
Abstract
We developed a simple method to culture two sessile rotifers, Limnias ceratophylli and Limnias melicerta, which should be applicable to other sessile species, and examined effect of the concentration of Chlorella vulgaris on population growth of these species. Limnias ceratophylli had higher population abundances at higher food levels. For both species, intrinsic rate of increase (r), derived from population growth study, varied from 0.12 to 0.16 day−1. Differences in r varied depending on food level for L. ceratophylli, but not for L. melicerta. Both species had little mortality during 2–3 weeks and thereafter survivorship declined until 5–7 weeks depending on food concentration. Mean life expectancy at birth for L. ceratophylli and L. melicerta was 29–34 days and 28–33 days, respectively. Generation time was shorter for L. ceratophylli. Gross and net reproductive rates were higher for L. ceratophylli. For both species, increase in food density resulted in significant decrease of average lifespan and life expectancy at birth. Gross reproductive rate and rate of population increase of both species were not significantly affected by food density. Generation time was significantly affected due to increase in algal food only for L. ceratophylli but not for L. melicerta.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
While we understand that temperature and food influence the population levels of planktonic rotifers (Edmondson, 1946; Galkovskaja, 1987; Miracle & Serra, 1989), we know little of the conditions that influence growth and reproduction of sessile species (Edmondson, 1945; Garcia, 2004). As with planktonic rotifers, food type no doubt plays an important role in population growth in the sessile taxa (King, 1967; Thor et al., 2002). Indeed, we do know that sessile taxa (e.g., Ptygura and Floscularia) appear to have food preferences, selecting certain algae, but not bacteria or yeast (Wallace & Starkweather, 1983, 1985; Wallace et al., 1998). When feeding on algae, most planktonic rotifers reach their population maxima in two to three weeks and thereafter begin to decline as the resources become limiting (Stemberger and Gilbert, 1985; Yoshinaga et al., 2001). We also know quite a bit about the conditions necessary to establish successful culture of plankton rotifers (e.g., Stemberger, 1981; Walz, 1987; Lubzens et al., 1989; Nandini et al., 2009; Xi et al., 2011; Yoshimatsu & Hossain, 2014). Unfortunately, there have been too few published studies of sessile rotifers to permit careful examination of the factors regulating their population growth in comparison to planktonic species.
Planktonic species have been cultured under a variety of conditions including in small (1 ml) (Walz, 1983) to very large (thousands of liters) (Yoshimatsu & Hossain, 2014) volumes. On the other hand, little information is available on culture methods for sessile species that have been removed from their natural habitat and in the absence of their natural substrata: e.g., Sinantherina socialis (Garcia, 2004). Additionally, it is not known whether the algal species commonly used to culture planktonic rotifers are adequate for sessile species. Having the ability to culture sessile rotifers in the laboratory would permit detailed analyses of their growth by employing life table techniques, a basic tool of demographic studies in rotifers (Nandini & Sarma, 2000; Wallace et al., 2006, 2015). For example, age-specific mortality and reproduction are best estimated using a cohort population and population maxima can be estimated using a population dynamics approach (Walz, 1993). In planktonic rotifers, these two methods usually are examined simultaneously (e.g., Sarma & Nandini, 2001). For sessile species, there are few such studies (e.g., Edmondson, 1945; Wallace & Edmondson, 1986). Here we report techniques to culture two sessile species Limnias ceratophylli Schrank, 1803 and Limnias melicerta Weisse, 1848 under laboratory conditions. In this study, we quantified effects of food concentration using Chlorella vulgaris Beijerinck, 1890 on the population growth and the life table demography of these rotifers. We hypothesized that higher food levels would result in higher survivorship, fecundity, and population growth rates in both species, and that there would be no significant differences in the demographic variables between both species considering that they have a similar body size.
Materials and methods
Sampling area
The sampling site for this study was the Ecological Reserve of Pedregal, San Angel (Mexico City). Covering >237.3 ha, this region has five lakes with a combined area of >11,900 m2. The area has five interconnected water bodies located at an altitude of 2270 m above sea level. During the summer months of 2013, we collected samples from the water bodies of the Ecological Reserve and isolated L. ceratophylli and L. melicerta.
Culture medium for sessile rotifer species
A few individuals of each species were separated from the macrophytes and placed into transparent jars containing 5 ml of the filtered pond water. These were fed green alga Chlorella vulgaris at low density (0.25 × 106 cells ml−1). After some days of growth, we established clonal populations of each species from this initial culture by removing one individual and transferring it to a new culture vessel. Chlorella vulgaris was batch-cultured using Bold’s basal medium (Borowitzka & Borowitzka, 1988) supplemented with sodium bicarbonate (0.5 g l−1) every third day. Algae in log phase was harvested by centrifugation at 4000 rpm for 5 min., rinsed, and re-suspended in distilled water to remove the algal culture medium which does not support rotifer growth. In all cases, algal density was determined by counting using a hemacytometer.
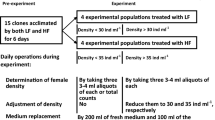
To develop an appropriate culture medium for our experiments, we conducted a preliminary growth study using L. ceratophylli and three different culture media: filtered lake water (FLW, from the water body from which the test rotifers were obtained), moderately hard water, EPA culture medium (EPA medium), and a mixture of both in equal proportions (50% FLW + 50% EPA). EPA medium was prepared each day dissolving 96 mg of NaHCO3, 60 mg of CaSO4, MgSO4 60 mg, and 4 mg of KCl in 1 l of distilled water (Weber, 1993; Lewis et al., 1994). From the clonal population, 50 individuals were placed into transparent glass vessels of 100 ml capacity containing 50 ml of the one of the three culture media. The experiments were conducted in 4 replicates at 22 ± 2 °C and with photoperiod of 12L:12D. Culture vessels received Chlorella at a density of 0.5 × 106 cells ml−1. The test media were replaced daily for 15 days after which all culture vessels received only EPA medium for another 5 days. Thereafter, the preliminary experiments were discontinued. Based on results from these trials, growth assessments and demographic evaluations were conducted for both species.
Population growth experiments
The experimental design for both species was similar to that in the preliminary tests except here we used a starting population of five individuals in 30 ml (about 0.167 ind. ml−1) EPA medium with the algal food concentrations at three levels (low: 0.25 × 106, medium: 0.5 × 106 and high 1.0 × 106 cells ml−1 of C. vulgaris). For each food concentration, we set up 4 replicates. As in the preliminary tests, the medium was replaced daily. The experiment was stopped after 35 days by which time rotifers in most treatments began to enter the stabilization phase. The rate of population increase (r) was derived using the exponential equation:
where N 0 = initial population density; N t = population density after time t; and t = time in days. In treatments where a peak of population abundance was not evident, the growth rate was calculated from the slope between ln N and time (Sibly & Hone, 2002).
Life table demography experiments
Life table demography experiments were conducted for each species by employing an experimental design similar to the population growth studies except here we used neonates (mean age: 3 h). On a daily basis, we counted and removed offspring and dead adults. The experiments were discontinued when the original cohort died in each replicate.
Data from the life table demography were used to derive life expectancy at birth (LEB, e 0), average of lifespan (ALS), gross reproductive rate (GRR), net reproductive rate (\(R_{0}\)), generation time (T), and population growth rate (r). The following equations described by Krebs (1985) were used to calculate these life table metrics.
where L x = number of individuals alive on the average during the age interval x to x + 1; Tx = cumulative number of individuals at age x; n x = number of living individuals at the age x; l x = proportion of surviving to start of age x; and m x = offspring produced per female at age x. The population growth rate was derived iteratively and the confidence intervals were obtained using a jackknife method (Meyer et al., 1986). We used one-way analysis of variance (ANOVA) and for multiple comparison, post hoc (Tukey) tests, to evaluate the differences in growth rates and other life history variables of the tested rotifer species.
Results
Effect of culture medium
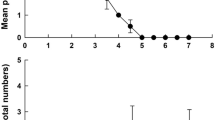
Both species easily adapted to the culture media. Limnias ceratophylli showed similar growth patterns on all the test media (EPA, EPA + FLW and FLW) up to day 12. When these populations were transferred to test vessels containing only the EPA medium, the populations increased from day 13 onwards, especially for the treatment initially involved filtered lake water and mixed with lake water + EPA medium. EPA medium alone did not result in higher population growth (Fig. 1). Therefore, all further experiments were conducted using individuals of the monoclonal population initially adapted to mixed filtered lake water + EPA medium. Subsequent experiments were conducted using only EPA medium.

Population growth of Limnias ceratophylli with different culture media, the vertical line indicates the beginning of use of EPA medium in all treatments. The means and standard errors (SE) are based on 3 replicates. In some cases, the SE bars are obscured by the symbols
Population growth
Population growth patterns of L. ceratophylli and L. melicerta were affected by algal concentration; both species had higher population abundances at the highest tested food level (Fig. 2). The lowest food density had resulted in lower population growth of L. ceratophylli. However for L. melicerta, the growth curves at the two higher algal densities were similar and food density-related differences were not evident. Growth curves of L. ceratophylli reached equilibrium after 3 weeks in different concentrations of food; however, it was observed that at a concentration of 1 × 106 cells ml−1, both species reached their highest densities.

Population growth of L. ceratophylli and L. melicerta at different concentrations of Chlorella vulgaris (X 106 cells ml−1). The initial density was 0.167 ind. ml−1. Values represent the mean and standard error based on 4 replicates
An increase in food availability resulted in a significant increase in the peak population density of both species. This was evident in L. ceratophylli where the algal concentration had significant effect on the peak population density (threefold increase at 1 × 106 cells as compared to that at 0.25 × 106 cells/ml) (one-way ANOVA, F = 14.08, df = 2, P < 0.05).
Population growth rate (r) followed a similar trend to that observed in the maximum densities; in L. ceratophylli, the r ranged from 0.12 to 0.16 per day and in L. melicerta it varied little (0.14–0.15 per day). There was a statistically significant increase in r of both species with increase in food availability (one-way ANOVA, F = 43.69, 12.35, df = 2, P < 0.001, respectively).
Life table demography
Both species had little mortality during weeks 2–3 and thereafter the survivorship gradually declined until weeks 5–7 depending on the food concentration. At a given food density L. melicerta had little mortality for the first 3 weeks when compared to L. ceratophylli. For both the rotifer species, increase in food concentration significantly decreased survival (one-way ANOVA, F = 19.44, 9.51, df = 2, P < 0.01, respectively) (Fig. 3).

Age-specific survivorship (closed circles) and fecundity (open circles) curves of L. ceratophylli and L. melicerta cultured under different concentrations of C. vulgaris (×106 cells ml−1). The mean values and standard error based on 4 replicates are shown
The age-specific fecundity curves of both species showed some significant differences in the pattern of offspring production. Reproduction started on the 3rd day for L. ceratophylli and on the 5th day for L. melicerta. Food density-related differences were evident for L. ceratophylli; at the higher food levels this species had higher offspring production. However, for L. melicerta effect of food density on offspring production was not evident. In addition, there was a distinct peak of offspring production in L. ceratophylli under the three food densities, while under comparable conditions the neonate production in L. melicerta was nearly continuous during the first two weeks of reproduction. Regardless of food density, the peak offspring production for both the rotifer species was observed between 7 and 21 days (Fig. 3).
Data on the life history variables of both L. ceratophylli and L. melicerta in relation to food density are presented in Table 1. At any given food density, average lifespan (23–29 days) and life expectancy at birth (23–28 days) of L. ceratophylli were slightly shorter than those of L. melicerta (29–34 days and 28–33 days, respectively). Generation time was also shorter for L. ceratophylli than for L. melicerta. However, both gross and net reproduction rates were higher for L. ceratophylli as compared to L. melicerta. The intrinsic rate of increase (r) was higher (0.362–0.383 per day) for L. ceratophylli as compared to that in L. melicerta (0.292–0.304 per day). For both species, increase in food density resulted in a significant decrease of average lifespan (one-way ANOVA, F = 30.97, df = 2, P < 0.001), life expectancy at birth (F = 30.96, df = 2, P < 0.001), and \(R_{0}\) (F = 6.95, df = 2, P < 0.05). Gross reproductive rate of both the rotifer species was not significantly affected by food density (F = 0.71, df = 2, P > 0.05). Generation time was significantly (Tukey test, df = 2, P < 0.05) affected due to increase in algal food only for L. ceratophylli, but not for L. melicerta. The r of both the rotifer species was significantly influenced by the offered food density (one-way ANOVA, F = 15.88, 13.77, df = 2, P < 0.001, respectively) (Table 1).
Discussion
Here, we demonstrate that sessile rotifers may be easily cultured using EPA medium, a common culture medium for many other species of zooplankton (Weber, 1993; Nandini & Sarma, 2000). Our data showed that mixing the EPA medium with filtered lake water was appropriate for an initial acclimation period of a few weeks. Thereafter, favorable growth was achieved when the population was transferred to EPA medium. Thus, it appears that sessile rotifers can adapt rapidly from the natural waters to synthetic medium (Wallace et al., 2006).
Flosculariid rotifers possess malleoramate trophi, which are able to crush food particles of up to a size of ~20 µm (Wallace & Starkweather, 1983; Monakov, 2003). In our study, we offered Chlorella (5 µm), a food item well below this size, and both species thrived.
Sessile rotifers are normally attached to a variety of substrata including algae, macrophytes, rocks, and floating twigs, and there are ample data to indicate that sessile rotifers have substrate specificity (e.g., Edmondson, 1944; Wallace, 1978; Wallace & Edmondson, 1986; Kuczyńska-Kippen, 2007; Wallace & Smith, 2013; Meksuwan et al., 2014). However, our data showed sessile species can be successfully cultured without the presence of their natural substrata. In culture jars L. ceratophylli and L. melicerta attached mainly on the bottom, but also to the sides.
Although food and feeding habits of Limnias are not well-known, field observations suggest that yeast is also consumed by the species of this genus (Wallace & Starkweather, 1983). Sessile rotifers live in the littoral zone attached to macrophytes (Meksuwan et al., 2014); their food is composed of planktonic algae such as Chlorella, Coelastrum, Scenedesmus, and Pediastrum (Nandini et al., 2005; Enríquez-García et al., 2009). Thus, it is not surprising that the food we provided was sufficient to permit good growth of these sessile rotifers.
The populations of both L. ceratophylli and L. melicerta continued to grow after an initial lag phase of about 7–10 days and the population growth was stabilized at approximately after four weeks in nearly all the offered algal concentrations. This trend is different in planktonic rotifers such as species of Anuraeopsis, Brachionus, and Plationus where the populations typically have a very short lag phase (2–5 days) reach a peak within 2 weeks and thereafter nearly decline in density (Dumont et al., 1995). On the other hand, non-planktonic rotifers of the genera Lecane and Euchlanis have a long lag phase (>1 week) and reach the stabilization phase after 3–4 weeks (Sarma et al., 2006). The trend with sessile rotifers is yet another instance where both the lag phase and the stabilization phase are long. This suggests that for a complete population cycle sessile rotifers need much longer time that either planktonic or periphytic species (Sarma et al., 2006; Espinosa-Rodríguez et al., 2012). In addition, during the course of our experiments sexual reproduction (presence of males and/or diapausing embryo production) was not observed, suggesting that the population consisted entirely of parthenogenetic females.
The maximum density reached by zooplankton typically depends on both the body size of the animal and the quantity of food available to it (Stemberger & Gilbert, 1985; Duarte et al., 1987). Smaller species reach higher abundances in comparison to larger species when cultured under comparable test conditions. For example, Anuraeopsis and Lepadella usually reach peak densities of >1000 ind. ml−1, while larger species such as Brachionus calyciflorus Pallas, 1766 may reach to about 100 ind. ml−1 (Dumont et al., 1995; Nandini et al., 2007). In the present study, both the species had relatively lower peak population densities (10–40 ind. ml−1) possibly due to their larger body sizes.
The intrinsic rate of increase (r) for most rotifers reach values that range from 0.5 to 1.5 day−1. Values lower than 0.5 day−1 are common in periphytic and benthic taxa, but values higher than 2 day−1 are rare, and have been reported for only a few species: B. calyciflorus and Brachionus plicatilis Müller, 1786, and Asplanchna sieboldii (Leydig, 1854) (Wallace et al., 2006). In this study r was much lower (0.01–0.38 day−1), which may be related to their life history. Non-planktonic and sessile rotifers seem to have lower growth rates compared to planktonic species. For example, under optimal food conditions, the non-planktonic species Lecane inermis (Bryce, 1892) and Lepadella rhomboides (Gosse, 1886) have growth rates >0.4 day−1 (Sarma et al., 2006).
Cohort life table studies yield information about age-specific survival and reproduction. In his study of a natural population of Floscularia conifera (Hudson, 1886), Edmondson (1945) found that the population experienced more than 50% mortality by the time the animals attained age 7 days and all the individuals died within a week thereafter. The survival curves of our laboratory populations of L. ceratophylli and L. melicerta had a rectangular shape—there was practically no mortality during the first two weeks and thereafter the population experienced gradual mortality. In addition, effect of food density on this pattern suggests that lowest food density resulted in almost no mortality during the first two weeks of age. This result supports research that indicates that food restriction enhances the duration of life (Sawada & Enesco, 1984; Snell et al., 2015). The statistical analysis of the average lifespan and life expectancy at birth also shows that food density had significant effect on the lifespan of both species.
Increase in food concentration significantly reduced the lifespan and \(R_{0}\) in these species. However, gross reproductive rate and r were not affected. Most planktonic rotifers show increased gross reproductive rate and \(R_{0}\), as well as in r per day with increase in food density (Sarma & Rao, 1991). However, in our study, both species showed lower output of embryos with increase in food density. It is possible that sessile species are most susceptible to higher levels of algae that may have adverse impact on their filter feeding. Certain species of planktonic rotifers (e.g., Brachionus variabilis Hempel, 1896) are also sensitive to increased food levels, which led to decreased survival and reproduction (Sarma & Nandini, 2001). The relationship between mean lifespan and T has received considerable attention in zooplankton research. King (1982) hypothesized that the mean lifespan of iteroparous species cultured under optimal conditions is twice the generation time. Indeed we also found a positive, significant relation between these two life history variables. However, the ratio was not 2× as King hypothesized, but was much less (1.03) suggesting a deviation as also reported in Sarma & Rao (1991).
Conclusions
It is difficult to quantify mortality and offspring production of sessile rotifers in macrophyte-dominated freshwaters (Edmondson, 1945). Here, we describe methods to culture sessile species that are generally employed to culture planktonic rotifers. With these methods, life table experiments can now be extended to assess the effects of environmental variables, as well as potentially toxic agents, on sessile rotifers.
References
Borowitzka, M. A. & L. J. Borowitzka, 1988. Micro-algal biotechnology. Cambridge University Press, London: 480 pp.
Duarte, C. M., S. Agustin & H. Peters, 1987. An upper limit to the abundance of aquatic organisms. Oecologia 74: 272–276.
Dumont, H. J., S. S. S. Sarma & A. J. Ali, 1995. Laboratory studies on the population dynamics of Anuraeopsis fissa (Rotifera) in relation to food density. Freshwater Biology 33: 39–46.
Edmondson, W. T., 1944. Ecological studies of sessile Rotatoria. Part I. Factors affecting distribution. Ecological Monographs 14: 31–66.
Edmondson, W. T., 1945. Ecological studies of sessile Rotatoria. Part II: dynamics of populations and social structures. Ecological Monographs 15: 141–172.
Edmondson, W. T., 1946. Factors in the dynamics of rotifer populations. Ecological Monographs 16: 357–372.
Enríquez-García, C., S. Nandini & S. S. S. Sarma, 2009. Seasonal dynamics of zooplankton in Lake Huetzalin, Xochimilco (Mexico City, Mexico). Limnologica 39: 283–291.
Espinosa-Rodríguez, C. A., S. S. S. Sarma & S. Nandini, 2012. Interactions between the rotifer Euchlanis dilatata and the cladocerans Alona glabra and Macrothrix triserialis in relation to diet type. Limnologica 42: 50–55.
Galkovskaja, G. A., 1987. Planktonic rotifers and temperature. Hydrobiologia 147: 307–317.
Garcia, M. A., 2004. The asexual life history of the colonial rotifer, Sinantherina socialis (Linnaeus). Department of Biology and Evolutionary Biology, Yale University, New Haven, CT: 129.
King, C. E., 1967. Food, age, and the dynamics of a laboratory population of rotifers. Ecology 48: 111–128.
King, C. E., 1982. The evolution of lifespan. In Dingle, H. & J. P. Hegmann (eds), Evolution and Genetics of Life Histories. Springer, New York: 121–138.
Krebs, C. J., 1985. Ecology. The Experimental Analysis of Distribution and Abundance, 3rd edn. Harper and Row, New York: 800 pp.
Kuczyńska-Kippen, N., 2007. Habitat choice in rotifer communities of three shallow lakes: impact of macrophyte substratum and season. Hydrobiologia 593: 27–37.
Lewis, P.A., D. J. Klemm, J. M. Lazorchak, T. J. Norberg-King, W. H. Peltier, & M. A. Heber. 1994. Short-term methods for estimating the chronic toxicity of effluents and receiving waters to freshwater organisms, 3rd edn. United States Environment Protection Agency Report EPA-600-4-91-002, U.S. EPA: Cincinnati, OH.
Lubzens, E., A. Tandler & G. Minkoff, 1989. Rotifers as food in aquaculture. Hydrobiologia 186: 387–400.
Meksuwan, P., P. Pholpunthin, E. J. Walsh, H. Segers & R. L. Wallace, 2014. Nestedness in sessile and periphytic rotifer communities: A meta-analysis. International Review of Hydrobiology 99: 48–57.
Meyer, J. S., C. G. Ingresoll, L. L. McDonald & M. S. Boyce, 1986. Estimating uncertainty in population growth rates: jackknife vs. bootstrap techniques. Ecology 67: 1156–1166.
Miracle, M. R. & M. Serra, 1989. Salinity and temperature influence in rotifer life history characteristics. Hydrobiologia 52: 81–102.
Monakov, A. V., 2003. Feeding of freshwater invertebrates. Kenovi Productions, Ghent: 373 pp.
Nandini, S. & S. S. S. Sarma, 2000. Life table demography of four cladoceran species in relation to algal food (Chlorella vulgaris) density. Hydrobiologia 435: 117–126.
Nandini, S., P. Ramírez-García & S. S. S. Sarma, 2005. Seasonal variations in the species diversity of rotifers from Lake Xochimilco (Mexico). Journal of Freshwater Ecology 20: 289–294.
Nandini, S., S. S. S. Sarma, R. J. Amador-López & S. Bolaños-Muñoz, 2007. Population growth and body size in five rotifer species in response to variable food concentration. Journal of Freshwater Ecology 22: 1–10.
Nandini, S., P. Ramírez-Garcia & S. S. S. Sarma, 2009. Evaluation of primary and secondary production using wastewater as a culture medium. Waste Management & Research 28: 928–935.
Sarma, S. S. S. & T. R. Rao, 1991. The combined effects of food and temperature on the life history parameters of Brachionus patulus Müller (Rotifera). Internationale Revue der gesamten Hydrobiologie und Hydrographie 76: 225–239.
Sarma, S. S. S. & S. Nandini, 2001. Life table demography and population growth of Brachionus variabilis Hampel, 1896 in relation to algal (Chlorella vulgaris) density. Hydrobiologia 446(447): 75–83.
Sarma, S. S. S., R. A. María-Isabel & S. Nandini, 2006. Population dynamics of littoral rotifers (Lecane inermis and Lepadella rhomboides) (Rotifera) in relation to algal (Chlorella vulgaris) food density. International Journal of Ecology and Environmental Sciences 32: 271–276.
Sawada, M. & H. E. Enesco, 1984. A study of dietary restriction and lifespan in the rotifer Asplanchna brightwelli monitored by chronic neutral red exposure. Experimental Gerentology 19: 329–334.
Sibly, R. M. & J. Hone, 2002. Population growth rate and its determinants: an overview. Philosophical Transactions of the Royal Society, London B: Biological 357: 1153–1170.
Snell, T. W., R. K. Johnston, K. E. Gribble & D. B. Mark Welch, 2015. Rotifers as experimental tools for investigating aging. Invertebrate Reproduction & Development 59: 5–10.
Stemberger, R. S., 1981. A general approach to the culture of planktonic rotifers. Canadian Journal of Fisheries and Aquatic Sciences 38: 721–724.
Stemberger, R. S. & J. J. Gilbert, 1985. Body size, food concentration, and population growth in planktonic rotifers. Ecology 66: 1151–1159.
Thor, P., G. Cervetto, S. Besiktepe, E. Ribera-Maycas, K. W. Tang & H. G. Dam, 2002. Influence of two different green algal diets on specific dynamic action and incorporation of carbon into biochemical fractions in the copepod Acartia tonsa. Journal of Plankton Research 24: 293–300.
Wallace, R. L., 1978. Substrate selection by larvae of the sessile rotifer Ptygura beauchampi. Ecology 59: 221–227.
Wallace, R. L. & W. T. Edmondson, 1986. Mechanism and adaptive significance of substrate selection by a sessile rotifer. Ecology 67: 314–323.
Wallace, R. L. & A. H. Smith, 2013. Rotifera. eLS. Wiley, Chichester.
Wallace, R. L. & P. L. Starkweather, 1983. Clearance rates of sessile rotifers: In situ determinations. Hydrobiologia 104: 379–383.
Wallace, R. L. & P. L. Starkweather, 1985. Clearance rates of sessile rotifers: In vitro determinations. Hydrobiologia 121: 139–144.
Wallace, R. L., J. J. Cipro & R. W. Grubbs, 1998. Relative investment in offspring by sessile Rotifera. Hydrobiologia 387(388): 311–316.
Wallace, R. L., T. W. Snell, C. Ricci & T. Nogrady, 2006. Rotifera Biology, Ecology and Systematics, 2nd ed. Kenobi Productions, Ghent.
Wallace, R. L., T. Snell & H. A. Smith, 2015. Rotifer: ecology and general biology. In Thorp, J. H. & D. C. Rogers (eds), Thorp and Covich’s freshwater invertebrates. Elsevier, Waltham, MA: 225–271.
Walz, N., 1983. Individual culture and experimental population dynamics of Keratella cochlearis (Rotatoria). Hydrobiologia 107: 35–45.
Walz, N., 1987. Comparative population dynamics of the rotifers Brachionus angularis and Keratella cochlearis. Hydrobiologia 147: 209–213.
Walz, N., 1993. Life history strategies of rotifers, Plankton regulation dynamics. Experiments and models in rotifer continuous cultures. In N. Walz (ed), Ecological Studies 98: 193–214. Berlin: Springer.
Weber, C.I., 1993. Methods for measuring the acute toxicity of effluents and receiving waters to freshwater and marine organisms. United States Environmental Protection Agency, Cincinnati, Ohio, EPA/600/4-90/027.
Xi, Y.-L., X.-F. Huang, H.-J. Jin & J.-K. Liu, 2011. The effect of food concentration on the life history of three types of Brachionus calyciflorus females. International Review of Hydrobiology 86: 211–217.
Yoshimatsu, T. & M. A. Hossain, 2014. Recent advances in the high-density rotifer culture in Japan. Aquaculture International 22: 1587–1603.
Yoshinaga, T., A. Hagiwara & K. Tsukamoto, 2001. Why do rotifer populations present a typical sigmoid growth curve? Hydrobiologia 446: 99–105.
Acknowledgements
SSSS and SN thank the authorities of FES I (Division of Investigation and Postgraduate Studies). MAJS received a scholarship from CONACyT. RLW was funded, in part, by NSF DEB 1257116.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: M. Devetter, D. Fontaneto, C. D. Jersabek, D. B. Mark Welch, L. May & E. J. Walsh / Evolving rotifers, evolving science
Rights and permissions
About this article

Cite this article
Sarma, S.S.S., Jiménez-Santos, M.A., Nandini, S. et al. Demography of the sessile rotifers, Limnias ceratophylli and Limnias melicerta (Rotifera: Gnesiotrocha), in relation to food (Chlorella vulgaris Beijerinck, 1890) density. Hydrobiologia 796, 181–189 (2017). https://doi.org/10.1007/s10750-017-3184-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-017-3184-5