
Abstract
The digestive capacity of jellyfish has barely been studied because of the complexity of their life stage body-forms and sizes. To assess the digestive capacity in the changes from the benthic polyp-form to the pelagic stage in the cannonball jellyfish Stomolophus meleagris, we used fluorometric and spectrophotometric techniques for measuring the activity of trypsin, chymotrypsin, aminopeptidase, amylase, lipase, phosphatase acid, and alkaline. The findings showed that at all stages were able to digest proteins, carbohydrates, and lipids, but the digestive capacity differs between the benthic–pelagic stage and final development of the ephyrae, suggesting a shift in nutrient requirements. The digestive capacity indicated that polyps better assimilate carbohydrates and proteins. During the pelagic stage, a shift in the digestive capacity occurred, where 5-day ephyrae better assimilate proteins, while 15-day-old ephyrae were better with lipids and proteins and juvenile medusae with lipids. This is the first study to describe changes in the digestive capacity of cannonball jellyfish and can help identify ecological features of an important fishing resource with potential aquaculture interest. Different diets for polyps, ephyrae, and juvenile medusae must be considered if the species is cultivated.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The cannonball jellyfish Stomolophus meleagris Agassiz, 1862, is one of the most globally appreciated edible jellyfish, found from New England to Brazil in the Atlantic and southern California to the equator in the Pacific (Kramp, 1961; Larson, 1976; Omori, 1978). The cannonball jellyfish has a metagenic life cycle with a benthic polypoid stage that can reproduce asexually, producing new polyps or giving rise to ephyrae by strobilation, which is a process that requires disk formation and metamorphosis in which structures of the polyp are lost and replaced in each disk with those of a developing ephyrae. Pelagic ephyrae develop into a juvenile medusoid stage with a radially symmetric bell or umbrella, in which contraction of the subumbrella muscles squeezes the bell to generate pulsations for swimming. It grows and matures into adult medusae that sexually reproduce to form planula larvae that settle and metamorphose into polyps, thus closing the life cycle (Calder, 1982; Arai, 1997).
Rhizostome scyphomedusae, like S. meleagris, lack tentacles; instead, they develop oral arms that filter zooplankton as the medusae swims (Costello & Colin, 1995). In contrast, polyps capture prey on their tentacles and bring the prey into the mouth (Calder, 1982). Scyphomedusae are primarily carnivores, using a wide selection of zooplankton when it is available (Mills, 1995). Some phytoplankton may be ingested, but the amount is not significant in comparison with zooplankton.
Less is known about feeding of scyphozoan polyps eating a variety of pelagic organism. In the laboratory, Artemia, copepods, decapod larvae, mollusks larvae, fish eggs, and larvae have been used (Cargo, 1975; Purcell et al., 2001). Larson (1991) indicates that S. meleagris selects against calanoid and cyclopoid copepods and their nauplii, while Padilla-Serrato et al. (2013) show that anchovy eggs were the most abundant prey, and Álvarez-Tello et al. (2016) show positive selection of fish eggs, gastropods, bivalves, and cirripeds and negative selection for copepods and appendicularians.
Cannonball jellyfish can be seasonally very abundant and could be an important ecological predator of zooplankton. This species has been commercially fished since 2010 for export to China (López-Martínez & Álvarez-Tello, 2013), although polyps have not yet been found in the field. Thus, no information is available on feeding or nutrient requirements for polyps of this species.
Digestive enzymes functionally link the process of ingestion and assimilation (Mayzaud, 1986). Digestive enzyme analysis helps to determine which dietary components are most effectively assimilated and dietary preferences, since a high activity of an enzyme reflects a diet high in the metabolic substrate in which it acts (Johnston & Freeman, 2005).
Estimates of the digestive enzyme activity have been frequently used to understand the digestive physiology and nutritional requirements in a great variety of marine animals, such as crustaceans (Biesiot & Capuzzo, 1990; Lovett & Felder, 1990; Lemos et al., 2002) and fish (Álvarez-González et al., 2005), but have rarely been used in jellyfish. Studies of enzymatic activity in scyphozoa are old, scarce, and focus on adult jellyfish such as S. meleagris, Mastigias papua (Lesson), Cassiopea frondosa (Pallas, 1774), Chrysaora quinquecirrha (Desor, 1848), Cyanea capillata (Linnaeus, 1758), and Aurelia aurita (Linnaeus, 1758) (Bodansky & Rose, 1922; Ohtsuki, 1930; Smith, 1937; Stewart & Lakshmanan 1975; Manchenko & Zaslavskaya, 1980; Båmstedt, 1988).
In some jellyfish, such as C. capillata, Lychnorhiza lucerna Haeckel, 1880, and S. meleagris, the ability to capture prey is related to the size of jellyfish, so changes in size and morphology are accompanied by alterations in feeding (Higgins et al., 2008; Álvarez-Tello et al., 2016, Nagata et al., 2016). In particular, the life cycle of cannonball jellyfish goes through a series of eco-physiological limitations and changes in morphology and feeding mechanisms that can influence the nutrient intake and affect the metabolic requirements of each developmental stage.
The objective of this study was to evaluate the digestive capacity of polyp, ephyrae, and medusae produced under controlled conditions, relating the digestive enzymatic activity to changes in the capacity to hydrolyze the main dietary nutrients. This approach is an essential step toward understanding the nutritional needs during the complex life cycle of a jellyfish.
Materials and methods
Origin and maintenance of experimental organisms
Thirty wild mature cannonball jellyfish (S. meleagris), with a mean wet mass of 338 g, were collected with commercial spoon nets (42 cm diameter, ≤ 5 mm mesh size) in Bahía de La Paz, B.C.S., Mexico (MFRS; 24°11′N, 110°27′W) in May 2015. Collection was obtained about 20 m from shore; water temperature was 24°C.
The jellyfish were transported to facilities at CIBNOR and were placed into a 2000-l oval fiberglass tank containing filtered (10 μm) seawater at 35 salinity. No aeration was provided, and temperature was maintained at 24°C in a controlled temperature research chamber. The captured jellyfish were allowed to spawn for 1 day, and no food was supplied; they were then returned to the sea. After about 2 days, larvae planula began to settle in 20-ml plastic containers placed at the bottom of the tank, where polyps subsequently developed.
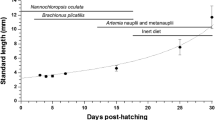
The initial polyps were fed a mixture of microalgae (Nannochloropsis oculata (Droop) D.J. Hibberd, 1981), rotifers (Brachionus plicatilis Müller, 1786), and Artemia franciscana Kellog, 1906 nauplii that were produced daily at our facilities. As soon as disk formation in the polyps was observed, the plastic containers were transferred to the experimental development system, where all stages (polyp, ephyra, and medusa) were maintained.
The experimental development system consisted of a 300-l conic fiberglass tank filled with filtered (1 μm) seawater at 35 salinity at three quarters of water content with constant aeration from the bottom and kept at 24°C, using aquarium heaters; photoperiod was 12:12 light/dark. The water quality was monitored daily to maintain constant conditions of salinity and temperature, and when necessary, salinity was adjusted with distilled water. Three perforated trays were suspended at the top of the tank, but completely immersed in the tank) where the plastic containers with the polyps were placed. Polyps, ephyrae, and medusae were fed ad libitum three times a week for 2 months with newly hatched Artemia franciscana nauplii.
Measurements and sampling
To describe the principal morphological traits of each group, observations were made under a stereomicroscope and recorded with digital photographs. From previous observations of morphological changes (Fig. 1), the jellyfish were separated into six groups: (1) polyp; (2) ephyra 1; (3) ephyra 5; (4) ephyra 10; (5) ephyra 15; and (6) juvenile medusa. Polyps consider only non-strobilating organisms with 16-tentacles; ephyrae were distinguished at 5-day intervals following strobilation, and juvenile medusae was defined as the stage when all morphological characteristics are similar to an adult, but no gonads are visible; this sequence was accomplished 20–23 days following strobilation. The samples were taken out of the experimental system; each was separated according to the appropriate group and placed in clean seawater to eliminate remains of food.

Specific enzymatic activities in the polyp, ephyra, and juvenile medusa stages of Stomolophus meleagris (mean and standard deviation); spectrophotometry methods used: a amylase and b aminopeptidase. Different letters on the columns indicate a significant difference (P < 0.05)
To estimate growth, five jellyfish in each group (except polyps), the central disk of each jellyfish was measured with the program AxionVision 4.2., without including the arms, as described by Båmstedt et al. (1999) (Table 1). Another five jellyfish were weighed in an ultra-balance after washing with de-ionized water; excess water was removed with a soft handkerchief.
Enzymatic assays
With the difference in size and tissue of the jellyfish of each group, preliminary work was done to establish the number of jellyfish necessary to get enough enzymatic extract for all analyzes. The results were: polyps (n = 198); ephyra 1 (n = 1174); ephyra 5 (n = 300); ephyra 10 (n = 78); ephyra 15 (n = 55); and juvenile medusae (n = 6). Whole jellyfish were washed three times with milli-Q water and homogenized in cold, using a tissue homogenizer (PRO Scientific, Oxford, CT) at a speed of 3000 cycles/s, two cycles of 15 s each, and then centrifuged (13,000 g at 4°C for 15 min). The supernatant was recovered, aliquoted in different microtubes, and stored at − 80°C until the analysis for enzyme activity was performed. Preliminary work done on medusa field samples indicated that enzymes were stable when frozen.
Trypsin, chymotrypsin, and lipase activities were determined in microplates, using the modified fluorometric method described in Toledo-Cuevas et al. (2011). Phosphatase activity was determined by the Molecular Probes protocol adapted by Toledo-Cuevas et al. (2011). Amylase (Vega-Villasante et al., 1993) and aminopeptidase N (Maraux et al., 1973) activities were determined by photometry. All assays were performed in triplicate and read in a multimode microplate reader (Varioskan LUX, Thermo Fisher Scientific, Waltham, MA).
Trypsin activity was measured using Boc-Gln-Ala-Arg-7 amido-4 methylcoumarin hydrochloride as substrate at a concentration of 0.8 mM in dimethyl sulfoxide (DMSO) and 50 mM Tris–HCl and 10 mM CaCl2 buffer at pH 7.5. Chymotrypsin was assayed using N-succinyl-Ala-Ala-Pro-Phe-7 amido-4-methylcoumarin at a concentration of 0.8 mM 194.8 ul in DMSO and 50 mM Tris–HCl and 10 mM CaCl2 buffer at pH 7.5. Fluorescence was measured at 460 nm emission and excitation at 355 nm. Aminopeptidase activity was measured using l-leucine p-nitroanilide as substrate at a concentration of 0.1 M in DMSO and 80 mM phosphate buffer at pH 7. Absorbance was read at 410 nm.
Lipase activity in extracts was determined using metillumbelliferyl heptanoate as substrate at a concentration of 50 mM in DMSO and 0.1 M phosphate buffer at pH 7.5. Fluorescence was measured at 460 nm emission and excitation at 355 nm. Amylase was assayed using starch as substrate at 1% in Tris–HCL 50 mM at pH 7.5 and read at 550 nm.
Alkaline phosphatase activity was quantified using DiFMUP (6,8 difluoro-4 methylumbelliferyl phosphate) at a concentration of 200 μM in DMSO and 100 mM glycine buffer, 1 mM MgCl2, 1 mM ZnCl2 at pH 10.4, which was read at 455 nm emission and excitation at 358 nm. Acid phosphatase was measured using DiFMUP at a concentration of 200 µM in DMSO and 100 mM sodium acetate buffer at pH 10.4. Fluorescence was measured at emission of 455 nm and excitation at 358 nm.
Digestive enzyme activity was expressed as U per mg protein−1 (specific). Enzymatic activity values were standardized to the concentration of soluble protein, using the analysis described by Bradford (1976). Bovine serum albumin was used as the standard (Sigma, A4503).
Statistical analysis
Normality and homoscedasticity of the data were tested using Shapiro–Wilk’s test and Levene’s test, respectively. Since the data were not normal, the Friedman test was used to determine the enzymatic activity of the different stages. Differences of P < 0.05 were considered statistically significant. From the data of enzymatic activity, 15 bootstrap pseudo-replicates were performed using the boot function in R software. Mass and size values from ephyra 1 to juvenile medusa were fitted to the best model that described growth rate.
Results
The main morphological characteristics of the changes in the ephyrae to medusae groups were observed (Table 1, 2). The polyp represents the benthic stage. It is sessile and has a cone-shaped calyx with 16 tentacles that capture its food and a small mouth in the middle. The polyp lacks gastric filaments (Table 1).
Ephyrae are the first free pelagic stage with eight arms and a small central mouth or manubrium. When the ephyrae swim, they generate currents with the arms that allow capture of prey in their subumbrellar region. As the ephyra grows, the manubrium becomes more complex, with folds and oral tentacles, finally merging to form the oral arms. The central disk grows, the mesoglea increases, and the bell is formed.
Table 2 shows mean mass and size of S. meleagris at each stage and soluble protein content. For the pelagic stage, mass increase fitted best to a logistic model: y = – 2.5/[1 + (–1.6]e−2.5x), while size increased exponentially: y = – 1.5(7.7–e−(−8.18)x), both with a correlation coefficient value of 0.99. The greatest growth was between ephyra 15 (E15) and juvenile medusae (JM), when the bell shape of a characteristic jellyfish was completed (Table 1).
All stages could hydrolyze carbohydrates, protein, and lipid substrates, as indicated by specific enzymatic activity, which showed significant differences (P < 0.01) between the stages (Figs. 1, 2). Aminopeptidase and amylase activity were significantly higher in polyps, decreasing thereafter to the lowest activity in the JM (P < 0.01) (Fig. 1a, b). A significantly higher amylase activity (P < 0.001) was found in polyps (26 U mg protein−1) and the lowest in JM (7 U mg protein−1). For aminopeptidase, a significant difference occurred in polyps, relative to E15 and JM (P < 0.001), where the activity in polyps was 114 U mg protein−1 and 13 U mg protein−1 in JM.

Specific enzymatic activities in the polyp, ephyra, and juvenile medusa stages of Stomolophus meleagris (mean and standard deviation). Fluorometric methods used: a trypsin, b chymotrypsin, c alkaline phosphatase, d acid phosphatase, and e lipase. Different letters on the columns indicate a significant difference (P < 0.05)
The highest trypsin activity (23,801 U mg protein−1) was found in ephyra 5 (E5), significantly decreasing (P < 0.01) thereafter to the lowest trypsin activity in JM (7820 U mg protein−1) (Fig. 2a). Chymotrypsin activity was significantly higher in E5 and different to ephyra 1 (E1) and JM (P < 0.01) (Fig. 2b). The lowest chymotrypsin activity occurred in E1 (406,672 U mg protein−1) and JM (407,093 U mg protein−1).
Alkaline and acidic phosphatase activity showed similar patterns (Figs. 2c, d), with significant lower activity in phosphatase and significant higher activity in E15 (P < 0.01). Lipase activity was significantly higher in E15 (357,780 U mg protein−1) and different to polyps (123,424 U mg protein−1) and E1 (155,353 U mg protein−1), which showed the lowest lipase activity (Fig. 2e).
Discussion
Digestive enzymes produced by the different stages of cannonball jellyfish give evidence of the types of usable metabolic fuels, thus connecting enzymatic activity and substrate availability (Johnston & Freeman, 2005). In general, greater enzymatic activity implies greater food intake (Boucher et al., 1975; Hoeger & Mommsen, 1984).
Enzyme synthesis is under genetic control in other invertebrates, i.e., lobster and shrimp larvae fed Artemia and can be modulated by the diet, metabolic changes and nutritional requirements to optimize digestive efficiency (Boucher et al., 1975; Samain et al., 1980; Biesiot, 1986; Lovett & Felder, 1990). In crustaceans, it has been shown that the activity of amylase and trypsin is regulated by trophic conditions, such as particle size, quantity, and biochemical composition of food (Boucher et al., 1975).
The digestive capacity in jellyfish has been barely studied because of the complexity of their life stage body-forms and sizes. No previous study was available that related to changes in the digestive enzyme activity in scyphozoan jellyfish. Digestion in scyphozoan involves an extracellular phase, as well as a following intracellular phase. The gastrovascular system of scyphozoan consists of a central stomach surrounded by stomach pouches and/or a canal system. The gastrodermis of the gastric cirri contains serous cells concentrated in the basal region of the cirri that produce digestive enzymes and can be directly applied to entangled prey (Arai, 1997).
Intracelullar digestion occurs in the gastrodermal cells, following endocytosis of particles derived from food. The enzymes carboxylesterase, N-acetylglucosaminidase, aminopeptidase, and phosphatases are related to intracellular digestion (Hoeger & Mommsen, 1984). These enzymes reduce food into small particles, then ingested through phagocytosis to finish with intracellular food digestion. Later the digested products reach the ectoderm by diffusing across the mesoglea (Chapman & Pardy, 1972).
The specific activity of amylase is positively correlated with the amount of phytoplankton cells ingested and assimilated, so a high enzymatic activity indicates that the phytoplankton are important in the diet of jellyfish (Boucher et al., 1975). In some crustacean larvae, high amylase activities could result from co-regulation with other enzymes, such as trypsin (Hirche & Anger, 1987).
High amylase activity at the polyp stage indicates major digestive capacity to assimilate simple meals, such as phytoplankton or dissolved organic material (Boucher et al., 1975). According to Östman (1997) the scyphopolyps, including S. meleagris, feed on bacteria, organic particulate matter, small crustaceans, microalgae, and tintinnids, using their tentacles. Moreover, the polyps absorbed free amino acids, such as glycine and alanine, which are a supplemental nutritional source, necessary for normal strobilation and development (Shick, 1975).
The higher activities of amylase and aminopeptidase found in polyps imply an advantage because they are sessile with limited opportunities to encounter prey, so have to capture very small particles and organisms with little mobility, and these enzymes easily assimilate foods, such as phytoplankton and organic particulate matter. In ephyrae and medusae, amylase and aminopeptidase activities decreased, which could be related to adaptation processes in the pelagic life and a diet shift.
Trypsin and chymotrypsin patterns in S. meleagris are similar to those observed in some larval stages of cephalopod (Villanueva et al., 2002; Moguel et al., 2010) and crustacean species, such as American lobster Homarus americanus H. Milne Edwards, 1837 and white shrimp Litopenaeus schmitti (Burkenroad, 1936) (Biesiot & Capuzzo, 1990; Lemos et al., 2002). In Octopus maya Voss & Solís, 1966 during the larval phase, erratic behavior in enzyme activity has been observed; trypsin and chymotrypsin showed activity peaks at days 3, 5, 9, and 14 after hatching (Moguel et al., 2010). Similarly, in experiments with Octopus vulgaris Cuvier, 1797 fed with Artemia, increased proteolytic activity (trypsin and chymotrypsin) occurred during the first days, but tended to decrease after day 10 (Villanueva et al., 2002). These patterns could be related to adaptation processes in diet shift, based on changes in nutritional requirements or feeding behavior and an increase in complexity during development.
The ephyra is a pelagic stage, but has limited swimming ability; thus, they need to capture slow-swimming organisms by creating fluid flows that allow capturing prey into the subumbrellar region (Kamiyama, 2017). According to Gómez-Aguirre (1991), the first feeding stages of S. meleagris ephyrae consist of small microplankton, such as diatoms, dinoflagellates, protozoa, and larvae of other invertebrates.
During development of the ephyrae, the manubrium became more complex and branched, which allows capture of more preys. The internal canals branch to allow food transport to the whole body; velar lappets developed between marginal lappets, and the central disk increases in size. By E15, ephyrae begin to acquire a hemispherical shape, and the width of the mesoglea (composed of water and collagen-like protein) increased. At this point the highest activity of lipase and phosphatase was observed.
Phosphatases are responsible for catalyzing the separation of inorganic phosphorus from organic phosphate (Lehninger, 1994) to produce energy and transport nutrients through the cells, such as glucose, proteins, and lipids, and even water and ions in the case of alkaline phosphatase (Dupuis et al., 1991; Álvarez-González et al., 2005). Van Praët (1976) showed an increase in acid phosphatase reaction product in endodermal cells of Actinia equina (Linnaeus, 1785) during 10 days of starvation, but a decrease at about 20 h after feeding, interpreting this as using the stock of acid phosphatase during digestion. In S. meleagris, the highest phosphatase activity, both acidic and alkaline, was found in E15.
The highest activity of phosphatase and lipase in E15 is related to exponential growth that is most marked in the transition between the ephyra and medusa stages. In this phase, the jellyfish needs more energy to maintain rapid growth because their feeding now focuses on food with high amounts of lipids, usually eggs of invertebrates, and fish.
Lipids have about twice the energy content as carbohydrates and proteins and are essential for egg production, since S. meleagris females of only 3.6 cm (~ 4.7 g) are already mature. Triglycerides are the main energy storage in S. meleagris that during vitellogenesis accumulate in oocytes (Carvalho-Saucedo et al., 2011).
Based on stomach content, juvenile and adult S. meleagris exhibit is a clear carnivorous specialist, capable of selecting food from the environment. Larson (1991) showed that S. meleagris select against calanoid and cyclopoid copepods and their nauplii, Padilla-Serrato et al. (2013) showed that anchovy eggs were the most abundant prey, and Álvarez-Tello et al. (2016) showed positive selection of fish eggs, gastropods, bivalves, and cirripeds and negative selection for copepods and appendicularians. The type and amount of prey ingested is related to the size of the jellyfish; copepods tend to decrease in the diet as it grows, while fish eggs tend to increase (Álvarez-Tello et al., 2016).
Medusa is the largest stage in the life cycle of S. meleagris, its high swimming capacity concentrates food around their oral parts and produces mucus to retain food items around the oral cavity (Southward, 1955); thus, medusae showed greater capacity to follow and catch prey, as their metabolism is higher and energy requirements increase.
Contrary to what was expected, the specific enzymatic activities for JM are very low. At this stage, there is the greatest increase in size, wet mass, and amount of protein, resulting from the increase in the mesoglea (composed of water and collagen-like protein). Protein concentration increased approximately tenfold from the E15 to the JM stage, which suggests underestimating the amount of enzymatic activity. Apart from this and considering the enzymatic activity, JM has the highest activity.
The same enzyme pattern was observed in other invertebrates, such as the cuttlefish Sepia officinalis Linnaeus, 1758, where trypsin, chymotrypsin, and acid phosphatase decrease the specific activity during the juvenile stages, up to 30 days post-hatching (Perrin et al., 2004); trypsin in Palaemon serratus (Pennant, 1777) (Van Wormhoudt & Sellos, 1980) and fish larvae of Paralichthys californicus (Ayres, 1858) also decrease at 30 days after hatching for trypsin, chymotrypsin, and alkaline phosphatase (Álvarez-González et al., 2005).
Our results indicate that the polyp, ephyra, and juvenile medusa stages are able to digest all metabolic fuels (protein, carbohydrates, and lipids) present in food, but major changes were observed when changing from benthic to pelagic activity and at the late ephyra (E15) stage, when the bell is completely formed and just before exponential mass growth. These changes suggest that the digestive capacity changes between the phases, according to their nutritional requirements and diet, and seems to be regulated internally, since the external factors were the same for each stage of jellyfish growth.
Boucher et al. (1975) state that there is an adaptation in activity of the digestive enzymes to the variations of available food, which allows the jellyfish to satisfy its metabolic needs when there is little available food in the environment. The presence of different types of enzymes (proteases, lipases, phosphatases, and amylases) in the stages of S. meleagris may be an adaptation to the variability in planktonic composition, and therefore their food, which favors high survival, which is reflected in the blooms that seasonally occur in this jellyfish.
Further work should study the ability to absorb and assimilate the principal dietary nutrients and their response to external factor as an essential step toward understanding the mechanism of digestion of scyphozoan jellyfish, which is of particular interest in blooming species, such as S. meleagris, when seasonal outbreaks occur.
In this work, fluorometric and spectrophotometric methods were used to provide new insights into jellyfish digestive capacity and detecting enzymatic activity. However, the fluorometric technique had a higher resolution because the substrates used in the analyses were more unstable and required less enzymes for their hydrolysis, besides requiring less biological material for analysis (Rotllant et al., 2008). Therefore, it was more useful for analyzing the small polyps and ephyrae. This work used a large number of polyps and ephyrae (see “Methods”) to avoid diluting the sample; but given the great sensitivity of the fluorometric technique (c.v. ca. 1%), fewer biological samples could be used in future studies.
In summary, S. meleagris has the capacity to digest all metabolic fuels (protein, carbohydrates, and lipids) in a meal. This ability changes throughout development and is probably tied to the genetic regulation of enzyme synthesis, according to feeding habits and morphological changes during growth. Polyps better hydrolyze carbohydrates, ephyrae better hydrolyze proteins, and this shifts to lipids at the juvenile medusa stage.
References
Álvarez-González, C. A., M. Cervantes-Trujano, D. Tovar-Ramírez, D. E. Conklin, H. Nolasco, E. Gisbert & R. Piedrahita, 2005. Development of digestive enzymes in California halibut Paralichthys californicus larvae. Fish Physiology and Biochemistry 31: 83–93.
Álvarez-Tello, F. J., J. López-Martínez & D. B. Lluch-Cota, 2016. Trophic spectrum and feeding pattern of cannonball jellyfish Stomolophus meleagris (Agassiz, 1862) from central Gulf of California. Journal of the Marine Biological Association of the United Kingdom 96: 1217–1227.
Arai, M. N., 1997. A functional biology of Scyphozoa. Chapman and Hall, London.
Båmstedt, U., 1988. Interspecific, seasonal and diel variations in zooplankton trypsin and amylase activities in Kosterfjorden, western Sweden. Marine Ecology Progress Series 44: 15–24.
Båmstedt, U., J. Lane & M. B. Martinussen, 1999. Bioenergetics of ephyra larvae of the scyphozoan jellyfish Aurelia aurita in relation to temperature and salinity. Marine Biology 135: 89–98.
Biesiot, P. M., 1986. Changes in midgut gland morphology and digestive enzyme activities associated with development in early stages of the American lobster. Doctoral thesis. Woods Hole Oceanographic Institution.
Biesiot, P. M. & J. M. Capuzzo, 1990. Changes in digestive enzyme activities during early development of the American lobster Homarus americanus Milne Edwards. Journal of Experimental Marine Biology and Ecology 136: 107–122.
Bodansky, M. & W. C. Rose, 1922. Comparative studies of digestion. American Journal of Physiology 62: 473–481.
Boucher, J., A. Laurec, J. F. Samain & S. L. Smith, 1975. Etude de la nutrition, du régime et du rythme alimentaire du zooplancton dans les conditions naturelles, par la mesure des activités enzymatiques digestives. Proceedings of the 10th European Symposium on Marine Biology 2: 85–100.
Bradford, M. M., 1976. A rapid and sensitive method for the quantization of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72: 248–254.
Calder, D. R., 1982. Life history of the cannonball jellyfish, Stomolophus meleagris L. Agassiz, 1860 (Scyphozoa, Rhizostomida). Biological Bulletin 162: 149–162.
Cargo, D. G., 1975. Comments on the laboratory culture of Scyphozoa. In Culture of Marine Invertebrate Animals. Plenum Press, New York.
Carvalho-Saucedo, L., J. López-Martínez, F. García- Domínguez, C. Rodríguez-Jaramillo & J. Padilla- Serrato, 2011. Biología reproductiva de la medusa bola de cañón Stomolophus meleagris en la laguna Las Guásimas, Sonora, México. Hidrobiológica 21: 77–88.
Chapman, B. Y. G. & R. L. Pardy, 1972. The movement of glucose and glycine through the tissues of Corymorpha palma torrey (Coelenterata, Hydrozoa). Journal of Experimental Biology 56: 639–645.
Costello, J. H. & S. P. Colin, 1995. Flow and feeding by swimming scyphomedusae. Marine Biology 124: 399–406.
Dupuis, Y., S. Tardival, Z. Poremska & P. Fournier, 1991. Effect of some alkaline phosphatase inhibitors of intestinal calcium transfer. International Journal of Biochemistry 23: 175–180.
Gómez-Aguirre, S., 1991. Larva éfira y diferenciación de Stomolophus meleagris (Scyphozoa: Rhizostomeae) en plancton de lagunas costeras de Tabasco, México. Anales del Instituto de Biología Universidad Nacional Autónoma de México Serie Zoológica 62: 383–389.
Higgins, J. E., M. D. Ford & J. H. Costello, 2008. Transitions in morphology, nematocyst distribution, fluid motions, and prey capture during development of the scyphomedusa Cyanea capillata. Biological Bulletin 214: 29–41.
Hirche, H. J. & K. Anger, 1987. Digestive enzyme activities during larval development of Hyas araneus (Decapoda, Majidae). Comparative Biochemistry and Physiology Part B 87: 297–302.
Hoeger, U. & T. P. Mommsen, 1984. Hydrolytic enzymes in the two North Sea ctenophores Pleurobranchia pileus and Beroe gracilis. Marine Biology 130: 123–130.
Johnston, D. & J. Freeman, 2005. Dietary preference and digestive enzyme activities as indicators of trophic resource utilization by six species of crab. Biological Bulletin 208: 36–46.
Kamiyama, T., 2017. Planktonic ciliates as food for the scyphozoan Aurelia coerulea: feeding and growth responses of ephyra and metephyra stages. Journal of Oceanography 74: 53–63.
Kramp, P., 1961. Synopsis of medusae of the worlds. Journal of the Marine Biological Association of the UK 40: 1–471.
Larson, R. J., 1976. Marine Flora and Fauna of the Northeastern United States. Cnidaria: Scyphozoa. NOAA Technical Report NMFS Circular 397 26 pp.
Larson, R. J., 1991. Diet, prey selection and daily ration of Stomolophus meleagris, a filter-feeding scyphomedusa from the NE Gulf of Mexico. Estuarine, Coastal and Shelf Science 32: 511–525.
Lehninger, A., 1994. Bioquímica, 2nd ed. Ediciones Omega. Barcelona, Spain.
Lemos, D., F. L. García-Carreño, P. Hernández & A. Navarrete del Toro, 2002. Ontogenetic variation in digestive proteinase activity, RNA and DNA content of larval and postlarval white shrimp Litopenaeus schmitti. Aquaculture 214: 363–380.
López-Martínez, J. & J. Álvarez-Tello, 2013. The jellyfish fishery in Mexico. Agricultural Sciences 4: 57–61.
Lovett, D. L. & D. L. Felder, 1990. Ontogenetic change in digestive enzyme activity of larval and postlarval white shrimp Penaeus setiferus (Crustacea, Decapoda, Penaeidae). Biological Bulletin 178: 144–159.
Manchenko, G. P. & N. I. Zaslavskaya, 1980. Genetic-variability of hexokinase and leucine aminopeptidase in the scyphozoan medusa Cyanea capillata. Biologiya Morya 6: 65–68.
Maraux, S., D. Louvard & J. Barath, 1973. The aminopeptidase from hog-intestinal brush border. Biochimica et Biophysica Acta-Enzymology 321: 282–295.
Mayzaud, P., 1986. Enzymatic measurements of metabolic processes concerned with respiration and ammonia excretion. The biological chemistry of copepods. Oxford University Press, Oxford.
Mills, C. E., 1995. Medusae, siphonophores, and ctenophores as planktivorous predators in changing global ecosystems. ICES Journal of Marine Science 52: 575–581.
Moguel, C., M. Mascaró, O. H. Avila-Poveda, C. Caamal-Monsreal, A. Sánchez, C. Pascual & C. Rosas, 2010. Morphological, physiological and behavioral changes during post-hatching development of Octopus maya (Mollusca: Cephalopoda) with special focus on the digestive system. Aquatic Biology 9: 35–48.
Nagata, R. M., A. C. Morandini, S. P. Colin, A. E. Migotto & J. H. Costello, 2016. Transitions in morphologies, fluid regimes, and feeding mechanisms during development of the medusa Lychnorhiza lucerna. Marine Ecology Progress Series 557: 145–159.
Ohtsuki, T., 1930. Digestive enzymes of some marine animals. Zoological Magazine 42: 411–421. (In Japanese).
Omori, M., 1978. Zooplankton fisheries of the world: a review. Marine Biology 48: 199–205.
Östman, C., 1997. Abundance, feeding behaviour and nematocysts of scyphopolyps (Cnidaria) and nematocysts in their predator, the nudibranch Coryphella verrucosa (Mollusca). Hydrobiologia 355: 21–28.
Padilla-Serrato, J. G., J. López-Martínez, A. Acevedo-Cervantes, E. Alcántara-Razo & C. H. Rábago-Quiroz, 2013. Feeding of the scyphomedusa Stomolophus meleagris in the coastal lagoon Las Guásimas, northwest Mexico. Hidrobiológica 23: 218–226.
Perrin, A., E. Le Bihan & N. Koueta, 2004. Experimental study of enriched frozen diet on digestive enzymes and growth of juvenile cuttlefish Sepia officinalis L. (Mollusca: Cephalopoda). Journal of Experimental Marine Biology and Ecology 311: 267–285.
Purcell, J. E., W. M. Graham & H. Dumont, 2001. Jellyfish blooms: ecosystem and societal importance Developments in Hydrobiology. Kluwer Academic Press, Dordrecht.
Rotllant, G., F. J. Moyano, M. Andrés, M. Díaz, A. Estévez & E. Gisbert, 2008. Evaluation of fluorogenic substrates in the assessment of digestive enzymes in a decapod crustacean Maja brachydactyla larvae. Aquaculture 282: 90–96.
Samain, J. F., J. Moal, J. Y. Daniel, J. R. Le Coz & M. Jezequel, 1980. The digestive enzymes amylase and trypsin during the development of Artemia: effect of food conditions. In Persoone, G., P. Sorgeloos, O. Roels & E. Jaspers (eds), The brine shrimp Artemia, Vol. 2. Universa Press, Wetteren: 427–443.
Shick, J. M., 1975. Uptake and utilization of dissolved glycine by Aurelia aurita scyphistomae: temperature effects on the uptake process; nutritional role of dissolved amino acids. Biological Bulletin 148: 117–140.
Smith, H. G., 1937. Contribution to the anatomy and physiology of Cassiopea frondosa. Papers from Tortugas laboratory of the Carnegie Institution of Washington 31:17–52.
Southward, A. J., 1955. Observations on the ciliary currents of the jellyfish Aurelia aurita L. Journal of the Marine Biological Association of the UK 34: 201–216.
Stewart, B. C. & S. Lakshmanan, 1975. Some properties of the acid phosphatase and the alkaline phosphatase of the summer jellyfish of the chesapeake bay, Chrysaora quinquecirrha desor. Comparative Biochemistry and Physiology Part A 50: 319–326.
Toledo-Cuevas, E. M., F. J. Moyano, D. Tovar-Ramírez, C. A. Strüssmann, C. A. Álvarez-González, C. C. Martínez-Chávez & C. A. Martínez-Palacios, 2011. Development of digestive biochemistry in the initial stages of three cultured Atherinopsids. Aquaculture Research 42: 776–786.
Van Praët, M., 1976. Acid phosphatase activities in Actinia equina L. and Cereus pedunculatus P. Bulletin de la Societé zoologique de France 101: 367–376.
Van Wormhoudt, A. & D. Sellos, 1980. Aspects biochimiques de la croissance: acides nucléiques et enzymes digestives chez Palaemon serratus (Crustacea Natantia). Oceanologica Acta 3: 97–105.
Vega-Villasante, F., H. Nolasco & R. Civera, 1993. The digestive enzymes of the Pacific brown shrimp Penaeus californiensis. 1. Properties of amylase activity in the digestive tract. Comparative Biochemistry and Physiology Part B 106: 547–550.
Villanueva, R., N. Koueta, J. Riba & E. Boucaud-Camou, 2002. Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding, using Artemia nauplii and compound diets. Aquaculture 205: 269–286.
Acknowledgements
The authors are grateful to Patricia Hinojosa Baltazar for enzyme analysis; Mónica Reza for maintenance and separation of organisms; Jorge Angulo for field assistance; Enrique Morales-Bojórquez and Emigdio Marin-Enriquez for statistics and R programming at CIBNOR. M.G.V. is a recipient of a fellowship from CONACYT, Mexico (Grant 376855).
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Juan Carlos Molinero
Rights and permissions
About this article

Cite this article
González-Valdovinos, M., Ocampo, L. & Tovar-Ramírez, D. Evaluation of digestive capacity in the polyp, ephyrae, and medusae stages of the cannonball jellyfish Stomolophus meleagris. Hydrobiologia 828, 259–269 (2019). https://doi.org/10.1007/s10750-018-3817-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-018-3817-3