
Abstract
The aim of this study was to assess urbanization effects on microbial communities from low-order streams. Artificial substrata were placed upstream (control) and downstream (urban) of the cities of the selected streams. Photosynthetic parameters derived from chlorophyll-a fluorescence were measured using a Pulse of Amplitude Modulated Fluorometer, and the bacterial biofilm and bacterioplankton were counted by microscopy after staining with 4′,6-diamidino-2-phenylindole. We found higher bacterial biofilm biomass together with higher concentration of nutrients in urban reaches. The biofilm total density of bacteria was negatively correlated with the humic acid concentration, while the bacterioplankton total density was positively correlated with soluble reactive phosphorus and nitrite concentration. Autotrophic Index reflected the predominance of heterotrophs in the three streams. The concentration of chlorophyll-a, the minimum fluorescence, and the other photosynthetic parameters showed variations between the streams. These would respond to environmental factors at local scale not included in this study and may be influenced by the low development of autotrophic biomass at least in two of the streams studied. The bacterial morphotypes Small rod, Large rod, and Vibrio shaped (large) allowed the differentiation of urban reaches and would be useful as indicators of urbanization effects in both biofilm and bacterioplankton of lowland streams.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The urbanization of landscapes causes substantial alterations to watershed processes and subsequent stream function. Understanding the ecology of urban streams is increasingly important as more watersheds are impacted by human activities (Epstein et al., 2016). Urbanization affects the natural landscape causing the elimination of habitats, fragmentation, and isolation in the riparian zone. In this sense, the most significant changes are the input of toxic substances associated with domestic and industrial waste and the increase in impervious surfaces (Schueler, 1987). Changes introduced by anthropogenic activity alter the physical and chemical characteristics of the basin’s channels and its waters, influencing the biota and the functioning of these ecosystems (Pozo & Elosegui, 2009). Consequently, a reduction in the ecological integrity of river ecosystems and the capacity to provide environmental services is observed (Sweeney et al., 2004; Domínguez et al., 2020). The processes of population growth and urbanization in Argentina have occurred since the end of the nineteenth century and were accentuated during the first decades after the Second World War (Ainstein, 2012). Compared to other Latin American countries, Argentina showed an incipient and accelerated urbanization process, with a trend toward the concentration of activities in the Pampean Region. This raised a process of inequality in the distribution of the population and economic activities (Velázquez, 2000). The Pampean Region is an area of 426,000 km2 and has the highest population density in the country (Morello et al., 2000), with approximately 18 million inhabitants that represent 46% of the total population (INDEC, 2010) but the integrity of their aquatic ecosystems remains poorly evaluated.
The urban streams of the Pampean region characterized by naturally high concentrations of nutrients and humic acids (Feijoó & Lombardo, 2007) receive domestic and industrial effluents, in many cases with little or no treatment (Frau et al., 2019). Tagliaferro et al. (2019) found that urbanization in Pampean streams has a negative effect on the decomposition of organic matter and that it generates a change in the taxonomic structure of macroinvertebrate communities.
In Pampean streams characterized by high availability of nutrients, biofilms have shown to respond to slight variations in water quality related to the effects of urbanization, like developing a higher biomass with a higher metabolic activity (Cochero et al., 2018, 2021; Vilches et al., 2014). Biofilms are communities of microorganisms embedded in a mucopolysaccharides matrix that develop on different types of substrates in aquatic environments (Sabater et al., 2007). These are complex communities with great taxonomic and functional biodiversity, as they are made up of algae, bacteria, archaea, fungi, protozoa, and viruses (Besemer, 2016). In streams, biofilms are key communities where metabolic and enzymatic activity determine fundamental ecosystem processes, such as primary production and respiration (Battin et al., 2016).
In addition, biofilm integrates the effects of environmental conditions, which is why it has been widely used as a bioindicator in monitoring river ecosystems. (Biggs, 1989; Pizarro & Alemanni, 2005; Sabater et al., 2007; Montuelle et al., 2010; Monti et al., 2020). Therefore, multiple structural and functional biofilm endpoints are employed in laboratory and field studies for direct and indirect assessment of environmental impacts (Proia et al., 2012). For example, degradation of water quality given by high levels of nutrients and chemical oxygen demand can lead to increased algal biomass and bacterial growth, respectively (Tien et al., 2009). Findlay (2010) sustained that microbial processes need to be considered for stream management, restoration, and response to possible future scenarios of climate change. The heterotrophic microbial biomass is recognized as a key portion of trophic transfer, so its quantification is important (Findlay, 2010). The main role of heterotrophic microbes is crucial since they participate in the cycling and reuse of organic material that comes from primary production and from terrestrial vegetation (Sabater et al., 2009).
Monitoring of ecological status of aquatic systems has traditionally been based on the determination of water quality through the determination of physical and chemical characteristics and / or the analysis of community structure and biological indicators (Pu et al., 2019). Functional parameters arise from the measurement of changes in different ecological processes (e.g., primary production) and their inclusion in the environmental monitoring protocols allows assessing functional alterations of the systems as a consequence of anthropic activities (Gessner & Chauvet, 2002; Young et al., 2008). In recent decades, the use of parameters derived from the fluorescence emitted by chlorophyll-a to study structural and functional indicators of autotrophic communities has increased (Schmitt-Jansen et al., 2008). This type of study allows fast and precise in vivo detection of physiological and structural changes of all photosynthetic organisms in the sample at once (Schmitt-Jansen & Altenburger, 2008). This tool has been used in ecotoxicology as an endpoint in tolerance tests (Tlili et al.. 2011) and to study the effect of metals (Serra et al., 2009; Corcoll et al., 2011), emerging pollutants (Ricart et al., 2010), and herbicides on the biofilm (Pesce et al., 2010; Feckler et al., 2018). Also, Ponsatí et al., 2016 used this tool as indicator of biofilm function to analyze relative importance of land uses and chemical pollutants in Mediterranean river basins during contrasting hydrological periods. In the Pampas region, this method has been used to evaluate the water quality of streams affected by diffuse contamination with phosphorus in the chronic presence of arsenic (Rodríguez Castro, 2015).
Previous studies in fluvial environments of the Pampean region have determined bacterial biomass in the biofilm (Cochero et al., 2013; Sierra et al., 2013), but no studies have used Pulse of Amplitude Modulated Fluorometer (PAM) fluorometry as a tool to determine structural and functional parameters of the community, nor the analysis of bacterial morphotypes as bioindicators.
The aim of this study was to assess urbanization effects on microbial communities in low-order streams. For this purpose, we measured the autotrophic and bacterial biofilm biomass, the bacterioplankton biomass and functional parameters of autotrophic biofilm in three Pampean streams running through urban zones. Also, water quality was estimated by means of physical and chemical variables and the relation between those and the biological parameters was evaluated. The urbanization effects were analyzed at each stream by comparing the parameters measured upstream from the city (control reach) with those measured downstream from the city (urban reach). We hypothesized that i) the microbial communities are negatively affected by nutrient enrichment resulting from urbanization. We predicted that in urban reaches (i) the biofilm and bacterioplankton biomass would be higher as a consequence of the increase in nutrient concentration; (ii) the autotrophic biofilm functional parameters would reflect a lower physiological state as a consequence of environmental stress; and (iii) the bacterial morphotypes structure would be affected by the environmental conditions related with nutrient enrichment.
Materials and methods
Study area
The Pampean region is warm temperate and fully humid, according to Köppen’s climate classification, with mean temperature of 14 and 20 °C during winter and summer, respectively, and mean annual precipitation of 939 mm, with the maximum rainfall occurring in late summer (Matteucci, 2012). Pampean streams are characterized by mild slopes (0.1–1 km/m) throughout their course, resulting in slow current velocities (Rodrígues Capítulo et al., 2010). Stream beds are formed by an homogeneous and hard substratum with fine sediments (silt and clay). Humic substances in Pampean streams are naturally high, as well as alkalinity, conductivity, dissolved oxygen, and nutrient concentration (Amuchastegui et al., 2016). The landscape is characterized by the absence of forested riverbanks, where the dominant vegetation is herbaceous and there are only a few isolated native trees, such as Salix humboldtiana or Celtis tala (Feijoó & Lombardo, 2007). Most streams present a diverse community of macrophytes, which play a very important role as the main primary producers and provide habitat heterogeneity to the system (Giorgi et al., 2005). Instead of native vegetation, urban streams present introduced vegetation in its riverbank.
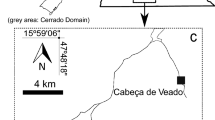
The study was carried out in three Pampean streams: La Choza, Giles, and Salgado, which cross the cities of General Rodríguez, San Andrés de Giles, and Lobos located in the Northeast area of Buenos Aires, Argentina (Fig. 1). The criteria for the selection of streams, previously described by Tagliaferro et al. (2019), were (1) Streams in association with small cities (< 100,000 people), (2) streams having similar hydrological characteristics, and (3) reference sites without cities on the riversides, nor close-by (at least 20 km distant from the riverside). At each stream, a “control reach,” upstream from the city, and an “urban reach,” downstream from the city, were selected. La Choza stream (34° 40′ S; 59° 09′ W) rises in an agricultural area south to Plomer Town and runs through an urban area where it receives wastewater from a chicken slaughterhouse and from residential neighborhoods (partially treated wastewaters and diffuse contamination). The Giles stream (34° 27′ S; 59° 27′ W) rises west to San Andrés de Giles city and runs through a natural area surrounded by native riparian vegetation with agricultural land use. Along the urban reach, the Giles stream receives partially treated wastewaters from a paper‐mill and residential diffuse contamination. This reach undergoes recurrent dredging on its channel margins. The Salgado stream (35° 08′ S; 59° 05′ W) belongs to the Salado river basin and delimits Lobos City in the western region. This stream was channelized at both control and urban reaches more than 15 years ago, and the channel has not received any additional management since then. However, the urban reach receives both partially treated wastewater from a part of the urban area and from a cattle slaughterhouse.

Location of the three streams studied in province of Buenos Aires, Argentina. The streams were shown in black lines. CABA: Ciudad Autónoma de Buenos Aires, Buenos Aires City
Environmental characterization
Samplings were carried out monthly between April and June of 2015. At each sampling site, a 100-m-long reach was selected in which wet width and depth were measured using a meter and water velocity with a propeller flowmeter (GM MFG co NYC). Discharge was calculated from these data by the area–velocity method (Elosegui et al., 2009). The percentage of coverage and the functional types of macrophytes present in each reach were also determined. At each sampling date (April, May and June) temperature, conductivity, pH, and dissolved oxygen concentration were measured using a probe (Thermo Scientific Orion Star A329) and water samples in triplicate were collected for nutrient analysis in polyethylene bottles (see Appendix S1). Water samples were stored at 4 °C and transported to the laboratory, where they were filtered within 2 h of collection using glass fiber filters (pore size = 0.7 mm). Analyses were conducted according to APHA (2005) and performed within 24 h from the sampling time. Soluble reactive phosphorus (SRP, mg P-PO4−3 l−1) was measured using the ascorbic acid method, nitrite (mg N-NO2− l−1) and nitrate (mg N-NO3− l−1) by reaction with sulfanilamide (with prior Cd reduction in the case of nitrate), and ammonium (µg N-NH4+ l−1) using the phenol-hypochlorite method. In addition, total suspended solids (TSS, mg l−1) were estimated by drying the glass fiber filters-previously weighed-to constant weight at 103–105 °C. The humic acids were estimated according to Lavado et al. (1982). All colorimetric determinations were analyzed with a Hitachi U-2001 spectrophotometer (Hitachi Ltd., Tokyo, Japan).
Biofilm and bacterioplankton
Artificial substrata were placed in each sampling site to evaluate the effect of urbanization on the communities of microorganisms. Forty substrata of etched glass (1 cm2) were adhered to the middle of both sides of concrete blocks. At the first sampling date the blocks were placed in the stream bed with the substrata parallel to the direction of the flow and thus the sediment could not cover them. The depth of the blocks was between 10 and 15 cm, so the light levels were sufficient to allow the development of an autotrophic community. The blocks were extracted from the sampling sites once the communities had developed, after 43 days of colonization. According to Tien et al. (2009), a one-month period of biofilm colonization is adequate for biomonitoring of water quality. At that point, the substrata were carefully peeled off the block, avoiding detachment of the biofilm layer. The colonized glasses were placed in compartmentalized containers with freshwater from the stream to avoid excessive movement, where they were transported to the laboratory (Stevenson & Bahls, 1999).
Once in the laboratory, photosynthetic parameters derived from chlorophyll fluorescence were measured using a Pulse of Amplitude Modulated Fluorometer (PAM) (FMS 1, Hansatech, England): minimum fluorescence yield (F0), maximum quantum yield (Fv/Fm), and effective quantum yield (ΦPSII). According to Rysgaard et al. (2001), F0 of dark-adapted cells at 665 nm is proportional to the chlorophyll-a concentration. F0 can be used as a structural parameter of the community, as it relates to algal biomass. The Fv/Fm parameter represents the maximum photosynthetic capacity and the ΦPSII represents the photosynthetic efficiency at a steady-state electron transport. These parameters are considered functional attributes of the autotrophic community since they arise from the measurement of the rates of different ecological processes and reflect their functioning (Maxwell & Johnson, 2000; Tambussi & Graciano, 2010). In this study we will refer to F0 parameter as minimum fluorescence, to Fv/Fm parameter as the maximal photosynthetic capacity, and to ΦPSII parameter as the photosynthetic efficiency of biofilm.
As PAM fluorometry is a fast and non-destructive technique, colonized glasses were also used to determine biofilm biomass after fluorescence measurements. Chlorophyll-a concentration (chl-a) was estimated by the Lorenzen method (Aminot, 1983) and the ash-free dry weight (AFDW) according to Aloi 1990. The Autotrophic Index (AI) which represents the ratio between AFDW (mg m−2) and chl-a values (mg m−2) (APHA, 2005) was calculated for each site.
For the analysis of the microbial structure of the bacterioplankton and the biofilm, samples were taken from the water column on each sampling occasion and from the artificial substrates at the end of the colonization. The samples were fixed with formalin (4%) and kept cold at 4ºC until analysis. Samples were filtered through 0.2-µm black polycarbonate filters, stained with 50 µl of 4′,6-diamidino-2-phenylindole (DAPI) (0.5 mg ml−1) for 10 min (Porter & Feig, 1980), and then mounted on a microscope slide with a drop of immersion oil for fluorescence. The inspection of samples was made at ×1000 magnification using Zeiss Axioplan microscope equipped with HBO 50 W lamp and a filter set for blue light, green light, and UV excitation. Bacteria were counted under UV light excitation and morphotypes were identified as bacterial cells that appeared at least once in each of the 25 fields observed per sample. The observed morphotypes were named according to their shape and size (Posh et al., 2009) (Fig. 2). The approximate size of each morphotype was estimated through photographs taken with a Canon Power Shot G10 camera and AxioVision software version 4.8 (Massana et al., 1997; Posch et al., 2009) (Appendix S2).

Length and schematic shape of the bacterial morphotypes found in the studied streams
Data analysis
Differences in nutrients concentration (SRP, N-NO2−, N-NO3− and N-NH4+), humic acids, TSS, algal biofilm metrics (chl-a, AFDW, F0, Fv/Fm, and ΦPSII), and bacterial metrics (total and morphotype densities) among reaches in each stream were analyzed by means of a one-way analysis of variance (ANOVA). Normality was tested with the Shapiro–Wilk’s goodness‐of‐fit test and transformed variables when necessary. Homogeneity of variance was tested by Levene’s test (Zar, 1999).
The association between the physical and chemical variables (SRP, N-NO2−, N-NO3− and N-NH4+, TSS, humic acids, conductivity, pH, dissolved oxygen, and temperature) and the reaches was performed using principal component analysis (PCA). The standardized mean values for each sampling occasion were used. Pearson correlations were performed to evaluate the relationship between the biological parameters (chl-a, AFDW, F0, Fv/Fm and ΦPSII, bacterial biofilm, and bacterioplankton total densities) and physical and chemical parameters, discharge, and macrophyte coverage. The mean values for each reach were used. The matrix of Pearson correlations can be found in the Supplementary Material (Appendix S3). All the analyses were conducted using R (3.6.0) in RStudio (1.4.1106) with the packages stats (R CoreTeam, 2019) car (Fox & Weisberg, 2019), vegan (Oksanen et al., 2020), and factoextra (Kassambara & Mundt, 2020).
Results
Environmental characterization
The pH and temperature were similar in all streams during the study. In Giles stream the conductivity was low and similar in both control and urban reaches, while in La Choza and Salgado streams, higher values were recorded in the control reaches (Table 1). These variables were correlated (R = 0.82. P < 0.05). The percentage of macrophyte coverage per section was low and similar between the La Choza reaches, while higher coverages were recorded in urban reach of Giles and control reach of Salgado. The Submerged and Emerging functional groups were present in the three streams in at least one of the sites, and the Floating group was only recorded in the urban reach of Salgado stream. (Table 1).
Nutrient concentrations were higher in urban reaches with the exception of nitrate in La Choza and Giles streams. For SRP and N-NO2− significant differences were found between reaches of the three streams (P < 0.05, n = 9). For N-NH4+ and N-NO3−, only significant differences were found between the La Choza and Salgado reaches (P < 0.05, n = 9). The N-NO2− was positively correlated with SRP (R = 0.95, P < 0.05), N-NH4+(R = 0.86, P < 0.005), and discharge (R = 0.87, P < 0.05). The N-NO3− was correlated with temperature (R = 0.83, P < 0.05) and N-NH4+ was negatively correlated with dissolved oxygen (R = -0.83, P < 0.05). Humic acids and TSS presented a different trend from the previous parameters, with higher average values in the control reaches and significant differences (P < 0.05, n = 9) only in Giles and Salgado streams (Table 2). Humic acids were positively correlated with pH (R = 0.87, P < 0.05) (see Appendix S3).
The ordination of the reaches based on the physical and chemical variables was carried out using a principal component analysis (PCA). The first two components of the PCA accumulated 63.73% of the total variance. The first component (percentage of variance explained: 37.1%) showed a positive correlation with N-NO2− (coefficient 0.84), SRP (0.78), and N-NH4+ (0.77) and a negative correlation with humic acids (-0.67) and TSS (-0.63). The second component (percentage of variance explained: 26.6%) showed a positive correlation with N-NO3− (0.79) and dissolved oxygen (0.76) and a negative correlation with conductivity (− 0.61) (Fig. 3). The urban reaches of La Choza and Salgado streams were associated with greater concentration of N-NO2−, SRP, and N-NH4+, while the control reaches of the three streams were associated with greater concentration of humic acids, TSS, and pH. The urban reaches of Giles stream were associated with greater concentration of N-NO3− and dissolved oxygen (Fig. 3).

Biplot of principal component 1 and 2 (Dim 1 and Dim 2, respectively) with the percentage of explained variance between parenthesis. The physical and chemical variables were represented by arrows. The sampling reaches were represented by different forms: circles, squares, and triangles correspond to La Choza, Giles, and Salgado streams, respectively. Green indicates control reach and red indicates urban reach, whereas the numbers 1 to 3 indicate the sampling occasion; temp: temperature, do: dissolved oxygen, nh4: ammonium, no2: nitrite, no3: nitrate, SRP: soluble reactive phosphorus, cond: conductivity, tss: total suspended solids, humic: humic acids
Biofilm and bacterioplankton
Biofilm biomass was estimated using Minimum Fluorescence (F0), chlorophyll-a concentration (chl-a), and ash-free dry weight (AFDW). Despite the great variability observed, the trend reflected by these three parameters is higher biomass in the control reaches compared to the urban ones in the three streams, with the exception of chl-a in the Giles and Salgado streams. Significant differences were found between reaches in La Choza and Salgado for chl-a (P < 0.05, n = 18, La Choza and Giles for F0 (P < 0.05, n = 40), and Salgado for the AFDW (P < 0.05, n = 18) (Table 3). The AFDW was positively correlated with TSS (R = 0.83, P < 0.05) and the macrophyte coverage (R = 0.87, P < 0.05). Significant correlations between F0 and chl-a concentration and the physical and chemical variables, discharge, and macrophyte coverage were not found.
Values of Autotrophic Index (AI) greater than 200 indicate a predominantly heterotrophic community with non-photosynthetic organisms and with organic debris. The results reflect the predominance of heterotrophs in the three streams (Table 3).
Maximal photosynthetic capacity (Fv/Fm) was higher in the control reaches compared to the urban ones of the La Choza and Salgado streams, while in the Giles stream, the situation was opposite (Fig. 4 A). For Fv/Fm, significant differences were found (P < 0.05, n = 40) between reaches of the three streams. A positive correlation was found between Fv/Fm, macrophyte coverage (R = 0.86, P < 0.05), and ɸPSII (R = 0.89, P < 0.05) (see Appendix S3). The ɸPSII was higher in the control reach than in the urban reach of the La Choza (P < 0.05, n = 9). Despite the same trend was observed in Salgado stream, significant differences were not found between reaches. In the Giles stream, ɸPSII was higher in the urban reach than in the control reach (P < 0.05, n = 9) (Fig. 4 B). Significant correlations between ɸPSII and the physical and chemical variables were not found.

A Maximum photosynthetic capacity (Fv/Fm, R.U.: random units) from biofilm samples in control (C) and urban (U) reaches. B: Photosynthetic efficiency (ɸPSII, R.U.: random units) from biofilm samples in control (C) and urban (U) reaches. The symbol ** represents significant differences between urban and control reaches, where P < 0.001
The total density of bacteria in the biofilm (cells cm−2) was significantly higher in the urban reaches than the controls for the three streams (P < 0.05, n = 6) (Fig. 5 A). A higher density of bacterioplankton was also recorded in urban reaches (Table 4), but significant differences were only found in the La Choza stream (P < 0.05, n = 18). The maximum total density recorded was 8.89.106 cells ml−1 and corresponds to the urban reach of Salgado, in which great variability can be observed (Fig. 5 B). The biofilm total density of bacteria was negatively correlated with the humic acid concentration (R = -0.84, P < 0.05). A positive and highly significant correlation between bacterioplankton total density and SRP was found (R = 0.98, P < 0.01) also with N-NO2− (R = 0.95, P < 0.01) (See Appendix S3).

A Biofilm total density (cell cm−2) in control and urban reaches. B Bacterioplankton total density (cell ml −1) in control and urban reaches. The symbols * and ** represent significant differences between urban and control reaches, where * indicates P < 0.05 and **P < 0.001
Significant differences (P < 0.05; n = 6) between reaches were found for all bacterial morphotypes present in the biofilm, between stream reaches. In all cases, except for the Cocci morphotype in the Salgado stream and the Filament morphotype in the La Choza stream, the number of cells cm−2 was higher in the urban reach. The Vibrio shape (large) morphotype was recorded at the control reach of La Choza stream and at all urban sites (Fig. 6 A). In the bacterioplankton, all the morphotypes registered higher average values in the urban reaches compared to the controls, except for the Cocci morphotype in Giles and Salgado streams and the Filament morphotype in Salgado stream (Table 4). In La Choza stream, significant differences were found for the morphotypes Small rod and Large rod, in Giles stream for Vibrio shaped (small), Small rod, Large rod, and Filament, and in Salgado stream for all morphotypes (P < 0.05, n = 18). The Vibrio-shaped (large) morphotype was only registered in the urban reaches of the streams (Fig. 6 B).

A Biofilm bacterial morphotypes (cell cm−2) in control (C) and urban (U) reaches. B Bacterioplankton morphotypes (cell ml−1) in control (C) and urban (U) reaches
Discussion
The present study aimed to assess the effects of urbanization on the microbial communities from low-order streams. We hypothesized that the microbial communities were negatively affected by nutrient enrichment resulting from urbanization. Our predictions were partially met: i) the bacterial biofilm biomass was higher in the three urban reaches in agreement with an increase of nutrient concentration (SRP, N-NO2−, and N-NH4+), while the autotrophic biofilm and bacterioplankton biomass were more variable between streams suggesting that other environmental variables could have influence in the structure of these communities; ii) the autotrophic biofilm functional parameters did not reflect a lower physiological state in urban reaches, they showed a variable trend according to the autotrophic biomass; and iii) the bacterial morphotypes structure were affected by nutrient enrichment in both biofilm and bacterioplankton communities, with a significantly higher biomass of the Small rod, Large rod, and Vibrio-shaped (large) morphotypes in the three urban reaches.
In urban reaches, the concentrations of SRP and most of the inorganic nitrogen compounds exceed the reference values for Pampean streams with low disturbance (Feijoó & Lombardo, 2007). This could be explained by the nutrient inputs from point sources related to the urban area nearby (industrial and municipal effluents, sewage treatment plants) and runoff and infiltration processes from areas with accumulation of domestic waste which are cited as the main anthropogenic sources of inorganic nitrogen in aquatic ecosystems (Camargo & Alonso, 2006). Our results are consistent with those of Tagliaferro et al., 2019 in a previous study carried out in these same streams, in which it was considered that eutrophication due to the high concentrations of SRP, nitrites, and ammonia was the main stressor associated with urban reaches. Also, Cochero et al., 2021 found a higher concentration of SRP in urban reaches of three Pampean streams and higher levels of nitrites, nitrates, and ammonium in the most strongly impacted site by urbanization. On the other hand, the higher concentrations of humic acids and TSS in control reaches could be related to the loss of riparian vegetation with the urban use of the surrounding land.
Biofilm is a key community in the functioning of river systems (Battin et al., 2016) and has been widely used to monitor its ecological status (Sabater et al., 2007; Serra et al., 2009; Montuelle et al., 2010; Monti et al., 2020). Structural parameters such as F0, chl-a concentration, and AFDW are of great importance for the analysis of the ecological integration of the Pampean streams affected by urbanization. In streams with urban influence, mainly as a consequence of a higher nutrient load, increases in the biomass of the biofilm were recorded (Taylor et al., 2004; Catford et al., 2007; O'Brien & Wehr, 2010). In the Pampas Region, higher concentrations of nutrients and chlorophyll-a were observed in the most urbanized streams (Sierra et al., 2013). However, Walsh et al. 2005 indicate that, in systems without nutrient limitation, in the absence of urban impact, such an increase is less likely. Our results show a higher bacterial biofilm biomass in urban reaches, mainly as a consequence of a higher nutrient load. The lower development of autotrophic biofilm in urban reaches could be attributed to other environmental variables related to urban influence. This difference in colonization is frequent in water courses polluted by wastewater discharges, where toxicity generated from substances released from sediments or directly from effluents with scarce or without water treatment (Ricart et al., 2010) explain a higher proliferation of the heterotrophic component of the biofilm, competing for substrata colonization (Masseret et al., 1998; Valdés et al., 2021). The presence of a higher density of bacterial biofilm and the heterotrophic condition suggested by the AI supports this idea.
The parameter F0 reflected a higher biomass of the autotrophic fraction in the control reaches of La Choza and Giles streams. This parameter is consistent with the concentrations of chl-a in La Choza and Salgado streams, while in Giles stream F0 of the urban reach underestimates the high concentration of chl-a. This indicates that it is not always possible to use F0 as an indicator of algal biomass because excessive development of the biofilm alters the linear relationship between F0 and chl-a concentration (Schmitt-Jansen & Altenburger, 2008; Corcoll et al., 2012).
The measurement of the fluorescence emitted by chlorophyll-a allows determining functional parameters of the community linked to the photosynthetic process and in particular to the primary reactions that take place in the PSII (Maxwell &Johnson 2000; Tambussi & Graciano, 2010). Different physical or chemical factors of environmental stress such as changes in light intensity, nutritional deficiencies, presence of heavy metals, and detergents, among others affect the photosynthesis and modify the emission of fluorescence (Moreno et al., 2008). Thus, these changes can be used to reveal response mechanisms, quantify responses to stress, and identify pollutants and their sources (Maxwell & Johnson, 2000). The maximum photosynthetic capacity (Fv/Fm) for the control reaches of the three streams is similar to those found by Corcoll et al. (2011) in biofilm of a non-polluted river. These results would reflect a greater integrity of the photosynthetic apparatus of the biofilm in the reaches that do not receive the impact of urbanization (Bilger et al., 1995; Maxwell & Johnson, 2000), except for Giles stream. In the Giles stream, the use of this parameter could be rejected, since the F0 value intervenes for its estimation, which indicated an underestimation of the biomass of the biofilm. It should be noted that the Giles urban reach was dredged prior to samplings, so the development of the autotrophic biomass could have been affected due to the higher concentration of available nutrients and the existence of new niches (Licursi & Gómez, 2009). The Photosynthetic Efficiency (ɸPSII) was higher in the control reach of La Choza stream, reflecting like Fv/Fm a better physiological state of the community in the reach that does not receive the influence of urbanization. While, in Giles stream it was higher in the urban reach, which would indicate that the community on this site is not affected in its photosynthetic functioning. In the Salgado stream, ɸPSII was not significantly different between control and urban reaches. Thus, this parameter could be less sensitive to detect the effects on algae of toxic compounds depending on the time of exposure of the toxicant substances (Corcoll et al., 2012). One of most important environmental factors that influence the photosynthesis of the autotrophic communities is the irradiance that reaches the streambed (Hill, 1996; Hill & Dimick, 2002). The Pampean low-order streams has a natural riparian vegetation mainly herbaceous (Feijoó & Lombardo, 2007). Without riverine shaded the solar radiation easily reaches the streambed allowing a high primary gross production given by the epiphyton and the bottom algae (Vilches & Giorgi, 2010). The light intensity it would not be a limiting factor for the photosynthesis in the studied streams. The variability of the results could be related to differences at local scale between streams and to the low development of the autotrophic biomass at least in Salgado stream. The use of parameters derived from chlorophyll-a fluorescence to detect changes in the structure and functioning of the biofilm and to evaluate the effect of a stressor such as urbanization in lowland Pampean streams has no precedents. Corcoll et al. (2012) recommend the implementation of these bioindicators in the study of river ecosystems due to their ability to provide warning signals to the effects of toxicity. Nevertheless, these parameters were variable between the streams suggesting the influence of others factors not included in our study so further studies will be necessary to explain the variability.
In low-order streams, microbial biomass in sediments or attached to surfaces is usually greater than planktonic biomass and is the main responsible for the processing of organic matter (Pusch et al., 1998; Findlay et al., 2002). However, in rivers with low current velocity, the development of the bacterial planktonic community can be important (Freese et al., 2006), establishing an intimate relationship with the particles transported by the system (Romani et al., 2009). The diversity of these communities and the variety of their metabolic capacities give them a key role in river ecological processes. For this reason, their study is of interest for determining the ecological status of fluvial courses, since they are considered indicators of their health, and of importance in systems that have suffered disturbances. (Burns & Ryder, 2001; Freese et al., 2006; Findlay, 2010). Bacterioplankton density exhibited a similar trend in the three streams, with higher values in the urban reaches. Schumann et al. (2003) determined bacterial densities between 4.0 and 10.9.106 cell.ml−1 in a shallow river with low current velocity and eutrophic. According to these authors, this range would be typical of several freshwater systems with different trophic degrees. Our results were within this range of densities. In urban reaches, the registered values were similar to those found by Freese et al. (2006) during the fall in an eutrophic river characterized by a low current speed. These authors found a correlation between bacterial density and water temperature, which explained the lower density recorded in cold seasons. In the present study, the temperatures at both sites of each stream were similar, so the differences observed were not due to temperature differences.
The bacterial density of the biofilm allowed the differentiation of the three reaches of streams, with higher values in the urban reaches. The estimated densities were comparable with those registered by Cochero et al. (2021) in the epipelic biofilm of urbanized Pampean streams. These authors also found higher bacterial densities in urban reaches compared with peri-urban sites at each stream studied. Cochero et al. (2013) recorded a positive correlation between biofilm bacterial density and soluble reactive phosphorus (SRP) concentration. In the present study, we found a positive correlation between SRP concentration and density of bacterioplankton but not with biofilm bacterial density. The urban sites of the three streams were characterized by presenting higher concentrations of PRS and some or all of the inorganic forms of nitrogen as a consequence of urbanization, which could be responsible for the increase in the bacterial density of the biofilm. Carr et al. (2005) found a positive correlation between bacterial abundance and the biomass and algal production of the biofilm, suggesting that algae and bacteria coexist in an association that offers the space and resources to sustain the production of both groups of organisms. Both Cochero et al. (2013) as Sierra et al. (2013) registered an increase in algal biomass associated with an increase in bacterial density. In this study, the algal biomass indicator parameters of the reaches impacted by urbanization were not greater than those not impacted. The higher bacterial density was not accompanied by a higher algal biomass, possibly due to the influence of factors that limit the development of the primary producers, but not of the bacterial biofilm, such as the presence of substances that are toxic to the algae from urban and industrial waste, as previously mentioned (Masseret et al., 1998; Valdés et al., 2021). From the results obtained, we consider that the bacterial density of the biofilm is an adequate parameter to detect the effect of urbanization in Pampean streams.
Bacterial abundance, size, and biomass are important variables in aquatic ecosystems (Cole et al., 1993). Size determines aspects of the metabolism of organisms and in the case of prokaryotes it is an easier parameter to determine than genetic identity and can provide important ecological information (Massana et al., 1997). The bacterial morphotypes can be distinguished by its shape and size. The dominant morphotypes in the bacterioplankton samples from the three streams were the smallest in size (Cocci and Small rod). In all cases, the density of the largest morphotypes (Filament, Large rod, and Vibrio shaped (large)) was much lower than the density of the dominant morphotype. These results are similar to those obtained by Quiroga et al. (2017) in the bacterioplankton of bog lagoons and those of Velimirov et al. (2011) in this same community along the Danube River. These authors found a dominance of small morphotypes with little representation of larger shapes. In general, the phenotypic structure of bacterioplankton is strongly influenced by selective size predation (Jürgens & Matz, 2002) by planktonic flagellates and bacterial ciliates, which preferentially consume bacterial cells with a size between 1 and 3 µm (Pernthaler, 2005). The morphotypes that made it possible to differentiate the reaches of the three streams were Small rod and Large rod, with higher densities in urban sites. In addition, the Vibrio-shaped (large) morphotype, characterized by presenting an intermediate size (3.2–4.8 µm), was only registered in the urban reach of the streams. The conditions for its development would only be given in the sites with urban influence and could be related both to the concentration of nutrients in these sites, since the abundance and biomass of the microorganisms increase with the increase in the availability of nutrients (Sommaruga & Psenner, 1995). In the biofilm samples from the three streams, the dominant morphotype was Small rod. In this case, the pressure exerted by selective predation that could have been responsible for the dominance of the Cocci morphotype in bacterioplankton would not be evident. But as in this one, the density of the larger morphotypes was much lower than the density of the smaller ones. These results are similar to those obtained by Romaní & Sabater (1999) in the epilithic biofilm of a Mediterranean river with high concentrations of nutrients, in which they registered the dominance of small morphotypes during an annual period. All the morphotypes found in the biofilm allowed the differentiation of stream reaches, with higher densities in urban reaches, with the exception of the Cocci morphotype in Salgado stream. The Vibrio-shaped (large) morphotype was not exclusively registered in the reaches with urban influence since it was observed in the control reach of the La Choza stream, although its density was very low. Although there is no history of studying the density of bacterial morphotypes as possible indicators of the effect of urbanization in aquatic systems, Quiroga et al. (2017) affirm that planktonic bacteria from peatlands and other wetlands respond rapidly to anthropogenic disturbances and can be considered as useful bioindicators of changes in these systems, since monitoring the structure and function of this community has management and conservation implications. The results of the present work indicate that the bacterial morphotypes, Small rod, Large rod, and Vibrio shaped (large), would be useful as indicators of urbanization effects in both biofilm and bacterioplankton of Pampean streams.
Conclusion
In conclusion, the ecological integrity of the streams was affected by urbanization, which can be observed trough studying biological indicators. The main change observed was the increase in the concentration of nutrients, in particular SRP and inorganic forms of nitrogen. Stream biofilm was predominantly heterotrophic, so autotrophic biomass estimators and functional parameters would not be the most appropriate parameters to evaluate the effects of urbanization. However, the bacterial density of biofilm and the structure of morphotypes in both biofilm and bacterioplankton reflected the urban influence and would be adequate to evaluate the effects of urbanization in the Pampean streams. Given that the Pampean Region supports a high demographic density (INDEC, 2010) and that worldwide the advance of urbanization is an accelerated phenomenon (Niemelä et al., 2010), these results could be used as base information for the realization of future studies in the zone.
References
Ainstein, L., 2012. Urbanización, medio ambiente y sustentabilidad en Argentina. Cuaderno Urbano Resistencia 12(12): 173–189.
Aloi, J. E., 1990. A critical review of recent freshwater periphyton field methods. Canadian Journal Fisheries Aquatic Science 47: 656–670.
Aminot, A., 1983. Dosage de la chlorophylleet des phéopigment par spectrophotométric. Pp. 177–189 pp en A. Aminot and M. Chansspied, (eds) Manual des analyses chimiques en Milienmarin Centre National pour L´Explotation des Océans, Brest, Francia.
Amuchástegui, G., L. di Franco & C. Feijoó, 2016. Catchment morphometric characteristics, land use and water chemistry in Pampean streams: a regional approach. Hydrobiologia 767: 65–79.
APHA, 2005. Standard Methods for the examination of water and waste water. A. Eaton, L. Clesceri, E. Rice, & A. Greenberg, (eds) American Public Health Association.
Basílico, G., L. de Cabo & A. Faggi, 2013. Impacts of composite wastewater on a Pampean stream (Argentina) and phytoremediation alternative with Spirodela intermedia Koch (Lemnaceae) growing in batch reactors. Journal of Environmental Management 115: 53–59.
Battin, T. J., K. Besemer, M. M. Bengtsson, A. M. Romani & A. I. Packmann, 2016. The ecology and biogeochemistry of stream biofilms. Nature Reviews Microbiology 14(4): 251.
Bauer, D. E., M. E. Conde & N. Gómez, 2002. Phytoplankton of a small lowland stream related to water quality and hydraulic discontinuities. Archiv Fur Hydrobiologie 153(3): 421–442.
Berggren, M., H. Laudon, M. Haei, L. Ström & M. Jansson, 2010. Efficient aquatic bacterial metabolism of dissolved low-molecular-weight compounds from terrestrial sources. The ISME Journal 4(3): 408–416.
Besemer, K., 2016. Microbial biodiversity in natural biofilms. In en Romaní, A., H. Guasch & M. Balaguer (eds), Aquatic Biofilms. Ecology, Water Quality and Wastewater Treatment Caister Academic Press, Norfolk, UK: 63–87.
Biggs, B. J., 1989. Biomonitoring of organic pollution using periphyton, South Branch, Canterbury, New Zealand. New Zealand Journal of Marine and Freshwater Research 23(2): 263–274.
Bilger, W., U. Schreiber & M. Bock, 1995. Determination of the quantum efficiency of photosystem II and of non-photochemical quenching of chlorophyll fluorescence in the field. Oecologia 102(4): 425–432.
Burns, A. & D. S. Ryder, 2001. Potential for biofilms as biological indicators in Australian riverine systems. Ecological Management & Restoration 2(1): 53–64.
Camargo, J. A. & A. Alonso, 2006. Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: a global assessment. Environment International 32(6): 831–849.
Carr, G. M., A. Morin & P. A. Chambers, 2005. Bacteria and algae in stream periphyton along a nutrient gradient. Freshwater Biology 50(8): 1337–1350.
Catford, J. A., C. J. Walsh & J. Beardall, 2007. Catchment urbanization increases benthic microalgal biomass in streams under controlled light conditions. Aquatic Sciences 69(4): 511–522.
Cochero, J., A. M. Romaní & N. Gómez, 2013. Delayed response of microbial epipelic biofilm to nutrient addition in a Pampean stream. Aquatic Microbial Ecology 69(2): 145–155.
Cochero, J., M. M. Nicolosi Gelis, M. B. Sathicq & N. Gomez, 2018. Biofilm early stage development in two nutrient-rich streams with different urban impacts. River Research and Applications 34(7): 755–764.
Cochero, J., Gelis, M. M. N., Donadelli, J., & Gomez, N. ,2021. Translocation of Epipelic Biofilms and Their Short-Term Responses to Urbanization Impacts in Nutrient Rich Streams. Anais da Academia Brasileira de Ciências, 93.
Cole, J. J., S. Findlay & M. L. Pace, 1988. Bacterial production in fresh and saltwater ecosystems: a cross-system overview. Marine Ecology Progress Series. Oldendorf 43(1): 1–10.
Cole, J. J., M. L. Pace, N. F. Caraco & G. S. Steinhart, 1993. Bacterial biomass and cell size distributions in lakes: more and larger cells in anoxic waters. Limnology and Oceanography 38(8): 1627–1632.
Corcoll, N., B. Bonet, M. Leira & H. Guasch, 2011. Chl-a fluorescence parameters as biomarkers of metal toxicity in fluvial biofilms: an experimental study. Hydrobiologia 673: 119–136.
Corcoll, N., Ricart, M., Franz, S., Sans-Piché, F., Schmitt-Jansen, M., & Guasch, H., 2012. The use of photosynthetic fluorescence parameters from autotrophic biofilms for monitoring the effect of chemicals in river ecosystems. In Emerging and priority pollutants in rivers (pp. 85–115). Springer, Berlin, Heidelberg.
Domínguez, E., Giorgi, A., Miserendino, M.A., Marchese, M., & Gómez N., 2020. Problemáticas de cuencas en Argentina. Recomendaciones para su gestión. Pp: 259–272. En Domínguez, E., Giorgi, A., & N. Gómez (eds). La bioindicacion en el monitoreo y evaluación de los sistemas fluviales de la Argentina.
Elosegi, A., Butturini, A. & Armengol J., 2009. El caudal circulante. Pp 51–69 en: A. Elosegui & S. Sabater (Eds), Conceptos y técnicas en ecología fluvial. Fundación BBVA, España.
Epstein, D., J. Kelso & M. Baker, 2016. Beyond the urban stream syndrome: organic matter budget for diagnostics and restoration of an impaired urban river. Urban Ecosystem 19: 1623–1643.
Feckler, A., J. Rakovic, M. Kahlert, R. Tröger & M. Bundschuh, 2018. Blinded by the light: Increased chlorophyll fluorescence of herbicide-exposed periphyton masks unfavorable structural responses during exposure and recovery. Aquatic Toxicology 203: 187–193.
Feijoó, C. & R. J. Lombardo, 2007. Baseline water quality and macrophyte assemblages in Pampean streams: a regional approach. Water Research 41: 1399–1410.
Findlay, S., 2010. Stream microbial ecology. Journal of the North American Benthological Society 29: 170–181.
Findlay, S., J. Tank, S. Dye, H. M. Valett, P. J. Mulholland, W. H. McDowell & W. B. Bowden, 2002. A cross-system comparison of bacterial and fungal biomass in detritus pools of headwater streams. Microbial Ecology 43(1): 55–66.
Fox, J.& Weisberg, S.,2019. An {R} Companion to Applied Regression, Third EditionThousand Oaks CA: Sage. https://socialsciences.mcmaster.ca/jfox/Books/Companion/.
Frau, D., J. Medrano, C. Calvi & A. Giorgi, 2019. Water quality assessment of a neotropical pampean lowland stream using a phytoplankton functional trait approach. Environmental Monitoring and Assessment 191(11): 681.
Freese, H. M., U. Karsten & R. Schumann, 2006. Bacterial abundance, activity, and viability in the eutrophic river Warnow, Northeast Germany. Microbial Ecology 51(1): 117–127.
Gabellone, N. A., M. C. Claps, L. C. Solari & N. C. Neschuk, 2005. Nutrients, conductivity and plankton in a landscape approach to a Pampean saline lowland river (Salado River, Argentina). Biogeochemistry 75(3): 455–477.
Gessner, M. O. & E. Chauvet, 2002. A case for using litter breakdown to assess functional stream integrity. Ecological Applications 12: 498–510.
Giorgi, A., C. Feijoó & H. G. Tell, 2005. Primary producers in a Pampean stream: temporal variation and structuring role. Biodiversity and Conservation 14(7): 1699–1718.
Hill, W., 1996. Effects of light. Pp. En: Stevenson, R.J.; Bothwell, M. L. & Lowe, R.L. (Eds) Algal ecology: Freshwater benthic ecosystems. Academic Press, USA.
Hill, W. R. & S. M. Dimick, 2002. Effects of riparian leaf dynamics on periphyton photosynthesis and light utilisation efficiency. Freshwater Biology 47(7): 1245–1256.
INDEC, 2010. Censo Nacional de Población, Hogares y Viviendas 2010. Instituto Nacional de Estadística y Censos. www.indec.gov.ar.
Jürgens, K. & C. Matz, 2002. Predation as a shaping force for the phenotypic and genotypic composition of planktonic bacteria. Antonie Van Leeuwenhoek 81(1–4): 413–434.
Kassambara, A., & Mundt, F., 2020. Package ‘factoextra’. Extract and Visualize the Results of Multivariate Data Analyses. http://www.sthda.com/english/rpkgs/factoextra
Lavado, R., O. Duymovich, J. Gimenez & L. Alvarez, 1982. Pérdidas de sustancias húmicas de suelos nátricos a través del río Samborombón. Revista Museo de La Plata IX, Geología 76: 97–103.
Licursi, M. & N. Gomez, 2009. Effects of dredging on benthic diatom assemblages in a lowland stream. Journal of Environmental Management 90(2): 973–982.
Massana, R., J. M. Gasol, P. K. Bjørnsen, N. Blackburn, Å. Hagström, S. Hietanen & C. Pedrós-Alió, 1997. Measurement of bacterial size via image analysis of epifluorescence preparations: description of an inexpensive system and solutions to some of the most common problems. Scientia Marina 61: 397–407.
Masseret, E., C. Amblard & G. Bourdier, 1998. Changes in the structure and metabolic activities of periphytic communities in a stream receiving treated sewage from a waste stabilization pond. Water Research 32(8): 2299–2314.
Matteucci, S. D., 2012. Ecorregion Pampa. Pp. 391–445 en J. Morello, S. Matteucci, & A. Rodriguez (eds), Ecorregiones y Complejos Ecosistémicos Argentinos. Orientación Grafica Editora, Buenos Aires.
Maxwell, K. & G. N. Johnson, 2000. Chlorophyll fluorescence-a practical guide. Journal of Experimental Botany 51(345): 659–668.
Monti, D., C. Hubas, X. Lourenço, et al., 2020. Physical properties of epilithic river biofilm as a new lead to perform pollution bioassessments in overseas territories. Sci Rep 10: 17309. https://doi.org/10.1038/s41598-020-73948-7.
Montuelle, B., U. Dorigo, A. Bérard, B. Volat, A. Bouchez, A. Tlili & S. Pesce, 2010. The periphyton as a multimetric bioindicator for assessing the impact of land use on rivers: an overview of the Ardières-Morcille experimental watershed (France). Hydrobiologia 657(1): 123–141.
Morello, J., G. Buzai, C. Baxendale, A. Rodriguez, S. Matteucci, R. Godagnone & R. Casas, 2000. Urbanización y consumo de tierra fértil. Ciencia Hoy 10: 50–61.
Moreno, S. G., H. P. Vela & M. O. Alvarez, 2008. La fluorescencia de la clorofila a como herramienta en la investigación de efectos tóxicos en el aparato fotosintético de plantas y algas. Revista De Educación Bioquímica 27(4): 119–129.
Niemelä, J., S. Saarela, T. Soderman, L. Kopperoinen, V. Yli-pelkonen, S. Vaare & D. Kotze, 2010. Using the ecosystem services approach for better planning and conservation of urban green spaces: a Finland case study. Biodiversity Conservation 19: 3225–3243.
O’Brien, P. J., & Wehr, J. D., 2009. Periphyton biomass and ecological stoichiometry in streams within an urban to rural land-use gradient. In Global Change and River Ecosystems—Implications for Structure, Function and Ecosystem Services (pp. 89–105). Springer, Dordrecht.
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., Mcglinn, D., Minchin, P. R., O’hara, R. B., Simpson, G. L., Solymos, P., Henry, M., Stevens, H., Szoecs, E., & Maintainer, H. W., 2020. Package “vegan” Community Ecology Package Version 2.5–7.
Pesce, S., S. Lissalde, D. Lavieille, C. Margoum, N. Mazzella, V. Roubeix & B. Montuelle, 2010. Evaluation of single and joint toxic effects of diuron and its main metabolites on natural phototrophic biofilms using a pollution-induced community tolerance (PICT) approach. Aquatic Toxicology 99(4): 492–499.
Pernthaler, J., 2005. Predation on prokaryotes in the water column and its ecological implications. Nature Reviews Microbiology 3(7): 537–546.
Piccinini, M. A., 2016. Capacidad del arroyo Salgado, Partido de Lobos, para depurar efluentes cloacales, Universidad Nacional de Luján, Tesis de Post-grado:
Pizarro, H. & M. Alemanni, 2005. Variables físico-químicas del agua y su influencia en la biomasa del perifiton en un tramo inferior del Río Luján (Provincia de Buenos Aires). Ecología Austral 15(1): 73–88.
Ponsatí, L., N. Corcoll, M. Petrović, Y. Picó, A. Ginebreda, E. Tornés & S. Sabater, 2016. Multiple-stressor effects on river biofilms under different hydrological conditions. Freshwater Biology 61(12): 2102–2115.
Porter, K. G. & S. Feig, 1980. The use of DAPI for identifying and counting aquatic microflora. Limnology and Oceanography 25: 943–948.
Posch, T., J. Franzoi, M. Prader & M. M. Salcher, 2009. New image analysis tool to study biomass and morphotypes of three major bacterioplankton groups in an alpine lake. Aquatic Microbial Ecology 54(2): 113–126.
Pozo, J. & Elosegui, A., 2009. El marco físico: la cuenca. Pp. 39–49 en: Elosegui, A. & S. Sabater (eds). Conceptos y técnicas en ecología fluvial. Fundación BBVA, España.
Proia, L., Cassió, F., Pascoal, C., Tlili, A., & Romaní, A. M., 2012. The use of attached microbial communities to assess ecological risks of pollutants in river ecosystems: the role of heterotrophs. In Emerging and Priority Pollutants in Rivers (pp. 55–83). Springer, Berlin, Heidelberg.
Pu, Y., W. Y. Ngan, Y. Yao & O. Habimana, 2019. Could benthic biofilm analyses be used as a reliable proxy for freshwater environmental health? Environmental Pollution 252: 440–449.
Pusch, M., D. Fiebig, I. Brettar, H. Eisenmann, B. K. Ellis, L. A. Kaplan & M. Traunspurger, 1998. The role of micro-organisms in the ecological connectivity of running waters. Freshwater Biology 40(3): 453–495.
Quiroga, M. V., G. Mataloni, B. M. Wanderley, A. M. Amado & F. Unrein, 2017. Bacterioplankton morphotypes structure and cytometric fingerprint rely on environmental conditions in a sub-Antarctic peatland. Hydrobiologia 787(1): 255–268.
R Core Team, 2019. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/.
Ricart, M., H. Guasch, M. Alberch, D. Barceló, C. Bonnineau, A. Geiszinger, M. la Farré, J. Ferrer, F. Ricciardi, A. Romani, S. Morine, L. Proia, L. Sala, D. Sureda & S. Sabater, 2010. Triclosan persistence through wastewater treatment plants and its potential toxic effects on river biofilms. Aquatic Toxicology 100(4): 346–353.
Rodríguez Castro, M. C., 2015. Capacidad de depuración de sustancias bioaprovechables en arroyos de llanura y su relación con el arsénico. Tesis Doctoral, Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires.
Rodrígues Capítulo, A., N. Gómez, A. Giorgi, & C. Feijoó, 2010. Global changes in pampean lowland streams (Argentina): Implications for biodiversity and functioning. Hydrobiologia 657: 53–70.
Romaní, A. M. & S. Sabater, 1999. Epilithic ectoenzyme activity in a nutrient-rich Mediterranean river. Aquatic Sciences-Research across Boundaries 61(2): 122–132.
Romaní, A. M., Artigas, J., Camacho, A., GraÇa, M. A. S. & C. Pascoal, 2009. La biota de los ríos: los microorganismos heterotróficos. Pp. 169–218 en: Elosegui A. & S. Sabater (eds). Conceptos y técnicas en ecología fluvial. Fundación BBVA, España.
Rysgaard, S., M. Kühl, R. N. Glud & J. W. Hansen, 2001. Biomass, production and horizontal patchiness of sea ice algae in a high-Arctic fjord (Young Sound, NE Greenland). Marine Ecology Progress Series 223: 15–26.
Sabater, S. & Elosegui, A., 2009. Otros factores físicos de importancia para los seres vivos: luz, temperatura, corriente. Pp: 133–140 en: A. Elosegui & S. Sabater (Eds), Conceptos y técnicas en ecología fluvial. Fundación BBVA, España.
Sabater, S., H. Guasch, M. Ricart, A. Romaní & G. Vidal, 2007. Monitoring the effect of chemicals on biological communities. The biofilm as an interface. Analytical and Bioanalytical Chemistry 387: 1425–1434.
Sabater, S., J. Donato, A. Giorgi & Elosegui, A., 2009. El río como ecosistema. Pp. 23–37 en: Elosegui A. & S. Sabater (eds). Conceptos y técnicas en ecología fluvial. Fundación BBVA, España.
Schmitt-Jansen, M. & R. Altenburger, 2008. Community-level microalgal toxicity assessment by multiwavelength-excitation PAM fluorometry. Aquatic Toxicology 86: 49–58.
Schmitt-Jansen, M., U. Veit, G. Dudel & R. Altenburger, 2008. An ecological perspective in aquatic ecotoxicology: approaches and challenges. Basic and Applied Ecology 9: 337–345.
Schueler, T., 1987. Controlling Urban Runoff: A Practical Manual for Planning and Designing Urban BMPs, Metropolitan Washington Council of Governments, Washington, DC:
Schumann, R., T. Rieling, S. Görs, A. Hammer, U. Selig & U. Schiewer, 2003. Viability of bacteria from different aquatic habitats. I. Environmental conditions and productivity. Aquatic Microbial Ecology 32(2): 121–135.
Serra, A., N. Corcoll & H. Guasch, 2009. Copper accumulation and toxicity in fluvial periphyton: the influence of exposure history. Chemosphere 74: 633–641.
Sierra, M. V., N. Gómez, A. V. Marano & M. A. Di Siervi, 2013. Caracterización funcional y estructural del biofilm epipélico en relación al aumento de la urbanización en un arroyo de la Llanura Pampeana (Argentina). Ecología Austral 23(2): 108–118.
Sierra, M. & N. Gomez, 2007. Structural characteristics and oxygen consumption of the epipelic biofilm in three lowland streams exposed to different land uses. Water, Air, and Soil Pollution 186(1–4): 115–127.
Sommaruga, R. & R. Psenner, 1995. Permanent presence of grazing-resistant bacteria in a hypertrophic lake. Applied and Environmental Microbiology 61(9): 3457–3459.
Stevenson, R. J., & Bahls, L. L., 1999. Periphyton Protocols. In: Barbour, MT, J. Gerritsen, BD Snyder, and JB Stribling. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish. EPA/841-B-99-002. US EPA, Office of Water, Washington, DC.
Sweeney, B. W., T. L. Bott, J. K. Jackson, L. A. Kaplan, J. D. Newbold, L. J. Standley, W. C. Hession & R. J. Horwitz, 2004. Riparian deforestation, stream narrowing, and loss of stream ecosystem services. Proceedings of the National Academy of Sciences of the United States of America 101: 14132–14137.
Tagliaferro, M., A. Giorgi, A. Torremorell & R. Albariño, 2019. Urbanisation reduces litter breakdown rates and affects benthic invertebrate structure in Pampean streams. International Review of Hydrobiology 105(1–2): 33–43.
Tambussi, E. A. & Graciano, C., 2010. La fluorescencia modulada de la clorofila. Método del pulso saturante. pp 117–127 en: Técnicas en medición en ecofisiología vegetal: conceptos y procedimientos (No. 581.1). Ediciones INTA. Buenos Aires, Argentina.
Taylor, S. L., S. C. Roberts, C. J. Walsh & B. E. Hatt, 2004. Catchment urbanisation and increased benthic algal biomass in streams: linking mechanisms to management. Freshwater Biology 49(6): 835–851.
Tien, C. J., W. H. Wu, T. L. Chuang & C. S. Chen, 2009. Development of river biofilms on artificial substrates and their potential for biomonitoring wáter quality. Chemosphere 76(9): 1288–1295.
Tlili, A., M. Maréchal, A. Bérard, B. Volat & B. Montuelle, 2011. Enhanced co-tolerance and co-sensitivity from long-term metal exposures of heterotrophic and autotrophic components of fluvial biofilms. Science of the Total Environment 409: 4335–4343.
Valdés, M. E., L. H. M. L. M. Santos, M. C. Rodríguez Castro, A. Giorgi, D. Barceló, S. Rodríguez-Mozaz & M. V. Amé, 2021. Distribution of antibiotics in water, sediments and biofilm in an urban river (Córdoba, Argentina, LA). Environmental Pollution 269: 116–133. https://doi.org/10.1016/j.envpol.2020.116133.
Velázquez, G., 2000. El proceso de urbanización en la Argentina: de la primacía a la fragmentación socio-espacial. Tiempo y Espacio 9–10: 5–22.
Velimirov, B., N. Milosevic, G. G. Kavka, A. H. Farnleitner & A. K. Kirschner, 2011. Development of the bacterial compartment along the Danube River: a continuum despite local influences. Microbial Ecology 61(4): 955–967.
Vilches, C. & A. Giorgi, 2010. Metabolism in a macrophyte-rich stream exposed to flooding. Hydrobiologia 654(1): 57–65.
Vilches, C., A. Giorgi, M. C. Rodriguez Castro & M. A. Casco, 2014. Periphyton responses to non-point pollution in eutrophic-humic environment: an experimental study. International Journal of Environmental Research 8: 523–530.
Walsh, C. J., A. H. Roy, J. W. Feminella, P. D. Cottingham, P. M. Groffman & R. P. Morgan, 2005. The urban stream syndrome: current knowledge and the search for a cure. Journal of the North American Benthological Society 24(3): 706–723.
Young, R., C. Mathaei & C. Towsend, 2008. Organic matter breakdown and ecosystem metabolism: functional indicators for assessing river ecosystem health. Journal of the North American Benthological Society 27(3): 605–625.
Zar, J. H., 1999. Biostatistical Analysis, 5th ed. Prentice Hall, New York: 931.
Acknowledgements
We would like to thank Eduardo Zunino, Sebastian Kravetz, and Marina Tagliaferro for their help in the field. We thank Laura Rigacci for help in the lab. We thank our funders, Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET); Universidad Nacional de Luján (UNLu); and Agencia Nacional de Promoción de la Investigación, el Desarrollo Tecnológico y la Innovación (AGENCIA)—Fondo para la Investigación Científica y Tecnológica (FONCYT).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be as a potential conflict interest.
Additional information
Handling Editor: Stefano Amalfitano
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Gorbarán, R., Vilches, C., Rodríguez Castro, M.C. et al. Structural parameters of biofilm and bacterioplankton are better indicators of urbanization than photosynthetic functional parameters in low-order streams. Hydrobiologia 850, 607–625 (2023). https://doi.org/10.1007/s10750-022-05110-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-022-05110-5