
Abstract
Since 2013, the pelagic zone of Upper Lake Constance (ULC) has been subject to a massive invasion of the non-native three-spined stickleback (Gasterosteus aculeatus Linnaeus, 1758). Data from monthly monitoring of pelagic whitefish (Coregonus wartmanni Bloch, 1784) were used to compare weight-at-age and abundance of pelagic whitefish for years before (1997–2012) and after the invasion (2013–2015). Growth and abundance of pelagic whitefish is shown to be heavily influenced by stickleback presence. Mean autumn weight-at-age of whitefish decreased by 33.3% after the invasion took place and a significant decline in autumn CPUE in otherwise unfished cohorts of the population was also recorded. The results imply direct effects of stickleback presence on pelagic whitefish, including interspecific competition for food leading to reduced growth and survival, and predation of eggs and larvae, hampering recruitment. These observations coincide with a sharp decline in whitefish yield. In conclusion, this study shows that the invasion of stickleback has substantially altered the pelagic fish community of ULC, with severe consequences for commercial fisheries.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Many lakes in the alpine region have experienced drastic changes in nutrient profile over recent years. Anthropogenic eutrophication of these naturally oligotrophic systems has been followed in many cases by managed re-oligotrophication (Jeppesen et al., 2005; Gerdeaux et al., 2006), and with consequent changes in fish community composition. Furthermore, the native fish communities of all lakes are subject to the impacts of climate change (Jeppesen et al., 2014; Winfield et al., 2016) and aquatic invasive species (AIS). Striking examples of the threat from invasive species in large lakes are those of the sea lamprey (Petromyzon marinus L. 1758) and alewife (Alosa pseudoharengus Wilson 1811) in the Great Lakes of North America, the arrival of which combined with overfishing causes a massive reduction in numbers of predatory salmonids and the collapse and ultimate disappearance of most deepwater ciscoe species (Brown et al., 1987; Zimmermann & Krueger, 2009). The three-spined stickleback (Gasterosteus aculeatus) is one of several AIS to have emerged in Upper Lake Constance (ULC). The species first established in ULC between 1940 and 1950, but recently expanded from an exclusively shoreline habitat into the pelagic zone (Alexander et al., 2016).
A quantitative measure of the stickleback population in the pelagic zone of ULC was provided by a large fishing survey in September 2014 (Alexander et al., 2016). At this time, sticklebacks made up more than 95% of fish abundance and about 28% of fish biomass in the pelagic zone and this population persists to date (Fisheries Research Station, unpublished data). The situation exhibits similarities with that in the Baltic Sea, where recent drastic increases in populations of the same stickleback species have coincided with a devastating decline in recruitment of pike and perch (Bergström et al., 2015). The whitefish yield of the professional fisheries in ULC has declined in recent years in line with managed re-oligotrophication (Rösch, 2014; Baer et al., 2016). However, the further steep decline of 40% recorded between 2014 and 2015 (www.ibkf.org) takes the yield well below that previously recorded in the original pre-eutrophication state and thus cannot be explained by reduced productivity alone. One possible explanation for this further and unexpected decrease, especially with respect to the pelagic whitefish (Coregonus wartmanni), is the combined effect of competition for food and predation associated with the emergence of invasive sticklebacks in the pelagic zone. Data from long-term monitoring on growth and relative abundance of pelagic whitefish (www.ibkf.org) were analysed in an effort to isolate possible stickleback invasion effects on growth and relative abundance of whitefish. Background age class composition, catch per unit effort (CPUE) and body weight at catch of C. wartmanni were compared for the periods before and after stickleback invasion.
Material and methods
Study area
Lake Constance is situated between Austria, Germany and Switzerland. It has a surface area of 536 km2, of which 472 km2 belongs to the Upper Lake and 63 km2 to the much smaller Lower Lake (LLC). It is part of the Rhine drainage basin. This study used data from ULC only. ULC has undergone intensive re-oligotrophication in recent years. The fish community comprises a minimum of 30 species (Eckmann & Rösch, 1998), of which about 10 are targeted by professional fishermen (Rösch, 2014). Of these, whitefish (Coregonus spp.) are the most economically important, and fisheries management is based on routine monitoring of this important group (www.ibkf.org). An overview of the fisheries situation is given by Baer et al. (2016).
Sampling
In total, data were available from 1877 monitoring catches in the pelagic zone of ULC between 1991 and 2015. The occurrence of sticklebacks as bycatch in these nets was evaluated.
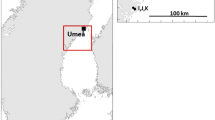
Monthly monitoring involves the setting of pelagic gillnets at three locations offshore from the towns of Langenargen, Lindau and Romanshorn (Fig. 1). The thread size of all nets is 0.12 mm. Monitoring nets were deployed in the mesh sizes routinely used by professional fishermen (38 and 40 mm) as well as smaller sizes (26, 32 and 36 mm) intended to sample the unexploited component of the whitefish population. The nets are set overnight at depths selected based on the experience of professional fishermen and on hydroacoustic surveys. From April to December, they are free floating, and from January to March they are fixed. The catch of each net is recorded according to the number of fish per species. According to IBKF (Internationale Bevollmächtigtenkonferenz für die Bodenseefischerei) definition, all whitefish caught in open water in ULC are presumed to be the pelagic type, C. wartmanni. Total body length (to the nearest 0.5 cm), body wet weight (to the nearest g) and sex are determined from a minimum of 20 whitefish from every net and scales are taken for age determination.

Map showing locations of sampling conducted in Upper Lake Constance: a Langenargen, b Nonnenhorn and c Romanshorn
Calculations of mean weight-at-age (MW) used only data from September to October, this being the time when 1+ fish first attain a size that can be captured in the gill nets deployed. The current analysis uses data from pelagic whitefish aged from 1.5 years (1+) to 4.5 years (4+) caught between 2001 and 2015. In order to eliminate examiner-based differences in age determinations between the three stations, the dataset from one location (Langenargen, where one trained person had performed the determination since 2001) was chosen for all statistical interrogation. In 2013 and 2014, autumn records were supplemented with data from November monitoring events to compensate for a lack of 1+ fish in the September or October catches.
Data treatment
Trends in stickleback occurrence in net fisheries between 1991 and 2015 were tested one-sided by the exact Cochran trend test.
The effect of stickleback occurrence on the age composition of the whitefish population was tested using an ordinal logistic model in the following form:
where j goes from age class 1 to age class 4 minus 1, β 0 is a constant, Xi is the vector of independent variables and β 0 is the parameter estimate for the ith independent variable with year, stickleback occurrence and net type as the independent variables.
Mean whitefish body weight in years with and without sticklebacks was compared using the following general linear model (GLM) (Sachs, 1997; Underwood, 1997):
where Y ijklm is the weight, µ denotes the year, α i is the age, γ k is the month of catch, δ l is the occurrence of sticklebacks [Yes/No], (µδ)l denotes the interaction between year and stickleback occurrence, (µα)j denotes the interaction between year and age class and ε ijklm is the random residual error.
Age 1+ whitefish in ULC are usually first caught in autumn, so this is also the time when highest abundances are usually reported in the unfished cohort. The datasets from September to October of each year were incorporated into a general linear model (GLM) (Sachs, 1997; Underwood, 1997) to compare years with and without sticklebacks. A Box-Cox transformation of the CPUE was necessary due to residual deviation from model assumptions:
where Y ijkl is the CPUE, µ denotes the year, α i is the mesh size, β j is the occurrence of sticklebacks [Yes/No], (αβ)jk is the interaction between the two effects and ε ijkl is the random residual error.
All data were analysed using JMP Pro (SAS Institute Inc.) version 13.1.0.
Results
Occurrence of sticklebacks
Sticklebacks were documented as bycatch in 2.9% (n = 54) of all analysed net catches (n = 1835), with between 1 and 76 individuals per net. Between 1991 and 2012, not one stickleback was noticed in 1500 nets (Fig. 2). The first stickleback bycatch was recorded in 2013, after which the proportion of sticklebacks in bycatch records increased significantly (P = 0.0001). In 2013, 13 of 102 nets (12.7%) contained sticklebacks, in 2014 the figure was 15 out of 109 nets (13.8%) and in 2015 it was 26 out of 124 nets (21.0%) (Fig. 2). Based on these results, analyses of mean body weight and CPUE were carried out on data divided into years before (2012 and earlier) and after (2013–2015) the emergence of stickleback.

Percentage of net fisheries with sticklebacks in the bycatch (grey columns) per year between 1991 and 2015; the number of net fisheries per year is written above the columns
Age class composition
The unfished cohort of the whitefish population comprised fish of age up to 4+ . Age 1+ whitefish were almost exclusively caught in 26-mm nets (Fig. 3). Before the stickleback invasion, 30.8 ± 2.6% (±standard error SE) of the catch in 26-mm monitoring nets was composed of 1+ whitefish, whereas after the arrival of sticklebacks in 2013 and 2014, no 1+ whitefish were recorded during September and October and in 2015 the 1+ age class was represented by just 3 individuals (6.5% of total catch; Fig. 3). In total, before the stickleback invasion around 11% of the catch comprised 1+ whitefish, and after the stickleback invasion below 1% (Table 1). Age 2+ fish were caught mainly in 32-mm mesh (Fig. 3) and accounted for 37.3 ± 3.1% (±SE) of total catch in all mesh sizes before the arrival of sticklebacks (Table 1). This value in all three mesh sizes fell to 24.7 ± 7.3% (±SE) when sticklebacks were present (Table 1). Small numbers of 2+ fish were also caught in nets with a mesh size of 36 mm in most years, but 2013 was the first year with no 2+ individuals in the 36-mm nets (Fig. 3). Age 3+ and 4+ fish were mainly caught in the 32- and 36-mm nets (Fig. 3). The logistic model shows that the catch data were significantly affected by all three independent variables, year (P = 0.0013), stickleback occurrence (P < 0.001) and mesh size (P < 0.001) (Table 1).

Age class composition of whitefish catches in three mesh sizes (26, 32 and 36 mm) during autumn (September, October) for the period before (1997–2012) and after stickleback appearance (2013–2015)
Body weight-at-age
For the period 2001–2012 (before stickleback appearance), the mean September/October weight (MW) of age 1+ to 4+ whitefish varied considerably between years (Fig. 4). For age classes 3+ and 4+, a significant (P < 0.05) negative trend was observed over the course of the years (Fig. 4). After the appearance of sticklebacks, a significant (P < 0.05, Table 2) drop in MW was observed for every age class (Fig. 4).

Mean wet weight (with standard deviation) of whitefish of different year classes (1+: above left, 2+: above right, 3+: blow left, 4+: below right) in years with (black squares) and without (open circles) sticklebacks
The model revealed that stickleback occurrence has a significant negative influence (P < 0.05) on MW of whitefish (Table 2). No influence of season (month) was observed (P > 0.05) (Table 1). As expected, differences between age classes and age class combined with year were significant (Table 2). The model also revealed an overall difference of 33.3% in mean body weights between years with and without sticklebacks (range over age classes 17.4–50.4%). MW for 1+ whitefish in years with sticklebacks averaged around 39.0 g less than in earlier years, while for older classes the discrepancies averaged 58.4 g for 2+ fish, 54.7 g for 3+ fish and 59.4 g for 4+ fish (Table 2).
CPUE
A negative trend in autumn CPUE values was observed for all mesh sizes for the period 1997–2012 (Fig. 5), but high variability meant that the trend could only be confirmed statistically for the 36 mm mesh size (P < 0.0001, analysis of variance). A sharp but not statistically significant decline in CPUE was observed for every mesh size in the period 2013–2015, when sticklebacks were present (P > 0.05, analysis of variance; Fig. 5, Table 3). Overall, the model (n = 335, R 2 adjusted = 0.27) revealed that for each mesh size category both year (P < 0.0001, GLM) and the interaction of year and stickleback presence had a strong negative influence (P = 0.012) on CPUE (Table 3).

Indirect measurement of whitefish abundance as CPUE (n per 100 m2 net per night) during autumn catches at the mesh sizes of 26 mm (a), 32 mm (b) and 36 mm (c) in years with sticklebacks (black squares) and without sticklebacks (open circles)
Discussion
The routine species monitoring that forms the basis of fishery management in ULC suggests that both weight-at-age and CPUE for pelagic whitefish exhibit an abrupt and significant downward trend from 2012 to 2013, and autumn-specific weight-at-age and CPUE were both significantly lower in years with sticklebacks present than in previous years. Such declines have previously been linked to a variety of factors, including the effects of climate change (Jeppesen et al., 2014; Winfield et al., 2016), trophic change (Gerdeaux et al., 2006) and changes in fisheries management (Kuparinen & Merilä, 2007). However, the effects of such drivers are normally rather subtle or enacted over the long term. The changes observed here indicate a sudden and rather drastic event in the lake, and the invasion of the pelagic zone of ULC by neozoic sticklebacks is a candidate.
Sticklebacks were first detected in ULC about 80 years ago (Nümann, 1972). A recent analysis suggesting that the species may have colonized the Lake Constance region via the upper Danube 2000–4000 years ago (Roesti et al., 2015; Marques et al., 2016) is not supported by historic (pre 20th century) fish survey data which record all the small-sized fish species now regarded as native in ULC without any mention of sticklebacks (Zeheter, 2015; Roch et al., submitted). Debate over the origin of the species in ULC notwithstanding, no sticklebacks were detected in the pelagic zone of ULC in the last 80 years, with the exception of a single short-lived occurrence in the 1970s, when the lake was eutrophic (Nümann, 1972). The recent emergence and persistence of sticklebacks in the pelagic zone of ULC is thus a new and apparently unprecedented phenomenon, made all the more remarkable by its suddenness and scale. The new population was first recorded as bycatch in 2013, when it was already a significant presence, and by autumn 2014 more than 95% of all pelagic fish recorded was a stickleback, making up 28% of recorded fish biomass (Alexander et al., 2016). Monitoring effort in the years immediately before 2013 was consistent with that from 2013 onwards, strongly suggesting the stickleback invasion of the pelagic zone took place in that year. Sticklebacks remain the significantly dominant species in the pelagic zone to this day (2017).
The stickleback invasion can be expected to have two direct effects on whitefish in ULC. Firstly, the newcomers represent direct competition for Daphnia, the principle food resource in the pelagic zone (Straile & Geller, 1998; Stich & Brinker, 2010; Roch et al., submitted). The sharp decrease in weight-at-age of whitefish recorded after the occurrence of sticklebacks may thus be indicative of reduced growth resulting from decreased food availability. Fish possessing smaller energy reserves are also more prone to natural mortality, especially when challenged by winter conditions (Hurst, 2007) or disease (Houston et al., 2007). The second major likely effect of invasion is that of predation, and a resulting negative impact on recruitment and year class strength, and thus abundance. Sticklebacks are commonly characterized as generalist feeders, foraging on zooplankton and benthic food (Hynes, 1950). They frequently consume eggs and juveniles of their own species (Smith & Reay, 1991; Manica, 2002; Mehlis et al., 2010) and prey on larvae of other fish species (Manzer, 1976; Delbeek & Williams, 1988; Kean-Howie et al., 1988; Gotceitas & Brown, 1993; Byström et al., 2015). A recent study by Roch et al. (submitted) indicates that Lake Constance sticklebacks are able to prey effectively on whitefish larvae in aquarium experiments. In addition, the whitefish larvae of ULC exhibit almost no predator avoidance behaviour (Roch et al., submitted). A survey close to a stocking site for hatchery-reared whitefish in ULC during spring 2016 revealed that the stomachs of sticklebacks contained massive numbers of whitefish larvae (Roch et al., submitted). The present study suggests that the relative abundance of 1+ whitefish during autumn (measured as CPUE) decreased drastically following the appearance of sticklebacks, especially in the 26-mm nets. Thus, a direct recruitment suppressing the effect of stickleback predation seems highly possible. Less explicable, however, is the decreasing relative abundance of larger whitefish, caught in nets with the mesh sizes of 32 and 36 mm and the high proportions of older fish (3+ to 4+) recorded in the first year of the stickleback invasion. Possible scenarios include the aforementioned increase in mortality due to strong interspecific competition for food or a change in whitefish behaviour resulting from decreasing food availability. It is possible that during autumn whitefish are opting to swim less or to occupy deeper, colder water layers to save energy. Both behaviours could lead to reduced catches in passive fishing gill nets, and an earlier study by Thomas et al. (2010) showed that in years with low food supply, ULC whitefish were harvested from greater depths, where water temperatures were up to Δ 4°C lower. A shift in whitefish distribution towards deeper and colder water might be a bioenergetic optimization strategy. Similar shifts are observed in other salmonids, including the Atlantic salmon Salmo salar L. 1758, in which starving individuals move to cooler (deeper) waters to reduce metabolic costs (Javaid & Anderson, 1967). Both explanations appear superficially plausible, but data are not available to support or refuse these hypotheses. Sticklebacks have caused problems for fish and fisheries in other waters. A recent massive increase in stickleback abundance in the Baltic Sea (Bergström et al., 2015; Byström et al., 2015) has coincided with a drastic decline in perch (Perca fluviatilis L. 1758) and pike (Esox lucius L. 1758) recruitment. Perch and pike are significant nearshore spawning species in ULC, alongside roach (Rutilus rutilus L. 1758) and pikeperch (Sander lucioperca L. 1758). As a highly valued and intensively fished species, perch stocks are routinely monitored and the 2015 yield was one of the lowest since records began (IBKF, 2016). Recruitment failure in this species is expected to impact yield with a 2- to 3-year delay, and thus the extremely low yield in 2015 could be a result of larval predation by newly emergent sticklebacks in 2013. In the case of other shore spawning species such as roach and pike, lower fishing intensity than for whitefish and perch (IBKF, 2016) means that commercial yield does not represent stock size or recruitment. In fact, occasional inspection of pike stomachs suggests that they prey successfully on stickleback and pike yield in ULC has increased in recent years (www.ibkf.org).
Long-term datasets from the professional fisheries suggest that in its former oligotrophic state ULC yielded an annual whitefish catch in the region of 300 metric tons (mt) (1910–1955 figures, mean yield ± standard deviation 289 ± 100 mt) (Baer et al., 2016). These values lie within the productivity range reported from other prealpine oligotrophic lakes (Müller et al., 2007). However, the recent decline in whitefish yield from 293 mt in 2013 to 152 mt in 2015 took place with no change in trophic state (www.ibkf.org), meaning that this exceptionally low yield cannot be explained by lake productivity alone.
In conclusion, the recent steep decline in whitefish weight-at-age and yield in ULC appear to be closely associated with the invasion of AIS sticklebacks and is likely via interspecific competition for food and a possible recruitment effect. This adds to the existing severe pressures on lake productivity resulting from trophic changes, and the potentially serious effects of transportation, tourism, heavy use of productive shallow water zones and the negative impacts of climate change (Straile et al., 2007; Stich & Brinker, 2010; Wahl & Peeters, 2014). Further data from the commercial fisheries suggest a negative influence on other species too, with perch yield at historically low levels, and yields of Arctic charr (Salvelinus sp) and burbot (Lota lota L. 1758) collapsing (IBKF, 2016).
The likely future of stickleback dominance in ULC is unclear. Elsewhere, the progress of invasive stickleback population has been controlled and finally brought to collapse by high prevalence of the tapeworm Schistocephalus solidus Müller 1776 (Heins et al., 2010), and it is possible that a similar story may unfold in ULC. However, monitoring of the parasite loads of ULC sticklebacks suggests that the prevalence of infection with S. solidus ranges between 15 and 60% (Gugele et al., unpublished data), well below the 80% that leads to population collapse, e.g. in Walby Lake, Alaska (Heins et al., 2010).
In the absence of any apparent decline in the stickleback population of ULC, active fisheries management options for control and containment are being considered. Possible actions might include intensified fishing (electric fishing, fyke nets, etc.) during the spawning season in tributaries, where pelagic stickleback may migrate to spawn in high numbers (Marques et al., 2016). Focusing action away from the lake itself may limit the risk of whitefish bycatch. Other options, such as the use of small mesh gill or trawl nets in the pelagic zone, are not considered viable because of the patchy distribution and mobility of the stickleback population (Gugele pers.com.). Furthermore, such actions would likely also impact whitefish juveniles sharing the same habitat, rendering them counterproductive. In an effort to solve this issue, the depth distributions of whitefish and stickleback are currently under investigation using experimental trawls and hydroacoustic surveys. Firm data differentiating species by depth may permit targeted trawling for stickleback in the pelagic zone without risking significant bycatch of juvenile whitefish.
Another management approach under consideration is to refine stocking practices to further support native fish. Stocking with whitefish larvae has been shown to boost stock levels when recruitment deficits exist (Wanke et al., 2016), and stocking with whitefish larvae in sizes exceeding the predatory gape of sticklebacks may stabilize future whitefish recruitment in ULC. Despite these possibilities, at present there is no direct or effective short-term solution to the problem for the commercial fishermen of ULC facing exceptionally low yields of whitefish and probably other species.
Furthermore, a convincing explanation is still lacking for the sudden and explosive invasion of the pelagic zone by sticklebacks, after 80 years of inconspicuous occupation of the littoral. Changing lake trophic state alone is insufficient to explain the phenomenon, given that in comparable pre-eutrophication conditions the species maintained an inconspicuous presence and kept away from the pelagial. The missing factor could be a new food component such as the invasive Limnomysis benedeni Czerniavsky 1882, which has been found in ULC in high densities for less than 10 years (Hanselmann, 2011). Another significant possible factor is the recent practice of massive stocking with artificially reared whitefish larvae, far exceeding anything attempted pre-eutrophication (Eckmann et al., 2007). This artificial abundance of small fish may support sticklebacks in the pelagic zone in early spring when crustacean food is still scarce and thus provide the species with a marked advantage over competing whitefish, which have no comparable starter feeding opportunity. It may also be that the current situation is exacerbated by a combination of factors including climate change (Wahl & Peeters, 2014) and invasion lag time, a common feature of invasive species dynamics (Sakai et al., 2001). Further studies are needed to shed light on the underlying mechanisms.
In conclusion, while this study does not explain the reason of the stickleback invasion, it highlights the potential for AIS to disrupt an entire existing food web and substantially alter existing fish communities with severe consequences for fishery yields.
References
Alexander, T. J., P. Vonlanthen, G. Périat, J.-C. Raymond, F. Degiorgi & O. Seehausen, 2016. Artenvielfalt und Zusammensetzung der Fischpopulation im Bodensee. Projet Lac, Eawag.
Baer, J., R. Eckmann, R. Rösch, R. Arlinghaus & A. Brinker, 2016. Managing Upper Lake Constance fishery in a multi-sector policy landscape: beneficiary and victim of a century of anthropogenic trophic change. In Song, A. M., S. D. Bower, P. Onyango, S. J. Cooke & R. Chuenpagdee (eds), Inter-Sectoral Governance of Inland Fisheries. TBTI Publication Series, Canada.
Bergström, U., J. Olsson, M. Casini, B. K. Eriksson, R. Fredriksson, H. Wennhage & M. Appelberg, 2015. Stickleback increase in the Baltic Sea – a thorny issue for coastal predatory fish. Estuarine Coastal Shelf Sciences 163: 134–142.
Brown Jr., E. H., R. N. Argyle, R. N. Payne & M. E. Holey, 1987. Yield and dynamics of destabilized chub (Coregonus spp.) populations in lakes Michigan and Huron. Canadian Journal of Fisheries and Aquatic Sciences 44: 371–383.
Byström, P., U. Bergström, A. Hjälten, S. Ståhl, D. Jonsson & J. Olsson, 2015. Declining coastal piscivore populations in the Baltic Sea: where and when do sticklebacks matter? AMBIO 44: 462–471.
Delbeek, J. C. & D. D. Williams, 1988. Feeding selectivity of four species of sympatric stickleback in brackish-water habitats in eastern Canada. Journal of Fish Biology 32: 41–62.
Eckmann, R. & R. Rösch, 1998. Lake Constance fisheries and fish ecology. Advances Limnology 53: 285–301.
Eckmann, R., M. Kugler & C. Ruhlé, 2007. Evaluating the success of large-scale whitefish stocking at Lake Constance. Advances Limnology 60: 361–368.
Gerdeaux, D., O. Anneville & D. Hefti, 2006. Fishery changes during the re-oligotrophication in 11 peri-alpine Swiss and French lakes over the past 30 years. Acta Oecologia 30: 161–167.
Gotceitas, V. & J. A. Brown, 1993. Risk of predation to fish larvae in the presence of alternative prey: effects of prey size and number. Marine Ecololgy Progress Series 98: 215.
Hanselmann, A. J., 2011. A review of spatio-temporal patterns of the colonisation of Lake Constance with alien Macrozoobenthos. Lauterbornia 72: 131–148.
Heins, D. C., E. L. Birden & J. A. Baker, 2010. Host mortality and variability in epizootics of Schistocephalus solidus infecting the threespine stickleback, Gasterosteus aculeatus. Parasitology 137: 1681–1686.
Houston, A. I., J. M. McNamara, Z. Barta & K. C. Klasing, 2007. The effect of energy reserves and food availability on optimal immune defence. Proceedings of the Royal Society B 274: 2835–2842.
Hurst, T. P., 2007. Causes and consequences of winter mortality in fishes. Journal of Fish Biology 71: 315–345.
Hynes, H. B. N., 1950. The food of fresh-water sticklebacks (Gasterosteus aculeatus and Pygosteus pungitius), with a review of methods used in studies of the food of fishes. Journal of Animal Ecology 19: 36–58.
IBKF, 2016. Internationale Bevollmächtigtenkonferenz für die Bodenseefischerei. [available on internet at http://www.ibkf.org/aktuelles/gesamtbericht/]. Access in June. 2016.
Javaid, M. Y. & J. M. Anderson, 1967. Thermal acclimation and temperature selection in Atlantic salmon, Salmo salar and rainbow trout, Salmo gairdneri. Journal of the Fisheries Research Board of Canada 93: 137–145.
Jeppesen, E., M. Sondergaard, J. P. Jensen, K. E. Havens, O. Anneville, L. Carvalho, M. F. Coveney, R. Deneke, M. T. Dokulil, B. Foy, D. Gerdeaux, S. E. Hampton, S. Hilt, K. Kangur, J. Köhler, E. H. H. R. Lammens, T. L. Lauridsen, M. Manca, M. R. Miracle, B. Moss, P. Noges, G. Persson, G. Phillips, R. Portielje, S. Romo, C. L. Schelske, D. Straile, I. Tatrai, E. Willen & M. Winder, 2005. Lake responses to reduced nutrient loading – an analysis of contemporary long-term data from 35 case studies. Freshwater Biology 5: 1747–1771.
Jeppesen, E., M. Meerhoff, T. A. Davidson, D. Trolle, M. Sondergaard, T. L. Lauridsen, M. Beklioglu, S. Brucet, P. Volta, I. Gonzales-Bergonzoni & A. Nielsen, 2014. Climate change impacts on lakes: an integrated ecological perspective based on a multi-faceted approach, with special focus on shallow lakes. Journal of Limnology 73: 88–111.
Kean-Howie, J. C., S. Pearre Jr. & L. M. Dickie, 1988. Experimental predation by sticklebacks on larval mackerel and protection of fish larvae by zooplankton alternative prey. Journal of Experimental Marine Biology and Ecology 124: 239–259.
Kuparinen, A. & J. Merilä, 2007. Detecting and managing fisheries induced evolution. Trends in Ecology and Evolution 22: 652–659.
Manica, A., 2002. Filial cannibalism in teleost fish. Biological Reviews of the Cambridge Philosophical Society 77: 261–277.
Manzer, J. I., 1976. Distribution, food, and feeding of the threespine stickleback, Gasterosteus aculeatus, in Great Central Lake, Vancouver Island, with comments on competition for food with juvenile sockey salmon, Oncorhynchus nerka. Fisheries Bulletin 74: 647–668.
Marques, D. A., K. Lucek, J. I. Meier, S. Mwaiko, C. E. Wagner, L. Excoffier & O. Seehausen, 2016. Genomics of rapid incipient speciation in sympatric threespine stickleback. PLOS Genetics 12: e1005887.
Mehlis, M., T. C. M. Bakker, L. Engqvist & J. G. Frommen, 2010. To eat or not to eat: egg-based assessment of paternity triggers fine-tuned decisions about filial cannibalism. Proceedings of the Royal Society B: Biological Sciences 277: 2627–2635.
Müller, R., M. Breitenstein, M. M. Bia, C. Rellstab & A. Kirchhofer, 2007. Bottom–up control of whitefish populations in ultra-oligotrophic Lake Brienz. Aquatic Sciences 69: 271–288.
Nümann, W., 1972. The Bodensee: effects of exploitation and eutrophication on the salmonid community. Journal of Fisheries Research Board Canada 29: 833–847.
Roch, S., L. von Ammon, J. Geist & A. Brinker, submitted. Foraging habits of invasive three-spined sticklebacks (Gasterosteus aculeatus) – impacts on fisheries yield in Upper Lake Constance.
Rösch, R., 2014. Lake Constance fish and fisheries. In Welcomme, R. L., J. Valbo-Jorgensen & A. S. Halls (eds) Inland Fisheries Evolution and Management – Case Studies from Four Continents, FAO Fisheries and Aquaculture Technical Paper 579: 21–31.
Roesti, M., B. Kueng, D. Moser & D. Berner, 2015. The genomics of ecological vicariance in threespine stickleback fish. Nature Communications 6: 8767.
Sachs, L., 1997. Angewandte Statistik. Springer, Berlin.
Sakai, A. K., F. W. Allendorf, J. S. Holt, D. M. Lodge, J. Molofsky, K. A. With, S. Baughman, R. J. Cabin, J. E. Cohen, N. C. Ellstrand, D. E. McCauley, P. O’Neil, I. M. Parker, J. N. Thompson & S. G. Weller, 2001. The population biology of invasive species. Annual Review of Ecology, Evolution, and Systematics 32: 305–332.
Smith, C. & P. Reay, 1991. Cannibalism in teleost fish. Reviews in Fish Biology and Fisheries 1: 41–64.
Stich, H. B. & A. Brinker, 2010. Oligotrophication outweighs effects of global warming in a large, deep, stratified lake ecosystem. Global Change Biology 16: 877–888.
Straile, D. & W. Geller, 1998. Crustacean zooplankton in Lake Constance from 1920 to 1995: response to eutrophication and re-oligotrophication. Advances Limnology 53: 255–274.
Straile, D., R. Eckmann, T. Jüngling, G. Thomas & H. Löffler, 2007. Influence of climatic variability on whitefish (Coregonus lavaretus) year-class strength in a deep, warm monomictic lake. Oecologia 151: 521–529.
Thomas, G., R. Rösch & R. Eckmann, 2010. Seasonal and long-term changes in fishing depth of Lake Constance whitefish. Fisheries Management and Ecology 17: 386–393.
Underwood, A. J., 1997. Experiments in Ecology – Their Logical Design and Interpretation Using Analysis of Variance. Cambridge University Press, Cambridge: 504.
Wahl, B. & F. Peeters, 2014. Effect of climatic changes on stratification and deep-water renewal in Lake Constance assessed by sensitivity studies with a 3D hydrodynamic model. Limnology and Oceanography 59: 1035–1052.
Wanke, T., U. Brämick & T. Mehner, 2016. Early detection of reproduction deficits and the compensatory potential of enhancement stocking for vendace, Coregonus albula, fisheries in German lakes. Fisheries Management and Ecology 22: 55–65.
Winfield, I. J., C. Baigún, A. Pavel, P. A. Balykin, B. Becker, Y. Chen, A. N. Filipe, Y. V. Gerasimov, A. L. Godinho, R. M. Hughes, J. D. Koehn, D. N. Kutsyn, V. Mendoza-Portillo, T. Oberdorff, A. M. Orlov, A. P. Pedchenko, P. Pletterbauer, I. G. Prado, R. Rösch & S. J. Vatland, 2016. International perspectives on the effects of climate change on inland fisheries. Fisheries 41: 399–405.
Zeheter, M., 2015. Order in the Lake: managing the sustainability of the Lake Constance fisheries, 1350–1900. Environment and History 21: 597–629.
Zimmermann, M. A. & C. C. Krueger, 2009. An ecosystem perspective on re-establishing native deepwater fishes in the Laurentian Great Lakes. North American Journal of Fisheries Management 29: 1352–1371.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: Nico Salmaso, Orlane Anneville, Dietmar Straile & Pierluigi Viaroli / Large and deep perialpine lakes: ecological functions and resource management
Rights and permissions
About this article

Cite this article
Rösch, R., Baer, J. & Brinker, A. Impact of the invasive three-spined stickleback (Gasterosteus aculeatus) on relative abundance and growth of native pelagic whitefish (Coregonus wartmanni) in Upper Lake Constance. Hydrobiologia 824, 243–254 (2018). https://doi.org/10.1007/s10750-017-3479-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-017-3479-6