
Abstract
Macrophytes are an essential biological element of freshwater aquatic ecosystems and are well known to reflect prevailing ecological conditions in rivers. Their use as bioindicators of nutrient status in river systems is widespread yet their reliability is hotly debated. The aim of this investigation was to assess whether macrophytes are reliable indicators of nutrient levels or better applied as indicators of other non-nutrient environmental conditions. The importance of two water-column nutrients; nitrates (N) and orthophosphates (P) were assessed in terms of their influence on macrophyte species richness and diversity in relation to other non-nutrient environmental factors using 395 river plots from Ireland. Then, in this context, the efficiency of macrophytes to detect nutrient levels was assessed by selecting two macrophyte-based water quality assessment tools; the Mean Trophic Rank (MTR) and the Predictions And Classification System for river macrophytes (LEAFPACS2). Finally, the ability of phytosociological communities within these rivers to reflect trophic levels was examined using the same two water quality assessment tools. It was shown that water-column nutrients N and P have a minor influence on macrophyte richness and no significant influence on macrophyte diversity and that MTR and LEAFPACS2 were only weakly correlated with N and P levels. It was concluded that macrophytes are sensitive to environmental changes but respond to a combination of ecological factors rather than N or P alone. Therefore, this study suggests that macrophytes are not the most efficient taxon group to apply when assessing trophic changes in isolation of other non-nutrient factors.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Macrophytes are an essential biological element of freshwater aquatic ecosystems and their multiple ecological functions are well-known (Aguiar et al., 2014) and as a result can be useful as biological indicators of prevailing ecological conditions (Baatrup-Pedersen et al., 2017). There is evidence to suggest that macrophytes are both reliable and useful at indicating levels of nutrients (e.g. Szoszkiewicz et al., 2009; Willby et al., 2012), particularly phosphorus (Szoszkiewicz et al., 2014) which is thought to be the principle nutrient that limits growth of aquatic plants (Thiébaut et al., 2002). Indeed, nutrients such as phosphorus can be carried in water-borne sediment in rivers (Mayora et al., 2018) and macrophytes have an important role in the retention of these fine sediments which can be closely related to increased dissolved nutrient availability in river systems (Jones et al., 2012). There is however, evidence that highlights the weaknesses and unreliability of using macrophytes as tools for water quality assessment (Demars et al., 2012; Baatrup-Pedersen et al., 2017). A key issue is that macrophytes respond to a combination of both physical and chemical environmental drivers (Schaumberg et al., 2004).
Despite the ongoing debate, macrophyte indices and metrics continue to be used, improved and developed internationally. Macrophytes are one of the mandatory biological factors of Europe’s Water Framework Directive (WFD) in the assessment and monitoring of ecological status of surface waters (WDF, 2000). Since the establishment of the WFD, many assessment methods have developed based on a variety of criteria such as macrophyte indicator species, vegetation structure, vegetation composition and species diversity.
This investigation aims to assess whether macrophytes and their associated phytosociological communities are indeed reliable indicators of nutrient levels or whether they are better utilised as indicators of other non-nutrient environmental conditions. There are relatively few studies that examine the use of vegetation units such as phytosociological communities or plant groups as bioindicators (Dawson & Szoszchiewicz, 1999; Steffen et al., 2014). This research will contribute to the continuing international debate whether macrophytes are efficient bioindicators of river water nutrient levels and consequently inform the application of macrophyte metrics for monitoring purposes while endeavouring to meet legislative requirements relating to water quality status of rivers. Two macrophyte nutrient assessment tools were selected to facilitate this investigation. Both were chosen because they are presently being evaluated by the Environmental Protection Agency for potential use in the Irish context (Catherine Bradley, Ecological Monitoring and Assessment Unit, EPA August 2018 pers. comm.). One index-based assessment tool is the Mean Trophic Rank (MTR) (Holmes et al., 1999) where selected indicator species (including vascular plants, bryophytes and macroalgae) were given a Species Trophic Rank (STR) value based on expert opinion. These values are used to calculate the MTR of a river site taking cover abundance of these trophic indicator species into account. A low MTR signifies nutrient enrichment (Holmes et al., 1999). The MTR was first developed in the UK to detect changes in trophic levels downstream from waste-water treatment plants (Holmes et al., 1999). Since then this tool was positively evaluated (Dawson et al., 1999, Szoszkiewicz et al., 2002) and has been modified, adapted and used in other European countries such as Poland (Szoszkiewicz et al., 2002) and Slovakia (Raven et al., 2011).
A multi-metric assessment tool that is used in the UK is the Predictions And Classification System for macrophytes (LEAFPACS2) (WFD-UKTAG, 2014), improved and updated from the original LEAFPACS tool (Willby et al., 2010, 2012). It is comparable to MTR in that the survey method is based on that of the MTR and the core trophic indicator species lists are similar, although the LEAFPACS2 species list includes a higher number of trophic indicator species. LEAFPACS2 also differs in that it combines the results of five calculated metrics (based on a deviance from reference conditions) into a final Ecological Quality Ratio (EQR). This EQR is then used to classify the river site into one of 4 ecological quality classes (poor/bad, moderate, good, high). Metrics include those that take alkalinity and slope into account, as these were viewed to have a strong influence on macrophyte community composition and productivity (Willby et al., 2012).
Macrophyte richness in rivers is thought to respond well to changes in environmental conditions (Scarlett & O’Hare, 2006; Szoszkiewicz et al., 2014). The first objective of this investigation was to ascertain the importance of water-column nutrients N (nitrates) and P (orthophosphates) in driving macrophyte richness and diversity in relation to other available non-nutrient chemical and physical factors. The second objective was to assess the efficiency of river macrophytes to detect water-column nutrient levels across a selection of Irish river plots by using the MTR and LEAFPACS2 macrophyte assessment tools. Finally, this study examined the ability of phytosociological communities within these rivers to reflect trophic levels using the same two water quality assessment tools and if any differences could be equated to nutrient levels alone. It should be noted that nutrient data available for this research were from water samples collated from various sources and did not necessarily include sediment-borne nutrient information, hence nutrients N and P are understood to be water-column nutrients.
Methods
Preparation of data
Collation of river vegetation plots and environmental information
A total of 395 river vegetation plots were extracted from the Irish River Macrophyte Database (RMD) (Weekes et al., 2018a). Each plot had the dimensions of 100 m river stretch multiplied by the wetted river channel width (m). Within each plot, macrophytes (vascular plants, bryophytes and common macroalgae, e.g. Cladophora spp.) had been recorded and each species given a % cover abundance value. Associated environmental data included substrate type % cover (bedrock, boulders, cobbles, gravel, sand, fine); slope (m/km); altitude (m a.s.l.); channel width; (m); channel depth (cm). Channel depth was measured as the average water depth within the river plot surveyed. Standard procedure is measuring 3 to 5 depths across the river channel (depending on channel width) at the top(upstream) middle and end (downstream) of each river plot. Water chemistry measurements included water alkalinity (CaCO3 mg/l) and water conductivity (μS/cm). Water nutrient factors included N (mg N/l) and P (mg P/l). Water chemistry results were based on averages from either annual or three-year data recorded bimonthly by the Environmental Protection Agency (EPA) for plots recorded in the Republic of Ireland and the Northern Ireland Environmental Agency (NIEA) for plots within Northern Ireland (N and P levels ranged from 5.8 × 10−3 – 6.652 mg N/l and 3.2 × 10−3–8.845 × 10−1 mg P/l respectively).
Additional environmental data were gathered for each plot from various GIS shapelayers using QGIS (QGIS Development Team version Essen 2.14.7 2016). These included geological bedrock (limestone, sandstone, siliceous, metamorphic) simplified from the GSI (Geological Survey of Ireland) (https://www.gsi.ie/en-ie/data-and-maps/Pages/Bedrock.aspx#); soil type of Northern Ireland from the Agri-Food and Biosciences Institute (AFBI) (Jordan & Higgins, 2009) and of the Republic from the Environmental Protection Agency (EPA) and Teagasc soils data (http://gis.teagasc.ie/soils/about.php). Details of the distances between the macrophyte plots and river source (km) and altitude (m a.s.l.) at source were needed for the LEAFPACS2 indices calculations, these were extracted from the EPA’s Rivers and Lakes data (http://gis.epa.ie/GetData/Download), in combination with NASA’s topography mission data (https://data.gov.ie/dataset/digitalelevation-model-of-ireland-from-nasas-shuttle-radar-topography-mission-srtm).
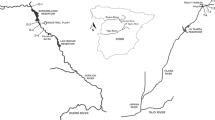
The river plots were from across the island of Ireland (Fig. 1) from a range of wadable rivers with Strahler order ranging from 1 to 7 and an altitude ranging from 1 to 205 m a.s.l.

Map showing the distribution of the 395 river vegetation plots with elevation (darkest shading > 450 m). These plots were extracted from the Irish River Macrophyte Database (RMD)
Calculation of water quality metrics for each plot
MTR calculations
The STR (score ranges from 1 to 10, eutrophic to oligotrophic respectively) and the Species Cover Value (SCV—uses % cover values converted to the 9-point scale) for each indicator species present in a river plot were used to calculate the MTR for each river plot using the formula:
LEAFPACS2 calculations
There are 4 metrics calculated based on what is observed in the field versus what would be expected if reference conditions existed. A breakdown of the calculations, formulae and set of rules used can be seen in detail in WFD-UKTAG (2014). Metrics specific for Northern Ireland rather than U.K. were used for this study that included data from both Northern Ireland and the Republic of Ireland. All calculations were carried out using the downloadable online LEAFPACS2 class calculator and survey metrics calculator (https://www.wfduk.org/resources/rivers-macrophytes). A summary explanation of each metric is described below:
RMNI River Macrophyte Nutrient Index is calculated using the trophic scores of indicator species present versus what would be expected. Scores range between 1 and 10, the lower the score, the less tolerant the species is of eutrophication. The metric takes alkalinity, slope, distance from source and altitude at source into consideration in the calculations.
NTAXA The total number of scoring aquatic taxa recorded in the field versus what would be expected. Slope is taken into consideration in the calculations. This acts as a diversity metric.
NFG Number of functional groups. There are a total 24 functional groups that are considered truly aquatic. Species belonging to these different groups are counted to give the NFG for a river plot versus what would be expected. Slope is taken into consideration in the calculations. This also acts a diversity index.
ALG The % cover of green filamentous algae in a river plot versus a global reference value of 0.05%
A final score is calculated for the surveyed river plot by normalising the 4 metrics and then combining them according to a set of rules into a final EQR (Ecological Quality Ratio).
Hereafter, the final LEAFPACS2 EQRs will be referred to as LPEQR in the text.
Calculation of macrophyte richness and Diversity Index
Macrophyte richness was evaluated as the total number of macrophytes recorded within each plot.
Simpson’s index (Gini coefficient) (Simpson, 1949; Pielou, 1969) was used to calculate the diversity index for each plot. The formula used was:
where: Pk proportion of species k in an infinite population, S total no. of species present, A high value indicates a large number of macrophytes with similar cover abundances (evenness), A low number indicates domination by a few macrophytes.
Statistical analysis
Testing for data auto-correlation
The data were initially tested using the Moran’s I within R version 3.5.0 (R Core Team, 2018) within the package ‘lctools’ (Kalogirou, 2016).
There was some autocorrelation detected, but the Moran’s I value was minimal (MI = 0.19; Z = 10.0; P < 0.05). MI values are interpreted as follows: 1 = clustering, − 1 = dispersion and 0 = random spatial pattern. This small degree of clustering was not considered to result in bias within the data.
Investigation into the importance of N and P in relation to other environmental factors
Negative binomial models were constructed within the R package ‘MASS’ (Venables & Ripley, 2002). These models were used to identify the effects of available environmental variables on macrophyte richness and macrophyte diversity and determine whether nutrients (N and P) had a significant role to play in either macrophyte richness and/or diversity. A measurement of influence of each environmental variable was then calculated using Model Averaging within the R package ‘MuMIn’ (Barton, 2016) to identify those factors that had the greatest influence.
Before proceeding, autocorrelation between variables was tested using the Spearman rank correlation coefficient (ρ) within the R package ‘Hmisc’ (Harrell, 2016). There were two environmental factors that were found to be highly correlated with each other, these were conductivity and alkalinity (ρ = 0.94, P < 0.05) and as a result, conductivity was removed from the negative binomial model analysis described above.
Assessment of the efficiency of macrophytes to detect changes in N and P using MTR and LEAFPACS2
Spearman rank correlations were investigated between MTR and LPEQR against N and P levels and other available chemical water properties and physical factors. The Bonferroni test was used to adjust P values to combat the effects of multiple testing. Spearman ρ > 0.7 was considered as a strong correlation; ρ < 0.7 > 0.5 was considered to be a moderate correlation and ρ < 0.5 > 0.3 was considered weak. Multiple pairwise plots were then constructed (Becker et al., 1988) to visually examine any relationships between the factors.
Investigation into whether phytosociological plant communities can reflect trophic differences using in MTR and LEAFPACS2
Each plot had been assigned to a phytosociological association or sub-association (community) during a previous related study when constructing a National River Vegetation Classification System for Irish Rivers (INRVC) (Weekes et al., 2018b). An unsupervised clustering method, K means (MacQueen, 1967) had been used to find groups within in the data. The number of K cluster centroids had been chosen based on expert opinion and then cluster composition was defined objectively using the K means algorithm. K means analysis was carried out using R software (R Core Team 2015 version 3.2.2) which is integrated within the vegetation software JUICE (Tichý, 2002) which is software specifically designed for editing and analyses of phytosociological data. Full details of the analytical procedure and resulting vegetation classification can be found in Weekes et al. (2018b). The plots in this investigation were distributed between 16 phytosociological associations/subassociations (hereafter referred to as communities), an overview of each community is presented in Table 1.
ANOVA (Analysis of Variance) and Tukey’s post hoc. tests were carried out within the R package “Hmisc” using transformed environmental data to investigate whether there were any significant differences in environmental conditions between the assigned vegetation communities. Similarly, this procedure was used to identify any significant differences in the MTR and LPEQR scores between the communities and then whether they corresponded to any significant differences in nutrient levels. Non-metric multidimensional scaling (NMDS) within the programme PCORD (McCune & Mefford, 2011) was carried out and resulting macrophyte community centroids were plotted to summarise assemblage structure across these environmental gradients.
Boxplots were constructed within the R package ‘ggplot2’ (Wickham, 2009) to visually indicate the direction of any differences between the macrophyte communities and their environmental conditions and MTR/LPEQR scores.
Results
The importance of N and P in relation to other environmental factors
Model averaging of the negative binomial identified the relative importance of all environmental factors (in relation to each other) on macrophyte richness (Table 2). The most influential factors were found to be altitude, slope, geology and alkalinity (> 0.82 (out of a score of 1) relative importance). P also had significant influence but to a lesser extent compared with the other factors (relative importance = 0.75), as did channel width (relative importance = 0.68) and depth (relative importance = 0.67). Levels of N and factors including substrate type, soil type and flow were not found to significantly influence macrophyte richness.
Nutrient levels and other environmental factors had no significant influence on species diversity as explained by the Simpson index (Appendix 1). Species diversity was not found to be significantly affected by any environmental factor.
Assessment of the efficiency of macrophytes to detect changes in N and P using MTR and LEAFPACS2
Pairwise tests showed that there was a moderate correlation between MTR and LPEQR (ρ = 0.51, P < 0.001), a higher correlation would have been expected on the premise that both tools are designed to detect nutrient levels within a river (Fig. 2).

Multiple pairwise comparisons showing the relationship between MTR, LPEQR and available water chemical factors with corresponding Spearman rho ρ and adjusted P values
Water chemistry factors were all at least weakly correlated with each-other (Fig. 2) with moderate correlations found between N and P (ρ = 0.61, P < 0.001) and between N and conductivity (ρ = 0.59, P < 0.001).
MTR was moderately negatively correlated with alkalinity but weakly negatively correlated with P and N levels. The LPEQR was weakly correlated with P but not correlated with other chemical factors (Fig. 2). A weak correlation between MTR and slope (ρ = 0.34, P < 0.001) and a weak correlation between LPEQR and substrate (gravel) (ρ = 0.36, P < 0.001) were found. Pairwise comparisons between nutrient levels and macrophyte richness and diversity show that, although significant, they were only weakly correlated (Fig. 3), i.e. a weak correlation between N and macrophyte richness and diversity (ρ = 0.14, P = 0.0258; ρ = 0.15, P = 0.0192 respectively) and a weak correlation between P and macrophyte richness and diversity (ρ = 0.0.15, P = 0.0168; ρ = 0.15, P = 0.0138 respectively).

Multiple pairwise comparisons showing the relationship between N and P and macrophyte richness and diversity with corresponding Spearman rho ρ and adjusted P values
Differences in environmental factors and MTR and LPEQR scores between the phytosociological plant communities
Significant differences in MTR and LPEQRs values and environmental factors (both physical and chemical) were found between some (not all) phytosociological communities using ANOVA. Tukey’s post hoc test identified which communities were significantly different from each other in terms of these factors, and an NMDS plot summarises the distribution of macrophyte communities across the environmental gradients (Fig. 4). Moss dominated communities are found where there is higher slope and coarser substrate towards the right side of the plot, vascular plants are more dominant in finer substrate and deeper waters towards the left side of the plot. Water chemistry plays as a role in community distribution, but N and P do not seem to have any major influence above non-nutrient factors. MTR appears to have a stronger influence within the plot, however its correlation with alkalinity must be taken into consideration here.

NMDS plot summarising macrophyte community distribution across environmental gradients. The centroids represent each of the 16 macrophyte communities and overlay vectors represent the environmental factors
Significant differences in water chemistry were found; N (NO3) levels (F = 7.63), P (F = 7.40), conductivity (F = 11.17) and alkalinity (F = 10.23) (where P < 0.05, DF = 15) (Fig. 5). MTR and LPEQR were significantly different between some communities (MTR: F = 19.91, P < 0.05, DF = 15; LPEQR: F = 7.19, P < 0.05, DF = 15) (Fig. 6). There were also significant differences in slope, channel depth and width, substrate type and altitude between some of the communities (where P < 0.05, DF = 15) (Fig. 7).

Boxplots of communities in relation to their associated chemical factors: A alkalinity; B nitrates; and C orthophosphates, (conductivity not included as it was highly correlated with alkalinity). Significant differences between these factors and corresponding communities are found in Appendix 2

Boxplots of communities and corresponding for: A MTR and B LEAFPACS2 EQRs. Significant differences between these factors and corresponding communities are found in Appendix 2

Boxplots of macrophyte communities and their associated physical factors: A Slope; B Channel depth; C Channel width; D Fine substrate and E Elevation. Significant differences between these factors and corresponding communities are found in Appendix 2
The significant differences found between the plant communities were mostly due to a combination of environmental factors, a full breakdown of where the significant differences lie is found in Appendix 2. There was rarely a significant difference between communities in terms of MTR and LPEQRs values that corresponded to nutrient levels alone. Exceptions however were found: (1). Between the two bryophyte communities, Leptodictyo riparii-Fissidentetum crassipedis and the Scapanietum undulatae where NO3 levels, MTR and LPEQR values were significantly different. (2). Between two vascular plant communities, Ranunculetum fluitantis and Rorippo-Phalaridetum arundinaceae where P levels, MTR and LPEQR values were significantly different (3). Between Myriophylletum alterniflori and Rorippo-Phalaridetum arundinaceae where NO3 levels, MTR and LPEQR values were significantly different. This equates to just 3 out of the 53 instances (5.6%) where MTR was found to be significantly different and indicating nutrient levels alone. The other instances where MTR was found to be significant corresponded to significant differences in non-nutrient factors, or where no other difference was found (52.8%; 28 instances) and the remaining 41.5% (22 instances) were corresponding to a combination of both nutrient and non-nutrient factors.
Similarly, the above examples equate to just 3 out of the 25 instances (12%) where LPEQRs were significantly different and indicating nutrient levels alone. Some 36% (9 instances) where LPEQRs were found to be significant corresponded to significant differences in non-nutrient factors, or where no other difference was found and the remaining 52% (13 instances) corresponded to a combination of both nutrient and non-nutrient factors.
Discussion
The importance of N and P in relation to other environmental factors
Model averaging of the negative binomial model indicated that although P levels had a significant influence on macrophyte richness within a river stretch, it was altitude, slope, geology, alkalinity and conductivity (being strongly correlated with alkalinity) that had the greatest significant influence. Other physical factors, channel width and depth, also played a significant role. N levels had no significant influence on macrophyte richness. These results would indicate that river macrophyte richness has a greater response to both physical and non-nutrient factors than to water -column nutrient levels such as N and P. This has also been found in other studies (e.g. Demars & Thiébaut, 2008; Szoszkiewicz et al., 2014). Alkalinity has been found to be an important chemical factor in distinguishing bryophyte species richness and community composition (Sekulová et al., 2011) and likewise for vascular plant distribution in rivers at lower elevation (Lewin & Szoszkiewicz, 2012; Baatrup-Pedersen et al., 2017).
No factors in this study were found to significantly influence macrophyte diversity. Macrophyte diversity did not reflect changes in either physical or chemical conditions, nor was an indicator of ecological status. This would suggest that there are changes in species composition rather than species diversity in response to environmental conditions, e.g. the more sensitive species are replaced when there are either physical or chemical changes, rather than a reduction in species diversity. Significant differences in macrophyte diversity have been found in other studies however, relating to anthropogenic pressures (Camargo, 2018), while others point out that biodiversity may be restricted by poor physical conditions rather than water quality (Baatrup-Pedersen & Riis, 1999). There was no evidence in this study that species diversity was affected in this way. Species diversity is not recommended as a reliable indicator of water quality in rivers (Thiébaut et al., 2002; Szoszkiewicz et al., 2017). Instead, diversity loss seems to occur when there is extreme nutrient enrichment over long periods of time (Steffen et al., 2013). It has been suggested that although macrophytes respond primarily to environmental factors other than nutrients, water quality assessment tools that are designed to detect nutrient enrichment are still useful and reliable (Szoszkiewicz et al., 2014). Other studies suggest that assessment tools that target phosphorus are ambiguous due to the stronger response of macrophytes to other factors (Demars & Thiébaut, 2008; Demars et al., 2012; Baatrup-Pedersen et al., 2017).
Assessment of the efficiency of macrophytes to detect changes in N and P through MTR and LEAFPACS2
The sensitivity of MTR was recognised as being more of an indicator of phosphate levels (Dawson et al., 1999) rather than nitrates although Demars and Harper (1998) did find a significant but weak correlation with N levels. A weak significant Spearman rank correlation was found in this present study between MTR and N levels (ρ = − 0.42) while there was no correlation found between N and LPEQR. Both MTR and LPEQR significantly correlated with P, but correlations were also weak (ρ = − 0.46 and ρ = − 0.39 respectively) as has been found in other studies on MTR (Demars & Harper, 1998) and LEAFPACS (Demars et al., 2012). MTR values in this study were more strongly correlated with alkalinity and conductivity than either N or P and there was also a weak correlation with slope. These have been recognised issues with MTR (Dawson et al., 1999; Thiébaut et al., 2002) and proposals were made to only compare rivers that were physically similar (Baatrup-Pedersen et al., 2006; Szoszkiewicz et al., 2017) to eliminate the response to physical and non-nutrient chemical changes. The LEAFPACS2 method attempted to address these issues and accounted for other factors such as alkalinity, slope and distance from source in the metric calculations to isolate the effects of nutrients in their metrics (WFD-UKTAG, 2014). The results here, would suggest that these measures were successful as no correlation was found between LPEQR and alkalinity or slope (although a weak correlation was found between LPEQR and gravel substrate). Other studies have shown that N and P still only accounted for a very small proportion of the variability in plant communities (Demars et al., 2012) and that intrinsic and geographical variability still prevails in methods that use either trophic indices (such as MTR) or more complex metrics (such as LEAFPACS2) (Baatrup-Pedersen et al., 2006). It is claimed that many indices (including LEAFPACS2 and MTR) have failed to isolate the effects of nutrient enrichment from other physical and non-nutrient chemical factors (Demars et al., 2012) and this seems to be the case in this study. If one had to choose between MTR or LEAFPACS2 however, LEAFPACS2 seems a more suitable option for water quality analysis in terms of nutrients than MTR as there are fewer co-correlations with other factors but remembering that the correlation to phosphate levels is weak.
The relationship between MTR, LEAFPACS and environmental conditions within Irish river vegetation communities
There were significant differences in MTR and LPEQR scores between some of the bryophyte communities however, there was little evidence of a relationship between the MTR/LPEQR scores and any significant differences in nutrient levels. This is surprising as bryophytes are considered important and sensitive bioindicators (Baláži & Hrivnák, 2015). There was one exception, the Leptodictyo riparii-Fissidentetum crassipedis was found to be significantly different in N levels compared to Scapanietum undulatae and also significantly different in both MTR and LPEQR scores. This community is characterised by Leptodictyum riparium (Hedw.) Warnst. which is known to be an indicator of nutrient-enriched conditions (Holmes et al., 1999; Porley & Hodgets, 2005) while the other bryophyte communities in this study including the Scapanietum undulatae tend to prevail in waters that are nutrient poor. A recognised weakness in MTR assessment is that there are a low number of trophic indicator species for upland streams (Szoszkiewicz et al., 2007; Demars et al., 2012) where bryophytes are more likely to prevail, a higher number of bioindicators decreases the risk of status miscalculation (Szoszkiewicz et al., 2007), this could be a contributory factor to the lack of correspondence between the assessment scores and nutrient levels in bryophyte-dominated streams. There were only a few factors that were found to be significantly different between the bryophyte communities, this is most likely due to the fact that most bryophyte communities in this study are from similar types of habitats of higher slope, rocky substrate and lower nutrient levels. The greatest significant differences in environmental conditions, e.g. slope and channel depth were between the bryophyte and the vascular plant communities. Most of the significant differences in MTR and LPEQR were also between bryophyte and vascular plant communities but these did not necessarily correspond to significant differences in N and P alone. Most significant differences were found to be due to a combination with other factors such as alkalinity and/or conductivity, and not necessarily nutrient status. This suggests that environmental factors, including N and P are not easily isolated from each other and that environmental factors effect macrophyte distribution synergistically (Friberg et al., 2011).
In this study, the greater number of significant differences were found between macrophyte communities that are known to be very different in terms of habitat, nutrient status and macrophyte morphology. Where nutrient levels were found to be significantly different between two communities, other non-nutrient factors were also found to play a significant role in community differentiation. There was a significant difference, for example, in the MTR and corresponding P levels between the Nymphaeo albae-Nupharetum luteae (lowland vascular plant community) and Scapanietum undulatae fontinalietosum squamosae (upland bryophyte community). However, other factors, both physical (slope) and chemical (alkalinity) also played a significant role in differentiating these two communities. Likewise, there were notable significant differences in MTR values and P levels between the two vascular plant communities, Myriophylletum alterniflori and Nymphaeo albae-Nupharetum luteae. These communities are known to prevail in very different conditions, the Myriophylletum alterniflori in fast-flowing, oligotrophic waters and the Nymphaeo albae-Nupharetum luteae in slow/still, nutrient-rich waters.
Significant environmental differences between other vascular plant communities were fewer. The likely reason for this is that a range of these vascular plant communities occur in the lower reaches where nutrient levels are generally higher and physical conditions similar, e.g. slower flow and finer sediment, e.g. the Rorippo-Phalaridetum arundinaceae, Sagittario sagittifoliae-Sparganietum emersi and Glycerio-Sparganietum neglecti. This makes distinction between the environmental factors including nutrients, within these communities more difficult.
Implications for using macrophytes for ecological quality assessment
Macrophytes and their associated communities provide a valuable indication of a combination of prevailing environmental conditions, but it is difficult to target one causal factor (Dawson & Szoszkiewicz, 1999; Thiébaut & Muller, 1999). It has been shown in this study and others that nutrient levels are not the most influential factors on macrophyte richness or distribution (Matson, 2006; Demars et al., 2012). Macrophytes show a high degree of natural variability (Schaumberg et al., 2004) making assessment tools based on macrophytes difficult to perfect.
Furthermore, indications from this study would suggest that MTR and LPEQRs and hence their macrophyte indicators are not necessarily picking up changes in nutrient levels alone but also reflect a combination of changes in prevailing conditions. As a result, one might be better served by using macrophytes as indicators of overall ecological status that includes a combination of factors, not just trophic levels alone (Schaumberg et al., 2004).
Communities that were very distinct from one another were effective at bioindication of a combination of environmental changes, these communities tended to be at opposite ends of the ecological spectrum (e.g. Myriophylletum alterniflori versus Nymphaeo albaeNupharetum luteae). More subtle changes between communities of similar habitat and conditions were not so easily detected, suggesting that macrophyte communities and indicator species give us an estimation of ecological changes on a broad scale but not on a fine scale (Demars & Harper, 1998).
To detect changes on a finer scale, the examination of growth form and plant traits may provide more information on prevailing conditions. For example, no apparent significant environmental differences were found between the Rorippo-Phalaridetum arundinaceae and the Sagittario sagittifoliae-Sparganietum emersi as both occur in lowland slower flowing rivers with finer substrate and higher nutrient levels. Studies would suggest that substrate type and flow is one of the key factors in macrophyte distribution (Baatrup-Pedersen & Riis, 1999; Matson, 2006) and if this is the case here, changes will be hard to detect between these two communities. If the macrophyte morphology is examined however, notable differences can be found. For example, the Rorippo-Phalaridetum arundinaceae is dominated by Phalarus arundinacea L. a tall emergent/marginal grass that tolerates occasional flooding, whereas the Sagittario sagittifoliae-Sparganietum emersi is either dominated by Sagittaria sagittifolia L. and/or Sparganium emersum L. both of which possess permanently submerged strap-like leaves and emergent leaves that are morphologically different. There is evidence to suggest that examining plant morphology and plant traits, along with species abundance and assemblages can be more successful in determining both anthropogenic disturbance and hydrological changes (Daniel et al., 2006; Moncão et al., 2012). Conversely, river plants can also have a direct effect on channel structure and increase habitat heterogeneity (O’Briain et al., 2017) and some can species act as landform engineers (Gurnell, 2014).
Another consideration would be to take aquatic algae into account, this group is not always recorded consistently during river vegetation surveys (Baláži & Hrivnák, 2015; Weekes et al., 2018a) despite algae being an essential component of river vegetation (Baláži & Hrivnák, 2015) and sensitive to nutrient enrichment (Blum, 1956). Anecdotal evidence from 3 years of river macrophyte surveying by the author would indicate that the proportion of growth of macroalgal species within both vascular plant and bryophyte communities is important in reflecting trophic changes in a river stretch. Algal cover is included in the metrics of some freshwater assessment tools, however, there is no consistency as to what % algae cover best reflects trophic changes. For example, the reference value (high water quality) for algal cover is set as 0.05% within the LEAFPACS2 method; moderate river water quality is indicated by an algal cover of > 50% in the CBAS (Canonical Correspondent Analysis (CCA)-Based Assessment System (NSSHARE, 2008), while a cover of 5% was set for the threshold of good/moderate in the ‘Rapid Assessment of PeriPhyton Ecology in Rivers’ (RAPPER method) (Kelly et al., 2016). This shows there is plenty of scope for further research on this topic.
Conclusions
It is difficult to attribute changes in macrophyte distribution to any one environmental factor as macrophytes and their communities tend to respond to a combination of factors. Macrophytes are not necessarily the most efficient taxonomic group to apply when focussing on trophic changes alone. This has implications for the use of water quality assessment tools such as the MTR and LEAFPACS2 that endeavour to indicate nutrient enrichment based on macrophytes alone. Both methods were found to be only weakly correlated with nutrient levels and tend to indicate a combination of environmental differences. The value of macrophytes in informing ecological assessment might be best focussed on either their use as indicators of overall quality or ‘health’ of a river channel (encompassing both physical structure and chemical properties of water) or enhancing their use as trophic indicators by combining information on plant species traits and integrating macroalgae genera/species more comprehensively into survey methods and assessment calculations.
Data availability
Some of the data for this research are freely available as they are part of the River Macrophyte Database held within the National Vegetation Database, National Biodiversity Data Centre, Waterford, Ireland. However, some data used in this research were shared with the authors with special permission from the owners of the data and are not freely available without direct contact with those owners.
References
Aguiar, F. C., P. Segurado, G. Urbanič, J. Cambra, C. Chauvin, S. Ciadamidaro, G. Dörflinger, J. Ferreira, M. Germ, P. Manolaki, M. R. Minciardi, A. Munné, E. Papastergiadou & M. T. Ferreira, 2014. Comparability of river quality assessment using macrophytes: a multistep procedure to overcome biogeographical differences. Science of the Total Environment 476–477: 757–767.
Baatrup-Pedersen, A. & T. Riis, 1999. Macrophyte diversity and composition in relation to substratum characteristics in regulated and unregulated Danish streams. Freshwater Biology 42: 375–385.
Baatrup-Pedersen, A., K. Szoszkiewicz, R. Nijboer, M. T. O’Hare & T. Ferreira, 2006. Macrophyte communities in unimpacted European streams: variability in assemblage patterns, abundance and diversity. Hydrobiologia 566: 179–196.
Baatrup-Pedersen, A., E. Göthe, T. Riis, D. K. Andersen & S. E. Larsen, 2017. A new paradigm for biomonitoring: an example building on the Danish Stream Plant Index. Methods in Ecology and Evolution 8: 297–307.
Baláži, P. & R. Hrivnák, 2015. Bryophytes and macro-algal growths as a part of macrophyte monitoring in rivers used for ecological assessment. Knowledge Management Aquatic Ecosystems. 416: 19.
Barton, K. 2016. MuMIn: Multi-Model Inference. R package version 1.15.6. https://CRAN.R-project.org/package=MuMIn.
Becker, R.A., J.M. Chambers & A.R. Wilks, 1988. The New S Language. A Programming Environment for Data Analysis and Graphics (Wadsworth & Brooks/Cole computer science series). Chapman & Hall, London, UK.
Blum, J. L., 1956. The ecology of river Algae. Botanical Review 22: 291–341.
Camargo, J., 2018. Responses of aquatic macrophytes to anthropogenic pressures: comparison between macrophyte metrics and indices. Environmental Monitoring and Assessment 190: 173.
Daniel, H., I. Bernez & J. Haury, 2006. Relationships between macrophytic vegetation and physical features of river habitats: the need for a morphological approach. Hydrobiologia 570: 11–17.
Dawson, F. H, J. R. Newman, M. J. Gravelle, K. J. Rouen & P. Henville, 1999. Assessment of the trophic status of rivers using macrophytes. Evaluation of the mean trophic rank. R & D Technical Report E39. Environment Agency, Bristol, UK.
Dawson, F. H. & K. Szoszkiewicz, 1999. Relationships of some ecological factors with the associations of vegetation in British rivers. Hydrobiologia 415: 117–122.
Demars, B. O. L. & D. M. Harper, 1998. The aquatic macrophytes of an English lowland river system: assessing response to nutrient enrichment. Hydrobiologia 384: 75–88.
Demars, B. O. L. & G. Thiébaut, 2008. Distribution of aquatic plants in the Northern Vosges rivers: implications for biomonitoring and conservation. Aquatic Conservation: Marine and Freshwater Ecosystems 18: 619–632.
Demars, B. O. L., J. M. Potts, M. Trémolières, G. Thiébaut, N. Gougelin & V. Nordmann, 2012. River macrophyte indices: not the Holy Grail! Freshwater Biology 57: 1745–1759.
Friberg, N., N. Bonada, D. C. Bradley, M. J. Dunbar, F. K. Edwards, J. Grey, R. B. Hayes, A. G. Hildrew, N. Lamouroux, M. Trimmer & G. Woodward, 2011. Biomonitoring of human impacts in natural ecosystems: the good, the bad and the ugly. Advances in Ecological Research 44: 1–68.
Gurnell, A., 2014. Plants as river system engineers. Earth Surface Processes and Landforms 39: 4–25.
Harrell, F.E. Jr 2016. Hmisc: Harrell Miscellaneous. R package version. 4.0-2. https://CRAN.R-project.org/package=Hmisc.
Holmes, N., J.R. Newman, S. Chadd, K.J. Rouen, L. Saint & F.H. Dawson, 1999. Mean Trophic Rank: A User’s Manual. R & D Technical Report E38, Environment Agency, Bristol, UK.
Jones, J. I., A. L. Collins, P. S. Naden & D. A. Sear, 2012. The relationship between fine sediment and macrophytes in rivers. River Research and Applications 28: 1006–1018.
Jordan, C & A. Higgins, 2009. The AFBI Soil Classification Map of Northern Ireland at 1:250,000 scale. Agri-Food and Biosciences Institute, Belfast. (Based on information from Cruickshank, J.G. (ed.) 1997. Soil and Environment: Northern Ireland. Agricultural and Environmental Science Department, Queen’s University, Belfast, NI.).
Kalogirou, S. 2016. lctools: Local Correlation, Spatial Inequalities, Geographically Weighted Regression and Other Tools. R package version 0.2-5. https://CRAN.Rproject.org/package=lctools.
Kelly, M. G., J. Krokowski & J. P. C. Harding, 2016. RAPPER: a new method for rapid assessment of macroalgae as a complement to diatom-based assessments of ecological status. Science of the Total Environment 568: 536–545.
Lewin, I. & K. Szoszkiewicz, 2012. Drivers of macrophyte development in rivers in an agricultural area: indicative species reactions. Central European Journal of Biology 7: 731–740.
MacQueen, J. 1967. Some methods for classification and analysis of multivariate observations. In L. M. L. Cam & J. Neyman (eds) Proceedings of the Fifth Berkeley Symposium on Mathematical Statistics and Probability. University of California Press, Berkeley, CA: 281–297.
Matson, R.P.H. 2006. The Use of Macrophytes as Indicators in the Bioassessment of Riverine Environmental Quality in Ireland. Unpublished Ph.D., University College, Dublin.
Mayora, G., B. Schneider & A. Rossi, 2018. Turbidity and dissolved organic matter as significant predictors of spatio- temporal dynamics of phosphorus in a large river floodplain system. River Research and Applications 34: 629–639.
McCune, B. & Mefford, M.J. 2011. PC-ORD. Multivariate analysis of Ecological Data, Version 6.0 for Windows. MjM Software, Gleneden Beach, Oregon, USA.
Moncão, F. S., A. Medeiros dos Santos & L. M. Bini, 2012. Aquatic macrophyte traits and habitat utilization in the Upper Paraná River floodplain, Brazil. Aquatic Botany 102: 50–55.
North South Shared Aquatic Resource (NS Share) 2008. Methods Manual I River Macrophytes. Department of Environment (DOE), Northern Ireland, and the Department of Environment Heritage and Local Government (DEHLG), IRL.
O’Briain, R., S. Shephard & B. Coghlan, 2017. Pioneer macrophyte species engineer finescale physical heterogeneity in a shallow lowland river. Ecological Engineering 102: 451–458.
Pielou, E. C., 1969. An Introduction to Mathematical Ecology. Wiley, New York.
Porley, R. & N. Hodgetts, 2005. Mosses and Liverworts. The New Naturalist Library. A Survey of British natural history. HarperCollins, London.
R Core Team (2018) R: A language and environment for statistical computing. R foundation for statistical computing, Vienna. https://www.R-project.org.
Raven, P., N. Holmes, H. Dawson, A. Ławniczak, E. Bulánková, J. Topercer & I. Lewin, 2011. River Habitat and Macrophyte Surveys in the High Tatra Mountains of Slovakia and Poland: Results from 2010. Environment Agency, Bristol.
Scarlett, P. & M. T. O’Hare, 2006. Community structure of in-stream bryophytes in English and Welsh rivers. Hydrobiologia 553: 143–152.
Schaumburg, J., C. Schranz, J. Foerster, A. Gutowski, G. Hofmann, P. Meilinger, S. Schneider & U. Schmedtje, 2004. Ecological classification of macrophytes and phytobenthos for rivers in Germany according to the Water Framework Directive. Limnologica 34: 283–301.
Sekulová, L., M. Hájek, P. Hájková, E. Mikulášková & Z. Fajmonová, 2011. Alpine wetlands in the West Carpathians: vegetation survey and vegetation–environment relationships. Preslia 83: 1–24.
Simpson, E. H., 1949. Measurement of diversity. Nature 193: 688.
Steffen, K., T. Becker, W. Herr & C. Leuschner, 2013. Diversity loss in the macrophyte vegetation of northwest German streams and rivers between the 1950s and 2010. Hydrobiologia 713: 1–17.
Steffen, K., C. Leuschner, U. Müller, G. Wiegleb & T. Becker, 2014. Relationships between macrophyte vegetation and physical and chemical conditions in northwest German running waters. Aquatic Botany 113: 46–55.
Szoszkiewicz, K., K. Karolewicz, A. Ławniczak & F. H. Dawson, 2002. An assessment of the MTR aquatic plant bioindication system for determining the trophic status of polish rivers. Polish Journal of Environmental Studies 11: 421–427.
Szoszkiewicz, K., S. Jusik, T. Zgola, M. Czechowska & B. Hryc, 2007. Uncertainty of macrophyte-based monitoring for different types of lowland rivers. Belgian Journal of Botany 140: 7–16.
Szoszkiewicz, K., J. Zbierska, R. Staniszewski & S. Jusik, 2009. The variability of macrophyte metrics used in river monitoring. International Journal of Oceanography and Hydrobiology. 38: 117–126.
Szoszkiewicz, K., H. Ciecierska, A. Kolada, S. C. Schneider, M. Szwabińska & J. Ruszczyńska, 2014. Parameters structuring macrophyte communities in rivers and lakes—results from a case study in North-Central Poland. Knowledge and Management of Aquatic Ecosystems 415: 08.
Szoszkiewicz, K., A. Budka, K. Pietruczuk, D. Kayzer & D. Gebler, 2017. Is the macrophyte diversification along the trophic gradient distinct enough for river monitoring? Environmental Monitoring and Assessment 189: 4.
Thiébaut, G. & S. Muller, 1999. A macrophyte communities sequence as an indicator of eutrophication and acidification levels in weakly mineralised streams in north-eastern France. Hydrobiologia 410:17–24.
Thiébaut, G., F. Guérold & S. Muller, 2002. Are trophic and diversity indices based on macrophyte communities pertinent tools to monitor water quality? Water Research 36: 3602–3610.
Tichý, L., 2002. JUICE, software for vegetation classification. Journal of Vegetation Science 133: 451–453.
Venables, W. N. & B. D. Ripley, 2002. Modern Applied Statistics with S, 4th ed. Springer, New York.
Water Framework Directive (WFD) 2000/60/EC Establishing a framework for Community action in the field of water policy. Official Journal of the European Communities L327: 1–73.
Water Framework Directive—United Kingdom Technical Advisory Group (WFD-UKTAG) 2014. UKTAG River Assessment Method Macrophytes and Phytobenthos: Macrophytes (River LEAFPACS2). Water Framework Directive–United Kingdom Advisory Group, Sterling, Scotland, UK.
Weekes, L., Ú. FitzPatrick, F. Kelly, R. Matson & M. Kelly-Quinn, 2018a. A review of the Irish River Macrophyte Database: informing future river macrophyte surveying. Biology and Environment: Proceedings of the Royal Irish Academy 118: 1–33.
Weekes, L., Z. Kącki, Ú. FitzPatrick, F. Kelly, R. Matson & M. Kelly-Quinn, 2018b. An Irish National Vegetation Classification System for aquatic river macrophytes. Applied Vegetation Science 21: 322–340.
Wickham, H., 2009. ggplot2: Elegant Graphics for Data Analysis. Springer, New York.
Willby, N. J., J. Pitt & G.L. Phillips, 2010. The ecological classification of UK rivers using aquatic macrophytes. Environment Agency Science Report, UK.
Willby, N., J. A. Pitt & G. Phillips, 2012. The Ecological Classification of UK Lakes Using Aquatic Macrophytes. Environment Agency, Bristol.
Acknowledgments
The authors are very grateful to the Irish Research Council for their funding for this research through their Employment-based Postgraduate Programme. We are also very grateful to all the data providers and the National Biodiversity Data Centre for providing the data, without which, this project would not have been possible. A special thank you to Dr Deirdre McClean and Dr Thomás Murray for their expertise and advice on the statistical analysis. Finally a special thank you to the anonymous reviewers that gave of their time to thoroughly review this paper.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Katya E. Kovalenko
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This paper has not been submitted elsewhere in identical or similar form, nor will it be during 21 the first three months after its submission to Hydrobiologia.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Weekes, L., FitzPatrick, Ú. & Kelly-Quinn, M. Assessment of the efficiency of river macrophytes to detect water-column nutrient levels and other environmental conditions in Irish rivers. Hydrobiologia 848, 2797–2814 (2021). https://doi.org/10.1007/s10750-021-04598-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-021-04598-7