
Abstract
Change or continuity of the human diet after the Spanish settlement in America is a topic mostly addressed in historical written documents with little use of the archaeological record and bioarchaeological or culture material. To counteract this weakness, this paper presents a study of the diet in individuals living in central-western Argentina between the seventeenth, eighteenth, and nineteenth centuries. The paper, focusing on historical bioarchaeology using stable isotopes (δ13C, δ15N, δ18O) from bone samples of human skeletal remains found in Mendoza, Argentina. The aim is to reconstruct the human diet and its residential mobility. Our results show little inclusion of maize in these populations’ diets, significantly less than those for the same region during pre-Hispanic times. The data do not indicate a historic continuity in dietary practices between pre-Hispanic and post Hispanic human population.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cultural change, collapse, and the establishment of new organization systems have been distinctive processes originated by the Spanish Conquest of the Americas. Most of the knowledge about this time period is based on written documents. Today, many archaeological studies complement this information from different regions of Latin America. The central- western region of Argentina (Fig. 1) is an interesting case study because it provides a unique example on the establishment of colonial cities in a region where small-scale groups of farmers and hunter-gatherers coexisted (Bárcena 2001; Gambier 2000). Within this framework, several research programs in central-western Argentina have attempted to develop a better understanding of how the Spanish settlement affected the previous local population. Recently, these studies have debated the ethnic structure, diet, and subsistence patterns during the historic time (after 1561 CE) (Bárcena 2001; Chiavazza 1999, 2001, 2010, 2013; Chiavazza and Mafferra 2007; Durán and García 1989; García 1999, 2011; García Llorca 2004; Lagiglia 1976; García Llorca and Cahiza 2007; Michieli 1998; Parisii 1991–92). Central points in this discussion have been the role of agriculture as an economic practice, and the relevance of maize for these populations’ diets. It has been generally accepted that once agriculture and the consumption of domestic resources were established, approximately 2000 years BP, farming practices were continuous until the historic period. This view implies a long-term farming base for the region (García 2011). An alternative model has been proposed (Chiavazza and Mafferra 2007; Gil et al. 2009; Mafferra 2010) which argues that once maize was incorporated into agricultural production it did not become important until 1000 years ago. In addition, the importance of maize varied highly within contemporary populations and among different areas from northern Mendoza (Chiavazza 2013; Gil et al. 2009, 2010, 2011). Chiavazza and Mafferra (2007) propose a similar model using historical data from the fifteenth century for the area of Mendoza city. Chiavazza and Mafferra (2007; also see Mafferra 2010) propose a low significance for American domestic plants during Hispanic times, supported by the lack of American domestic plants in the archaeobotanical record. Therefore, the native domestic plants would not have been significant after the Spanish arrival. Chiavazza and Mafferra’s (2007) paper was strongly questioned and discussed by García (2011; see answers in Chiavazza and Mafferra 2011). García (2011) affirms that the evidence does not permit any doubt about the engagement of agricultural practices by the historic-period Huarpes. García (2011) confirms the traditional model arguing about the existence of a local economy based on extractive and productive activities that persisted through the last 2000 years.

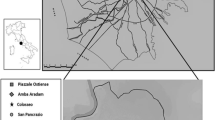
Location of archaeological sites mentioned in the study; San Francisco, (RSF), Alberdi and Ituzaingo (AEI), Plaza Huarpe Building (EPH), Santo Domingo (SD), San Agustín (SA) and La Caridad (LC)
The objective of this article is to improve the characterization of human diets from central-western Argentina, mainly from northern Mendoza (see Fig. 1). Focus is placed on the importance of maize as a dietary resource during historic times and on mobility patterns. In order to discuss the models explained above, we present the results of the first stable isotope (δ13C, δ15N and δ18O) studies on human bone from historical times for the region.
It has been clearly proven that the archaeological record of macrobotanical remains does not directly reflect subsistence and that an archaeological record has the potential to indicate processes on different spatial and temporal scales (Schiffer 1987; Ugan and Coltrain 2012). Records from archaeological deposits usually show a history of formation and temporal and spatial resolutions different to that indicated by chemical bio-indicators on human remains. Within this framework, the potential of stable isotopes to reconstruct human diets is a useful tool for understanding trends on a more fine-grained, individual scale.
Human Samples and Their Contexts
This study uses human bones remain recovered in colonial contexts from Mendoza city. The sample consists of human bones from excavations of Catholic churches: ruins of San Francisco (RSF), Santo Domingo (SD), La Caridad (LC) and San Agustín (SA) (see Fig. 1). The sample includes adults, both male and female, dated in a temporal interval of ca. 250 years, from the beginning of the seventeenth to the middle of the nineteenth centuries. Since the colonial churches were destroyed—as well as the rest of Mendoza city—by an earthquake on March 20, 1861, the integrity of the record and the temporal resolution is low. Chronological discriminations are difficult to make because of the great amount of debris generated in the successive demolitions. In addition, churches were confined places that received citizens’ remains continuously for about 250 years. The space crisis is visible in the movements of bones and the relocation of deposits to generate space for future inhumations. In this context there is yet another distortive process for most of the cases considered: the churches were rebuilt from humble buildings made of earth (adobe and/or tapia walls) during the seventeenth century to brick structures in the eighteenth century (Chiavazza 2008). Furthermore, churches became part of the settlements at a later stage. Early colonial and even pre-Hispanic domestic contexts can usually be found in different stratigraphic levels below the church floors or in contact with graves.
The sample from the Jesuit church (1608–1767), which then became San Francisco (1789–1861), has three building stages: a first temple (ca. 1611–1716) placed where the transept of the second building was raised (1716/1731–1861). In this setting, corpses from victims of the earthquake were documented as being buried around the ruins (Chiavazza 2005). The cases from Santo Domingo and San Agustín correspond to individuals buried under the main naves of the churches built during the eighteenth century. The sample of the individual from La Caridad is difficult to ascribe to a burial from the Jesuit or the Franciscan Order, because the Franciscans built the church and then gave it to the other brotherhood of La Caridad when the Jesuits were expelled (Mansegosa and Chiavazza 2010). This subject is relevant because not only the position establishes the social origin of the dead person on religious grounds, but also with the order entrusted with the funeral rites and disposition of the body (see the documentary study for Santo Domingo in Bárcena (2009). The temporal differences of the studied individuals were defined by their contextual associations and by the materials within stratigraphic levels (Chiavazza and Cortegoso 2001; Chiavazza and Zorrilla 2005).
What Does the Recent Archaeological Record Tell Us About Human Diet?
The study of archaeological middens at these sites (Chiavazza 2010, 2013; Chiavazza and Mafferra 2007) shows a pattern in which the colonial occupations in contact with native populations consumed a wide variety of both animal and plant products (Tables 1 and 2). A rapid incorporation of European subsistence elements, mostly Euro-Asiatic domestic vegetables, can be seen. From the fifteenth- and sixteenth-century contexts, both wild-native and domestic-foreign resources stand out (Chiavazza 2013). Urban sites (AEI, EPH, and RSF, see Fig. 1, Tables 1 and 2) present a record dominated by non-American domesticates. Other contemporary archeological sites, further away from this urban sector (PA70, see Fig. 1, Tables 1 and 2), present a minor presence of these non-local resources. In the non-urban archaeological site (PA70), there is a high importance of fish and low significance of camelids. In addition, the record of native domestic plants (such as maize) is not prominent.
Stable Isotopes, Diet and Mobility
Stable isotope analysis has become a solid line of evidence that has been applied to a variety of subjects such as palaeodiet, palaeoecology, and human mobility, among others (Koch 1998; Panarello et al. 2010). The use of stable isotopes to study palaeodiet began in the 1960s (Bender 1968). Van der Merwe and Vogel (1978) was the first high-impact example of an attempt to use this methodology to discuss the importance of maize in the human diet. Today, stable isotope analysis is a well-established practice in archaeological research (Ambrose 1993; Schwarcz and Schoeninger 1991). The method is based on the property of differential isotope discrimination. For instance, in the case of carbon, this discrimination (13C/12C) enables to differentiate vegetables with different photosynthetic mechanisms (C3, C4 and CAM). Since maize is a C4 plant, it can be easily discriminated in ecological contexts where resources have other photosynthetic pathways. Isotopes from other elements such as N (15N/14N) and O (18O/16O) allow researchers to discuss topics as trophic chains and residential mobility (Panarello et al. 2010).
It is difficult to measure the absolute abundance of a determined isotopic species. Analysts therefore have chosen to use the ratio between the number of molecules containing the heaviest isotope and the number of molecules containing the lightest isotope. In the case of carbon, with stable isotopes 12C and 13C, this ratio called R13 is equal to 13C/12C. Because the values of this ratio are very small, they are expressed in a simpler way based on isotopic deviation (δ). The δ indicates how much the isotope ratio of the sample under study departs from that presented by an already defined international standard (Newton 2010; Panarello et al. 2010)
where RM is the isotope ratio from the sample and RR is the same ratio but from the international standard. This same equation is applied to δ15N for the 15N/14N ratio and to δ18O for the 18O/16O ratio. The international standards to which these ratios refer are V-PDB for carbon, AIR for nitrogen, and V-SMOW for oxygen (Panarello et al. 2010).
The distribution of C isotope values in C3 plants has a mode of −27 ‰ and a range between −34 and −22 ‰ (Heaton 1999). The distribution of values of carbon in plants with a C4 photosynthetic pathway ranges from −16 to −7 ‰ with a mode value of −13 ‰ (Tessone 2011; Pate 1994). This is why it is accepted for central-western Argentina that bone collagen from an individual whose diet was based on C3 resources will have δ13C values of around −21.5 ‰ V-PDB, with a range from −17 to −23 ‰ V-PDB (Gil 2003; Gil et al. 2010). Those whose diet was based on C4 resources will have a bone collagen value of about −7.5 ‰ V-PDB and a range from −14 to −7 ‰ V-PDB, with values for mixed human diets falling in between. These values of δ13C are obtained in relation to pre-established international standards (Ambrose 1993; Tykot 2006). According to experimental advances (Ambrose and Norr 1993), the δ13C values based on bone collagen mainly reflect the protein fraction of the diet. On the other hand, the inorganic fraction (hydroxyapatite or just apatite) reflects the fraction of the diet that provides energy and metabolizes. δ13C from hydroxyapatite is in isotopic equilibrium with bicarbonate in blood (and also with dissolved CO2) that indicates the macronutrients that are being metabolized. The values obtained from bones show an isotope fractionation in 13C related to diet, which is named as the difference between substrate and base. In the case of collagen, an enrichment of around 5 ‰ is estimated, although this value may differ according to species and types of diets (Tessone 2011). The fractionation of the inorganic component (hydroxyapatite) is still under discussion but the generally stipulated value of 12 ‰ is adopted, although it may present values as far as 9 ‰ (Ambrose and Norr 1993; Tykot et al. 2009). This figure has been recently questioned and some authors argue that the relationship between diet and mineral vs. collagen is more complicated (see Kellner and Schoeninger 2007 for a detailed discussion). Other consideration places the temporal resolution as related to the analyzed tissue. In the case of bone, it is presumed that the value obtained is an average of the food ingested and assimilated within the last 7–10 years, but recently it has been suggested that the time span can be even more extensive (Hedges et al. 2007).
Nitrogen stable isotope analysis has been generally used to discriminate differences in the trophic levels between individuals (Hedges and Reynard 2006), but several researchers suggest that the values shown by those isotope values may reflect different ecological, climatic, nutritional, and metabolic variables (Ambrose and De Niro 1986; Hedges and Reynard 2006; Petzke et al. 2010; Sealy et al. 1987). On the other hand, the relationship between the increases in δ15N values directly associated to trophic levels is well founded and does not seem to be challenged (Hedges and Reynard 2006).
Another stable isotope recently incorporated into archaeological studies, particularly in Argentina, is δ18O. In archaeology, oxygen isotopes (18O/16O) are used to investigate mobility and patterns of residence (Buzon et al. 2011; Dupras and Schwarcz 2001; Knudson 2009; Sanhueza and Falabella 2010; Sponheimer and Lee-Thorp 1999; Turner et al. 2009). These isotopes are registered in phosphate and carbonate from hydroxyapatite and mainly reflect the isotopic composition of bodies of water chiefly determined by the water ingested during bone and teeth formation (Longinelli 1984; Longinelli and Nuti 1973; Luz et al. 1984; Sponheimer and Lee-Thorp 1999; Turner et al. 2009; White et al. 2004). The water ingested varies in a regular way with respect to latitude, altitude, rain patterns, and other environmental factors (Dansgaard 1964; Gat 1996; White et al. 1998, 2004). Therefore, if variations in 18O from water from a region are found, there is potential to explore human mobility and patterns of residence (Dupras and Schwarcz 2001; Turner et al. 2009; White et al. 2000, 2002, 2004). In order to understand human values, δ13C, δ15N and δ18O are collected from fauna, plants, and water of the same region, generating aspects of/for isotopic ecology (Martínez Del Rio et al. 2009).
Environment, Resources, and Isotopic Ecology
The current city of Mendoza (see Fig. 1) is located on the western border of the oriental plain, close to the piedmont that joins it with the Cordillera de los Andes. Mendoza city lies in the Monte desert. Most of the rain falls in the summer with an estimated mean annual average of 250 mm. The Río Mendoza, which originates in the Cordillera de los Andes during the melting season, is the main water resource. Several springs, brooks, and swamps are associated with it.
In order to adjust and discuss the results from stable isotope analyses, it is necessary to know the isotopic values of potential resources as well as surface water and rainfall. The isotopic values from resources improve the interpretation of isotopes values in human bone samples, allowing more accurate estimates for paleodiets. Values generated from water samples inform the δ18O and enabled us to adjust the results from the same isotope on human samples, both in terms of residential mobility and/or hypothetical geographic origin of the analyzed individuals. This isotopic information on resource and water contributes to the elaboration of a regional isotopic ecology.
Figure 2 shows the regional tendency of the δ13C and δ15N values available for the most commonly exploited resources during the late Holocene in the center west of Argentina (Durán et al. 2014; Gil et al. 2010). Although the variability of resources is lower than the potentially exploited one, significant trends can be observed in Fig. 2. On the one hand, δ13C values for maize show a significant segregation of this species from the other resources. This allows the expectation for populations that based their diet on this resource to be clearly differentiated from those who did not. This species shows a significant enrichment of δ13C and also of δ15N. With regard to this latter isotope, maize resembles the tendency shown by Cucurbita maxima (squash) the other domestic plant included in Fig. 2. Other significant vegetal resource, algarrobo/ mesquite (Prosopis sp.), also differentiated itself by presenting very low values of both isotopes. The fauna presents more uniform values. Its δ13C values point to a mixed composition of C3 and C4 resources, together with a δ15N mean value of around 7 ‰ (see Fig. 2).

Isotopic trends in δ13C and δ15N in the regional resources (Gil et al. 2010)
Many researchers have used stable isotopes of oxygen (16O/18O) to determine mobility and residence patterns of human populations (Buzon et al. 2011; Dupras and Schwarcz 2001; Knudson 2009; Sanhueza and Falabella 2010; Sponheimer and Lee-Thorp 1999; Turner et al. 2009). The isotopes are registered in phosphate or hydroxyapatite from tooth and/or bone show water composition at the time the tooth and/or bone were formed. In human beings, the isotopic composition of water in the body is mainly but not exclusively defined by the water they drank (Longinelli 1984; Longinelli and Nuti 1973; Luz et al. 1984; Sponheimer and Lee-Thorp 1999; Turner et al. 2009; White et al. 2004). Drinking water itself, in turn, varies according to latitude, altitude, rainfall pattern, and other environmental variables (Dansgaard 1964; Gat 1996; White et al. 1998, 2004). Thus, if the region presents a pattern of variation in 18O, the value of this isotope in bone and teeth can indicate where the individual consumed water. This variation can be used to evaluate mobility and residence of both individuals and populations (Dupras and Schwarcz 2001; Turner et al. 2009; White et al. 2000, 2002, 2004).
For δ18O isotopic trends from water, the data available comes from samples from the Río Mendoza (Hoke et al. 2009), groundwater, and rainfalls on its basin (Hoke et al. 2013). Data from nearby areas helps to contextualize this information and define it on a macro-regional scale (Gil et al. 2011; IAEA/WMO 2006; Panarello and Dapeña 1996; Sanhueza and Falabella 2010; Ugan et al. 2012; Vogel et al. 1975). Previous research propose an altitudinal relationship with δ18O from river waters: the Río Mendoza, central to the basin, presents estimated values of −19 ‰ V-SMOW at 4000 masl, diminishing 3 ‰ every 1000 m according to Vogel et al. (1975) or 4.8 ‰ according to Hoke et al. (2009). According to these gradients, in the city of Mendoza, the Río Mendoza is expected to present δ18O values between 8 and 10 ‰ V-SMOW. Besides, data from groundwater indicates depleted values for the plain, showing its association with the Río Mendoza. Vogel et al. (1975) obtained values of −18 ‰ V-SMOW, confirming that the Río Mendoza is the recharging source of the aquifers and is not rainfall. Rainfall samples in this area present values of −4 ‰ V-SMOW (Hoke et al. 2009).
Methodological Aspects
This paper analyzes the δ13C and δ15N stable isotope values of the human bone collagen and δ13C and δ18O values obtained from the carbonate fraction of these same bones. The δ13C and δ18O values of apatite were calculated using the V-PDB standard and δ15N using AIR. Bone collagen and hydroxyapatite were obtained from laboratory procedures carried out at the Museum of Natural History of San Rafael following the protocol described in Coltrain and Leavitt (2002) and presented below. δ13C, δ15N and δ18O measurements were made at the University Of Wyoming’s Stable Isotope Facility.
Specimens were selected after the bioanthropological description (Mansegosa and Chiavazza 2010). From a visual perspective, the bone specimens presented good conditions regarding preservation, without signs of thermal alteration or advanced weathering. Approximately 1 g of bone was obtained from each individual. Samples processed at MSR follow the laboratory procedures as describe below and were measured at the University of Wyoming’s Stable Isotope Facility. Collagen extraction started with 1 g of cortical bone cleaned of surface contaminants. Samples were demineralized whole in 0.6N HCl at 4 °C and progress was recorded daily. After demineralization, the collagen pseudomorph was rinsed to neutrality, treated with 5 % KOH to remove organic contaminants, and soaked 24 h in a 2:1:0.8 mixture of methanol, cloroform, and water to remove residual lipids. The acid and base extracted collagen pseudomorph were again rinsed to neutrality, then lyophilized and weighed to obtain a collagen yield. Approximately 100 mg of lyophilized collagen were gelatinized in 5 ml of acidified water (pH 3) for 24 h at 120 °C. Water-soluble and -insoluble phases were separated by filtration and the water-soluble phase lyophilized. Collagen13C and 15N were determined by flash combustion to produced CO2 and N2 and measured on a Costech 4010 Elemental Analyzer coupled to a Thermo Delta Plus XP IRMS (analytical precision 0.3 ‰). Both stable isotope measurements and sample weight percent carbon and nitrogen were obtained from single sample combustion. Collagen preservation was evaluated by atomic carbon to nitrogen ratios (atomic C:N). The samples all fall within the 2.9–3.6 range, indicative of adequately preserved bone collagen (Ambrose 1990).
To obtain the isotopic values from carbonate hydroxyapatite, 100 mg of bone were processed, ground, and weighed and then were transferred to an Eppendorf tube. This sample was placed in 3 % hydrogen peroxide for 15 min to remove organic components. Then the sample was washed three times to neutrality and dried. It was then washed in acetic acid 0.1 M for 15 min to remove labile carbonates and washed again to neutrality and dried. Finally, the sample of ground bone was sent to the University of Wyoming’s Stable Isotope Facility (Gas Bench online with a Finnigan Delta Plus XP) for analysis. The values of stable isotopes for carbon and oxygen were determined there and reported in values related to the V-PDB standard (analytical precision 0.3 ‰).
Results
Twenty human bones samples from the seventeenth, eighteenth, and nineteenth centuries were processed (Table 3). From those samples, 13 results based on bone collagen (δ13Ccol and δ15N) and 19 on bone hydroxyapatite (δ13Cap and δ18O) were obtained. Table 3 shows the basic variables of the human bones and the results from the isotopic analyses obtained from each sample. Table 4 summarizes the basic statistics. Our set has a mean value of −16.9 ‰ for δ13Ccol. When the estimation is made on the temporal groups already mentioned, these tendencies vary from −17 to −16.4 ‰, showing a slight enrichment of δ13C from bone collagen toward more recent occupations. The difference between the groups is not statistically significant (one way ANOVA F = 0.37, p = 0.698; Table 4). These values point out that the contributed protein comes mainly from a mixture from C3 and C4 resources, with a slight predominance of C3. This same isotope, when measured in hydroxyapatite, presents an average value of −12.2 ‰ and similar values (between −11.9 and −12.5 ‰) when it is estimated on temporal groups, although with a tendency to deplete (see Table 4). In this case, the results indicate that the energy fraction of the diet would come almost exclusively from C3 resources with a low to null supply of C4 resources. Only one individual shows values departing from this trend, (ca. −8 ‰ NC5 from San Francisco Ruins site), which implies an important energetic supply from C4. The range of variation is greater in δ13C values from collagen (2.9 ‰; between −18 and −15.1 ‰) than from hydroxyapatite (1.5‰; between ca. −12 and −13.5 ‰). The values from hydroxyapatite are significantly homogeneous except for the individual NC5, already mentioned (Fig. 3, also see Table 4). When analyzing according to sex, proteins sources (inferred from δ13Ccol) seem to have been isotopically similar between male and female. But females present greater internal variability, whereas for carbohydrates and fat (inferred from δ13Cap) the greater variation is found in males. In average, however, they present similar trends. The correlation between δ13C for bone collagen and apatite (see Fig. 3) shows a weak correlation of −0.20 (Spearman correlation p = 0.54). This indicates a weak and non-significant relation associating the enrichment of one with the depletion of the other. These results indicate that C4 resources, such as maize, were not a central source either of energy or of proteins. Besides, it shows that the isotopic sources of protein have varied more than the sources of carbohydrates and fat (energy). Individuals from the seventeenth century present a greater homogeneity in values both from bone collagen and hydroxyapatite (DS = 0.29 and 0.10 respectively) than those from eighteenth and nineteenth centuries (see Table 4). However, statistically significant variations between these two isotopes were not detected for temporal sets; that is, as regarding isotopes, there would be no significant variation in the average diet during those ca. 300 years.

Relation between the δ13C values of bone collagen and apatite
The δ15N values show an average of 10.7 ‰, indicating a high trophic level in the food chain. There are no statistical significant variations when analyzing this isotope according to temporal sets, although average samples from the seventeenth century are more enriched (12.2 ‰) and this set presents the smallest variability (DS = 0.1). No significant variation was found between the sexes.
The isotopic composition of oxygen in hydroxyapatite from mammalian bones is directly related to corporeal water composition, which reflects the isotopic composition of the oxygen that enters the body. The isotopic composition of oxygen in turn is defined by a complex relationship between climate, diet, and physiology (Longinelli 1984; Luz et al. 1984; Sponheimer and Lee-Thorp 1999). The main sources of bodily oxygen are atmospheric O2, consumed water, and oxygen found in food. From these three sources, the drinking water has the most significant role in determining human corporeal values in δ18O.
The analyzed samples indicate an average of −11.1 ‰ with a range between −8 and −14 ‰, that is, a variation between individuals of 6 ‰. No statistical differences were detected among temporal sets, although samples from the eighteenth century present smaller variability (SD = 0.75) than the other two temporal sets (SD between 1.8 and 2 respectively; see Table 4).
Discussion
The δ13C values from bone collagen indicate a mixed composition of C3 and C4 resources in human diet but a significantly higher composition of C3. Bone collagen fraction mainly indicates the source/s origin of the proteins on the diet (Ambrose and Norr 1993). But hydroxyapatite, the component that indicates the source origin of the energy fraction on the diet, presents even more depleted values than bone collagen. This reflects a diet almost exclusively based on C3 resources as the energy source/s. Only one individual presents values that contradict this trend (NC5). These data support the low importance maize had among human diets during the seventeenth, eighteenth, and nineteenth centuries. However, it does not mean that the people did not know this resource, which is mentioned in documents and is found in archaeological records dating after about 500 years BP.
To improve the meaning of this trend, these isotopic data were compared with those available from other pre-Hispanic human samples from the same region (Gil et al. 2009, 2010, 2011). To do it, the Froehle et al. (2012) model was applied. This is a new version based on Kellner and Schoeninger (2007), where the model defines the diet composition of each sample according to the origin of the energy and proteins based on δ13Cap and δ13Ccol values. The bivariate carbon model (δ13Cap vs. δ13Ccol) provides detailed information on the isotopic signatures of whole diet and dietary protein but is limited in its ability to distinguish between C4 and marine protein. Here, using cluster analysis and discriminant function analysis, Froehle et al. (2012) generate a multivariate diet reconstruction model incorporating δ13Cap, δ13Ccol, and δ15N holistically. Inclusion of the δ15N data prove to be useful in solving protein-related limitations of the bivariate carbon model, and splits the sample into five distinct dietary clusters (Froehle et al. 2012). Five clusters with different diets had been identified statistically by Froehle et al. (2012). Each cluster indicates a diet characterized in terms of C3, C4, and marine/terrestrial resources. We include the human samples presented in this paper as “CWA Historic” and use two group of archaeological human samples (“CWA Group I” and “CWA Group II”) from pre-Hispanic times and recorded in the same region (data from Gil et al. 2009) as comparative strategies. In Fig. 4 “CWA Group I” diet (human samples with chronology between 2000 and 1000 years BP) falls almost entirely into Cluster 4, which barely overlaps Cluster 1. Therefore, “CWA GROUP I” is a result of an overall diet that was about 70 % C3, as well as getting at least 65 % of their protein from C3 sources (definition of each cluster in Froehle et al. 2012). In other words, both their overall diet and protein were overwhelmingly C3, with a smaller contribution from C4 sources. “CWA Historic” (human samples presented in this paper) falls mostly in the Cluster 1 and Cluster 4 overlapping area. Thus, they either ate a 100 % C3 diet, or a 70 % C3 diet similar to that of “CWA GROUP I” (definition of each cluster in Froehle et al. 2012). CWA GROUP II, with chronology between 1000 and 500 years BP, falls mostly in Cluster 2 and seems to have obtained a large portion of their overall diet and their protein from C4 sources (definition of each cluster in Froehle et al. 2012). “CWA Historic” group diet is much more similar to “CWA GROUP I” than to “CWA GROUP II,” and maize was not a significant part of the diet in “CWA Historic,” at most it would have made up 30 % of the diet, but possibly it was not eaten at all.

In the samples analyzed, it has been found that δ18O values have an important range of variation. In order to compare the values obtained from humans bone samples with those from water samples, the formulae presented in Knudson (2009; also see Gil et al. 2014a, b) was used. The values from humans converted to V-SMOW indicate they range from −19 to −11 ‰ V-SMOW, with an average of −15.5 ‰. The 8 ‰ of variation in the δ18O values is a substantial range, suggesting that these individuals consumed different water sources during the last decade of their lives. This indicates differences in residential mobility and/or geographic origin between individuals. On the other hand, these values are isotopically more depleted than those expected for individuals who drank water from the Río Mendoza in the city area (expected value −8 ‰ V-SMOW). However, the values in human bone samples are expected to be in this region at altitudes ranging from 5500 to 2000 − 1500 masl, or alternatively are similar to results obtained from groundwater aquifers near the city of Mendoza (−18 ‰ V-SMOW, Vogel et al. 1975). In this regard, data would point to a significant use of local groundwater or, less likely, the migration of individuals from other sectors such as the highland in the Cordillera de Los Andes. This last possibility is rather unlikely due to the limited temporal availability of this sector, which can only be inhabited during the summer. Figure 5 compares the δ18O obtained for human bone samples on pre-Hispanic and historic times from the lowlands and the highlands in central- western Argentina (ca. 30–32° SL; Gil et al. 2014a). Figure 5 shows a statistically significant difference, the values of the samples studied in the present paper are significantly lower than the others mentioned above (Gil et al. 2014a). The data show a change in the water source of supplies and/or geographic origin of the historical individuals from pre-Hispanic times.

Another significant topic concerns the analysis of individuals as either indigenous or Spanish or “Criollos” (creoles). The correlation between different cultural identities and cultural consumption patterns is difficult to assess from the archaeological record. The topic is relevant but interpreting the information from the written documents is also formidable. Thus, the intention of this paper is not to focus on the ethnic association of diet patterns. We assume that if a person was buried in a temple that he or she adhered, at least formally, to the ideology of Western society as expressed in the seventeenth-nineteenth centuries. A change in their diet and consumption pattern may or may not be inferred. In general, historic funerary documents provide information about cultural identity and geographic origin of individuals (Indian, African, mulatto, creole), but lacks spatial information about the individuals’ burial locations. The historic record itself is fragmentary and generally has low spatial resolution. Based on this observation, we avoid the ethnic assignation of individuals, but assume they ascribed to a Western lifestyle.
Conclusion
The results presented in this paper support the arguments of a human diet with low emphasis on maize during historical times (seventeenth-nineteenth centuries) in central-western Argentina. These trends are basically confirmed in colonial contexts recorded in Mendoza city. Agricultural practices centered on maize became important during part of the late Holocene (Gil et al. 2014b). Counter intuitively, during colonial times maize did not play such a central role to the urban population. The preliminary isotopic analysis of human individuals in rural/non urban contexts close to Mendoza city shows a similar pattern (Gil et al. 2009, 2014a, b). Maize was a highly significance resource during pre-Hispanic times, at least until 1400 CE (Gil et al. 2014b). Maize could be displaced by the early incorporation of animal (cattle, poultry) and plant (wheat and barley) products in the Mendoza valley. The zooarchaeological and archaeobotanical records in Mendoza-colony contexts are consistent with the isotopic data obtained in this study. It is debated today whether this dietary change was the result of Spanish invasion alone and/or a consequence of a new environmental structure associated with climatic change that preceded it, called the Little Ice Age (Gil et al. 2014b).
The incorporation of new, foreign products may not have taken place on a preexistent economic basis from the Huarpe ethnic group as proposed by the written documentation. The residential mobility of populations during colonial times is being debated. The δ18O results are not as would be expected for a population consuming water from the Río Mendoza in the city. This river, running along canals and ditches, has always been thought to be the main source of the city’s water. However, toward the end of the eighteenth century, the installation of a system of aqueducts and fountains provided water from a pre-Andean spring (El Challao) to the inhabitants of the city. In the historic reconstruction, the records from sources of drinking water have not been sufficiently analyzed. Yet, consuming water from the Río Mendoza has been taken for granted. The lack of evidence of water storage in wells or by other means provides analytical lines of research in historical documents for more concrete information about the origin of water consumed by the inhabitants of the City of Mendoza between the seventeenth and nineteenth centuries.
References
Ambrose, S. H. (1990). Preparation and characterization of bone and tooth collagen for isotopic analysis. Journal of Archaeological Science 17: 431–451.
Ambrose, S. (1993). Isotopic analysis of paleodiets: methodological and interpretive considerations. In Sandford, M. (ed.), Investigations of Ancient Human Tissue, Gordon and Breach Science, New York, pp. 59–129.
Ambrose, S., and De Niro, M. (1986). Reconstruction of African human diet using bone collagen carbon and nitrogen isotope ratios. Nature 319: 321–324.
Ambrose, S., and Norr, L. (1993). Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. In Lambert, J., and Grupe, G. (eds.), Prehistoric Human Bone: Archaeology at the Molecular Level, Springer, New York, pp. 1–37.
Bárcena, R. (2001). Prehistoria del Centro Oeste Argentino. In Berberián, E., and Nielsen, A. (eds.), Historia Argentina Prehispánica, vol. 2, Ediciones Brujas, Córdoba, pp. 561–634.
Bárcena, R. (2009). La Iglesia y Convento de Santo Domingo Soriano del Área Fundacional de Mendoza: Investigaciones Arqueológicas e Históricas en la Antigua Manzana de los Dominicos. In Targa, J. G. (ed.), Arqueología Colonial Latinoamericana: Modelos de Estudio, Archaeopress, Oxford, British Archaeological Reports, pp. 269–293.
Bender, M. (1968). Mass spectrometric studies of carbon-13 variations in corn and other grasses. Radiocarbon 10: 468–477.
Buzon, M. R., Conlee, C. A., and Bowen, G. J. (2011). Refining oxygen isotope analysis in the Nasca region of Peru: an investigation of water sources and archaeological samples. International Journal of Osteoarchaeology 21: 446–455.
Chiavazza, H. (1999). Por las arenas bailan los remolinos: arqueología en los cauces del río Mendoza. Acta de Resúmenes del XIII° Congreso Nacional de Arqueología Argentina. Universidade Nacional de Córdoba, Córdoba, pp. 320–322.
Chiavazza, H. (2001). Las Antiguas Poblaciones de las Arenas. Arqueología en las Tierras Áridas del Noreste Mendocino, Ediciones Culturales, Mendoza.
Chiavazza, H. (2005). Los templos coloniales como estructuras funerarias. Arqueología en la iglesia jesuita de Mendoza, British Archaeological Reports, Oxford.
Chiavazza, H. (2008). Bases teóricas para el análisis arqueológico de la espacialidad religiosa y los procesos de transformación cultural en la ciudad de Mendoza durante la colonia. Revista de Arqueología Americana 25: 225–244.
Chiavazza, H. (2010). Ocupaciones en antiguos ambientes de humedal de las tierras bajas del norte de Mendoza: sitio Tulumaya (PA70). Intersecciones en Antropología 11: 41–57.
Chiavazza, H. (2013). “No tan simples”: pesca y horticultura entre grupos originarios del norte de Mendoza. Comechingonia Virtual 7: 27–45.
Chiavazza H. and Cortegoso V. (2001). 2001. Arqueología urbana en Mendoza: arqueología en predios eclesiales del área fundacional de Mendoza. In: Arqueología Uruguaya hacia el fin del milenio (Actas del IX Congreso Nacional de Arqueología Uruguaya), Part II: 481–489.
Chiavazza, H., and Mafferra, L. (2007). Estado de las investigaciones arqueobotánicas en Mendoza y sus implicancias en la arqueología histórica. Revista de Arqueología Histórica Argentina y Latinoamericana 1: 127–154. Buenos Aires.
Chiavazza, H., and Mafferra, L. (2011). Comentario. Revista de Arqueología Histórica Argentina y Latinoamericana 5: 164–168.
Chiavazza, H., and Zorrilla, V. (2005). Conclusiones sobre la arqueología del predio mercedario de la ciudad de Mendoza. In Chiavazza, H., and Zorrilla, V. (eds.), Arqueología en el predio mercedario de la ciudad de Mendoza, Facultad de Filosofía y Letras, UNCuyo, Mendoza, pp. 357–364.
Coltrain, J., and Leavitt, S. (2002). Climate and diet in Fremont prehistory: economic variability and abandonment of maize agriculture in the Great Salt Lake basin. American Antiquity 67: 453–485.
Dansgaard, W. (1964). Stable isotopes in precipitation. Tellus 16: 436–468.
Dupras, T., and Schwarcz, H. (2001). Strangers in a strange land: stable isotope evidence for human migration in the Dakhleh Oasis, Egypt. Journal of Archaeological Science 28: 1199–1208.
Durán, V., and García, C. (1989). Ocupaciones Agroalfareras en el Sitio Agua de La Cueva Sector Norte (N.O. de Mendoza). Revista de Estudios Regionales CEIDER 3: 29–64.
Durán, V., Novellino, P., Gil, A., Menéndez, L., Bernal, V., and Pérez, I. (2014). Estudios Arqueológicos y Bioarqueológicos en el Valle del Río Gualcamayo (Norte De San Juan, Argentina). In Cortegoso, V., and Durán, V. (eds.), Arqueología de ambientes de altura de Mendoza y San Juan (Argentina), Universidad Nacional de Cuyo, Mendoza, pp. 163–201.
Froehle, A. W., Kellner, C. M., and Schoeninger, M. J. (2012). Multivariate carbon and nitrogen stable isotope model for the reconstruction of prehistoric human diet. American Journal of Physical Anthropology 147: 352–369.
Gambier, M. (2000). Prehistoria de San Juan, 2nd ed, Ansilta Editora, San Juan.
García, A. (1999). Economía y movilidad de las comunidades huarpes prehispánicas. Revista de Estudios Regionales Centro Interdisciplinario de Estudios Regionales 20: 7–32.
García, A. (2011). Agricultura huarpe y conquista española: discusión de recientes propuestas. Revista de Arqueología Histórica Argentina y Latinoamericana 5: 147–163.
García Llorca, J. (2004). Análisis arqueológico del sitio “Allayme y Gorriti” en el distrito de Pedro Molina, Guaymallén, Mendoza. Terceras Jornadas de Arqueología Histórica y de Contacto del Centro Oeste de la Argentina y Seminario de Etnohistoria. IV Jornadas de Arqueología y Etnohistoria del Centro Oeste del País I: 161–173. UNRC, Río Cuarto.
García Llorca, J., and Cahiza, P. (2007). Aprovechamiento de recursos faunísticos en las Lagunas de Guancache (Mendoza, Argentina). Análisis zooarqueológico de la Empozada y Altos Melién II. Chungará 39(1): 117–133.
Gat, J. R. (1996). Oxygen and hydrogen isotopes in the hydrologic cycle. Annual Review of Earth and Planetary Sciences 24: 225–262.
Gil, A. (2003). Zea mays on the South American periphery: chronology and dietary importance. Current Anthropology 44: 295–300.
Gil, A., Neme, G., Tykot, R. H., Novellino, P., Cortegoso, V., and Durán, V. (2009). Stable isotopes and maize consumption in central western Argentina. International Journal of Osteoarchaeology 19: 215–236.
Gil, A., Neme, G., and Tykot, R. H. (2010). Isótopos estables y consumo de maíz en el centro occidente Argentino: tendencias temporales y espaciales. Chungará 42: 497–513.
Gil, A., Neme, G., and Tykot, R. H. (2011). Stable isotopes and human diet in central western Argentina. Journal of Archaeological Science 38: 1395–1404.
Gil, A., Neme, G., Ugan, A., and Tykot, R. H. (2014a). Oxygen isotopes and human mobility in central-western Argentina. International Journal of Osteoarchaeology 24: 31–41.
Gil, A., Villalba, R., Ugan, A., Cortegoso, V., Neme, G., Michieli, T., Novellino, P., and Durán, V. (2014b). Isotopic evidence on human bone for declining maize consumption during the Little Ice Age in central western Argentina. Journal of Archaeological Science 49:213–227.
Heaton, T. (1999). Spatial, species, and temporal variations in the 13C/12C ratios of C3 plants: implications for palaeodiet studies. Journal of Archaeological Science 26: 637–649.
Hedges, R., and Reynard, L. (2006). Nitrogen isotopes and the trophic level of humans in archaeology. Journal of Archaeological Science 34: 1240–1251.
Hedges, R., Clement, J. G., Thomas, C., O’Connell, and Tamsin, C. (2007). Collagen turnover in the adult femoral mid-shaft: Modeled from anthropogenic radiocarbon tracer measurements. American Journal of Physical Anthropology 133: 808–816.
Hoke, G., Garzione, C., Araneo, D., Latorre, C., Strecker, M., and Kendra, J. (2009). The stable isotope altimeter: do Quaternary pedogenic carbonates predict modern elevations? Geology 37: 1015–1018.
Hoke, G. D., Aranibar, J. N., Viale, M., Araneo, D. C., and Llano, C. (2013). Seasonal moisture sources and the isotopic composition of precipitation, rivers, and carbonates across the Andes at 32.5–35.5°S. Geochemistry, Geophysics, Geosystems 14: 962–978.
IAEA/WMO. (2006). Global network of isotopes in precipitation. The GNIP database. <http://isohis.iaea.org>.
Kellner, C., and Schoeninger, M. (2007). A simple carbon isotope model for reconstructing prehistoric human diet. American Journal of Physical Anthropology 133: 1112–1127.
Knudson, K. J. (2009). Oxygen isotope analysis in a land of environmental extremes: the complexities of isotopic work in the Andes. International Journal of Osteoarchaeology 19: 171–191.
Koch, P. L. (1998). Isotopic reconstruction of past continental environments. Annual Review of Earth Planetary Sciences 26: 573–613.
Lagiglia, H. (1976). La Cultura de Viluco del Centro Oeste Argentino. Revista del Museo de Historia Natural 3: 227–265.
Longinelli, A. (1984). Oxygen isotopes in mammal bone phosphate: a new tool for paleohydrological and paleoclimatological research? Geochimica et Cosmochimica Acta 48: 385–390.
Longinelli, A., and Nuti, S. (1973). Oxygen isotope measurements of phosphate from fish teeth and bones. Earth and Planetary Science Letters 20: 337–340.
López, J., Quiroga, M.; Frías, Carlos; Anzorena, J., and Araujo, E. (2011). Análisis zooarqueológico en el Área Fundacional de la Ciudad de Mendoza correspondiente al período colonial temprano (siglos XVI-XVII). Poster presented in II Congreso Nacional De Zooarqueología Argentina. Olavarría; Argentina.
Luz, B., Kolodny, Y., and Horowitz, M. (1984). Fractionation of oxygen isotopes between mammalian bone-phosphate and environmental drinking water. Geochimica et Cosmochimica Acta 48: 1689–1693.
Mafferra, L. (2010). La problemática en torno al registro arqueobotánico en el norte de Mendoza. In Chiavazza, H., and Bárcena, R. (eds.), Actas del XVII Congreso Nacional de Arqueología Argentina, Facultad de Filosofía y Letras. Universidad Nacional de Cuyo, Mendoza, pp. 2083–2088.
Mansegosa, D., and Chiavazza, H. (2010). Consecuencias del proceso de colonización en la salud de la población urbana de Mendoza (Argentina): un estudio desde evidencias paleopatológicas (S. XVIII–XIX). Revista Española de Paleopatología 8: 1–18.
Martínez Del Rio, C., Wolf, N., Carleton, S., and Gannes, Z. (2009). Isotopic ecology ten years after a call for more laboratory experiments. Biological Review 84: 91–111.
Michieli, C. T. (1998). Aproximaciones a la identificación de una cerámica indígena posthispánica del sur de San Juan. Publicaciones 22: 55–76. Instituto de Investigaciones Arqueológicas y Museo UNSJ. San Juan.
Newton, J. (2010). Stable Isotope Ecology. In Encyclopedia of Life Sciences John Wiley, Chichester. DOI: 10.1002/9780470015902.a0021231.
Panarello, H.O., and Dapeña, C. (1996). Mecanismos de recarga y salinización en las cuencas de los ríos Mendoza y Tunuyán, Mendoza, República Argentina evidenciados por isótopos ambientales. XII Congreso Geológico De Bolivia. Memorias, Tarija, Sociedad Geológica Boliviana, pp. 1–18.
Panarello, H., Tessone, A., and Zangrando, A. (2010). Isótopos estables en arqueología: principios teóricos, aspectos metodológicos y aplicaciones en Argentina. Xama 19–23: 115–133.
Parisii, M. (1991). Algunos datos de las poblaciones prehispánicas del Norte y Centro Oeste de Mendoza y su relación con la dominación Inca del área. Xama 4–5: 51–69.
Pate, D. (1994). Bone chemistry and paleodiet: reconstructing prehistoric subsistence settlement systems in Australia. Journal of Anthropological Archaeology 16: 103–120.
Petzke, K. J., Benjamin, T., Fullerb, C., and Metges, C. (2010). Advances in natural stable isotope ratio analysis of human hair to determine nutritional and metabolic status. Current Opinion in Clinical Nutrition and Metabolic Care 13: 532–540.
Sanhueza, L., and Falabella, F. (2010). Analysis of stable isotopes: from the Archaic to the horticultural communities in central Chile. Current Anthropology 50: 127–136.
Schiffer, M. (1987). Formation Processes of the Archaeological Record, University of New Mexico Press, Albuquerque.
Schwarcz, H., and Schoeninger, M. (1991). Stable isotope analysis in human nutritional ecology. Yearbook of Physical Anthropology 34: 283–321.
Sealy, J., Van Der Merwe, N., Lee-Thorp, J., and Lanham, J. (1987). Nitrogen isotopic ecology in southern Africa: implications for environmental and dietary tracing. Gecochemica et Comochinica Acta 51: 2707–2717.
Sponheimer, M., and Lee-Thorp, J. (1999). Oxygen isotopes in enamel carbonate and their ecological significance. Journal of Archaeological Science 26: 723–728.
Tessone, A. (2011). Arqueología y Ecología Isotópica, Estudio de Isótopos Estables de Restos Humanos del Holoceno Tardío en Patagonia Meridional. Doctoral dissertation, Universidad de Buenos Aires, Buenos Aires.
Turner, B., Kamenov, G., Kingston, J. D., and Armelagos, G. J. (2009). Insights into immigration and social class at Machu Picchu, Peru based on oxygen, strontium, and lead isotopic analysis. Journal of Archaeological Science 36: 317–332.
Tykot, R. H. (2006). Isotope analyses and the histories of maize. In Staller, J., Tykot, R., and Benz, B. (eds.), Histories of Maize, Academic, Waltham, pp. 131–142.
Tykot, R. H., Falabella, F., Planella, T., Aspillaga, E., Sanhueza, L., and Becker, C. (2009). Stable isotopes and archaeology in central Chile: methodological insights and interpretive problems for dietary reconstruction. International Journal of Osteoarchaeology 19: 156–170.
Ugan, A., and Coltrain, J. (2012). Stable isotopes, diet, and taphonomy: a look at using isotope-based dietary reconstructions to infer differential survivorship in zooarchaeological assemblages. Journal of Archaeological Science 39: 1401–1411.
Ugan, A., Neme, G., Gil, A., Coltrain, J., Tykot, R., and Novellino, P. (2012). Geographic variation in bone carbonate and water δ18O values in Mendoza, Argentina and their relationship to prehistoric economy and settlement. Journal of Archaeological Science 39: 2752–2763.
Van der Merwe, N., and Vogel, J. C. (1978). 13C content of human collagen as a measure of prehistoric diet in Woodland North America. Nature 276: 815–816.
Vogel, J., Lerman, J., and Mook, W. (1975). Natural Isotopes in Surface and Groundwater from Argentina. Hydrological Sciences-Bulletin-des Sciences Hidrologiques 20: 203–221.
White, C. D., Spence, M. W., Stuart-Williams, H., and Schwarcz, H. P. (1998). Oxygen isotopes and the identification of geographical origins: the Valley of Oaxaca versus the Valley of Mexico. Journal of Archaeological Science 25: 643–655.
White, C. D., Spence, M. W., Longstaffe, F. J., and Law, K. R. (2000). Testing the nature of Teotihuacán imperialism at Kaminaljuyú using phosphate oxygen-isotope ratios. Journal of Anthropological Research 56: 535–558.
White, C. D., Spence, M. W., Longstaffe, F. J., Stuart-Williams, H., and Law, K. R. (2002). Geographic identities of the sacrificial victims from the Feathered Serpent Pyramid, Teotihuacan: implications for the nature of state power. Latin American Antiquity 13: 217–236.
White, C. D., Longstaffe, F. J., and Law, K. R. (2004). Exploring the effects of environment, physiology and diet on oxygen isotope ratios in ancient Nubian bones and teeth. Journal of Archaeological Science 31: 233–250.
Acknowledgments
We wish to thank the Centro de Investigaciones Ruinas de San Francisco Team (Area Fundacional, Municipalidad de Mendoza), SECTyP (UNCuyo, Mendoza), Museo de Historia Natural de San Rafael, and CONICET. We also thank Luciano Valenzuela for his comments on a preliminary version. Two reviewers noted significant problems and they made comments and observation that we considered in a new version. We appreciate the help of Andy Froehle and his willingness to discuss the interpretation of the multivariate model in our study case. We also thank Cristina Ducos for the translation of the Spanish version into English, Fernando Franchetti and Mariana Gimenez for reviewing the text, and Miguel Giardina for making some of the figures for this paper.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Chiavazza, H., Mansegosa, D. & Gil, A. Human Diet and Residential Mobility in the Central Western Argentina Colony: Stable Isotopes (13C, 15N, 18O) Trends in Archaeological Bone Samples. Int J Histor Archaeol 19, 289–308 (2015). https://doi.org/10.1007/s10761-015-0288-3
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10761-015-0288-3