about the effects of ILs on algae, the IL-mediated effects were investigated on algae cultured in media with slight alterations of salinity (30 and 35 ‰). In addition, given that phototrophic algae synthesize not only chlorophylls but also carotenoids, which are bound to peptides to form pigment–protein complexes in the thylakoid membrane (Takaichi 2011), it was of great interest to investigate the concentration range at which ILs alter algal growth and chlorophyll a (Chl a) content, as well as whether alterations of carotenoid (in terms of carotene content) content observed in IL-treated algae could have an important role during algal growth and survival under different stress conditions (Del Campo et al. 2007; Telfer 2002; Tsiaka et al. 2013). Finally, a linear correlation analysis (Pearson test, p < 0.05) between parameters measured in the alga was performed, in order to elucidate any relationship among IL-mediated biological responses in algae, providing thus a more comprehensive view on ILs’ effects on algae cultured in media with different salinities. To our knowledge, the present study is the first to show imidazolium-based IL ability to affect D. tertiolecta growth and survival and the attribution/contribution of photosynthetic and accessory pigments (Chl a and carotenoids, respectively) under different salinities on the obtained IL-mediated effects.
Materials and methods
Chemicals and reagents
1-butyl-3-methylimidazolium tetrafluoroborate ([bmim][BF4]) and 1-octyl-3-methylimidazolium tetrafluoroborate ([omim][BF4]) were purchased from Sigma-Aldrich (USA) (purity ≥97 %). The selected ILs consist of the same anion [BF4 −] and different cation length chain. The choice of the current imidazolium ILs was based on their popularity and application in many scientific fields, as well as the increased likelihood of migrating from industry to the environment. All other reagents used were of high analytical grade and purity.
Stock solutions of ILs
In all experiments, serial dilutions of freshly prepared stock solutions of ILs were used. Each stock was prepared by primarily dissolving the appropriate IL in ultrapure water, double distilled passed through a reverse osmosis system and further purified by Milli-Q quality water system. All stock solutions were kept in the dark at 4 °C, before being used for further analysis.
Algal biotest
IL-mediated effects on D. tertiolecta were investigated according to well-known protocols and guidelines (OECD 2011). In brief, D. tertiolecta (strain CCAP 19/6B, from Scottish Marine Institute, Oban, Argyll, Scotland) was grown in f/2 medium without Si (24 ± 1 °C, pH 8.3 ± 0.3, 86 ± 8.6 μmol photons m−2 s−1 fluorescent lighting) and different salinities (30 and 35 ‰). Different levels of salinity were achieved by the supplementation of proper amounts of NaCl (Aravantinou et al. 2015), while salinities were daily measured with the use of a HQ40 multi-meter (Hach Lange, Germany). The selection of the salinities was based on data obtained by preliminary tests which revealed no alterations of both algal cell number and growth rate; thus, the obtained results would have no interferences due to salinity effects. At late logarithmic phase, an amount of culture (1 × 104 cells mL−1) was transferred to conical sterilized flasks, containing f/2 medium (final volume 200 mL), and finally exposed to different concentrations of [bmim][BF4] and [omim][BF4] and to [bmim][BF4]–[omim][BF4] (dilution ratio 1:1) (0, 1, 10, 100, and 200 mg L−1) for 96 h, under conditions previously mentioned. In this context, it is important to highlight that, due to the absence of data regarding the solubility of each IL in culture media, the concentrations of ILs currently used refer to “nominal” concentrations, thus suggesting that the obtained toxic endpoints (e.g., growth rate) could be over- or underestimated in some extent. Moreover, it should be also mentioned that ILs could probably bind to culture medium compounds, such as serum and proteins, as well as to culture flask walls (Kramer et al. 2012), thus decreasing their free or effective concentration. According to the latter, IL-mediated effects on algal cells were investigated after their exposure to such high IL concentrations.
Every 24 h, the cell number was counted, using a Neubauer hemocytometer, while the growth (μ) and the inhibition rate (% I) were determined according to Eqs. (1) and (2).
where μ n = algal growth rate (day−1) after n days (24, 48, 72, or 96 h), X 0 = number of cells mL−1 at time 0 (t 0), X n = number of cells mL−1 at t n , t 0 = time of first measurement after beginning of test, and t n = time of nth measurement after beginning of test.
where % I = percent inhibition in average specific growth rate, μ c = mean value for average specific growth rate (μ) in the control group, and μ n = average specific growth rate for the treatment replicate.
The aforementioned procedure was repeated four times, and the results are expressed as the mean ± SD from four measurements.
In parallel, the amounts of carotenoids and Chl a were determined in algal cultures exposed to each concentration of the ILs tested in both salinities.
Determination of carotenoids (carotene content)
The determination of carotenoids was performed via quantification of carotenes using the spectrophotometric method initially proposed by Shaish et al. (1992) and later modified (in terms of sampling volume) by Garcia-Gonzalez et al. (2005). In brief, every 24 h, a culture aliquot of 1 mL (containing known number of cells) was centrifuged at 1000×g for 15 min and the cell pellet was extracted with 3 mL of ethanol/hexane (2:1, v/v). Thereafter, 2 mL of water and 4 mL of hexane were added, and the samples were thoroughly mixed, vigorously shaken, and finally centrifuged at 1200×g for 10 min. The hexane layer was separated, and its absorbance at 450 nm was determined: A 450 × 25.2 equals the micrograms of carotene in the sample (Shaish et al. 1992). The results (expressed as pg carotenoids cell−1, based on the known number of cells, previously measured in each sample) are the mean ± SD from four independent measurements in each case.
Determination of Chl a
The content of chlorophyll a (Chl a) was determined according to the method of Aminot and Ray (2000) which is based on the well-established trichromatic method described in the Standard Methods for the Examination of Water and Wastewater (APHA 1996, section 10200H). Specifically, every 24 h, a culture aliquot of 10 mL was filtered through glass microfiber filters (Whatman GF-F, nominal pore size 0.7 μm) and filters were kept at −21 °C till extraction. Extraction of chlorophyll was performed with 10 mL of 90 % v/v acetone, at 4 °C for 5 h. Extracts were separated by centrifugation at 1350×g for 15 min, and absorption was measured at 630, 647, 664, and 750 nm, using acetone as blank. The levels of Chl a were calculated according to the following equation:
The results (expressed as pg of Chl a per cell) are mean ± SD from four independent measurements in each case.
Statistical analysis
The estimation of IC50 endpoints and their 95 % confidence intervals (CI) in each case was performed with the use of probit analysis (p < 0.05, IBM SPSS 19 Inc. software package). In some cases, 95 % CI could not be calculated due to high variability of the algal response. After checking for homogeneity of variance (Levene’s test of equality of error variances), the significant differences among parameters were tested with the use of Mann–Whitney U test (p < 0.05). Univariate analysis (two-way ANOVA, p < 0.05) was performed for estimating the effect of salinity and/or incubation period and IL concentration on algal growth rate with time. Simple linear correlation (Pearson test, p < 0.05) analysis was conducted with the mean values of each parameter tested for investigating significant relationships among the biological responses obtained in both salinities.
Results
The effect of algal culture medium salinity on algal cells
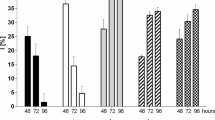
The physiological behavior of algal species in terms of growth rate, carotenoid content, and chlorophyll a (Chl a) cultivated in media with slight alterations of salinity was investigated. Despite the different cell numbers at 24 h, algae incubated in culture media with 30 and 35 ‰ of salinity followed similar growth rates over time (48, 72, and 96 h) (Fig. 1a). Moreover, almost similar trends of carotenoids and Chl a content were observed in both cases (Fig. 1b, c). On the other hand, although salinity did not seem to affect algal growth rate over time (4 days of incubation period) (Table 1(A)), it was significantly related with algal growth rates obtained at almost all cases of IL-treated cells (two-way ANOVA, p < 0.05) (Table 1(B)).

Growth rate (a), carotenoids (in terms of carotene content) (b), and Chl a (c) content in Dunaliella tertiolecta incubated in culture media with different salinities (30 and 35 ‰). Carotenoids and/or Chl a expressed as pg cell−1 (based on the known number of cells, previously measured in each sample). The results are mean ± SDs from four independent experiments in each case. Values that share the same letter represent significant difference among cells cultivated in culture media with different salinities (Mann–Whitney U test, p < 0.05)
[bmim][BF4]- and/or [omim][BF4]-mediated toxicity on D. tertiolecta
Algae treated with [bmim][BF4], or [omim][BF4], as well as with a binary mixture of [bmim][BF4]–[omim][BF4], showed differences in their growth rates. Specifically, a slight but significant growth arrest was observed in algae treated with [bmim][BF4] in either 30 or 35 ‰ culture media for 24–96 h, compared with the respective values of control cells in each case (Fig. 2a). Regarding [bmim][BF4]-mediated inhibitory effect in both salinities, there was no significant difference over time (for more details SM Figure 2.1A), as also shown by the IC50 values (85.49 mg L−1 at 24 h and >200 mg L−1 at the rest of the incubation period) in each case (Table 2). On the other hand, algae treated with [omim][BF4] showed significantly lower growth rates than the [bmim][BF4]-treated algae, thus reaching an almost total growth arrest in case of the higher concentration of [omim][BF4] (200 mg L−1) tested in both salinities (Fig. 2b, SM Figure 2.1B). The latter was further reinforced by the significantly high inhibition rate values (% I, see also SM Figure 2.1B) and the concomitant low [omim][BF4] IC50 values currently observed; the lower IC50 values observed in algae treated with [omim][BF4] in 30 ‰ medium for 24 and 48 h were attenuated over time (72 and 96 h), while opposite results were obtained in case of algae treated with [omim][BF4] in 35 ‰ medium (Table 2, see also SM Table 2.1 for number of cells in each case). In case of [bmim][BF4]–[omim][BF4], significantly lower values of growth rate and increased inhibition rates (in terms of both % I and IC50 values) were observed in algae incubated in 30 ‰ medium, compared to those incubated in 35 ‰ culture in all cases (Fig. 2c, Table 2, see also SM Figure 2.1C).

Growth rate (μ, day−1) of algae treated with a [bmim][BF4], b [omim][BF4], and c [bmim][BF4]–[omim][BF4] in culture media with different salinities (30 and 35 ‰, respectively). The results are mean ± SDs from four independent experiments in each case. Number sign (#) indicates 100 % algal growth arrest. Values with
 differ significantly from the respective control in each case (Mann–Whitney U test, p < 0.05).
differ significantly from the respective control in each case (Mann–Whitney U test, p < 0.05).
Carotenoid content
Algae treated with each IL showed IL’s type-, dose- and time-dependent variations of carotene content in both culture media. Specifically, a slight but significantly higher content of carotene was observed in algae treated in 30 ‰ medium with high concentrations of [bmim][BF4] (100 and 200 mg L−1), while no alterations (p > 0.05) were observed in case of algae treated with the respective IL in 35 ‰ medium (Fig. 3a). On the other hand, excessive amounts of carotenes were determined in algae treated with the aforementioned concentrations of [omim][BF4] in 35 ‰ medium, compared with those observed in both [bmim][BF4]- and [bmim][BF4]–[omim][BF4]-treated algae (Fig. 3b). Algae treated with [bmim][BF4]–[omim][BF4] in 30 ‰ medium showed elevated levels of carotenes compared to those measured in algae treated in 35 ‰ medium, in all cases (Fig. 3c).

Carotenoid content (in terms of carotenes expressed as pg cell−1) in algal cells treated with a [bmim][BF4], b [omim][BF4], and c [bmim][BF4]–[omim][BF4] (dilution ratio 1:1) for 24, 48, 72, and 96 h, in culture media with different salinities (30 and 35 ‰). Values with asterisk and  differ significantly from the respective control in each case (Mann–Whitney U test, p < 0.05, n = 4)
differ significantly from the respective control in each case (Mann–Whitney U test, p < 0.05, n = 4)
Chlorophyll a
The exposure of algae to different concentrations of each IL showed significant alterations in the levels of Chl a measured in each cell. Specifically, algae treated with at least 10, 100, and 200 mg L−1 of [bmim][BF4] for 24 and 72 h in 30 ‰ medium showed significantly higher levels of Chl a than those in algae treated with the respective IL concentrations in 35 ‰ medium, while the opposite was observed at 48 and 96 h (Fig. 4a). On the other hand, elevated values of Chl a were measured in algae treated with each concentration of [omim][BF4] in 30 ‰ medium for 24 and 48 h. Similarly, Chl a levels were high in algae treated with 10, 100, and 200 mg L−1 of [omim][BF4] in 35 ‰ medium for 24 h. In both cases, Chl a levels showed a significant attenuation over time (72 and 96 h), except in the case of algae treated with 100 and 200 mg L−1 of [omim][BF4] in 30 ‰ medium for 96 h (Fig. 4b). In the case of [bmim][BF4]–[omim][BF4], significantly higher values of Chl a content were observed in algae treated with 10, 100, and 200 mg L−1 of the binary IL mixture in 30 ‰ medium, compared with the respective values measured in control cells in any case, while similar trend of Chl a content increase was observed only at higher concentrations of the ILs, in case of algae treated in 35 ‰ medium (Fig. 4c).

Determination of chlorophyll a (Chl a, expressed as pg cell−1) content in algal cells treated with a [bmim][BF4], b [omim][BF4], and c [bmim][BF4]–[omim][BF4] (dilution ratio 1:1) for 24, 48, 72, and 96 h, in culture media with different salinities (30 and 35 ‰). Values with asterisk and  differ significantly from the respective control in each case (Mann–Whitney U test, p < 0.05, n = 4)
differ significantly from the respective control in each case (Mann–Whitney U test, p < 0.05, n = 4)
Correlations among biological parameters
In order to investigate whether there is any relationship among biological responses in IL-exposed algae, a linear correlation analysis (Pearson test, p < 0.05) was performed (Table 3(A, B)). A strong negative correlation was observed between growth rate and Chl a content measured in algae treated with either [omim][BF4] or [bmim][BF4]–[omim][BF4] in both salinities, as well as in algae treated with [bmim][BF4] in 35 ‰ medium. Similarly, algal growth rate and carotene content showed a significantly negative correlation, while opposite results (positive correlations) were obtained between carotenes and Chl a levels in case of algae treated with either [omim][BF4] or [bmim][BF4]–[omim][BF4] in 30 ‰ medium.
Discussion
Given that algal growth and protective system response over time could differ when being subjected to different salinities (Ben-Amotz et al. 2009), the results of the present study showed that slight, but significant, alterations of salinity could affect IL-mediated effects on D. tertiolecta. In fact, D. tertiolecta was more vulnerable to [omim][BF4] than [bmim][BF4] in both salinities, while algae treated with a binary mixture of [bmim][BF4] and [omim][BF4] in 30 ‰ medium revealed significantly higher growth arrest than those occurred in case of their treatment in 35 ‰ medium. Although there are a lot of studies regarding the inhibitory effects of ILs on green algae, the adverse effects of their mixtures remain still unknown. The latter becomes more complicated to be explained, since IL conjugated action, ion paring, and/or clustering as well as synergistic effects, which could play an important role on the observed IL adverse effects, are not easy to predict (Cvjetko Bubalo et al. 2014; Petkovic et al. 2011; Tsarpali and Dailianis 2015; Ventura et al. 2013).
The different extent of [bmim][BF4] and/or [omim][BF4] effects on algae species could be related to the ILs’ different length of alkyl chain (Bernot et al. 2005; Pham et al. 2010). In fact, most ILs show similar structural and functional properties with pesticides and antibiotics (Docherty and Kulpa 2005; Li et al. 2012) and their mode of action takes place through membrane disruption. Specifically, [omim][BF4] with longer alkyl chain than [bmim][BF4] is characterized by higher lipophilicity (Ma et al. 2010; Ranke et al. 2003; Stepnowski et al. 2004), a fact that could increase its interaction with cell membrane phospholipid bilayers and hydrophobic domains of membrane proteins, thus enhancing more toxic effects than [bmim][BF4], as revealed by the results of the present study. On the other hand, the contribution of [BF4 −] anions on the obtained ILs’ effects should not be excluded. Specifically, [BF4 −] anion was reported to be less stable and more toxic than other anions, such as [DCNA−], [TFMS−], [Br−], and [Cl−], due to the formation of strong hydrogen bonds between water and its atom (Latała et al. 2009; Pham et al. 2010; Samori et al. 2010; Shimizu et al. 2010; Zhang et al. 2011). Given that [BF4 −] hydrolysis rate could be indirectly affected by the cation’s chain length (Cammarata et al. 2001; Saha and Hamaguchi 2006), [omim][BF4], having longer alkyl chain than [bmim][BF4], could merely decrease the cation–anion interaction strength, thus extending anion hydrolysis and the obtained toxicity in algae (Freire et al. 2007). However, more studies should be performed for determining the role of ILs’ anions on the obtained effects, especially in algal species.
Although previous studies have reported a significant attenuation of imidazolium IL-mediated toxic effects on the freshwater species O. submarina and C. vulgaris, following an increase of salinity (Kulacki and Lamberti 2008; Latała et al. 2005; 2010), the present study showed that this is not always the case. In fact, it has been reported that various abiotic and biotic parameters, such as differences in algal sensitivity and response to stress conditions, the potential interference of the culture media with the tested ILs, and ILs’ dilution to each algal species culture medium (freshwater-saltwater), could differentiate the toxic behavior of ILs (DeLorenzo 2009; Kulacki and Lamberti 2008; Latała et al. 2009; Oukarroum et al. 2012; Aravantinou et al. 2015). D. tertiolecta, in contrast to O. submarina and C. vulgaris, does not have a cell wall which could reduce the permeability of IL cations (Latała et al. 2005), a fact that could affect algal vulnerability to ILs. The latter should be further investigated in a more detailed/extended study, due to difficulties in measuring IL nominal and initial concentrations, as well as potential metabolites, because of the interferences of both culture media and their salinity values with the obtained results.
According to the results of the present study, algal cells treated in 30 ‰ medium with [omim][BF4] or [bmim][BF4]–[omim][BF4] mixture showed a significant relationship among growth arrest/inhibition with the levels of carotenoids, at least in case of carotene content, and Chl a within algal cells. Specifically, the algal growth arrest attenuation over time, after exposure to ILs, was followed by a time-dependent increase of the Chl a content within algal cells, suggesting that the tested ILs cannot act against Chl a synthesis in algal cells. Similar results were also obtained in algal cells treated within 35 ‰ medium, with [bmim][BF4], [omim][BF4], or [bmim][BF4]–[omim][BF4] mixture in any case. The latter seemed to be more evident in the case of [omim][BF4] and [bmim][BF4]–[omim][BF4] mixture, which showed to be more toxic than [bmim][BF4] in both culture media conditions (salinity levels 30 and 35 ‰). Although the exact mechanism is not known, previous studies have reported that levels of Chl a within cells may increase under stress conditions (El-Sheekh et al. 2003; Nikookar et al. 2005; Tsiaka et al. 2013), probably via the existence of a cross-linking among carotenoids and photosynthetic pigments. The latter is based on the fact that carotenoids could merely serve as a protective mechanism during phototropic algal growth (Cantrell et al. 2003; Jeffrey and Skarstad 2009; Kumar et al. 2010; Melegari et al. 2013; Niyogi 2000; Telfer 2002). Actually, the role of carotenoids against chemical stressor ability to mediate algal growth rate over time, as well as their ability to act as antioxidant molecules, is well documented (Cantrell et al. 2003; Jeffrey and Skarstad 2009; Kumar et al. 2010; Li et al. 2005; Melegari et al. 2013; Telfer 2002; Tsiaka et al. 2013; Woodall et al. 1997). According to this and bearing in mind ILs’ ability to induce oxidative stress (for more details, see Kumar et al. 2011), the high levels of carotenoids measured in both [omim][BF4] and [bmim][BF4]–[omim][BF4] mixture-treated cells could reveal the protective role of these molecules against the oxidative-like events of ILs. In fact, since there is no data regarding the detection of 9-cis isomer which seemed to be more efficient in protecting against oxidative damage (Jimenez and Pick 1993), more studies are needed for elucidating the aforementioned thesis. Moreover, taking into account that ILs with longer alkyl chain could interfere with the cell membranes, thus leading to oxidative stress by-products, such as lipid peroxides, the enhancement of carotenoids could play an important role for the restriction of such phenomenon, via their reaction with free radicals and their ability to terminate chain reactions of lipid peroxidation by reacting with its products (Kumar et al. 2010; Melegari et al. 2013; Niyogi 2000; Woodall et al. 1997).
Despite the absence of data regarding IL ability to cause oxidative-like effects on D. tertiolecta, the significant correlations occurred between carotene content, growth rate, and/or Chl a content measured at least in algae treated in 30 ‰ medium with either [omim][BF4] or [bmim][BF4]–[omim][BF4] mixture could give further evidence for the presence of such a mechanism in IL-treated algae. On the other hand, the existence of a cross-linking among carotenes and photosynthetic pigments should not be excluded for the case of algae exposed to ILs in 35 ‰ medium. In this context, the absence of any direct correlation between carotene content enhancement and Chl a within cells could be due to a time delay response of Chl a or due to differences in IL-mediated individual, synergistic or antagonistic effects on algae pigments, which could regulate their production. Since the latter is only a hypothesis, more studies are needed in order to clearly elucidate the exact mechanism being involved in the enhancement of those pigments in D. tertiolecta faced with imidazolium ILs under slight variations of abiotic conditions, such as salinity.
In conclusion, the present study showed that [bmim][BF4] and [omim][BF4], as well as their binary mixture, could differentially affect the physiological behavior of the green alga D. tertiolecta, incubated/treated in culture media with slight but significant alterations of salinity (30 and 35 ‰). It was shown that imidazolium-based ILs can act against algal growth rate rather than Chl a synthesis, merely due to the existence of a cross-linking with carotenoids, whose enhancement prior or at least in parallel with Chl a could play an important role against the restriction of Chl a synthesis in challenged algal cells. Alterations of the tested parameters under different salinities could be due to differences in algae’s sensitivity and/or response to stress conditions, as well as to potential interference of the culture media with the tested ILs. Although the obtained significant correlations between the growth rate, carotene, and Chl a content in algal cells treated with the tested ILs further reinforce the aforementioned conclusive remark, more studies should be carried out in order to clearly elucidate the role of Chl a and carotenoids under stress conditions and the existence of either individual or synergistic/antagonistic effects among ILs.
References
Aminot A, Ray F (2000) Standard procedures for the determination of chlorophyll a by spectroscopic methods. ICES Tech Mar Environ Sci 30:1–18
APHA (1989) Toxicity testing with phytoplankton. Standard methods for examination of water and wastewater, 17th Ed, Suppl. Washington, DC, USA.
APHA (1996) Standard methods for the examination of water and wastewater. 20th Ed. Am. Pub. Health Assoc, Washington, DC. USA
Aravantinou AF, Tsarpali V, Dailianis S, Manariotis ID (2015) Effect of cultivation media on the toxicity of ZnO nanoparticles to freshwater and marine microalgae. Ecotox Environ Safe 114:109–116
ASTM (American Society for Testing and Materials) (1996) Standard guide for conducting static 96-h toxicity tests with microalgae. Vol. 11.05. ASTM, West Conshohocken, PA. USA
Bae MJ, Park YS (2014) Biological early warning system based on the responses of aquatic organisms to disturbances: a review. Sci Total Environ 466–467:635–649
Ben-Amotz A, Polle JEW, Subba Rao DV (eds) (2009) The alga Dunaliella: biodiversity, physiology, genomics and biotechnology. Science Publishers, Enfield
Bernot RJ, Kennedy EE, Lamberti GA (2005) Effects of ionic liquids on the survival, movement and feeding behavior of the freshwater snail, Physa acuta. Environ Toxicol Chem 24:1759–1765
Boyle TP (1984) The effect of environmental contaminants on aquatic algae. In: Shubert LE (ed) Algae as ecological indicators. Academic Press, New York, pp 237–256
Cammarata L, Kazarian SG, Salterb PA, Welton T (2001) Molecular states of water in room temperature ionic liquids. Phys Chem Chem Phys 3:5192–5200
Campanella L, Cubadda F, Sammartino MP, Saoncella A (2000) An algal biosensor for the monitoring of water toxicity in estuarine environments. Water Res 25:69–76
Cantrell A, McGarvey DJ, Truscott TG, Rancan F, Böhm F (2003) Singlet oxygen quenching by dietary carotenoids in a model membrane environment. Arch Biochem Biophys 412:47–54
Cho CW, Pham TPT, Jeon YC, Vijayaraghavan K, Choed WS, Yun YS (2007) Toxicity of imidazolium salt with anion bromide to a phytoplankton Selenastrum capricornutum: effect of alkyl-chain length. Chemosphere 69:1003–1007
Cvjetko Bubalo MC, Radošević K, Redovniković IR, Halambek J, Srček VG (2014) A brief overview of the potential environmental hazards of ionic liquid. Ecotoxicol Environ Saf 99:1–12
Del Campo JA, Garcia-Gonzalez M, Guerrero MG (2007) Outdoor cultivation of microalgae for carotenoid production: current state and perspectives. Appl Microbiol Biot 74:1163–1174
DeLorenzo AM (2009) Utility of Dunaliella in ecotoxicity testing. In: Ben-Amotz A, Polle JEW, Rao DVS (eds) The alga Dunaliella: biodiversity, physiology, genomics and biotechnology. Science Publishers, Enfield, pp 495–512
Docherty KM, Kulpa CFJ (2005) Toxicity and antimicrobial activity of imidazolium and pyridinium ionic liquids. Green Chem 7:185–189
El-Sheekh MM, El-Naggar AH, Osman MEH, El-Mazaly E (2003) Effect of cobalt on growth, pigments and the photosynthetic electron transport in Monoraphidium minutum and Nitzchia perminuta. Braz J Plant Physiol 15:159–166
Freire MG, Santos LMNBF, Fernandes AM, Coutinho JAP, Marrucho IM (2007) An overview of the mutual solubilities of water–imidazolium-based ionic liquids systems. Fluid Phase Equilibr 261:449–454
Garcia-Gonzalez M, Moreno J, Manzano JC, Florencio FJ, Guerrero MG (2005) Production of Dunaliella salina biomass rich in 9-cis-β-carotene and lutein in a closed tubular photobioreactor. J Biotechnol 115:81–90
Grishina EP, Ramenskaya LM, Gruzdev MS, Kraeva OV (2013) Water effect on physicochemical properties of 1-butyl-3-methylimidazolium based ionic liquids with inorganic anions. J Mol Liq 177:267–272
Jeffrey SW, Skarstad E (2009) Pigments of green and red forms of Dunaliella, and related chlorophytes. In: Ben-Amotz A, Polle JEW, Rao DVS (eds) The alga Dunaliella: biodiversity, physiology, genomics and biotechnology. Science Publishers, Enfield, pp 111–145
Jimenez C, Pick U (1993) Differential reactivity of β-carotene isomers from Dunaliella bardawil toward oxygen radicals. Plant Physiol 101:385–390
Kramer NI, Krismartina M, Rico-Rico A, Blaauboer BJ, Hermens JLM (2012) Quantifying processes determining the free concentration of phenanthrene in basal cytotoxicity assay. Chem Res Toxicol 25:436–445
Kulacki KJ, Lamberti GA (2008) Toxicity of imidazolium ionic liquids to freshwater algae. Green Chem 10:104–110
Kumar M, Kumari P, Gupta V, Anisha PA, Reddy CRK, Jha B (2010) Differential responses to cadmium induced oxidative stress in marine macroalga Ulva lactuca (Ulvales, Chlorophyta). Biometals 23:315–325
Kumar M, Trivedi N, Reddy CRK, Jha B (2011) Toxic effects of imidazolium ionic liquids on the green seaweed Ulva lactuca: oxidative stress and DNA damage. Chem Res Toxicol 24:1882–1890
Latała A, Stepnowski P, Nędzi M, Mrozik W (2005) Marine toxicity assessment of imidazolium ionic liquids: acute effects on the Baltic algae Oocystis submarina and Cyclotella meneghiniana. Aquat Toxicol 73:91–98
Latała A, Nędzi M, Stepnowski P (2009) Toxicity of imidazolium and pyridinium based ionic liquids towards algae. Chlorella vulgaris, Oocystis submarina (green algae) and Cyclotella meneghiniana, Skeletonema marinoi (diatoms). Green Chem 11:580–588
Latała A, Nędzi M, Stepnowski P (2010) Toxicity of imidazolium ionic liquids towards algae. Influence of salinity variations. Green Chem 12:60–64
Li X, Ping X, Xiumei P, Zhenbin W, Liqiang X (2005) Toxicity of cypermethrin on growth, pigments, and superoxide dismutase of Scenedesmus obliquus. Ecotoxicol Environ Saf 60:188–192
Li H, Yu C, Chen R, Li J, Li J (2012) Novel ionic liquid-type Gemini surfactants: synthesis, surface property and antimicrobial activity, colloids and surfaces. Colloids Surf A 395:116–124
Ma JM, Cai LL, Zhang BJ, Hu LW, Li XY, Wang JJ (2010) Acute toxicity and effects of 1-alkyl-3-methylimidazolium bromide ionic liquids on green algae. Ecotoxicol Environ Saf 73:1465–1469
Matzke M, Stolte S, Thiele K, Juffernholz T, Arning J, Ranke J, Welz-Biermann U, Jastorff B (2007) The influence of anion species on the toxicity of 1-alkyl-3-methylimidazolium ionic liquids observed in an (eco)toxicological test battery. Green Chem 9:1198–1207
Melegari SP, Perreault F, Costa RHR, Popovic R, Matias WG (2013) Evaluation of toxicity and oxidative stress induced by copper oxide nanoparticles in the green alga Chlamydomonas reinhardtii. Aquat Toxicol 142–143:431–440
Nikookar K, Moradshahi A, Hosseini L (2005) Physiological responses of Dunaliella salina and Dunaliella tertiolecta to copper toxicity. Biomol Eng 22:141–146
Niyogi KK (2000) Safety valves for photosynthesis. Curr Opin Plant Biol 3:455–460
OECD (2011) Test no. 201: freshwater alga and cyanobacteria, growth inhibition test, OECD Guidelines for the Testing of Chemicals, Section 2, OECD Publishing, Paris. DOI: http://dx.doi.org/10.1787/9789264069923-en
Oren A (2005) A hundred years of Dunaliella research: 1095-2005. Sal Systems 1:2. doi:10.1186/1746-1448-1-2
Oukarroum A, Bras S, Perreault F, Popovic R (2012) Inhibitory effects of silver nanoparticles in two green algae, Chlorella vulgaris and Dunaliella tertiolecta. Ecotoxicol Environ Saf 78:80–85
Perreault F, Matias MS, Oukarroum A, Matias WG, Popovic R (2012) Okadaic acid inhibits cell growth and photosynthetic electron transport in the alga Dunaliella tertiolecta. Sci Total Environ 414:198–204
Petkovic M, Seddon KR, Rebelo LPN, Silva Pereira C (2011) Ionic liquids: a pathway to environmental acceptability. Chem Soc Rev 40:1383–1403
Pham TPT, Cho CW, Min J, Yun YS (2008) Alkyl-chain length effects of imidazolium and pyridinium ionic liquids on photosynthetic response of Pseudokirchneriella subcapitata. J Biosci Bioeng 105:428–428
Pham TPT, Cho CW, Yun YS (2010) Environmental fate and toxicity of ionic liquids: a review. Water Res 44:352–372
Ranke J, Mőlter K, Stock F, Bottin-Weber U, Poczobutt J, Hoffmann J, Ondruschka B, Filser J, Jastorff B (2003) Biological effects of imidazolium ionic liquids with varying chain lengths in acute Vibrio fischeri and WST-1 cell viability assays. Ecotoxicol Environ Saf 28:396–404
Saha S, Hamaguchi HO (2006) Effect of water on the molecular structure and arrangement of nitrile-functionalized ionic liquids. J Phys Chem B 110:2777–2781
Samori C, Malferrari D, Valbonesi P, Montecavalli A, Moretti F, Galletti P, Sartor G, Tagliavini E, Fabbri E, Pasteris A (2010) Introduction of oxygenated side chain into imidazolium ionic liquids: evaluation of the effects at different biological organization levels. Ecotoxicol Environ Saf 73:1456–1464
Shaish A, Ben-Amotz A, Avron M (1992) Biosynthesis of β-carotene in Dunaliella. Methods Enzymol 213:439–444
Shimizu K, Gomes MFC, Pádua AAH, Rebelo LPN, Lopes JNC (2010) Three commentaries on the nano-segregated structure of ionic liquids. J Mol Struc-Theochem 946:70–76
Stepnowski P, Składanowski AC, Ludwiczak A, Łączyńska E (2004) Evaluating the cytotoxicity of ionic liquids using human cell line HeLa. Hum Exp Toxicol 23:513–517
Takaichi S (2011) Carotenoids in algae: distributions, biosyntheses and functions. Mar Drugs 9:1101–1118
Telfer A (2002) What is β-carotene doing in the photosystem II reaction centre? Phil Trans R Soc B 357:1431–1439
Tsarpali V, Dailianis S (2015) Toxicity of two imidazolium ionic liquids, [bmim][BF4] and [omim][BF4], to standard aquatic test organisms: role of acetone in the induced toxicity. Ecotoxicol Environ Saf 117:62–71
Tsiaka P, Tsarpali V, Ntaikou I, Kostopoulou MN, Lyberatos G, Dailianis S (2013) Carbamazepine-mediated pro-oxidant effects on the unicellular marine algal species Dunaliella tertiolecta and the hemocytes of mussel Mytilus galloprovincialis. Ecotoxicology 22:1208–1220
U.S. EPA (U.S. Environmental Protection Agency) (1974) Marine algal assay procedure bottle test: eutrophication and lake restoration. Branch National Environmental Research Center, Corvallis, OR, USA
Ventura SPM, Gonçalves AMM, Gonçalves F, Coutinho JAP (2010) Assessing the toxicity on [C3mim][Tf2N] to aquatic organisms of different trophic levels. Aquat Toxicol 96:290–297
Ventura SPM, Gurbisz M, Ghavre M, Ferreira FMM, Gonçalves F, Beadham I, Quilty B, Coutinho JAP, Gathergood N (2013) Imidazolium and pyridinium ionic liquids from mandelic acid derivatives: synthesis and bacteria and algae toxicity evaluation. ACS Sustain Chem Eng 1:393–402
Wasserscheid P, Keim W (2000) Ionic liquids-new “solutions” for transition metal catalysis. Angew Chem Int Ed 39:3772–3789
Wong PK (2000) Effects of 2, 4-D, glyphosate and paraquat on growth, photosynthesis and chlorophyll-a synthesis of Scenedesmus quadricauda Berb 614. Chemosphere 4:177–182
Woodall AA, Britton G, Jackson MJ (1997) Carotenoids and protection of phospholipids in solution or in liposomes against oxidation by peroxyl radicals: relationship and protective ability. Biochim Biophys Acta 7:617–635
Zhang C, Malhotra SV, Francis AJ (2011) Toxicity of imidazolium- and pyridinium-based ionic liquids and the co-metabolic degradation of N-ethylpyridinium tetrafluoroborate. Chemosphere 82:1690–1695
Acknowledgments
This study was supported by the annual research grant sanctioned to the Section of Animal Biology by the University of Patras, Greece. The present work is included in the doctoral thesis of Ms. V. Tsarpali. Further data associated with the current article can be found within the supplementary file.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 114 kb)
Rights and permissions
About this article

Cite this article
Tsarpali, V., Harbi, K. & Dailianis, S. Physiological response of the green microalgae Dunaliella tertiolecta against imidazolium ionic liquids [bmim][BF4] and/or [omim][BF4]: the role of salinity on the observed effects. J Appl Phycol 28, 979–990 (2016). https://doi.org/10.1007/s10811-015-0613-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-015-0613-6