
Abstract
Bolboschoenus planiculmis is a key wetland plant in the semi-arid regions of Songnen Plain, China, providing food and habitat for the endangered Siberian crane. B. planiculmis wetlands are being degraded by water shortages caused by anthropogenic activities and climate changes. However, the vegetation dynamics under hydrological fluctuations have been rarely reported. We conducted a two-year field investigation in B. planiculmis wetlands to determine the vegetation response to water level changes, and a comparative control experiment to examine the restoration effect of supplemental water on degraded community. We found that the community biomass was positively related with water depth under natural conditions, whereas plant diversity and richness were negatively related. The population density, biomass, and root/shoot ratio of B. planiculmis exhibited quadratic relationships with water depth. The optimum ecological threshold of water depth determining B. planiculmis populations was 11.2–36.1 cm. The control experiment demonstrated that water depth at 0–10 cm during the early growing season and 10–25 cm at later growing stages can enhance the dominance of B. planiculmis and help it outcompete other macrophyte species. Understanding the water requirement cycles of B. planiculmis is valuable for guiding water management plans for the rehabilitation of degraded natural wetlands.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Hydrological regimes in natural wetlands have been increasingly disturbed by drainage, channelizing, and reclamation for agricultural development projects (Mitsch & Gosselink, 2015). Disturbed hydrological regimes in combination with climate changes can directly affect wetland vegetation by changing community types, productivity, and plant species composition (Rongoei et al., 2014; Hong & Kim, 2016). Water level in wetlands is a key variable of hydrological regimes that can affect the plant community (Boar, 2006). During the maintenance and restoration of natural wetlands, water level change is the most important driver of plant species diversity (Konar et al., 2013; An et al., 2019). For instance, species richness significantly decreased along soil water content in an alpine wetland ecosystem (Wu et al., 2013). Also, plant species richness of restored plant communities decreased with water level increasing in fresh water wetlands (Wang et al., 2010). The plant composition in passively restored wetlands often dynamically changes with water level fluctuations (De Steven et al., 2010). Therefore, empirical studies on the effects of hydrological fluctuations on the key response traits (including biomass, reproduction, and plant species diversity) in aquatic ecosystems are critical for better quantifying the responses of the plant community and designing proper management measures for disturbed wetlands.
Water depth is a key hydrological regime factor impacting the early growth, seedling survivorship, and subsequent development of wetland plants (Magee & Kentula, 2005; Hussner et al., 2009; Daoud-Bouattour et al., 2014). Understanding species-specific differences in the ecological adaptation of target plants to water depth could help improve the resilience of plant communities. Recently, the effects of water depth fluctuations on individual aquatic plant species have been widely addressed. For example, reduced water levels negatively affect the biomass of Egeria najas Planch., whereas water level recovery is beneficial for the rapid growth of the floating species Eichhornia crassipes (Mart.) Solms (Thomaz et al., 2006). The responses of Phormium tenax J. R. Forst. & G. Forst, Carex secta Boott, and Typha orientalis C. Presl to water depth differ both quantitatively and qualitatively, and are not linear (Sorrell et al., 2012). As water depth increases, Typha domingensis Pers. increases its shoot height, produces fewer but larger ramets, and exhibits reduced flowering (Grace, 1989). Deeper water conditions may enhance the growth of cattails but inhibit the seedling emergence of many wetland species (Kim et al., 2013; Liu et al., 2016). Different plant species have developed various morphological and physiological mechanisms to adapt to hydrological conditions. Therefore, the ecological adaptation of plants and optimum water depths should be fully considered when developing hydrological strategies to restore the population of target species.
In the semi-arid region of the Songnen Plain, China, Bolboschoenus planiculmis (F. Schmidt) T. V. Egorova is the main wetland plant that provides habitat and food for the endangered Siberian crane (Grus leucogeranus Pallas, 1773). Recently, B. planiculmis wetlands in this area have suffered from water shortages and drought caused by ditching and draining for farmland irrigation (An et al., 2018). To cope with this degradation, some management measures, such as the “river–lake connectivity” project, have been implemented to promote the restoration of wetlands (Li et al., 2016). However, the relationships between water level and wetland vegetation are not adequately considered, leading to a lower recovery efficiency and uncertainty of vegetation succession (An et al., 2019). As a result, mesophytes including Suaeda salsa (L.) Pall. and Echinochloa caudata Roshev. and emergent aquatic plants such as Phragmites australis (Cav.) Trin. ex Steud. and Typha angustifolia L. have begun to dominate plant communities in rewetted sites (Wen et al., 2020). Concurrently, the presence and dominance of B. planiculmis in wetlands has decreased (An et al., 2018). The different sensitivities to changes in hydrology and disturbance among individual species can result in variations in plant community composition (De Cauwer & Reheul, 2009; Wang et al., 2016). Thus, a comprehensive and quantitative study on the water requirements and optimal water level for the B. planiculmis community and population would facilitate the proper management and effective restoration of degraded wetlands.
We hypothesized that water depth fluctuations affected the B. planiculmis community and population characteristics under natural condition. We also theorized that a certain water level would meet with the requirements for the establishment of B. planiculmis, and enhance its recruitment and development in degraded wetlands. In this study, we conducted a two-year field investigation and a comparative control experiment. The specific objectives of this study were as follows: (1) to assess the variations in community characteristics and plant species diversity in B. planiculmis wetlands according to water level gradients, (2) to determine the ecological threshold of water depth for B. planiculmis populations, and (3) to examine the restoration effects of different depths of supplemented water on degraded B. planiculmis wetlands. Results of this study can provide the basic data for estimating ecological water requirements of B. planiculmis wetland, and contribute to improving the restoration efficiency of degraded B. planiculmis wetlands in semi-arid regions.
Materials and methods
Research area
The research area is in the Momoge National Natural Reserve (MNNR) (45°42′25″–46°18′0″ N, 123°27′0″–124°04′33″ E) on the Songnen Plain, China. It is an alluvial plain of the Nenjiang tributary, as well as the main stopover site for Siberian cranes (Wang et al., 2013). The climate is semi-arid with a continental monsoon season in the North Temperate Zone. The annual average temperature, precipitation, and average evaporation are 4.2 °C, 392 mm, 1,472 mm, respectively. There are large areas of cultivated fields, scattered marshes, and temporary pools. The main soil types are marsh and meadow soils. With increasing water depth, the marsh wetlands are characterized by wetland plants such as B. planiculmis, Phragmites australis, Carex spp, Scirpus validus Vahl, and other flood-tolerant species. The dominant vegetation in the meadows and wet meadows of this area are mesophytes such as Suaeda salsa and Echinochloa caudata.
Experimental design
Field investigation
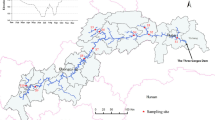
Thirteen B. planiculmis wetland sites in the MNNR were selected (Fig. 1). We conducted a two-year field investigation in these wetland sites. Water depth was measured monthly from May 2016 to September 2017 in each site using a graduated portable staff gauge. The water level was recorded as zero when the water level was at or below the ground surface. At each wetland site, water depth was measured to the nearest 0.1 cm at several locations. The average water level was then calculated. These averages were recorded as the representative monthly water depths for each wetland site.

Locations of B. planiculmis wetland sites in the Momoge National Natural Reserve (MNNR). The sampling sites of the two-year field investigation in 2016 and 2017 are indicated by black triangles labeled S1 through S13
We conducted a two-year vegetation investigation of the 13 B. planiculmis wetland sites in early August of 2016 and 2017. This is the season when the plants are experiencing rapid growth and reproduction, and the water requirement is highest. A sampling transect (50 m long × 10 m wide) was randomly selected for each site, and five quadrats (1 m × 1 m) were randomly selected within each transect. The community characteristics of B. planiculmis were then investigated. In each quadrat, community height was measured as the average height of 10 plants, Canopy coverage of the community was estimated as the surface covered by each species in the vertical projection and expressed as a percentage of the sampling surface. All individual plants were classified into species. Plant density per species was calculated from the number of individuals within each quadrat. All green plants from the entire quadrat were cut at ground level and collected. To determine the belowground biomass, one sub-quadrat (30 cm × 30 cm) was set in each quadrat and excavated to a depth of 30 cm. The excavated material was dried, crushed, and passed through a 2-mm sieve to isolate the roots (including root tubers, rhizomes, and fibrous roots) within each sample. Fine roots that remained in the soil samples were further isolated by spreading the samples in shallow trays and overfilling the trays with distilled water. The outflow from the trays was allowed to pass through a 0.5-mm sieve. All aboveground and belowground parts were oven-dried at 65°C until their weights remained constant. The final dried weights were to determine aboveground and belowground biomasses. The total biomass was the sum of aboveground and belowground biomasses of the living plants.
To determine the population characteristics of B. planiculmis, another 30 × 30 cm sub-quadrat was randomly selected in each quadrat. Next, all plants with roots were excavated to a depth of 30 cm and all B. planiculmis plants were selected. The population density of B. planiculmis was determined by calculating the number of individuals in each sub-quadrat. Subsequently, all plant samples were taken to the laboratory and the aboveground and belowground parts were separated. The belowground parts, including root tubers, rhizomes, and fibrous roots, were carefully rinsed with tap water, and wiped with filter paper. Afterward, the root tubers were separated and counted. The aboveground and belowground parts of B. planiculmis plants were oven-dried at 65°C for 72 h to achieve constant weights. The total B. planiculmis biomass was the sum of aboveground and belowground biomasses. The root/shoot ratio was calculated by dividing the belowground biomass by aboveground biomass.
Control experiment
In 2018, a control experiment was conducted at the experiment site (45°53′27″ N, 123°39′26″ E) near the degraded B. planiculmis wetlands in MNNR. On April 28, 2018, 15 plots (100 cm long × 100 cm wide × 30 cm deep) in the degraded B. planiculmis wetlands were randomly selected, excavated, and moved into the excavations of same dimensions in the experimental site. Before replacement of these plots, the excavations were covered with plastic sheets to prevent water loss and retain the desired water levels. Afterward, a transparent rain shelter was established above the experimental plots. These plots were watered based on five water level treatments (W1–W5): soil water content of 50% ± 5%, and above ground water levels with depths of 0, 10, 25, and 40 cm, respectively. Each treatment was replicated three times. A water level corresponding to a soil water content of 50% ± 5% matched the natural water condition in the degraded B. planiculmis wetlands. Any evaporated water was replaced daily to meet the experimental requirements, and the lowest water level was measured using a moisture meter before and after supplemental water was added to the plots.
The control experiment began on May 1, 2018, and ended on September 1, 2018. The vegetation was analyzed on June 1, July 1, August 1, and September 1. At each sampling date, one quadrat (20 cm long × 20 cm wide) was randomly selected within each plot. The plant height, density, aboveground biomass, and belowground biomass of B. planiculmis and its associated plants were measured using the above-mentioned methods. In addition, the newborn root tubers were separated from the stock plants, counted, and weighted after being oven-dried at 65°C for 72 h.
Data analysis
Plant species diversity indexes were examined based on vegetation data from the field investigation. The Shannon–Wiener diversity index (H), richness index (R), and evenness index (E) were calculated as follows:
where Pi is the density proportion of species i, S is the total number of species observed in the B. planiculmis community, and N is the sum of total species individuals.
The relationships between the plant biological indicators and environmental factors under natural conditions generally agrees with the Gauss equations. Therefore, the trend describing changes in plant biological indicators with the gradient of environmental factors can be fitted by the Gauss model equation (Gauch et al., 1974; Terbraak & Looman, 1986; Luan et al., 2013; Xu et al., 2015), as follows:
where y is a biological indicator of plant species; c is the maximum of y; x is the value of environment factor; u is the optimum ecological threshold of the species to environment factor; and t is the species tolerance, which is an index for describing the ecological threshold. The optimum ecological threshold of species to environmental factors is [u − t, u + t]. Prior to obtaining these parameters, a natural logarithm was calculated for both sides of the Gauss model equation to fit the univariate quadratic equation.
Analysis of variance (ANOVA) was used to assess differences in the vegetation characteristics of various wetland sites in MNNR at the P ≤ 0.05 levels. Linear regressions were performed to identify the relationships between water depth and community characteristics. To assess the influence of water depth on B. planiculmis development, we tested the relationships between population characteristics and water depth using non-linear regressions. The coefficient of determination (R2) of regressions at the P ≤ 0.05 level was considered significant. One-way ANOVA was conducted to analyze the differences in vegetation characteristics and biomass under water supplemented at different depths. Multiple comparisons were performed via the least significant difference (LSD) at the P ≤ 0.05 level. All statistical analyses were performed using SPSS (version 16.0; SPSS, Chicago, IL, USA).
Results
Plant community characteristics of B. planiculmis wetlands with different water depths under natural condition
The results showed significant differences in plant community characteristics among the wetland sites in 2016 and 2017 (P < 0.05; Table 1). The average plant height of B. planiculmis communities in 2016 was slightly lower than that in 2017. The average plant coverage changed slightly, while maximum values decreased from 2016 to 2017. The density of B. planiculmis communities in 2017 was higher than that in 2016. The averages of aboveground, belowground, and total biomasses decreased from one year to the next. However, maximum values of them increased from 2016 to 2017. The Shannon–Wiener index increased slightly from 2016 to 2017, but the richness index remained constant at 0.9. In the two investigations, the evenness index of B. planiculmis communities ranged from 0.5 to 0.9 in 2016, and from 0.6 to 0.8 in 2017.
Population characteristics of B. planiculmis with different water depths under natural conditions
The results from the two investigations showed that the mean values for the characteristics of B. planiculmis populations varied substantially among different wetland sites (P < 0.05; Table 2). Average plant height decreased only slightly from 2016 to 2017. The minimum measured coverage percentage increased from 2016 to 2017, while the maximum coverage decreased. The population density and mean tuber density of the plants increased from one year to the next. The biomass of the plants decreased from 2016 to 2017. The minimum measured tuber biomass decreased from 2016 to 2017, though the maximum and average tuber biomass increased. The average root/shoot ratio remained unchanged (1.2) from one year to the next.
Relationships between water depth and plant community characteristics of B. planiculmis wetlands
There was no relationship between plant height, coverage, or density and water depth (P > 0.05, Fig. 2A–C). However, the biomass of B. planiculmis communities increased with increasing water depth. The aboveground biomass, belowground biomass, and total biomass of the communities were positively related with water depth (P < 0.05; Fig. 2D–F), and the coefficients were 0.190, 0.199, and 0.245, respectively. The regressions showed that the Shannon–Wiener index and richness index were negatively related with water depth (P < 0.05, Fig. 3A, B), with coefficients of 0.281 and 0.378, respectively. There was no significant relationship between the evenness index of B. planiculmis community and water depth (P > 0.05, Fig. 3C).

Linear regressions between water depth and community characteristics of B. planiculmis wetlands in MNNR. A plant height, B coverage, C plant density, D aboveground biomass, E belowground biomass, and F total biomass. Values (mean ± SE, n = 5) are obtained from the two-year field investigation in 2016 and 2017. Vertical lines indicate error bars. Coefficient of determination (R2) and significance (P) of linear regression are indicated. Ns indicates no significant relationship

Linear regressions between water depth and plant diversity in B. planiculmis wetlands in MNNR. A Shannon–Wiener index, B richness index, and C evenness index. Values (mean ± SE, n = 5) are obtained from the two-year field investigation in 2016 and 2017. Vertical lines indicate error bars. Coefficient of determination (R2) and significance (P) of linear regression are indicated. Ns indicates no significant relationship
Relationships between water depth and population characteristics of B. planiculmis
The results show no relationship between water depth and the height, coverage, and tuber density of B. planiculmis populations (P > 0.05, Fig. 4A, C, G). However, the significant non-linear regressions demonstrate the relationships between water depth and the population density, aboveground biomass, belowground biomass, total biomass, tuber biomass, and root/shoot ratio (P < 0.05, Fig. 4B, D–F, H, I). These univariate quadratic equations agreed with the Gauss model, using the following equations:

Non-linear regressions between water depth and population characteristics of B. planiculmis in MNNR. A plant height, B population density, C coverage, D aboveground biomass, E belowground biomass, F total biomass, G tuber density, H tuber biomass, and I root/shoot ratio. Values (mean, n = 5) are obtained from the two-year field investigation in 2016 and 2017. Coefficient of determination (R2) and significance (P) of non-linear regression are indicated. Ns indicates no significant relationship
respectively. Based on the above population characteristics, the optimum water depths for B. planiculmis populations were 23.0, 28.0, 23.2, 23.8, 27.0, and 20.8 cm, respectively. The optimum ecological threshold ranges were [0.6 cm, 45.4 cm], [5.6 cm, 50.4 cm], [10.3 cm, 36.1 cm], [8.0 cm, 39.6 cm], [11.2 cm, 42.8 cm], and [5.0 cm, 36.6 cm], respectively. By calculating the intersection of the above threshold ranges, the optimum ecological threshold determining the B. planiculmis population was [11.2 cm, 36.1 cm].
Restoration effects of supplemented water on degraded B. planiculmis wetlands
The different water level treatments of the experimental plots had a significant effect on the degraded B. planiculmis community across the growing season (Table 3). At the early growth stage (June 1), plant density and growth were promoted under treatment W2 when compared with those under natural condition (W1) and greater water depths (W3–W5). The density and biomass of plants under W1 and W5 water levels were relatively lower than with other water depths. The July 1 results showed that the W3 level increased the density, belowground biomass and total biomass, although plant height was greatest under the W4 water level. Like the tendency observed on July 1, plant density, belowground biomass, and total biomass were generally increased by the W3 water level on August 1, whereas plant height was higher under W4 and W5 levels. From July 1 to September 1 (late growing stage), supplemental water treatments W3 and W4 increased biomass production, with no significant difference between the results from these two water levels.
The population density of B. planiculmis on June 1 under the W2 level (1180.0 plant m−2) was the greatest among the five water levels for that date (Fig. 5A). However, with the progression of the growth stages, the greatest population density at the late growth stage (September 1) was under W3 level. Aboveground biomass was greatest (134.9 g m−2) under the W2 condition at the early growth stage but in later growth stages (July 1, August 1, and September 1) the W4 levels produced greater results (Fig. 5B). The W3 water level significantly improved belowground biomass and total biomass at the early growth stage (June 1), whereas both the W3 and W4 water levels exhibited the greatest improvement in belowground and total biomass during later growth stages (July 1 through September 1, see Fig. 5C, D). For the reproduction of B. planiculmis, the newborn tuber density under the W2 water level was greatest (532.0 no. m−2) among the five water levels on June 1 (Fig. 5E). On July 1 the W3 water level showed a slightly greater density, but the density progressively increased under W2 through the later stages (August 1 and September 1) and the W2 water level again produced the greatest newborn tuber densities. Similarly, the biomass of newborn tubers was higher (18.5 g m−2) under W2 level on June 1 (Fig. 5F). Then, the W3 water levels achieved higher values (176.4 g m−2 and 748.3 g m−2) than the other water levels for July1 and August 1, respectively. There was no significant difference in newborn tubers among W2, W3, and W4 water levels on September 1.

Effects of supplementing water at different levels on population density (A), aboveground biomass (B), belowground biomass (C), total biomass (D), newborn tuber density (E), and newborn tuber biomass (F) of B. planiculmis across a growing season in the control experiment. W1, W2, W3, W4, and W5 indicate a soil water content of 50% ± 5%, and 0, 10, 25, and 40 cm water depths above the soil surface. Values are mean ± SE (n = 3). Vertical lines indicate error bars. Different letters indicate the significant differences among the five water levels at P ≤ 0.05
In addition to B. planiculmis, water supplement affected the growth of other mesophyte and macrophyte species (Fig. 6). In the early growing season (June 1 and July 1), the density and biomass of associated plants under W2 water levels were higher than those under other levels. At later growth stages (August 1 and September 1), W1 and W2 water levels generally increased the density and biomass. In particular, the plant densities under W1 and W2 water levels on September 1 (517.0 and 468.0 plant m−2, respectively) were much greater than those under other water levels (Fig. 6A). By the late growing season, the aboveground biomass of associated plants under the W1 water level (351.6 g m−2) was greater than under the other water levels (Fig. 6B). Belowground biomass was relatively higher under W2 and W3 water levels across the growing season (Fig. 6C). Total biomass was lower under W3, W4, and W5 water levels at the late growth stage (Fig. 6D).

Effects of supplementing water at different levels on plant density (A), aboveground biomass (B), belowground biomass (C), and total biomass (D) of associated plants of B. planiculmis across the growing season in the control experiment. W1, W2, W3, W4, and W5 indicate a soil water content of 50% ± 5%, and 0, 10, 25, and 40 cm water depths above the soil surface. Values are mean ± SE (n = 3). Vertical lines indicate error bars. Different letters indicate the significant differences among the five water levels at P ≤ 0.05
Discussion
In aquatic ecosystems, water depth commonly fluctuates with different frequencies and amplitudes depending on precipitation or human activities (Deegan et al., 2007; Rongoei et al., 2014). Many studies have shown that water depth has a remarkable effect on biomass production as plants are becoming established (Dai et al., 2020; Overbeek et al., 2020). Deeper water inhibits plant growth by decreasing the effectiveness of light (Strand & Weisner, 2001; Zhu et al., 2012) and by affecting pathways of nutrient cycling (Pinay et al., 2002; Yuckin & Rooney, 2019). Unlike previous results, the biomass of B. planiculmis communities increased with increasing the water depth in our study. A likely reason for this is that B. planiculmis communities commonly occur in habitats with shallow water, such as mudflats around lakes and water bodies in river floodplains. In this study, we conducted a two-year field investigation in the wetlands in MNNR having water depth of 0–40 cm. Although the wetland vegetative community productivity increased with increasing water depth, the dominance of B. planiculmis decreased. Water levels up to approximately 40 cm promoted the invasion of other macrophyte species, and emergent vegetation dominated. Therefore, greater water depths are unfavorable to the maintenance of B. planiculmis community.
Results of the field investigation show that water depth not only influences the productivity of the B. planiculmis community, but also alters plant species richness and diversity in natural condition. In a previous pot experiment, the lowest species richness and biomass results were produced under continuous flooding conditions, and the highest species richness and biomass results were recorded under conditions of no flooding (Casanova & Brock, 2000). The negative relationships between water depth and plant species richness shown by these experimental results agree with the results of this study. As water depth increased, emergents such as Phragmites australis and Typha orientalis became the dominant species. Higher water levels resulted in lower species diversity because the greater water depth led to the disappearance of mesophytes and overgrowth of aquatic macrophytes. Rapid growth and flood tolerance may enhance the competitive ability of macrophytes and thereby decrease plant diversity. These explanations are supported by earlier studies that showed species richness of plant communities decreased with the improvement of hydrological conditions in a freshwater wetland ecosystem (Wang et al. 2010) and an alpine wetland ecosystem (Wu et al., 2013). Conversely, at lower water depths, wetland and meadow plants such as B. planiculmis and S. salsa became the dominant species. A former field survey found that the meadow communities with lower water levels were composed of a mixture of grasses and forbs, with the greatest number of species (Dwire et al., 2006). Similarly, plant species diversity increased along the hydrological gradient from permanently inundated zones to the seasonally inundated zones in Nyando wetlands (Rongoei et al., 2014). Therefore, water level changes and hydrological gradients act as the determinants for plant diversity in B. planiculmis communities.
The application of ecological threshold theory quantifies the changing trend of plant ecological characteristics with environmental factors. This provides a theoretical basis for understanding the sensitivity of biology to environmental stress, for screening key environmental factors, and for the ecological restoration of wetlands. The biomass of B. planiculmis increased initially with depth but then decreased rapidly, indicating that there was a threshold of water depth for facilitating population growth. The Gauss model further demonstrated that the optimum water depth for B. planiculmis growth and reproduction was 11.2–36.1 cm. This result is consistent with existing reports regarding the non-linear regressions between the growth performances of other wetland plants and water depths (Sorrell et al., 2012). It was also reported that shallow water depths (i.e., 20–30 cm) usually favor the dominance of Typha species in wetlands (Newman et al., 1996), and high-water levels were not conducive to biomass accumulation in Typha domingensis (Chen et al., 2010). According to the physiological and ecological responses of Phragmites australis to water depth gradients in the Momoge wetland, the optimum water level of P. australis is 35–45 cm (Deng et al., 2012), which is likely because the relative plant height of P. australis is greater than that of B. planiculmis and exhibits greater adaptability to deeper water. Therefore, water depth had a significant impact on the growth of establishing plants, and the application of an ecological threshold could provide theoretical guidance for wetland vegetation restoration.
In our study, the 0 cm deep water condition produced higher plant density, biomass, and reproduction of B. planiculmis at the early growth stage, whereas these characteristics were all reduced during dry and standing water conditions. As the growth stage advanced, B. planiculmis exhibited vigorous growth at water depths of 10 cm and 25 cm, and then reduced growth with greater water depths, indicating a strategy for avoiding excessive submergence. This finding was supported by a report that indicated Juncus ingens N. A. Wakef. seedlings initially submerged in shallow water continued to grow well at a water depth of 20 cm (Vivian et al., 2014). In an earlier field experiment, water depths > 10 cm limited the growth of mesophytes, possibly because of restricted light availability (Bornette & Puijalon, 2011) or competitive exclusion by aquatic macrophyte species in deeper standing water (Fleming & Dibble, 2015). The results of our study further indicate that B. planiculmis plants produced more newborn tubers under 0 cm and 10 cm deep water conditions, producing fewer but larger tubers as the water depth increased to 40 cm. This agrees with a previous study where propagation characteristics were decreased under complete submergence conditions (Grace, 1989; Carter et al., 2018). This can be also explained by the fact that emergent vegetation may allocate more resources to aboveground parts in deeper water (Duke et al., 2015). Thus, water depth changes appear to be critical for plant growth, biomass allocation, and vegetative propagation. Flooding tolerance and adaptation in wetland plants should be fully considered when supplemental water is applied to restore target plants.
It has been well demonstrated that the growth characteristics of plants, including growth form, shoot and root biomass, and reproductive output (Zhang et al., 2020), generally reflect the competitiveness of plants (Cronk & Fennessy, 2001; Byun et al., 2017). Among many response traits, biomass is a key trait that is closely correlated with competitive ability (Byun et al., 2017). In our study, B. planiculmis populations gained considerable biomass and reproduction under 10 cm and 25 cm water depths, giving them a greater ability to compete for natural resources with associated plants. Expectedly, the establishment and development of associated mesophytes was negatively affected at these two water depths and facilitated at a low water level (soil water content of 50% ± 5%). In a competition experiment, Zizania latifolia (Griseb.) Hance ex F.Muell. appeared to grow rapidly and produce considerable biomass, and ultimately enhance its dominance under standing water conditions by propagating seedlings and by rapid early growth (Hong et al., 2014). This may explain our results that show B. planiculmis outcompeted the associated plants by increasing its density under a 0 cm deep water level at the early growth stage, and by increasing B. planiculmis biomass under optimal water depths of 10 cm and 25 cm at later growth stages. Therefore, the growth response of plants to water depth verifies the principle that the B. planiculmis community is initially assembled by flooding tolerance in natural conditions with shallow water and by competition in deep water.
According to the water requirements of different species, the manipulation of flooding conditions can be used as an effective measure for plant community assembly and maintenance of target species (Tanentzap et al., 2014). The results of our study also suggest that water depth regulation can substantively restore B. planiculmis populations and manage undesirable species. Begum et al. (2006) obtained similar results, whereby more Fimbristylis miliacea (L.) Vahl seedlings survived delayed flooding than survived earlier flooding. Thus, delaying flooding until the end of the growing season would effectively improve the control of undesired species if flooding water depths were optimum for B. planiculmis populations. The expansion of associated species is often related to changes in the flooding environment. Reducing the wet-dry cycles in natural wetlands can greatly enhance the encroachment of undesired species that are unable to adapt to prolonged and deep flooding environments (Tanentzap et al., 2014; Liu et al., 2016). Manipulation of water depths based on life cycle requirements of target plants can be used for the development of proper practices for restoring a B. planiculmis community and maintaining its dominance in the environment.
Conclusion
Results of the present study show that the community characteristics in B. planiculmis wetlands varied with water depth. In particular, the biomass and plant species diversity exhibited positive and negative linear regressions with the water depth gradient, respectively. The B. planiculmis density, aboveground biomass, belowground biomass, total biomass, root tuber biomass, and root/shoot ratio had a quadratic relationship with water level. According to the Gauss model, the optimum ecological threshold range of water depth was 11.2–36.1 cm under natural conditions.
For B. planiculmis restoration, population density and growth were increased under the water supplemented at 0–10 cm depths in the early growth stage, and subsequent growth was generally facilitated at water depths of 10–25 cm. Simultaneously, the water depths of 0–10 cm favored the reproduction of B. planiculmis across the growth season. In addition, a water supplement with optimal depths for B. planiculmis restoration can effectively decrease the presence of mesophytes and enhance its competitive ability against the associated macrophytes.
During the process of ecological restoration, a balance between water requirements and water supplement is essential for restoring and maintaining the stability of B. planiculmis communities. In particular, the water requirements of vegetation growth and reproduction should be fully considered across the entire life cycle to optimize the water supplement strategy and improve ecological function of degraded wetlands in semi-arid regions.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
An, Y., Y. Gao & S. Z. Tong, 2018. Emergence and growth performance of Bolboschoenus planiculmis varied in response to water level and soil planting depth: implications for wetland restoration using tuber transplantation. Aquatic Botany 148: 10–14.
An, Y., Y. Gao, X. H. Liu & S. Z. Tong, 2019. Interactions of soil moisture and plant community properties in meadows restored from abandoned farmlands on the Sanjiang Plain, China. Community Ecology 20(1): 20–27.
Begum, M., A. S. Juraimi, R. Amartalingam, A. Bin Man & S. O. B. Rastans, 2006. The effects of sowing depth and flooding on the emergence, survival, and growth of Fimbristylis miliacea (L.) Vahl. Weed Biology and Management 6(3): 157–164.
Boar, R. R., 2006. Responses of a fringing Cyperus papyrus L. swamp to changes in water level. Aquatic Botany 84(2): 85–92.
Bornette, G. & S. Puijalon, 2011. Response of aquatic plants to abiotic factors: a review. Aquatic Sciences 73(1): 1–14.
Byun, C., J. M. Nam & J. G. Kim, 2017. Effects of flooding regime on wetland plant growth and species dominance in a mesocosm experiment. Plant Ecology 218(5): 517–527.
Carter, C., J. D. Madsen & G. N. Ervin, 2018. Effects of initial propagule size and water depth on Butomus umbellatus L. growth and vegetative propagation. Aquatic Botany 150: 27–32.
Casanova, M. T. & M. A. Brock, 2000. How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecology 147(2): 237–250.
Chen, H. J., M. F. Zamorano & D. Ivanoff, 2010. Effect of flooding depth on growth, biomass, photosynthesis, and chlorophyll fluorescence of Typha domingensis. Wetlands 30(5): 957–965.
Cronk, J. K. & M. S. Fennessy, 2001. Wetland Plants: Biology and Ecology. Lewis Publishers, Boca Raton.
Dai, X., Z. B. Yu, G. S. Yang & R. R. Wan, 2020. Role of flooding patterns in the biomass production of vegetation in a typical herbaceous wetland, Poyang Lake wetland, China. Frontiers in Plant Science 11: 1547.
Daoud-Bouattour, A., M. Bottollier-Curtet, H. F. -B. Jamaa, Z. Ghrabi-Gammar, S. B. Saad-Limam, L. Rhazi & S. D. Muller, 2014. Effects of hydrology on recruitment of Pilularia minuta Durieu (Marsileaceae), an endangered plant of Mediterranean temporary pools. Aquatic Botany 112: 76–83.
De Cauwer, B. & D. Reheul, 2009. Impact of land use on vegetation composition, diversity and potentially invasive, nitrophilous clonal species in a wetland region in Flanders. Agronomy for Sustainable Development 29(2): 277–285.
De Steven, D., R. R. Sharitz & C. D. Barton, 2010. Ecological outcomes and evaluation of success in passively restored Southeastern depressional wetlands. Wetlands 30(6): 1129–1140.
Deegan, B. M., S. D. White & G. G. Ganf, 2007. The influence of water level fluctuations on the growth of four emergent macrophyte species. Aquatic Botany 86(4): 309–315.
Deng, C. N., G. X. Zhang & X. L. Pan, 2012. Eco-physiological responses of Phragmites australis to different water depth in Momoge Wetland. Ecologic Science 31(4): 352–356. (In Chinese)
Duke, S. T., S. N. Francoeur & K. E. Judd, 2015. Effects of Phragmites australis invasion on carbon dynamics in a freshwater marsh. Wetlands 35(2): 311–321.
Dwire, K. A., J. B. Kauffman & J. E. Baham, 2006. Plant species distribution in relation to water-table depth and soil redox potential in Montane riparian meadows. Wetlands 26(1): 131–146.
Fleming, J. P. & E. D. Dibble, 2015. Ecological mechanisms of invasion success in aquatic macrophytes. Hydrobiologia 746(1): 23–37.
Gauch, H. G., G. B. Chase & R. H. Whittaker, 1974. Ordination of vegetation samples by Gaussian species distributions. Ecology 55(6): 1382–1390.
Grace, J. B., 1989. Effects of water depth on Typha latifolia and Typha domingensis. American Journal of Botany 76(5): 762–768.
Hong, M. G. & J. G. Kim, 2016. Effects of initial density, nutrient, and water level regime on the seedling survival and growth of Typha orientalis Presl. Journal of Plant Biology 59(4): 369–376.
Hong, M. G., C. Y. Son & J. G. Kim, 2014. Effects of interspecific competition on the growth and competitiveness of five emergent macrophytes in a constructed lentic wetland. Paddy and Water Environment 12: S193–S202.
Hussner, A., C. Meyer & J. Busch, 2009. The influence of water level and nutrient availability on growth and root system development of Myriophyllum aquaticum. Weed Research 49: 73–80.
Kim, D. H., H. T. Kim & J. G. Kim, 2013. Effects of water level and soil type on the survival and growth of Persicaria thunbergii during early growth stages. Ecological Engineering 61: 90–93.
Konar, M., M. J. Todd, R. Muneepeerakul, A. Rinaldo & I. Rodriguez-Iturbe, 2013. Hydrology as a driver of biodiversity: Controls on carrying capacity, niche formation, and dispersal. Advances in Water Resources 51: 317–325.
Li, X. N., J. Q. Li, X. Fang, Y. W. Gong & W. L. Wang, 2016. Case studies of the sponge city program in China. World Environmental and Water Resources Congress: 295–308.
Liu, B., M. Jiang, S. Z. Tong, W. G. Zhang, H. T. Wu, Y. Liu & X. G. Lu, 2016. Differential flooding impacts on Echinochloa caudata and Scirpus planiculmis: implications for weed control in wetlands. Wetlands 36(5): 979–984.
Luan, Z. Q., Z. X. Wang, D. D. Yan, G. H. Liu & Y. Y. Xu, 2013. The ecological response of Carex lasiocarpa community in the riparian wetlands to the environmental gradient of water depth in Sanjiang Plain, Northeast China. Scientific World Journal 402067.
Magee, T. K. & M. E. Kentula, 2005. Response of wetland plant species to hydrologic conditions. Wetlands Ecology and Management 13: 163–181.
Mitsch, W. J. & J. G. Gosselink, 2015. Wetlands. Wiley, New Jersey.
Newman S, J. B. Grace & J. W. Koebel, 1996. Effects of nutrients and hydroperiod on Typha, Cladium, and Eleocharis: implications for everglades restoration. Ecological Applications 6(3): 774-783.
Overbeek, C. C., S. F. Harpenslager, J. P. van Zuidam, E. E. van Loon, L. P. M. Lamers, M. B. Soons, W. Admiraal, J. T. A. Verhoeven, A. J. P. Smolders, J. G. M. Roelofs & H. G. van der Geest, 2020. Drivers of vegetation development, biomass production and the initiation of peat formation in a newly constructed wetland. Ecosystems, 23(5): 1019–1036.
Pinay, G., J. C. Clement & R. J. Naiman, 2002. Basic principles and ecological consequences of changing water regimes on nitrogen cycling in fluvial systems. Environmental Management 30(4): 481–491.
Rongoei, P. J. K., J. Kipkemboi, S. T. Kariuki & A. A. van Dam, 2014. Effects of water depth and livelihood activities on plant species composition and diversity in Nyando floodplain wetland, Kenya. Wetlands Ecology and Management 22(2): 177–189.
Sorrell, B. K., C. C. Tanner & H. Brix, 2012. Regression analysis of growth responses to water depth in three wetland plant species. Aob Plants pls043.
Strand, J. A. & S. E. B. Weisner, 2001. Morphological plastic responses to water depth and wave exposure in an aquatic plant (Myriophyllum spicatum). Journal of Ecology 89(2): 166–175.
Tanentzap, A. J., W. G. Lee, A. Monks, K. Ladley, P. N. Johnson, G. M. Rogers, J. M. Comrie, D. A. Clarke & E. Hayman, 2014. Identifying pathways for managing multiple disturbances to limit plant invasions. Journal of Applied Ecology 51: 1015–1023.
Terbraak, C. J. F. & C. W. N. Looman, 1986. Weighted averaging, logistic regression and the Gaussian response model. Vegetatio 65(1): 3–11.
Thomaz, S. M., T. A. Pagioro, L. M. Bini & K. J. Murphy, 2006. Effect of reservoir drawdown on biomass of three species of aquatic macrophytes in a large sub-tropical reservoir (Itaipu, Brazil). Hydrobiologia 570: 53–59.
Vivian, L. M., R. C. Godfree, M. J. Colloff, C. E. Mayence & D. J. Marshall, 2014. Wetland plant growth under contrasting water regimes associated with river regulation and drought: implications for environmental water management. Plant Ecology 215(9): 997–1011.
Wang, L., C. C. Song, J. M. Hu & T. Yang, 2010. Response of regeneration diversity of Carex Lasiocarpa community to different water levels in Sanjiang Plain, China. Chinese Geographical Science 20(1): 37–42.
Wang, Y., J. Feng, Q. X. Lin, X. G. Lyu, X. Y. Wang & G. P. Wang, 2013. Effects of crude oil contamination on soil physical and chemical properties in Momoge wetland of China. Chinese Geographical Science 23: 708–715.
Wang, P., Q. Zhang, Y. S. Xu & F. H. Yu, 2016. Effects of water level fluctuation on the growth of submerged macrophyte communities. Flora 223: 83–89.
Wen, D., Y. M. Hu, Z. P. Xiong, Y. Chang, Y. H. Li, Y. Wang, M. Liu & J. H. Zhu, 2020. Potential suitable habitat distribution and conservation strategy for the Siberian Crane (Grus leucogeranus) at spring stopover sites in Northeastern China. Polish Journal of Environmental Studies 29(5): 3375–3384.
Wu, G. L., G. H. Ren, D. Wang, Z. H. Shi & D. Warrington, 2013. Above- and below-ground response to soil water change in an alpine wetland ecosystem on the Qinghai-Tibetan Plateau, China. Journal of Hydrology 476: 120–127.
Xu, X. L., Q. Zhang, Z. Q. Tan, Y. L. Li & X. L. Wang, 2015. Effects of water-table depth and soil moisture on plant biomass, diversity, and distribution at a seasonally flooded wetland of Poyang Lake, China. Chinese Geographical Science 25(6): 739–756.
Yuckin, S. & R. Rooney, 2019. Significant increase in nutrient stocks following Phragmites australis invasion of freshwater meadow marsh but not of cattail marsh. Frontiers in Environmental Science 7: 112.
Zhang, X. B., K. D. Guo, C. Lu, R. M. Awais, Y. F. Jia, L. Q. Zhong, P. Z. Liu, R. Dong, D. Liu, W. K. Zeng, G. C. Lei & L. Wen, 2020. Effects of origin and water depth on morphology and reproductive modes of the submerged plant Vallisneria natans. Global Ecology and Conservation, 24: e01330.
Zhu, G. R., W. Li, M. Zhang, L. Y. Ni & S. R. Wang, 2012. Adaptation of submerged macrophytes to both water depth and flood intensity as revealed by their mechanical resistance. Hydrobiologia 696(1): 77–93.
Funding
This work was financially supported by the National Key Research and Development Program of China (2019YFA0607101), the Strategic Priority Research Program of the Chinese Academy of China (XDA23060402), the National Natural Science Foundation of China (41871102; U19A2042; 41771108), and the Science and Technology Development Program of Jilin Province of China (20180201010SF; 20200201016JC).
Author information
Authors and Affiliations
Contributions
YA and TS: Writing and Editing; ST: Data analysis; YZ and BL: Sample collection. All authors contributed to the writing to additions and comments to the text.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interests.
Informed consent
The author consent to the publication of this article.
Additional information
Handling editor: Chris Joyce
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
An, Y., Song, T., Zhang, Y. et al. Optimum water depth for restoration of Bolboschoenus planiculmis in wetlands in semi-arid regions. Hydrobiologia 849, 13–28 (2022). https://doi.org/10.1007/s10750-021-04704-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-021-04704-9