
Abstract
For arboreal primates, ground use may increase dispersal opportunities, tolerance to habitat change, access to ground-based resources, and resilience to human disturbances, and so has conservation implications. We collated published and unpublished data from 86 studies across 65 localities to assess titi monkey (Callicebinae) terrestriality. We examined whether the frequency of terrestrial activity correlated with study duration (a proxy for sampling effort), rainfall level (a proxy for food availability seasonality), and forest height (a proxy for vertical niche dimension). Terrestrial activity was recorded frequently for Callicebus and Plecturocebus spp., but rarely for Cheracebus spp. Terrestrial resting, anti-predator behavior, geophagy, and playing frequencies in Callicebus and Plecturocebus spp., but feeding and moving differed. Callicebus spp. often ate or searched for new leaves terrestrially. Plecturocebus spp. descended primarily to ingest terrestrial invertebrates and soil. Study duration correlated positively and rainfall level negatively with terrestrial activity. Though differences in sampling effort and methods limited comparisons and interpretation, overall, titi monkeys commonly engaged in a variety of terrestrial activities. Terrestrial behavior in Callicebus and Plecturocebus capacities may bolster resistance to habitat fragmentation. However, it is uncertain if the low frequency of terrestriality recorded for Cheracebus spp. is a genus-specific trait associated with a more basal phylogenetic position, or because studies of this genus occurred in pristine habitats. Observations of terrestrial behavior increased with increasing sampling effort and decreasing food availability. Overall, we found a high frequency of terrestrial behavior in titi monkeys, unlike that observed in other pitheciids.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The ability or propensity of arboreal primates to use the ground varies widely among species (Napier and Napier 1967; Wu 1993; Wu et al.1988). For some primates, this behavior appears to be linked to a species’ capacity to disperse between forest fragments by crossing roads or open and disturbed areas, to gain access to vital resources, such as fruit and water, or to a reduction in predation risk associated with the long-lasting presence of human observers (Ancrenaz et al.2014; Anderson et al.2007; Barnett et al.2012a; Campbell et al.2005; Cheyne et al.2018; Eppley et al.2016; Grueter et al.2009; Mourthé et al.2007; Nowak et al.2014; Tabacow et al.2009). In this sense, ground use potentially enhances species resilience and its long-term persistence in fragmented landscapes (Jones 2005).
Lower canopy forests are likely to provide less food for upper canopy specialists and result in a closer proximity to the ground compared to more stratified forest (Takemoto 2004). Furthermore, by using the ground arboreal primates can expand their niche, allowing them access to a greater diversity of resources (Boyer et al.2006; Mesa-Sierra and Pérez-Torres 2017). Strata use is also linked to body size. Small-bodied primates usually concentrate their activities in the lower and middle levels of the forest, thus increasing the chance that ground use will occur (Fleagle 1999). For example, ground use is more common in species of Pithecia, which use the forest understory more often and have less specialized diets than in the larger-bodied species of Chiropotes and Cacajao (Barnett et al.2012a; Boyle et al.2015).
Increases in study duration and the length of time devoted to fieldwork activities by researchers (e.g., increased sampling effort) can increase the probability of detecting rare events and unusual behaviors, such as terrestriality (Weatherhead 1986). For example, white-faced sakis (Pithecia pithecia) showed high rates of terrestrial behavior during systematic monitoring at Isla Redonda, Lago Guri, Venezuela, but low rates in shorter-term studies (see Table III in Barnett et al.2012a). Similarly, spider monkeys (Ateles geoffroyi) showed high rates of terrestrial behavior per hour during ca. 2000 h of monitoring, but low rates in ca. 500 h of monitoring (Table I in Campbell et al.2005). Although other factors may influence ground use (such as geophagy and drinking water in spider monkeys: Campbell et al.2005), testing the influence of the extent of sampling effort would improve our understanding of terrestrial behavior in primates.
Despite the absence of morphological specializations for terrestriality (Aversi-Ferreira et al.2013), ground use is widespread in Neotropical primates as an occasional, although potentially important, part of their behavioral repertoire. The frequency, duration, and context of terrestriality can vary substantially between Neotropical primate species, and such behavior is relatively more common in genera such as Cebus and Sapajus (Ottoni and Izar 2008; Porfírio et al.2017), and rarer in such genera as Cacajao, Chiropotes, and Pithecia (Barnett et al., 2012a) (Table I). As in other species, the availability of arboreal food resources and forest strata potentially influences the nature and extent of terrestrial behaviors in Neotropical primates (Campbell et al.2005; Cant 1992).
Among species in the family Pitheciidae, terrestrial behavior in pitheciines (Cacajao, Chiropotes, and Pithecia) is unusual and almost completely restricted to the exploitation of alternative food resources when the availability of highly used arboreal items is low (Barnett et al.2012a). In addition, it can differ substantially among genera, field sites and populations (Barnett et al.2012a). Variation between study sites is compatible with the hypothesis that local variables, such as food availability, predator density, and traditions influence terrestrial activities and their frequencies (Barnett et al.2012a, 2013). In another pitheciid group, the titi monkeys, the frequency of terrestrial behavior, and variables that potentially contribute to this behavior, remain unknown.
Titi monkeys (Pitheciidae, Callicebinae) are small-bodied (ca. 1 kg) platyrrhines that live in groups of two to five individuals, typically including a male–female adult pair and their offspring (Norconk 2011). Previously included in a single genus, Callicebus, titi monkeys were recently split into three genera (Callicebus, Cheracebus, and Plecturocebus) based on phylogenetic and divergence-time analyses using molecular data (Byrne et al.2016), and in accordance with long-recognized geographically based species groupings (van Roosmalen et al.2002). The 35 species currently described (Boubli et al.2019; Byrne et al.2016; Serrano-Villavicencio et al.2017; van Roosmalen et al.2002) are distributed throughout Amazonia into Paraguay, with a discontinuous distribution in eastern Brazil. Titi monkeys inhabit a variety of habitats, ranging from Andean premontane forests (e.g., Plecturocebus oenanthe: Bóveda-Penalba et al.2009), lowland rainforests (e.g., Plecturocebus lucifer: Kinzey et al.1977), and dry semideciduous forests (e.g., Callicebus barbarabrownae: Printes et al.2011), to semiarid Chaco forests (e.g., Plecturocebus pallescens: Rumiz 2012). Many species occur in landscapes severely fragmented by human activities (e.g., Callicebus coimbrai: Chagas and Ferrari 2010; Plecturocebus moloch: Michalski and Peres 2005; Plecturocebus oenanthe: Bóveda-Penalba et al.2009; Shanee et al.2011; Plecturocebus grovesi: Boubli et al.2019). The fruit-based diet of titi monkeys is seasonally complemented with leaves, seeds, flowers, animal prey, or other items depending on the species and population (Bicca-Marques and Heymann 2013). These small-bodied monkeys use all levels of the forest, but are often found in the lower strata (up to 10 m) (Bicca-Marques and Heymann 2013). No comprehensive analysis of terrestrial behavior in titi monkeys has been conducted to date; although members of the group have long been reported to use the ground (Kinzey 1977; Mason 1966). In this study, we collated published and unpublished records of terrestrial behavior on the Callicebinae, aiming to identify important ecological correlates, general patterns, and similarities and differences among taxa and regions. We hypothesized that: 1) opportunities to observe unusual behaviors increase with study effort, and we thereby tested the prediction that the number of observations of terrestrial behavior in titi monkeys would be positively correlated with study duration (a proxy for sampling effort); 2) the frequency of terrestrial behavior in titi monkeys increases when arboreal food resources are scarce, and we thereby tested the prediction that the frequency of ground use would be negatively correlated with rainfall level (a proxy for habitat-wide fruit availability); and 3) opportunities for ground use by titi monkeys increase with a reduction in vertical niche dimension, and we thereby tested the prediction that terrestrial behavior would be negatively correlated with forest height (a proxy for vertical niche dimension).
Methods
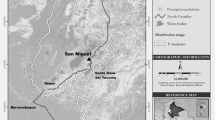
We collated published and unpublished data on terrestrial activity by wild titi monkeys from 86 studies conducted at 65 locations in South America (Electronic Supplementary Material [ESM] Table SI; Fig. 1). Our dataset contains considerable methodological variation in data recording and reporting, a common limitation of collaborative studies using collated, multiauthor data (e.g., Barnett et al.2012a; Boyle et al.2015).

Location of (a) the 19 study sites where Callicebus spp. and (b) the 46 study sites where Cheracebus spp. and Plecturocebus spp. were observed engaging in terrestrial behavior (listed in ESM Table SI). White circles correspond to sites where we recorded up to 10 terrestrial records, white triangles from 11 to 50 records, and white squares are ≥51 records. Black symbols represent nonsystematic records where terrestrially was recorded, but titi monkeys were not the study targets.
We divided terrestrial activity into seven categories (Table II). We did not include accidental falling to the ground as this is not an intentional act. However, we considered intentional plummeting to the ground a predator-avoidance strategy.
Data Analysis
We treated each study site as a sampling unit, irrespective of study duration and the number of individuals involved, allocating each one to a behavioral category. Following Mourthé and collaborators (Mourthé et al. 2007), we defined a terrestrial event as that in which one (or more) individual was observed to descend to the ground (or was sighted when already on the ground). For each species, we calculated the number of records of each type of terrestrial activity as a percentage of total terrestrial records (Table III). Although the frequency of terrestrial behaviors would be a better response variable, we did not have accurate data on sampling effort (i.e., hours of observation) for each study to allow calculation of such rates. Instead, we used study duration (in months) as a proxy for sampling effort.
We used rainfall in the driest quarter of the year (available at WorldClim: Hijmans et al.2005) as a proxy for fruit availability (following Hawes and Peres 2016; Mendoza et al.2017) because for most study sites we lacked comparable data on floristic composition and plant phenology needed to assess the influence of a lean season on terrestrial behavior. We considered driest quarter rainfall a reasonable proxy for seasonality and availability of fruits for most titi monkey species in our study. We obtained information on mean forest height for each study site from the Woods Hole Research Center (http://whrc.org/publications-data/datasets/detailed-vegetation-height-estimates-across-the-tropics/) to assess the influence of forest height on terrestrial behavior. We extracted estimates of forest height across the tropics using the geographic coordinates of each study site plotted on a 30 × 30 m grid.
We computed a chi-squared test to compare the frequency of different activities performed on the ground between Plecturocebus and Callicebus. Subsequently, we applied post hoc chi-squared tests to detect which activities differed between genera by comparing the frequency of each activity against the frequency of all the other activities. Because we used the same variables in multiple tests, we applied a sequential Bonferroni correction to assess statistical significance and reduce the chance of type I errors (Holm 1979). We did not compare Cheracebus with the other genera because of the small number of terrestrial records obtained for members of this genus.
We used a generalized linear model (GLM) with a Poisson response distribution to examine whether study duration, rainfall level, and forest height were correlated with the total number of terrestrial records reported in each study. Then, to plot the effect of one variable while controlling for the other, we ran partial models (Velleman and Welsch 1981). We computed partial regressions in three steps: first, we computed GLMs of our response variable (terrestrial activity) against two of our predictors, excluding a given predictor that was explored separately (e.g., terrestrial activity vs. study duration + vegetation height, terrestrial activity vs. rainfall level + vegetation height). In the second step, we computed GLMs of the given independent variable that we explored separately against the other two independent variables. Finally, we plotted the residuals from the first step against the residuals from the second step. We checked visually for compliance with model assumptions through diagnostic plots (Zuur et al.2010), and checked residuals for homoscedasticity using a Shapiro–Wilk test (W = 0.932, P = 0.133). We also checked for multicollinearity via a variance inflation factor (keeping all variables with VIF <3.0; Zuur et al.2010), using the package car (Fox and Weisberg 2011). We included in these analyses only the 19 systematic studies that contained sufficient information on all variables aforementioned and performed all analyses using R 3.3.0 (R Core Team 2016).
Data Availability
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
Ethical Note
All contributors declare that the studies adhered to the legal requirements of the countries in which the fieldwork was conducted and complied, in each case, with the appropriate ethical requirements of the institutions and governments concerned and adhered to the Code of Best Practices for Field Primatology of the American Society of Primatologists and International Primatological Society (www.asp.org/resources/docs/Code%20of_Best_Practices%20Oct%202014.pdf). The authors declare that they have no conflict of interest.
Results
Terrestrial behaviors (N = 764 records) were recorded in 72 of the 88 study populations (71%); this includes all five species of Callicebus (N = 20 studies, N = 333 records), 48 of 57 populations (84%) of 18 species of Plecturocebus spp. (N = 57 studies, N = 425 records), and 5 of 6 (83%) populations of four species of Cheracebus spp. (N = 6 studies, N = 6 records) (ESM Table SI). Most data (67%) came from nonsystematic studies, whereas the remaining (33%) came from systematic studies of titi monkeys. Terrestrial activity was most commonly associated with feeding/foraging and moving/traveling (Table III). In general, behaviors performed when the titi monkeys were on the ground differed between Callicebus spp. and Plecturocebus spp. (χ2 = 77.823, df = 7, P < 0.0001).
Feeding/Foraging, Moving/Traveling, Resting, Geophagy, and Infant Retrieval
Feeding and foraging represented 37% of all terrestrial activity records (Table III). Callicebus spp. devoted most of their time exploiting leaves on the ground (new leaves = 49%, mature leaves = 16%), while Plecturocebus spp. primarily consumed invertebrates (46%), and Cheracebus spp. ate fallen fruits, leaves, seeds, and invertebrates (N = 1 record each). Callicebus spp. engaged more frequently in feeding/foraging on the ground (48%) than Plecturocebus spp. (28%; χ2 = 29.84, df = 1, P = 0.001).
Plecturocebus spp. showed a higher investment in moving/traveling on the ground than did Callicebus spp. (40% vs. 16%; χ2 = 29.70, df = 1, P = 0.001). This stems from frequent observations of Plecturocebus modestus, and especially Plecturocebus olallae individuals (17.7% of study records) traveling on the ground to reach feeding sites in naturally fragmented forests.
Resting on the forest floor was uncommon in all three genera, and did not differ between Callicebus spp. and Plecturocebus spp. (4% vs. 3%; χ2 = 1.08, df = 1, P = 0.297). Rates of geophagy (χ2 = 3.32, df = 1, P = 0.068) and infant retrieval (χ2 = 0.18, df = 1, P = 0.665) also did not differ between the two genera.
Social Interactions
Several titi monkeys were recorded playing directly on the forest floor and/or on fallen tree trunks. Plecturocebus oenanthe descended to the ground to chase and play with tamarins (Leontocebus leucogenys) in the San Martin region, Peru, while Plecturocebus toppini behaved similarly with tamarins (Leontocebus weddelli and Saguinus imperator) at Los Amigos Biological Station, Peru. Three titi species (Callicebus nigrifrons, Callicebus personatus, and Callicebus coimbrai) were observed playing with marmosets (Callithrix aurita, Callithrix geoffroyi, and Callithrix jacchus, respectively) in southeastern and northeastern Brazil. Playing behavior was observed in nearly identical proportions in Callicebus spp. and Plecturocebus spp. (13% and 11%, respectively; χ2 = 1.70, df = 1, P = 0.191).
Antipredator Behaviors
Predators were reported as common at 16 of 20 sites (80%) of Callicebus spp., 4 of 5 Cheracebus spp. sites (80%), and 33 of 56 Plecturocebus spp. sites (59%) (ESM Table SI). Individuals of all three genera were observed using the forest floor to escape from potential aerial predators (e.g., hawks, eagles) and humans. Frequencies of antipredator behavior on the ground were similar for Callicebus spp. and Plecturocebus spp. (1% for both; χ2 = 0.52, df = 1, P = 0.467).
Correlates of Terrestrial Behavior in Titi Monkeys
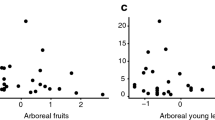
We found that, overall, study duration and rainfall level correlated with terrestrial activity (R2 = 0.29). As predicted, the longer a study, the higher the number of records of titi monkeys on the ground (Fig. 2a). In addition, the lower the rainfall (high seasonality and low fruit availability during lean seasons), the higher the frequency of ground use (Fig. 2b). Forest height, however, did not correlate with ground use (Table IV).

Partial regressions of the number of terrestrial records in titi monkeys against (a) study duration (controlled for rainfall level and vegetation height) and (b) rainfall level (controlled for study duration and vegetation height).
Discussion
Our findings demonstrated that, overall, callicebines are more likely to use the ground than are other pitheciine genera (Barnett et al.2012a), although there was considerable variation in the extent and nature of terrestrial behaviors. While more than half of the species of each genus was observed using the ground level, there was substantial between-species, population-specific, and context-dependent variation in the frequencies at which titi monkeys performed activities on the ground. These findings should be interpreted with caution due to the limitations of our collated data set, including variation in sampling efforts and methods. However, we found interesting patterns of ground use in titi monkeys. Such variation could result from several factors, including phylogenetic, seasonal, and biogeographic differences between study sites, local density of predators (Campbell et al.2005), and local traditions (sensu Tabacow et al.2009).
Feeding/foraging was the most frequent activity performed on the ground by Callicebus spp. and Cheracebus spp. and the second most frequent activity by Plecturocebus spp. Fleshy fruit is the main food type in the diet of titi monkeys, whereas new and mature leaves and invertebrates are typically exploited during lean periods, and thus are characterized as alternative foods (Bicca-Marques and Heymann 2013). Leaves and invertebrates are protein-rich foods, with high concentrations of lipids in the latter (Lambert 2011). Therefore, we suggest that descending to the ground to search for these resources during lean periods is likely to be linked to the need to obtain nutrients. The high number of records of feeding on the ground by titi monkeys suggests that the benefits of descending from the canopy to access food resources outweighs the potential costs of increased predation risk and handling difficulties (Treves 2000).
The geophagy recorded for Plecturocebus spp. and Callicebus spp. can be related to one of several benefits (or a combination of them) resulted from ingestion of soil, including mineral supplementation, antacid action, toxin absorption, endoparasite control, and/or antidiarrheal agents (Krishnamani and Mahaney 2000; Setz et al.1999). Forest ground levels have a higher concentration than the canopy of old and dead bark under which insects can hide (Li 2007), and may be locally rich in clay and mud areas (e.g. mineral licks) (Blake et al.2010; Lee et al.2010; Voigt et al.2008). Geophagy is also relatively common among other Amazonian pitheciids, such as Chiropotes spp. and Pithecia spp. (Ferrari et al.2008), possibly due to high levels of dietary plant-based tannins, which is linked to the generally poor soils of the region. Other animals, such as frugivorous bats, nonpitheciid primates, and parrots also eat clay to alleviate the effects of plant alkaloids or as an alternative source of nutrients (Blake et al.2010; Bravo et al.2008; Brightsmith et al.2008).
Moving/traveling on the ground is a major component of terrestriality in primates. Primates may travel through open areas to disperse between habitat patches or feeding sites (Li 2007). Such behaviors, however, are often accompanied by the enhanced risk of predation (Barnett et al.2015; Galetti and Sazima, 2006) and exposure to parasites (Nunn and Altizer 2006). Moreover, vegetation type, diet, and distribution of food resources influence arboreal primate decisions to use the ground, for example, Bolivian endemic titi monkeys occur in naturally fragmented forests, particularly Plecturocebus olallae and regularly travel on the ground to move between forest patches (Kirkpatrick and Long 1994; Li 2007; Martínez and Wallace, 2011; Su et al.1998). When necessary, titis travel to find new suitable habitat or to reunite with a groups (Ferrari et al.2013a,b). However, the nature of the causative factors behind the higher frequency of moving/traveling in Plecturocebus spp. compared to Callicebus spp. remain unclear. Overall, more detailed analyses are required to assess which habitat attributes and matrix elements affect the likelihood of ground use by titi monkeys for moving/traveling activities.
The rarity of resting on the ground by titi monkeys may also result from increased predation risk (Eppley et al.2016; Mourthé et al.2007) and/or exposure to parasites (Nunn and Altizer 2006), although resting on the ground can also provide thermoregulatory benefits, as suggested for bamboo lemurs (Hapalemur meridionalis: Eppley et al.2016) and chimpanzees (Pan troglodytes: Takemoto 2004). At all study sites, terrestrial predators were potentially present (ESM Table SI). For example, felids and tayra can be found at Manu National Park, Peru (Endo et al.2010); cougars, boa snakes, and pit-vipers at Fazenda Trapsa, northeastern Brazil (Chagas et al.2010); and tayra, ocelots, and cougars at RPPN Santuário do Caraça, southeastern Brazil (Talamoni et al.2014). Furthermore, the forest floor may also harbor a variety of infective parasite stages released in feces, vomit, blood, or urine that increase the risk of infection (Nunn et al.2000).
Similar to other activities performed on the ground, playing exposes titi monkeys to terrestrial predators while hampering their ability to be vigilant. This limitation can be compensated for by an increase in vigilance by those group members that remain in the canopy, as suggested for squirrel monkeys (Saimiri boliviensis: Biben et al. 1989), golden lion tamarins (Leontopithecus rosalia: Oliveira et al. 2003), and black-fronted titi monkeys (Callicebus nigrifrons, C. Gestich, pers. obs.). The interspecific play observed between Plecturocebus spp. and various callitrichids may also benefit interacting individuals in a similar way, in addition to a dilution effect resulting from the increase in the number of potential prey (Delm 1990). Although we do not have data on predator density at each study site, carnivorous mammals and snakes were present at all sites where play was recorded. Despite these risks, play on the ground is often recorded in titi monkeys (Kinzey 1981), other pitheciids (Barnett et al.2012a), and atelids (Campbell et al.2005; Mourthé et al.2007).
Finally, despite the increased risk of predation by terrestrial carnivores on the ground, the forest floor can also serve as an escape route for titis from arboreal and aerial predators, conspecific chasing, and humans (Table III), as reported for other Neotropical primates (Ateles spp.: Julliot 1994; Brachyteles hypoxanthus: Mourthé et al.2007; Cacajao spp.: Chiropotes spp. and Pithecia spp.: Barnett et al.2012a,b; Cebus spp.: Gilbert and Stouffer 1995). This escape strategy can be an extension of plummeting into the lower vegetation, a common response of small and medium-sized primates (ca. 2–8 kg) to the presence of aerial predators (Barnett et al.2017, 2018; Mourthé and Barnett 2014). Under such circumstances, the additional danger of meeting other predators is likely to be temporarily offset in the presence of an imminent threat.
Correlates of Terrestrial Behavior in Titi Monkeys
We found that both study duration and rainfall level (surrogates for sampling effort and resource availability, respectively) correlated with ground use by titi monkeys. Longer study duration increased likelihood of observing rare behaviors (Weatherhead 1986). Nevertheless, 5 out of 19 systematic studies did not report ground use in Callicebinae, suggesting that the extended contact with human observers does not always facilitate observations of terrestriality. Such variation is compatible with the hypothesis that other factors (probably local ones) are more influential than human contact. Changes in resource distribution and availability influence habitat choice (Camaratta et al.2017; Mourthé 2014), resource selection and foraging strategies (Nagy-Reis and Setz 2017), and forest strata use (Ding and Zhao 2004) by primates. Titi monkeys may also adjust diet composition in response to variations in fruit availability and often use the lower forest strata (0.5–10 m) during periods of fruit scarcity (Acero-Murcia et al.2018; Bicca-Marques and Heymann 2013; Caselli and Setz 2011; Souza-Alves et al.2011). While terrestrial behavior in titi monkeys was correlated with fruit scarcity (this study), some primate species do not seem to follow this pattern. Although significant, the estimates of our models were low (Table IV), implying that an increase of 200 mo in study duration or a decrease of 200 mm of rainfall in the driest quarter is required to record one additional observation of terrestrial behavior in titi monkeys. However, such a decrease in rainfall in the driest quarter may not be feasible in regions where it is lower than 200 mm. In addition, the more common presence of open-canopy forests in these regions probably explains why their titis use the ground more frequently (Deguchi et al.2006).
Titi monkeys have a wide distribution in South America, occurring in forests that vary greatly in height and canopy connectivity (ESM Table SI). Independent of height and canopy connectivity, titi monkeys usually explore the lower strata in disturbed forests (Bicca-Marques and Heymann 2013). This proximity to the ground together with food scarcity in the canopy can help to explain their terrestriality. However, alternative factors, such as predation risk, might explain why titi monkeys seem to avoid the ground at some sites. The identification of the drivers of terrestriality in New World monkeys remains a subject for future continued research.
Conclusion
Titi monkeys engage in a variety of activities on the ground, which are more frequent in populations inhabiting more marked seasonal environments and those studied for longer periods. There is a clear difference in the pattern of ground use between Callicebus spp. (Atlantic Forest titi monkeys) and Plecturocebus spp. (Amazonian titi monkeys). Whereas Callicebus spp. showed a higher frequency of feeding/foraging for food resources on ground, Plecturocebus spp. moved/traveled more frequently on the ground, probably to find alternative food sources and to cross forest clearings. Although Callicebus spp. occur in highly fragmented landscapes more frequently than do Plecturocebus spp., Callicebus species appear to move/travel less than Plecturocebus on the ground. For Cheracebus spp., we cannot assess whether infrequently observed ground use reflects lower sampling efforts or other more subtle methodological differences between studies, or instead represents a genuine genus-specific propensity for less-frequent terrestriality. The possible ecological and behavioral specialization of Cheracebus spp. to terra firme forests (van Roosmalen et al.2002), which have high and well-stratified canopies (Defler 1994; Lawler et al.2006), along with the basal position of this clade within callicebines (Byrne et al.2016), suggests that they share a low level of terrestriality, similar with pitheciines. Further investigation is needed to appropriately address differences in moving/traveling behavior between Plecturocebus spp. and Callicebus spp. and the rarity of terrestrial behavior in Cheracebus spp. and to assess whether all titi monkeys share similar levels of behavioral flexibility in disturbed habitats.
References
Acero-Murcia, A., Almario, L. J., García, J., Defler, T. R., & López, R. (2018). Diet of Caquetá titi (Plecturocebus caquetensis) in a disturbed forest fragment in Caquetá, Colombia. Primates Conservation, 32, 1–17.
Almeida-Silva, B., Guedes, P. G., Boubli, J. P., & Strier, K. B. (2005). Deslocamento terrestre e o comportamento de beber em um grupo de barbados (Alouatta guariba clamitans Cabrera, 1940) em Minas Gerais, Brasil. Neotropical Primates, 13, 1–3.
Ancrenaz, M., Sollmann, R., Meijaard, E., Hearn, A. J., Ross, J., et al (2014). Coming down from the trees: Is terrestrial activity in Bornean orangutans natural or disturbance driven? Scientific Reports, 4, 4024.
Anderson, J., Rowcliffe, J. M., & Cowlishaw, G. (2007). Does the matrix matter? A forest primate in a complex agricultural landscape. Biological Conservation, 135, 212–222.
Aversi-Ferreira, R. A. G. M. F., de Abreu, T., Pfrimer, G. A. Silva, S. F., Ziermann, J. M., et al. (2013). Comparative anatomy of the hind limb vessels of the bearded capuchins (Sapajus libidinosus) with apes, baboons, and Cebus capucinus: With comments on the vessels’ role in bipedalism. BioMed Research International, ID 737358. https://doi.org/10.1155/2013/737358.
Aximoff, I., & Vaz, S. M. (2016). Bugio-ruivo (Primates, Atelidae) em campos de altitude e com anomalia na coloração no Parque Nacional do Itatiaia, Sudeste do Brasil. Oecologia Australis, 20, 122–127.
Barnett, A. A., Almeida, T., Spironello, W. R., Sousa Silva, W., MacLarnon, A., & Ross, C. (2012b). Terrestrial foraging by Cacajao melanocephalus ouakary (Primates) in Amazonian Brazil: Is choice of seed patch size and position related to predation-risk? Folia Primatologica, 83, 126–139.
Barnett, A. A., Andrade, E. S., Ferreira, M. C., Garcia Soares, J. B., Fonseca da Silva, V., et al (2015). Primate predation by black hawk-eagle (Spizaetus tyrannus) in Brazilian Amazonia. Journal of Raptor Research, 49, 105–107.
Barnett, A. A., Bezerra, B. M., Oliveira, M., Queiroz, H., & Defler, T. R. (2013). Cacajao ouakary in Brazil and Colombia: Patterns, puzzles and predictions. In L. M. Veiga, A. A. Barnett, S. F. Ferrari, & M. A. Norconk (Eds.), Evolutionary biology and conservation of titis, sakis and uacaris (pp. 179–195). Cambridge: Cambridge University Press.
Barnett, A. A., Boyle, S., Norconk, M., Palminteri, S., Santos, R. S., et al (2012a). Terrestrial activity in Pitheciins (Cacajao, Chiropotes and Pithecia spp.). American Journal of Primatology, 74, 1106–1127.
Barnett, A. A., de Oliveira, T., Soares da Silva, R. F., de Albuquerque Teixeira, S., Todd, L. M., et al (2018). Honest error, precaution or alertness advertisement? Reactions to vertebrate pseudopredators by red-nosed cuxiús (Chiropotes albinasus), a high-canopy Neotropical primate. Ethology, 124, 177–187.
Barnett, A. A., Silla, J. M., de Oliveira, T., Boyle, S. A., Bezerra, B. M., Spironello, W. R., Setz, E. Z. F., da Silva, R. F. S., de Albuquerque Teixeira, S., Todd, L. M., & Pinto, L. P. (2017). Run, hide or fight: Anti-predation strategies in endangered red-nosed cúxiu (Chiropotes albinasus, Pitheciidae) in South-Eastern Amazonia. Primates, 58, 353–360.
Biben, M., Symmes, D., & Bernhards, D. (1989). Vigilance during play in squirrel monkeys. American Journal of Primatology, 17, 41–49.
Bicca-Marques, J. C. (1992). Drinking behavior in the black howler monkey (Alouatta caraya). Folia Primatologica, 58, 107–111.
Bicca-Marques, J. C., & Heymann, E. (2013). Ecology and behavior of titi monkeys (genus Callicebus). In L. M. Veiga, A. A. Barnett, S. F. Ferrari, & M. A. Norconk (Eds.), Evolutionary biology and conservation of titis, sakis and uacaris (pp. 196–207). Cambridge: Cambridge University Press.
Bicca-Marques, J. C., Muhle, C. B., Prates, H. M., Oliveira, S. G., & Calegaro-Marques, C. (2009). Habitat impoverishment and egg predation by Alouatta caraya. International Journal of Primatology, 30, 743–748.
Blake, J. G., Guerra, J., Mosquera, D., Torres, R., Loiselle, B. A., & Romo, D. (2010). Use of mineral licks by white-bellied spider monkeys (Ateles belzebuth) and red howler monkeys (Alouatta seniculus) in eastern Ecuador. International Journal of Primatology, 31, 471–483.
Boubli, J. P., Byrne, H., da Silva, M. N. F., Silva-Júnior, J., Costa-Araújo, R., et al (2019). On a new species of titi monkey (Primates: Plecturocebus Byrne et al., 2016), from Alta Floresta, southern Amazon. Brazil. Molecular Phylogenetics and Evolution, 132, 117–137.
Bóveda-Penalba, A., Vermeer, J., Rodrigo, F., & Guerra-Vásquez, F. (2009). Preliminary report on the distribution of Callicebus oenanthe on the eastern feet of the Andes. International Journal of Primatology, 30, 467–480.
Boyer, D., Ramos-Fernández, G., Miramontes, O., Mateos, J. L., Cocho, G., Larralde, H., Ramos, H., & Rojas, F. (2006). Scale-free foraging by primates emerges from their interaction with a complex environment. Proceedings of the Royal Society of London B: Biological Sciences, 273(1595), 1743–1750.
Boyle, S. A., Thompson, C. L., DeLuycker, A., Alvarez, S. J., Alvim, T. H. G., et al (2015). Geographic comparison of plant genera used in frugivory among the pitheciids Cacajao, Callicebus, Chiropotes and Pithecia. American Journal of Primatology, 78, 493–506.
Bravo, A., Harms, K. E., Stevens, R. D., & Emmons, L. H. (2008). Collpas: Activity hotspots for frugivorous bats (Phyllostomidae) in the Peruvian Amazon. Biotropica, 40, 203–210.
Brightsmith, D. J., Taylor, J., & Phillips, T. D. (2008). The roles of soil characteristics and toxin adsorption in avian geophagy. Biotropica, 40, 766–774.
Byrne, H., Rylands, A. B., Carneiro, J. C., Lynch Alfaro, J. W., Bertuol, F., et al (2016). Phylogenetic relationship of the New World titi monkeys (Callicebus): First appraisal of taxonomy on molecular evidence. Frontiers in Zoology, 13, 10. https://doi.org/10.1186/s12983-016-0142-4.
Camaratta, D., Chaves, Ó. M., & Bicca‐Marques, J. C. (2017). Fruit availability drives the distribution of a folivorous–frugivorous primate within a large forest remnant. American Journal of Primatology, 79(3), e22626. https://doi.org/10.1002/ajp.22626
Campbell, C. J., Aureli, F., Chapman, C. A., Ramos-Fernández, G., Matthews, K., Russo, S. E., Suarez, S., & Vick, L. (2005). Terrestrial behavior of Ateles spp. International Journal of Primatology, 26, 1039–1051.
Cant, J. G. (1992). Positional behavior and body size of arboreal primates: A theoretical framework for field studies and an illustration of its application. American Journal of Physical Anthropology, 88, 273–283.
Caselli, C. B., & Setz, E. Z. F. (2011). Feeding ecology and activity pattern of black-fronted titi monkeys (Callicebus nigrifrons) in a semideciduous tropical forest of southern Brazil. Primates, 52, 351–359.
Chagas, R. R. D., & Ferrari, S. F. (2010). Habitat use by Callicebus coimbrai (Primates: Pitheciidae) and sympatric species in the fragmented landscape of the Atlantic Forest of southern Sergipe, Brazil. Zoologia (Curitiba), 27, 853–860.
Chagas, R. R. D., Santos Jr., E. M., Souza-Alves, J. P., & Ferrari, S. F. (2010). Fazenda Trapsa, a refuge of mammalian diversity in Sergipe, northeastern Brazil. Revista Nordestina de Biologia, 19, 35–43.
Cheyne, S. M., Supyansyha, A., Neale, C. J., Thompson, C., et al (2018). Dowon from the treetops: Red langur (Presbytis rubicunda) terrestrial behavior. Primates, 59, 437–448. https://doi.org/10.1007/s10329-018-0676-5.
Defler, T. R. (1994). Callicebus torquatus is not a white-sand specialist. American Journal of Primatology, 33, 149–154.
Deguchi, A., Hattori, S., & Park, H.-T. (2006). The influence of seasonal changes in canopy structure on interception loss: Application of the revised gash model. Journal of Hydrology, 318, 80–102.
Delm, M. M. (1990). Vigilance for predators: Detection and dilution effects. Behavioral Ecology and Sociobiology, 26(5), 337–342.
Dib, L. R. T., Oliva, A. S., & Strier, K. B. (1997). Terrestrial travel in muriquis (Brachyteles arachnoides) across a forest clearing at the Estação Biológica de Caratinga, Minas Gerais, Brazil. Neotropical Primates, 5, 8–9.
Ding, W., & Zhao, Q. K. (2004). Rhinopithecus bieti at Tacheng, Yunnan: Diet and daytime activities. International Journal of Primatology, 25(3), 583–598.
Endo, W., Peres, C. A., Salas, E., Mori, S., Sanchez-Vega, J. L., Shepard, G. H., Pacheco, V., & Yu, D. W. (2010). Game vertebrate densities in hunted and nonhunted forest sites in Manu National Park, Peru. Biotropica, 42(2), 251–261.
Eppley, T. M., Donati, G., & Ganzhorn, J. U. (2016). Determinants of terrestrial feeding in an arboreal primate: The case of the southern bamboo lemur (Hapalemur meridionalis). American Journal of Physical Anthropology, 161(2), 328–342.
Ferrari, S. F., Boyle, S. A., Marsh, L. K., Port-Carvalho, M., Santos, R. R., Silva, S. S. B., Vieira, T. M., & Veiga, L. M. (2013a). The challenge of living in fragments. In L. M. Veiga, A. A. Barnett, S. F. Ferrari, & M. A. Norconk (Eds.), Evolutionary biology and conservation of titis, sakis and uacaris (pp. 350–358). Cambridge: Cambridge University Press.
Ferrari, S. F., & Hilário, R. R. (2012). Use of water sources by buffy-headed marmosets (Callithrix flaviceps) at two sites in the Brazilian Atlantic Forest. Primates, 53, 65–70.
Ferrari, S. F., Santos, E. M., Freitas, E. B., Fontes, I. P., Souza-Alves, J. P., et al. (2013b). Living on the edge: Habitat fragmentation at the interface of the semiarid zone in the Brazilian northeast. In L. Marsh & C. A. Chapman (Eds.), Primates in fragments: Complexity and resilience (pp. 121–135). Developments in primatology: Progress and prospects. New York: Springer science+business media.
Ferrari, S. F., Veiga, L. M., & Urbani, B. (2008). Geophagy in New World monkeys (Platyrrhini): Ecological and geographic patterns. Folia Primatologica, 79, 402–415.
Fleagle, J. G. (1999). Primate adaptation and evolution (3rd ed.). San Diego: Academic Press.
Fox, J., & Weisberg, S. (2011). Packcage “car” for R software.
Galetti, M., & Sazima, I. (2006). Impact of feral dogs in an urban Atlantic Forest fragment in southeastern Brazil. Natureza and Conservacação, 4, 146–151.
Gilbert, K. A., & Stouffer, P. C. (1995). Variation in substrate use by white-faced capuchins. Human Evolution, 10, 265–269.
Grueter, C. C., Li, D., Ren, B., Wei, F., Xiang, Z., & van Schaik, C. P. (2009). Fallback foods of temperate-living primates: A case study on snub-nosed monkeys. American Journal of Physical Anthropology, 140, 700–715.
Hawes, J. E., & Peres, C. A. (2016). Patterns of plant phenology in Amazonian seasonally flooded and unflooded forest. Biotropica, 48, 465–475.
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G., & Jarvis, A. (2005). Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25, 1965–1978.
Holm, S. (1979). A simple sequentially rejective multiple test procedure. Scandinavian Journal of Statistics, 65–70.
Jones, C. B. (2005). Behavioral flexibility in primates: Causes and consequences. Developments in primatology: Progress and prospects. New York: Springer Science+Business Media.
Julliot, C. (1994). Predation of a young spider monkey (Ateles paniscus) by a crested eagle (Morphnus guianensis). Folia Primatologica, 63, 75–77.
Kinzey, W. G. (1977). Positional behavior and ecology in Callicebus moloch. Yearbook of Physical Anthropology, 20, 248–480.
Kinzey, W. G. (1981). The titi monkey, genus Callicebus. In A. F. Coimbra-Filho & R. A. Mittermeier (Eds.), Ecology and behavior of Neotropical primates (pp. 241–276). Rio de Janeiro: Academia Brasileira de Ciências.
Kinzey, W. G., Rosenberger, A. L., Heisler, P. S., Prowse, D. L., & Trilling, J. S. (1977). A preliminary field investigation of the yellow handed titi monkey, Callicebus torquatus torquatus, in northern Peru. Primates, 18(1), 159–181.
Kirkpatrick, R. C., & Long, Y. C. (1994). Altitudinal ranging and terrestriality in the Yunnan snub-nosed monkey (Rhinopithecus bieti). Folia Primatologica, 63(2), 102–106.
Krishnamani, R., & Mahaney, W. C. (2000). Geophagy among primates: Adaptive significance and ecological consequences. Animal Behaviour, 59, 899–915.
Lambert, J. E. (2011). Primate nutritional ecology: Feeding biology and diet at ecological and evolutionary scales. In C. J. Campbell, A. Fuentes, K. C. MacKinnon, M. Panger, & S. K. Bearder (Eds.), Primates in perspective (2nd ed., pp. 513–522). New York: Oxford University Press.
Lawler, R. R., Ford, S. M., Wright, P. C., & Easley, S. P. (2006). The locomotor behavior of Callicebus brunneus and Callicebus torquatus. Folia Primatologica, 77, 228–239.
Lee, A. T., Kumar, S., Brightsmith, D. J., & Marsden, S. J. (2010). Parrot claylick distribution in South America: Do patterns of “where” help answer the question “why”? Ecography, 33(3), 503–513.
Li, Y. (2007). Terrestriality and tree stratum use in a group of Sichuan snub-nosed monkeys. Primates, 48, 197–207. https://doi.org/10.1007/s10329-006-0035-9.
Link, A., & Di Fiore, A. (2013). Effects of predation risk on the grouping patterns of white-bellied spider monkeys (Ateles belzebuth belzebuth) in Western Amazonia. American Journal of Physical Anthropology, 150, 579–590.
Link, A., Galvis, N., Fleming, E., & Di Fiore, A. (2011). Patterns of mineral lick visitation by spider monkeys and howler monkeys in Amazonia: Are licks perceived as risky areas? American Journal of Primatology, 73, 386–396.
Martínez, J., & Wallace, R. B. (2011). First observations of terrestrial travel for Olalla’s titi monkey (Callicebus olallae). Neotropical Primates, 18, 49–52.
Mason, W. A. (1966). Social organization of the south American monkey, Callicebus moloch: A preliminary report. Tulane Studies Zoology, 13, 23–28.
Mendoza, I., Peres, C. A., & Morellato, L. P. C. (2017). Continental-scale patterns and climatic drivers of fruiting phenology: A quantitative Neotropical review. Global and Planetary Change, 148, 227–241.
Mesa-Sierra, N., & Pérez-Torres, J. (2017). Calidad estructural y funcional de espacios usados por Alouatta seniculus em fragmentos de bosque seco tropical (Córdoba, Colombia). Neotropical Primates, 23, 9–15.
Michalski, F., & Peres, C. A. (2005). Anthropogenic determinants of primate and carnivore local extinctions in a fragmented forest landscape of southern Amazonia. Biological Conservation, 124, 383–396.
Mourthé, I. M. C. (2014). Response of frugivorous primates to changes in fruit supply in a northern Amazonian forest. Brazilian Journal of Biology, 74, 720–727.
Mourthé, I. M. C., & Barnett, A. A. (2014). Crying tapir: The functionality of errors and accuracy in predator recognition in two Neotropical high-canopy primates. Folia Primatologica, 85, 379–398.
Mourthé, I. M. C., Guedes, D., Fidelis, J., Boubli, J. P., Mendes, S. L., & Strier, K. B. (2007). Ground use by northern muriquis (Brachyteles hypoxanthus). American Journal of Primatology, 69, 706–712.
Nadjafzadeh, M. N., & Heymann, E. W. (2008). Prey foraging of red titi monkeys, Callicebus cupreus, in comparison to sympatric tamarins, Saguinus mystax and Saguinus fuscicollis. American Journal of Physical Anthropology, 135, 56–63.
Nagy-Reis, M. B., & Setz, E. Z. F. (2017). Foraging and ranging behavior of black-fronted titi monkeys (Callicebus nigrifrons) and their relation to food availability in a seasonal tropical forest. Primates, 58, 149–158. https://doi.org/10.1007/s10329-016-0556-9.
Napier, J. R., & Napier, P. H. (1967). Handbook of living primates. London: Academic Press.
Norconk, M. A. (2011). Sakis, uakaris, and titi monkeys: Behavioral diversity in a radiation of primate seed predators. In C. J. Campbell, A. Fuentes, K. C. MacKinnon, M. Panger, & S. K. Bearder (Eds.), Primates in perspective (2nd ed., pp. 122–139). New York: Oxford University Press.
Nowak, K., le Roux, A., Richards, S. A., Scheijen, C. P., & Hill, R. A. (2014). Human observers impact habituated samango monkeys’ perceived landscape of fear. Behavioral Ecology, 25, 1199–1204.
Nunn, C. L., & Altizer, S. (2006). Infectious diseases in primates: Behavior, ecology and evolution. Oxford: Oxford University Press.
Nunn, C. L., Gittleman, J. L., & Antonovics, J. (2000). Promiscuity and the primate immune system. Science, 290, 1168–1170.
Oliveira, C. R., Ruiz Miranda, C. R., Kleiman, D. G., & Beck, B. B. (2003). Play behavior in juvenile golden lion tamarins (Callitrichidae: Primates): Organization in relation to costs. Ethology, 109(7), 593–612.
Ottoni, E. B., & Izar, P. (2008). Capuchin monkeys tool use: Overview and implications. Evolutionary Anthropology, 17, 171–178.
Pinheiro, T., Ferrari, S. F., & Lopes, M. A. (2013). Activity budget, diet, and use of space by two groups of squirrel monkeys (Saimiri sciureus) in eastern Amazonia. Primates, 54(3), 301–308.
Porfirio, G., Santos, F. M., Foster, V., Nascimento, L. F., Macedo, G. C., et al. (2017). Terrestriality of wild Sapajus cay (Illiger, 1815) as revealed by camera traps. Folia Primatologica, 88(1), 1-8.
Pozo-Montuy, G., & Serio-Silva, J. C. (2007). Movement and resource use by a group of Alouatta pigra in a forest fragment in Balancán, México. Primates, 48(2), 102–107. https://doi.org/10.1007/s10329-006-0026-x
Prates, H. M., & Bicca-Marques, J. C. (2008). Age-sex analysis of activity budget, diet, and positional behavior in Alouatta caraya in an orchard forest. International Journal of Primatology, 29, 703–715.
Printes, R. C., Rylands, A. B., & Bicca-Marques, J. C. (2011). Distribution and status of critically endangered blond titi monkey Callicebus barbarabrownae of north-East Brazil. Oryx, 45, 439–443.
R Core Team (2016). R: A language and environment for statistical computing. Version 3.3.0. Vienna, Austria: R Foundation for Statistical Computing.
Rumiz, D. I. (2012). Distribution, habitat and status of the white-coated titi monkey (Callicebus pallescens) in the Chaco-Chiquitano forest of Santa Cruz, Bolivia. Neotropical Primates, 19, 8–15.
Serrano-Villavicencio, J. E., Vendramel, R. L., & Garbino, G. S. T. (2017). Species, subspecies, or color morphs? Reconsidering the taxonomy of Callicebus Thomas, 1903 in the Purus–Madeira interfluvium. Primates, 58, 159–167.
Setz, E. Z. F., Enzweiler, J., Solferini, V. N., Amendola, M. P., & Berton, R. S. (1999). Geophagy in golden-faced saki monkey, Pithecia pithecia chrysocephala, in Central Amazon. Journal of Zoology (London), 247, 91–103.
Shanee, S., & Shanee, N. (2011). Observations of terrestrial behavior in the Peruvian night monkey (Aotus miconax) in an anthropogenic landscape, La Esperanza, Peru. Neotropical Primates, 18, 55–58.
Shanee, S., Tello-Alverado, J. C., Vermeer, J., & Boveda-Penalba, A. J. (2011). GIS risk assessment and GAP analysis for the Andean titi monkey (Callicebus oenanthe). Primate Conservation, 26, 17–24.
Soini, P. (1987). Ecology of the saddle-back tamarin Saguinus fuscicollis illigeri on the Rio Pacaya, northeastern Peru. Folia Primatologica, 49, 11–32.
Souza, F. S. C., & Calouro, A. M. (2018). Predation of army ants by Toppin’s titi monkey, Plecturocebus toppini Thomas 1914 (Primates: Pitheciidae), in an urban forest fragment in eastern Acre. Brazil. Primates, 1–6. https://doi.org/10.1007/s10329-018-0677-4
Souza-Alves, J. P., Fontes, I. P., Chagas, R. R. D., & Ferrari, S. F. (2011). Seasonal versatility in the feeding ecology of a group of titis (Callicebus coimbrai) in the northern Brazilian Atlantic Forest. American Journal of Primatology, 73, 1199–1209.
Su, Y., Ren, R., Yan, K., Li, J., Zhou, Y., Zhu, Z., Hu, Z., & Hu, Y. (1998). Preliminary survey of the home range and ranging behavior of golden monkeys (Rhinopithecus [Rhinopithecus] roxellana) in Shennongjia National Natural Reserve, Hubei, China. In N. G. Jablonski (Ed.), The natural history of the doucs and snub-nosed monkeys (pp. 255–268). London: World Scientific Publishing.
Tabacow, F. P., Mendes, S. L., & Strier, K. B. (2009). Spread of a terrestrial tradition in an arboreal primate. American Anthropologist, 111, 238–249.
Takemoto, H. (2004). Seasonal change in terrestriality of chimpanzees in relation to microclimate in the tropical forest. American Journal of Physical Anthropology, 124, 81–92.
Talamoni, S. A., Amaro, B. D., Cordeiro-Júnior, D. A., & Maciel, C. E. M. A. (2014). Mammals of Reserva particular do Patrimônio natural Santuário do Caraça, state of Minas Gerais, Brazil. Check List, 10(5), 1005–1013.
Treves, A. (2000). Theory and method in studies of vigilance and aggregation. Animal Behaviour, 60(6), 711–722.
van Roosmalen, M. G. M., van Roosmalen, T., & Mittermeier, R. A. M. (2002). A taxonomic review of the titi monkeys, genus Callicebus Thomas, 1903, with the description of two new species, Callicebus bernhardi and Callicebus stephennashi, from Brazilian Amazonia. Neotropical Primates, 10 (Suppl.), 1–52.
Velleman, P. F., & Welsch, R. E. (1981). Efficient computing of regression diagnostics. The American Statistician, 35(4), 234–242.
Voigt, C. C., Capps, K. A., Dechmann, D. K. N., Michener, R. H., & Kunz, T. H. (2008). Nutrition or detoxification: Why bats visit mineral licks of the Amazonian rainforest. PLoS One, 3(4), e2011. https://doi.org/10.1371/journal.pone.0002011.
Weatherhead, P. J. (1986). How unusual are unusual events? The American Naturalist, 128, 150–154.
Wu, B. Q. (1993). Patterns of spatial dispersion, locomotion, and foraging behavior in three groups of Yunnan snub-nosed landue (Rhinopithecus roxellana) on Baimaxue Mountains, northwestern Yunnan Province, China. Folia Primatologica, 60, 63–71.
Wu, B. Q., Tai, Z., & Ji, W. (1988). A preliminary survey of ecology and behavior on Yunnan snub-nosed monkey (Rhinopithecus bieti) group. Zoological Research, 9, 373–384.
Zuur, A. F., Ieno, E. N., & Elphick, C. S. (2010). A protocol for data exploration to avoid common statistical problems. Methods in Ecology and Evolution, 1, 3–14.
Acknowledgments
All authors thank the numerous sources that funded, helped, and supported the various research projects from which the data were compiled and analyzed. We are grateful the Joanna Setchell and two anonymous reviewers for the valuable consideration in the manuscript.
Author information
Authors and Affiliations
Contributions
JPS-A and AAB conceived and designed the experiments. RRH and IM analyzed the data. JPS-A and AAB wrote the first draft of manuscript. All the authors revised the manuscript.
Corresponding author
Additional information
Handling Editor: Joanna M. Setchell.
Electronic supplementary material
ESM 1
(DOCX 73 kb)
Rights and permissions
About this article

Cite this article
Souza-Alves, J.P., Mourthe, I., Hilário, R.R. et al. Terrestrial Behavior in Titi Monkeys (Callicebus, Cheracebus, and Plecturocebus): Potential Correlates, Patterns, and Differences between Genera. Int J Primatol 40, 553–572 (2019). https://doi.org/10.1007/s10764-019-00105-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10764-019-00105-x