
Abstract
The present study offers several management strategies in order to improve the performance of a free water surface constructed wetland that treats mainly eutrophic water and which is also designed to improve and increase the biodiversity of habitat and wildlife. To attain these goals, it has been necessary to analyze the influence of certain operational parameters and environmental factors on the mass removal rates (MRRs) and the mass removal efficiencies (MREs), depending on if the objective is to maximize nutrient removal or to achieve low effluent concentrations. The system, referred to as FG, operated in a range of hydraulic loading rates (HLRs) from 7 to 58 m year−1 and removed phosphorus (P) and nitrogen (N) at an average rate of 7.15 g P m−2 year−1 and 60.07 g N m−2 year−1. P and N removal varied seasonally, mainly due to input concentrations (C in), but inlet mass loading and HLRs also strongly influenced MRRs. Based on these results, we propose to maintain a mean HLR of 58 m year−1 in winter and 25 m year−1 in summer to increase annual nutrient removal and thereby barely affecting pumping costs.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
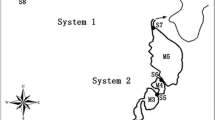
Cultural eutrophication is the main problem facing most surface waters nowadays (Smith & Schindler, 2009). Successful eutrophication mitigation is based mainly on the restriction of nutrient inputs to the waterbody, and this can be achieved by a wide variety of external and internal controls (Cooke et al., 1993; Smith et al., 1999). Although external nutrient inputs tend to be the main cause of eutrophication in shallow lakes and other water bodies, internal nutrient loads, i.e., nutrients released into the water column from sediments and from phytoplankton biomass decomposition, could be a major nutrient source that delays their recovery (Søndergaard et al., 2003; Jeppesen et al., 2005). There are several methods to reduce internal loads, such as the use of phosphates inactivation agents (Cooke et al., 1993; Zamparas & Zacharias, 2014) or removal of sediment layers (Phillips et al., 1999). Recently, a new approach to reduce these loads, based on the use of FWSCWs (free water surface constructed wetlands), has been tested in l’Albufera de València Lake (AV Lake) (Martin et al., 2013; Rodrigo et al., 2013). The Lake is enclosed in the l’Albufera de València Natural Park (AVNP) (Fig. 1), which is a wetland of International Importance (Ramsar Convention, 1990) and a Zone of Special Protection for Birds. The main characteristics of AV Lake are that it is shallow (around 1 m deep), highly eutrophic, and it is surrounded by rice fields.

Location of AV Lake and Tancat de la Pipa and map of FWSCWs showing FG cells
Constructed wetlands (CWs) are widely used for the removal of pollutants from wastewaters, for urban storm water treatment, for industrial wastewater treatment, for mine water treatment, and for field runoff treatment (Kadlec & Wallace, 2009). These systems present several advantages compared to conventional treatments (easy and low construction cost, relative lower-energy requirements, low operational and maintenance cost, and provide habitats for a wide diversity of plants and animals). These characteristics make them ideal candidates for the improvement of water quality in aquatic ecosystems (Pomogyi, 1993; Spieles & Mitsch, 2000; Kadlec et al., 2010), and as result of this, wildlife biodiversity is also enhanced. Comín et al. (2001) proved that the restoration of wetlands belts around lagoons increases spatial heterogeneity and diversity of the landscape, as well as improving their water quality. Fleming-Singer & Horne (2006) reported that it is possible to achieve a suitable habitat for birds as well as achieving high rates of nitrogen removal.
Recently, this technology has been tested in the treatment of eutrophic waters (Coveney et al., 2002; Li et al., 2008; Dunne et al., 2012, 2013 ). However, there are scarce studies that report information concerning the management of these systems to enhance their performance, and in addition, these are, mostly, focused on the phosphorus removal. This is the case of the study carried out in Lake Apopka (Florida) by Dunne et al. (2012, 2015), the goal of which was to maximize P removal from the eutrophic lake water working at high HLR (30 m year−1). Researchers found that P removal performance increased during cool periods and decreased during warm periods while operating cost remained constant. Drawing from these results, a seasonal operating regime with low warm-season flows to increase cost-effectiveness was adopted.
The role of the FWSCWs built in AVNP is multiple: to treat the lake water, to improve and increase the biodiversity of habitat and wildlife (from the bottom to the top of the food web), while ensuring its public use compatible with the above. These desired outcomes imply that some operational or design parameters, such as water depth, vegetation cover, or the lining of the substratum, besides enhancing nutrient removal, should also provide a suitable habitat for wildlife, specially when the surrounding rice fields are dry, given that there are times when FWSCWs play an important role as a food source and refuge for birds.
Following on from this, the hypothesis of this study is that it is possible to enhance the system’s performance, based on knowledge and management of the main factors that influence it. In addition, our premise is that if the abovementioned analysis is found to be feasible, we propose the establishment of a range of optimum values for these factors depending on the goal, namely to maximize nutrient and phytoplankton removal from eutrophic water and point sources or to achieve the lowest possible effluent concentrations.
The efficiency obtained during the first 2 years of operation (April 2009–March 2011) by these FWSCWs has already been reported (Martín et al., 2013). The objectives of this study are (1) to determine the feasibility and the efficiency of a given FWSCWs to remove and retain P and N from eutrophic lake water operating over a longer period, (2) to analyze the influence of different operational parameters and environmental factors on nutrient removal, and (3) to establish optimum operating conditions and propose recommendations for the management and design of artificial wetlands to treat eutrophic waters.
The operational parameters studied are IML, HLR, water column depth, and vegetation cover. Otherwise, the environmental factors are C in, temperature, pH, dissolved oxygen concentration (DO), and presence of birds into the FWSCW.
Materials and methods
Site description and operation
The research described in this paper was conducted at Tancat de la Pipa, an area of approximately 40 ha, located on the north of AVNP (Valencia, Spain), and that was formerly a rice field. Site details are described in Martin et al. (2013). Briefly, three FWSCWs (named FG, fp, and F4) were built for treating eutrophic water from the AV Lake. This paper focuses on the operation of one of them, the system called FG, which is divided into three cells arranged in series and named FG1, FG2, FG3, respectively (Fig. 1). This design allows the independent operation and management of each cell.
Lake water enters to FG from a gully named Barranco del Poyo (BP) (Fig. 1). This flow is continuous, by gravity and it is measured daily through a V notch weir. The inlets to each cell are four small sluices (Fig. 1), 0.3 m wide, spaced every 30 m, with wooden or riser boards that can be raised or lowered to manipulate the water column depth and, therefore, the hydrological gradient between inlets and outlets. The outlets from FG are two sluice gates (Fig. 1) of 0.5 m width. The flows in each of these sluices are regularly measured with a properly calibrated mini current meter (specifically with SEBA F1). The treated water is pumped back into the AV Lake. Environmental restrictions prevent the cells from being artificially waterproofed; nevertheless, substantial percolation was not obtained from the water balance (only about 5% with respect to inflow). This low percolation is related to the low hydraulic conductivity of the soil, since its texture is silty-clay/silty-clay-loam.
The initial planting period was January–February 2009. The kind of plants and the initial plant density are shown in Martin et al. (2013). The vegetation cover in each treatment cell is displayed in Fig. 2. In early summer of 2009, FG1 and FG2 had already reached complete vegetation cover, but in October 2009, FG1 was harvested and after that, a suitable plant cover was never recovered. This was mainly due to strong herbivore predation of soft shoots. FG operation was interrupted again in March 2011, for planting Phragmites spp. in FG1, and in August 2011, to allow the spread of these macrophytes. Nevertheless, the reed that had been planted did not grow properly. The gradual disappearance of vegetation cover in FG2 was also caused by herbivore predation. Unlike previous cells, in FG3 the vegetation cover increased, though slightly, over time. This was mainly due to the invasion of Phragmites spp. and the low-density planting of Iris pseudacorus in FG3 in March 2011. However, FG3 never reached a high cover probably due to the low initial plant density (Fig. 2).

Vegetation cover in FG1, FG2, FG3, and FG. Plant cover was estimated as the area occupied by vegetation considering the projection of the aboveground biomass on the ground. FG is the weighted average, where the weight assigned is the cell area
Sample collection and analysis
In this paper, wetland operation data from April 6, 2009 to October 29, 2012, are reported. A total of 304 water samples were collected in 76 samplings carried out at the inflow and outflow of each cell, between 9 a.m. and 14 p.m. From April 2009 to October 2011, the samplings were carried out once a fortnight and from November 2011, on a monthly basis.
Water temperature, pH, conductivity and DO were measured in situ using portable field measurement equipment (WTW® probes) and water samples were taken with 2 L bottles placed to mid-water depth and preserved in cold storage until arrival at the laboratory. DO continuous measurements were recorded every 15 min for 24 h at each sampling point. The measurements were carried out on consecutive days for each point and in the following seasons: early autumn 2011, early winter 2012, early spring 2012, and early summer 2012. The days were randomly selected.
Water samples were filtered according to Standard Method (APHA, 1991) and analyzed for total nitrogen (TN), ammonium (NH4 +–N), nitrite (NO2 −–N), nitrate (NO3 −–N), total phosphorus (TP), and orthophosphates (DIP-P), using the Spectroquant® Analysis System by Merck. The difference between TP and DIP measured is called non-orthophosphate phosphorus (n-DIP). The organic nitrogen (ON) was estimated as difference between TN and dissolved inorganic nitrogen (DIN, the sum of NH4 +–N, NO3 −–N, NO2 −–N). Nutrient loads from dry and wet atmospheric deposition were estimated from samples obtained with a total atmospheric deposition sampler. This consists of a 35-cm-diameter funnel and a 5-L container to collect such deposition. After each rainfall event, the sample collected is analyzed.
Calculations
Data of rainfall and evapotranspiration were obtained from a regional agricultural research center (IVIA, http://riegos.ivia.es), for proximity at study area. The inlet mass loading (IML), mass removal rates (MRRs), and annual mass removal efficiencies (MREs) were calculated as follows:
where Q in is the inflow from BP (m3 days−1), C in is the input concentration (g m−3), Q P is the precipitation flow (m3 d−1), C P is the atmospheric deposition concentration (g m−3), ET is the evapotranspiration (m3 days−1), C out is the output concentration (g m−3), and A is the wetland area (m2). These variables were considered constant between measurements, except for precipitation, since it is highly variable in this area. The overall MRE were calculated as accumulated mass removed during the period of the study and divided by accumulated mass input during the same period.
We used the results obtained by Hernández-Crespo et al. (2016), in relation to aboveground biomass and the amount of nutrient storage in plant tissue (NPT) at the end of each growing season, to calculate the nutrient mass removed by plants uptake in FG system.
Statistics
We report descriptive statistics such as mean, maximum, minimum, and standard deviations. The normal distribution of each variable studied in this paper was ascertained using the Kolmogorov–Smirnov test (N > 30) or Shapiro–Wilk test (N < 30), and the Levene test was used for ascertaining homoscedasticity of data. After confirming normality and homoscedasticity, one-way ANOVA test was applied; otherwise, the Kruskal–Wallis non-parametric test was used to compare seasonal variations. When the ANOVA was significant, the post hoc Bonferroni test was used to identify different groups and for non-parametric samples Mann–Whitney U test. Student’s t test and Wilcoxon test for non-parametric variables were used to compare the influent and effluent pollutant concentrations. Spearman’s correlation coefficients were computed to study the relationship between variables. Multiple linear regression models were performed to predict nutrient mass removal rate as function of inflow concentration and hydraulic loading rate. In the present study, the forward stepwise method was used to build the linear regression models. Other non-linear models were performed (logarithmic). A level of P < 0.05 was considered statistically significant in all comparisons. Analyses were performed in SPSS 15.0 for Windows (SPSS Inc. Chicago, USA), except linear regression models that were carried out with Statgraphics Plus 5.1.
Results
Hydraulics, temperature, dissolved oxygen, and pH
During the study period, FG treated 5.4 × 106 m3 of water from BP (that is approximately 20% of the lake’s water volume) and a water volume from the rainfall lower than 0.1 Hm3. The evapotranspiration represented about 4% of the treated water, justifying its inclusion in the mass balance. The HLR (Fig. 3) was increased in several steps with mean values of 7, 25, 32, and 58 m year−1 from April 2009 to March 2011. However, from April 2011 the HLR had to be decreased to 27 m year−1 in order to minimize the pumping costs. The HLR tested were always within a suitable range value for wildlife habitat. The water depth oscillated between 0.08 and 0.25 m (Table 1), because of concern for the health of the newly emerging vegetation and the bird habitat. Nonetheless, the design limit is 0.35 m. The mean theoretical hydraulic residence time (HRT) was 3 days, but the values were variable (Table 1).

Time series (April 2009 through October 2012) of hydraulic loading rates measured. The solid line represents the mean value tested in FG
Temperature and DO measured at the outflow were significantly lower than inflow ones (P < 0.01). Effluent temperatures ranged among 3.7 and 30.7°C, with a mean value of 17.5°C, which is lower than mean inflow value (19.7°C). DO outflow concentrations ranged between 0.14 and 15.43 mg L−1, with a mean value of 3.71 mg L−1. Nevertheless, in the case of DO, its behavior in each FG cell was different. FG1 was oxygen producer, especially in autumn and winter, which is depleted into the following cells (FG2 and FG3). As expected, temperature and DO at the all points showed seasonal variation and inverse patterns. The lowest DO values in FG3 effluent were recorded in summer and the highest values were registered in winter and early spring (in accordance with the lowest temperatures and the largest DO production in FG1). Figure 4 shows daily oxygen oscillations registered in FG1 and FG3 effluents in each season. It shows higher DO concentrations in FG1 and FG3 effluents during the day in winter than in summer. The higher values were usually measured after midday (coinciding with a higher photosynthetic activity) and the lowest values just before sunrise. Note the low DO concentrations, almost zero, measured in FG3 in summer. Over the study period, DO concentrations showed a continuous decrease in FG3 effluent and an increasing trend in BP and FG1 and FG2 effluents.

Daily cycles of dissolved oxygen observed in each season for FG1 and FG3 effluents
The pH values at the outflow oscillated between 6.91 and 8.20, but in FG1 and FG2 cells it reached values of 9.
Input–output concentrations and seasonal variations
The main component of TP in the inflow was the n-DIP (average of 77%), which responds to a high eutrophication level (Fig. 5). In contrast, TN in the inflow was mostly DIN (61% of TN) and NO3 − (76% of DIN) in particular (Fig. 5). Nevertheless, in samples taken after rainfall events the DIP was the main TP component (reaching up to 76%) and the ammonium for TN. In these cases, the TP and the ammonium reach values higher than 1 mg L−1.

Mean and standard deviation observed in each seasons for a TP, b DIP, c n-DIP, d TN, e NH4 +, f NO3 −, and g ON. For significant differences: 1 spring, 2 summer, 3 autumn, 4 winter
For all the studied parameters, C in were significantly greater than outflow concentrations (P < 0.01), being the differences greater in n-DIP and NO3 −. In addition, seasonal differences were observed in both inflow and outflow concentrations (Fig. 5). At the inflow, the highest mean values were registered in autumn and winter for P and in spring and winter for N. The low levels of DIN recorded in summer are noteworthy. At the outflow, the minimum values for P were observed in winter and for N in spring, while the highest values were measured in summer for P and in winter for N (mainly due to high levels of nitrates). The ON outflow concentrations were higher than the inflow ones in summer.
A relevant result was that DIP outflow concentrations were significantly correlated with DO concentrations (r = −0.747, P < 0.01).
Nutrient removal
Inlet mass loadings were highly variable (Table 2) and significant correlation between these and the outflow concentrations were not found [except weakly for NO3 − (r = 0.237, P < 0.05)]. Atmospheric loads accounted around 2% with respect to the input from the BP. P and N mass removal rates (Table 3) and mass removal efficiencies (Table 4) ranged also widely over time, decreasing in the case of efficiencies, except for NO3 −. For P, the highest removal rates were obtained for n-DIP and the highest MREs for DIP. For N, both were achieved for NO3 −.
Regarding to seasonal variation, for P the highest MRRs and MREs were achieved in autumn and winter (P < 0.05). For N, the highest removal rates were reached in winter and spring, and the lowest in summer (P < 0.01). The lowest N efficiencies were obtained in winter and summer (P < 0.05). Therefore, the nutrient removal rates in FG are lower in summer and higher in winter.
The average annual mass removed for FG from the AV Lake was 306 kg year−1 of TP and 2558 kg year−1 of TN (2111 kg as DIN) from April 2009 to October 2012, representing between 0.1 and 0.9% and between 0.1 and 0.2% of input annual P and N loads to the lake (data not shown here).
Influence factors on nutrient removal
For almost all parameters, C in and IMLs were positive and significantly correlated with both MRRs and MREs (Table 5). In the case of MRRs, the relationships followed a linear trend, so the highest ones were obtained with the highest C in and IMLs, although for TN and NO3 − a logarithmic trend also fitted well to data with IMLs (Fig. 6). For MREs, the relationship was logarithmic approaching the asymptote at 100%.

TN and NO3 − mass loading versus TN and NO3 − mass removal rates. The solid line represents the linear fit and the dotted line the logarithmic
In addition, TP, n-DIP, and TN mass removal rates were positive, significant, and linearly correlated with HLRs, while DIP and DIN mass removal rates were not (Table 5). From the results shown in Table 5, it might be suspected that the HLR did not influence on P removal efficiency. However, it increased slightly with HLR up to 32 m year−1, although there were no significant differences working among 7–58 m year−1 (P > 0.05) (Fig. 7). Conversely, the MREs obtained operating at 27 m year−1, and coinciding with lower plants presence (Fig. 2), were significantly lower than the MREs achieved with the previous HLRs (P < 0.05) (Fig. 7). As will be discussed later, the tested HLRs did not influence TN removal efficiencies (Fig. 7).

Mean and standard deviation of mass removal efficiencies (%) obtained in each HLR tested (m year−1). For significant differences: a 7, b 25, c 32, d 58, e 27
With respect to temperature, only TN, NO3 −, and DIP mass removal rates and DIP mass removal efficiencies were significant and negatively correlated with it (Table 5). These relations were linear, although really weak, so that the highest removal coincided with the lower effluent temperatures. The lack of relation may indicate that the temperature does not significantly influence in removal or that other factors are interfering with this influence.
Other factors studied were DO and pH of water and the vegetation cover (not shown in Table 5). TP, DIP, and NH4 + removal efficiencies were positive and significantly correlated with vegetation cover within the CW (r = 0.347, P < 0.01, r = 0.307, P < 0.01, r = 0.413, P < 0.01, respectively). DIP removal efficiencies were correlated non-linearly with DO concentrations and with pH values (r = 0.506, P < 0.01, r = 0.477, P < 0.01, respectively).
Two models for P and N retention were developed with the main factors analyzed: C in and HLR (Model 1 and Model 2). The multiple linear regression models were significant (P < 0.0001) and for TP it explains about 83% of the variable-dependent variance and for TN the 58%. The C in was the most important independent variable in both models.
The main factor that influenced nutrient removal efficiencies was clearly the C in (Table 5). “Threshold C in,” below which is unlikely to achieve positive efficiencies, was estimation by fitting the data to a logarithmic curve (Table 6).
Plant biomass and nutrient storage
The estimated values of aboveground biomass in FG varied between 0.61 and 0.73 kg d m m−2 (Table 7). Nonetheless, these values increase appreciably if the vegetated area alone is considered (Table 7). Furthermore, these values were not homogeneous between cells and over time.
N and P accumulation in aboveground standing crop varied between 0.1 and 16 gN m−2, and 0.01 and 2.3 gP m−2. Annual mass balance calculations showed that plant storage represented between 7 and 31% of the N and 9 and 41% of the P removed by FG. However, these nutrients were not definitively removed from the FG, as only aboveground biomass of FG1 was harvested at the end of the first growing season, representing the 9% of N and 14% of P removed that year for this cell. In the whole of the study period, nutrient removal by plants only accounted 5% of N and P removed in total by FG.
Discussion
The FWSCWs located in Tancat de la Pipa (Valencia, Spain) are efficient in removing nutrients and phytoplankton biomass from AV Lake (Martin et al., 2013). This study has focused on the operation of one of them, FG system, and analyzes the main factors that influence nutrient removal from eutrophic water.
The main aim is to establish optimum operating conditions and propose recommendations for the management and design of FWSCWs to treat eutrophic waters. These have to guarantee the removal of high amount of nutrients, obtaining low outflow concentrations and creating suitable habitat for the wildlife, conditions that are usually difficult to achieve simultaneously. As removing high quantities of nutrients implies working at high HLRs and the efficiency in nutrient removal is inversely related to the loading (Nichols, 1983), it should be prioritized getting maximized removal or alternatively low concentrations in the outlet, depending on the objective pursued in each case. In this work, the main parameters that influence in both performance indicators (removal rates and efficiencies) are analyzed.
Dissolved oxygen and temperature
DO and temperature are among the main factors that can influence on wetland efficiency, since they affect several biogeochemical processes. In FG, both variables show a decrease from inflow to outflow; in the case of DO this was mainly related with organic matter degradation and nitrification. Temperature decreases an average of 2.2°C, affecting microbiological process rates. This is an important issue to consider in the design of new CWs with similar characteristics.
On the other hand, seasonal variations at inflow and outflow were observed for both variables, and this could imply differences in nutrient removal depending on the season. In FG, there was a difference of almost 30°C between the maximum temperature in summer and the minimum temperature in winter (4°C). In fact, the low temperatures reached in winter could have affected processes sensitive to temperature, e.g, nitrification (Jing & Lin, 2004). DO oscillated inversely with temperature, due to the lower solubility and the greater biochemical oxygen demand (carbonaceous and nitrogenous) when the temperature increases (Kadlec & Reddy, 2001).
Since FG1 was harvested, this cell operated as an oxygen producer because of algal growth. This same trend was also observed in FG2. In fact, the algal photosynthesis in FG1 provoked wide oscillations of DO and high concentrations of this as shown in Fig. 4. In seasons with high phytoplankton biomass, as in the autumn, the concentrations oscillated from oversaturation to values close to zero. These daily fluctuations can affect biological and physical–chemical processes during the day, e.g., nitrification–denitrification, adsorption/desorption (Picot et al., 1993; Garcia et al., 2006). In FG3, daily DO oscillations were also observed, but not as pronounced as FG1, mainly due to the lower phytoplankton biomass.
Input–output concentrations and seasonal variations
Compared to FWSCWs that treat industrial or urban wastewater (Kadlec & Wallace, 2009), FG operated at low input nutrient concentrations and the main TP component was n-DIP instead of DIP (Fig. 5). Otherwise, in contrast with systems that treat eutrophic water (Dunne et al., 2013), in this study most of TN was DIN, and more concretely NO3 − (Fig. 5), showing a great influence of the rice fields drainage on BP, mainly from January to March. (Fig. 5f). Nevertheless, this water composition changed after intense Mediterranean rainfall events (normally in winter and autumn), when DIP and NH4 + concentrations strongly increased as a consequence of the arrival of urban runoff and the flushing effect on phytoplankton (Fig. 5b). During spring and autumn the particulate forms (n-DIP and ON) were high (Fig. 5c, g), related with the phytoplankton blooms (data not shown) in the AV Lake. Thus, this study demonstrates the versatility of the CW in the treatment of highly variable influents, both in terms of composition and pollution level.
FG successfully laminated the peaks of DIP, NH4 +, and NO3 −, being the mean reductions in autumn and winter of 88–92% for DIP and 72–47% for NH4 + (Fig. 5b, e) and in spring and winter of 93–47% for NO3 − (Fig. 5f). Therefore, building CWs around AV Lake would help reduce the impact of storm water and agricultural pollution on the water body, as it has been observed in other shallow eutrophic lakes (Jiang et al., 2007; Sollie et al., 2008). This role is especially important in sensitive aquatic systems, since the excess of these nutrients can lead to its eutrophication and, in the case of NH4 +, also because of its toxic form at high pH levels and its oxygen demand (Kadlec & Wallace, 2009).
The highest TP outflow concentrations were observed in summer due to a slight increase of the levels of DIP into the water column, which is inversely related with DO. When soils are under anaerobic conditions, iron is reduced from ferric to the ferrous state, releasing phosphorus that was previously held as insoluble ferric phosphate compounds (Mitsch & Gosselink, 2000). Moreover, the higher temperatures observed in this season could have favored the mineralization of organic matter, thus increasing the release of P from sediments to the water column.
For TN, the outflow concentrations were higher in winter and its reduction moderate (34%, Fig. 5d); this is probably due to the limitation of the nitrification (low temperatures), denitrification (high DO), and low macrophyte uptake (spring–summer phenomenon (Vymazal, 2007)). Nevertheless, the N reduction in summer was also low mainly due to high levels of ON, related with an increase of the plant litter decomposition and a rise of phytoplankton biomass in warmer months.
Nutrient removal and factors that affected them
Mass removal rates
The mean TN removal rate obtained (Table 3) is almost twice the value obtained by Dunne et al. (2013), with similar TN load, likely because of nitrates are the main inflow component in FG and these are more easily removed than organic forms in CWs (Phipps & Crumpton, 1994).
The main factors that affected TN mass removal rates were C in and IML (Table 5). MRRs appeared to increase linearly up to the highest loads (Fig. 6), in line with other authors (Tanner et al., 1995; Kadlec & Knight, 1996; Spieles & Mitsch, 2000). Nevertheless, some data may be indicating a logarithmic trend, and therefore a likely upper limit for TN loading (Fig. 6), but more data at high loading would be necessary to define a maximum MRR. HLR seems to weakly affect the TN removal rates, but not DIN (Table 5). For TN and NO3 −, the negative rates were obtained mainly in summer, being MRRs in this season significantly lower than that of the remaining seasons (P < 0.05). Owing to TN input concentrations and temperature data presented an inverse relationship, removal rates were higher in winter than summer, and it is reflected in the negative correlation found between temperature and MRRs (Table 5). It follows then, that the influence of temperature is obscured by the strong influence of C in in TN removal rates. For NH4 +, seasonality was not found.
The mean TP mass removal rate obtained (Table 3) in this study was higher than the values obtained for other CWs treating natural waters (Nairn & Mitsch, 2000; Dunne et al., 2012) likely because of the higher C in and IMLs in FG. In this survey, TP mass removal rates appeared to increase linearly up to the highest C in, IMLs, and HLRs (Table 5), according with other authors (Reddy et al., 1999; Nairn & Mitsch, 2000; Dunne et al., 2012). So, the results obtained suggest that the maximum removal rate has not been reached, and FG could work at higher loadings. This maximum is reached when soil exchange sites are filled, biological uptake is inhibited, or water velocities limit physical and chemical retention processes (Nairn & Mitsch, 2000). However, it is likely that the limit for HLR is not far in FG since the maximum limit in the water column depth for this CW is 0.35 m, and HLR much higher than 58 m year−1, without increasing the water column depth, would imply very low hydraulic residence time, which could limit the effective sedimentation and denitrification processes within CW. In addition, greater depth of water column than 0.3-0.35 m could affect the communities of emergent vegetation and the birds’ habitat. Kadlec (1999) suggested that the upper HLR limit is 100 m year−1.
Seasonality was found for TP and DIP. MRRs obtained in winter and autumn were significantly higher than those obtained in summer and spring (P < 0.05). This was associated with a higher C in of DIP in the coldest months, a higher DO concentration into the wetland (greater retention of PID in the sediment), a lower organic matter decomposition rate (the temperature values are at their lowest), and a DIP uptake by phytoplankton present. For n-DIP, seasonality was not found.
The C in was the most important independent variable in the multiple linear regression models developed. These models are really useful as management tools, for example, to establish the HLR that permits achieving an optimal MRR when the C in decreases or in the estimation of the removals achieved without measuring the effluent concentration, hence enabling significant cost savings.
Mass removal efficiencies
Overall nutrient mass removal efficiencies achieved in FG (Table 4) are close to the highest values found in CWs treating surface waters (Pomogyi, 1993; Moustafa et al., 1998; Hey et al., 1994; Coveney et al., 2002; Li et al., 2008; Kadlec et al., 2011).
C in was the main factor that influenced nutrient removal efficiencies (Table 5). HLRs did not influence in these, at least in the range tested in this study. The decrease in TP removal efficiencies observed at 27 m year−1 (Fig. 7) was related with a worsening of the sedimentation process owing to loss of vegetation (Fig. 2), since it led to the appearance of hydraulic short-circuiting and the exposure of the sediments to the re-suspension (Brix, 1997). In fact, the TP removal efficiencies were correlated with the percentage of vegetation cover in the wetland. In relation to TN removal efficiencies, the negative correlation found with HLRs (Table 5) corresponds to the fact that the highest C in were measured when the system worked at 7 m year−1, and it was shown that HLR did not really influence N mass removal efficiencies, as can be observed in Fig. 7. Nevertheless, it is likely that higher HLRs could negatively affect DIN removal because of the decrease in the contact time between the pollutants and microorganisms, as long as the depth is not increased.
Apart from the factors analyzed previously, there were other ones that had a negative influence, though to a lesser extent, on MREs obtained. One of them was the presence of birds within FG. Particularly in the period from late March to early May, a high density of birds use the CW for feeding, nesting, and roosting, and they provoke a decrease in MREs, specially for TP. This has been related with the re-suspension of material previously settled and with the input loads of nutrients through bird’s excrements. Fortunately, these loads only represent in average (annual basin) 0.5% of TN loading and 2.2% of TP load from the lake (CHJ, 2012), although they are greater between March and May, and thus, they were not considered for performances. Finally, the influence of DO concentrations and pH on DIP efficiencies is noteworthy. Although phosphorus inorganic precipitation has not been directly measured in this study, it probably occurred thanks to the CW’s high planktonic primary production and the alkalinity and calcium concentration of the water.
Plant role
The values obtained regarding nutrient storage in plants are in the range of values found in the literature for slightly loaded CWs (Coveney et al., 2002; Kadlec, 2006). Nevertheless, if the biomass is not harvested prior to fall senescence, most of the nutrients storage could return to the water column in the decomposition process (Brix, 1997). In general, it has been reported that removal of nutrients via plant harvesting is low, but it could be substantial for lightly loaded systems such as FG (Vymazal, 2007), and so there is much controversy in the literature about the fact of recommending the harvest or not. In this case, the annual storage of nutrients in aboveground biomass represents a substantial percentage of the nutrients removed from the inflow, particularly when input loadings are lower than 90 g m−2 year−1 of TN and 8 g m−2 year−1 of TP, thus being the plant harvesting an important via for permanent nutrient removal in FG, according to other studies treating eutrophic water (Tang et al., 2009).
In addition to the aforementioned, plant cover is also important in FG because it promotes the sedimentation and decreases the re-suspension (P removal), provides DO to the sediment (nitrification) and attachment sites for microorganisms, it is an organic matter source (denitrification), attenuates solar radiation (reduces algal growth), increases wildlife diversity (specially, it is a food source and refuge for birds), and offers aesthetic appearance to the system. Moreover, vegetated wetlands can act as a carbon sink (Mander et al., 2008; Mitsch et al., 2013), playing hence an important role in climate change.
Nevertheless, the main mechanism identified in FG for nutrient removal was sedimentation for TP and nitrification–denitrification process for TN (Martin et al., 2013). Sedimentation is usually the dominant TP removal mechanism FWSCWs (Coveney et al., 2002; Dunne et al., 2012) and the denitrification process is frequently considered as the major mechanism for removing N in most types of CWs (Vymazal, 2007), including some that treat river/lake water (Pomogyi, 1993; Reilly et al., 2000); notwithstanding this, other mechanisms, such as N uptake by macrophytes, algae and microorganism, or N sedimentation, have also been identified as main mechanisms in CWs treating eutrophic water (Dunne et al., 2013).
Efficiencies trend over time
MRE decreased over time, with the exception of nitrates (Table 4). Probable causes include the decrease in input concentrations (except for NO3 −), the accumulation of organic matter in sediments (which is mineralized releasing nutrients to the water column), the strong reduction of vegetation cover, and the global DO reduction. In the case of P, another cause could be that the load was continually greater than 1 g m−2 year−1, so P adsorption capacity of the soil could become saturated and then lose the capacity of retain P (Nichols, 1983; Richardson & Quian, 1999).
Conversely, the loss of vegetation could have improved the NO3 − removal because of the decrease of the oxygen production and the increase of phytoplankton biomass. The latter provoke daily DO cycles within FG, reaching anoxic conditions at night (Fig. 4). Moreover, the DO measured at FG3 effluent decreased over time, which responds to wetland maturation. However, it has been established that plants play an important role in the nitrogen removal via sequential nitrification–denitrification (Gersberg et al., 1986) since they supply organic carbon and act as a support to surface area for microbial growth.
Management strategies
In the present study, HLR and IML have been identified as the main operational parameters and C in as the main environmental factor that influence nutrient removal rates. Thus, increasing the flow when inflow concentrations are higher, in autumn and winter for P and in spring and winter for N in this case, presents an excellent opportunity for increasing the nutrients removed from BP (inlet), which leads to further eutrophication in AV Lake. In both cases, mass removal rates are higher in winter due to mainly random pollution events from storm water and agricultural runoff.
So, it could be interesting to raise the flow during winter and decrease it during summer (the season with the lowest nutrient mass removal rates). This change increases annual nutrients removal, scarcely affecting pumping costs. In summer, a mean HLR of 25 m year−1 could be maintained and 58 m year-1 in winter. In this case, approximately 3 g m−2 year−1 of TP and 16 g m−2 year−1 of TN would be removed in summer and 12 g m−2 year−1 of TP and 106 g m−2 year−1 of TN in winter. Moreover, with a mean water depth of 0.2 m, the mean hydraulic residence time would be about 3 days in summer, avoiding an excessive HRT, which would favor, together with warm temperatures, algal blooms into the CW. Nevertheless, taking into account that the limiting nutrient in AV Lake is the P, it could also be interesting to increase the HLR in autumn. For example, if a HLR of 58 m year−1 was applied in this season, the mass removal rate would be approximately of 18 g m−2 year−1 of TP and 98 g m−2 year−1 of TN.
Another strategy could be to stop flow during the summer months and increase it in the other seasons. In summer, draw down water cells stimulate the growth and spread of vegetation and the mineralization of organic matter in sediments. Nonetheless, it can be expected that if suitable recovery of vegetation cover takes place in the wetland in forthcoming years, the support of oxygen to the wetland and N and P plant uptake will increase in this season, thereby achieving better nutrient removal.
In regard to removal efficiencies, the main parameter that influences these is C in, which cannot be managed in our case. Moreover, HLRs in the range of 7–58 m year−1 do not affect them. Therefore, MRRs can be maximized at 58 m year−1 without affecting to MRE, as long as the vegetation cover remains high.
Conclusions
FWSCWs are efficient at removing nutrients from eutrophic lake water and they are presented as a feasible alternative for the recovery of highly degraded surface waters. This study demonstrates that FG is efficient at removing NO3 −, DIP, and NH4 + from agricultural and urban runoff. In these cases, it operates as nutrient trap that protect the AV Lake.
The main factors that affected the TN and TP MRRs were the C in, the IMLs, and the HLRs. MRRs increased linearly with these factors suggesting that the maximum mass removal rates were not achieved in FG. On the other hand, the MREs mainly depended on C in, whereas HLRs between 7 and 58 m year−1 did not influence on nutrient MREs.
From the results obtained, to reach the desired balance between mass removal rates and efficiency, it is recommended working with HLR at a rate between 32 and 58 m year−1, depending on monetary constraints of pumping water into the lake. This study provides recommendations of operation, based on seasonal variation of C in. In CWs, that treats highly variable nutrient input concentrations (for example, due to point inputs from storm water or agricultural runoff), is interesting to manage the flow in order to increase the removal of nutrients and optimize the pumping cost. For that, we propose to increase the flow when the C in are high and reduce it when C in are low.
In the first year of operation, the cattail grew rapidly, attaining complete cover in the middle of the first growing season, and high nutrient removals. So, the monoculture of cattails is recommended when the herbivorous depredation is not a relevant problem. Otherwise, we recommend mixed systems with reeds and yellow iris since they are less attractive for depredation. The drawback is that reed would need approximately two growing seasons with a minimum water depth (lower than 0.1 m) for reaching a proper standing stock. Once the minimum cover required for system start-up (normally 60–80%, Kadlec & Wallace, 2009) is reached, the water flow can be gradually increased. In both cases, the harvest should be done by cells, thereby ensuring the presence of vegetated zones for water treatment and for bird habitat. As a general rule, each cell should be harvested every three years. The importance of having a high vegetation cover to achieve good results at removing nutrients has been demonstrated.
Abbreviations
- FWSCWs:
-
Free water surface constructed wetlands
- FG:
-
The FWSCW studied
- MRRs:
-
Mass removal rates
- MREs:
-
Mass removal efficiencies
- HLRs:
-
Hydraulic loading rates
- P:
-
Phosphorus
- N:
-
Nitrogen
- C in :
-
Input concentrations
- IML:
-
Inlet mass loading
- AV Lake:
-
l’Albufera de València Lake
- AVNP:
-
l’Albufera de València Natural Park
- CWs:
-
Constructed wetlands
- BP:
-
Barranco del Poyo
- DIN:
-
Dissolved inorganic nitrogen
- DIP:
-
Orthophosphates
- n-DIP:
-
Non-orthophosphate phosphorus
- DO:
-
Dissolved oxygen
References
APHA, 1991. Standard Methods for the Examination of Water and Wastewater, 17th ed. American Publish Health Association, Washington DC.
Brix, H., 1997. Do macrophytes play a role in constructed treatment wetlands? Water Science and Technology 35(5): 11–17.
CHJ, 2012. (Ed.). Memoria de investigación del Tancat de la Pipa. Technical report for the Spanish Water Authorities.
Comín, F. A., J. A. Romero, O. Hernández & M. Menéndez, 2001. Restoration of wetlands from abandoned rice fields for nutrient removal, and biological community and landscape diversity. Restoration Ecology 9(2): 201–208.
Cooke, G. D., E. B. Welch, S. A. Peterson & P. R. Newroth, 1993. Restoration and Management of Lakes and Reservoirs, 2nd ed. Lewis Publishers, Boca Raton, FL.
Coveney, M. F., D. L. Stites, E. F. Lowe, L. E. Battoe & R. Conrow, 2002. Nutrient removal from eutrophic lake water by wetland filtration. Ecological Engineering 19: 141–159.
Dunne, E. J., M. F. Coveney, E. R. Marzolf, V. R. Hoge, R. Conrow & R. Naleway, 2012. Efficacy of a large-scale constructed wetland to remove phosphorus and suspended solids from lake Apopka, Florida. Ecological Engineering 42: 90–100.
Dunne, E. J., M. F. Coveney, E. R. Marzolf, V. R. Hoge, R. Conrow, R. Naleway, E. F. Lowe, L. E. Battoe & P. W. Inglett, 2013. Nitrogen dynamics of a large-scale constructed wetland used to remove excess nitrogen from eutrophic lake water. Ecological Engineering 61: 224–234.
Dunne, E. J., M. F. Coveney, V. R. Hoge, R. Conrow, R. Naleway, E. F. Lowe, L. E. Battoe & Y. Wang, 2015. Phosphorus removal performance of a large-scale constructed treatment wetland receiving eutrophic lake water. Ecological Engineering 79: 132–142.
Fleming-Singer, M. S. & A. J. Horne, 2006. Balancing wildlife needs and nitrate removal in constructed wetlands: the case of the Irvine Ranch Water District’s San Joaquin Wildlife Sanctuary. Ecological Engineering 26: 147–166.
García, J., B. F. Green, T. Lundquist, R. Mujeriego, M. Hernández-Mariné & W. J. Oswald, 2006. Long term diurnal variations in contaminant removal in high rate ponds treating urban wastewater. Bioresource Technology 97: 1709–1715.
Gersberg, R. J., B. V. Elking, S. R. Lyong & C. R. Goldman, 1986. Role of aquatic plants in wastewater treatment by artificial wetlands. Water Research 20: 363–367.
Hernández-Crespo, C., N. Oliver, J. Bixquert, S. Gargallo & M. Martín, 2016. Comparison of three plants in a surface flow constructed wetland treating eutrophic water in a Mediterranean climate. Hydrobiologia 774: 183–192.
Hey, D. L., A. L. Kenimer & K. R. Barret, 1994. Water quality improvement by four experimental wetlands. Ecological Engineering 3: 381–397.
Jeppesen, E., M. Søndergaard, J. P. Jensen, K. Havens, O. Anneville, L. Carvalho, M. F. Coveney, R. Deneke, M. T. Dokulil, B. Foy, D. Gerdeaux, S. E. Hampton, S. Hilt, K. Kangur, J. Kohler, E. Lammens, T. L. Lauridsen, M. Manca, R. Miracle, B. Moss, P. Noges, G. Persson, G. Phillips, R. Portielje, S. Romo, C. L. Schelske, D. Straile, I. Tatrai, E. Willén & M. Winder, 2005. Lake responses to reduced nutrient loading – an analysis of contemporary long-term data from 35 case studies. Freshwater Biology 50: 1747–1771.
Jiang, C., X. Fan, G. Cui & Y. Zhang, 2007. Removal of agricultural non-point source pollutants by ditch wetlands: implications for lake eutrophication control. Hydrobiologia 581: 319–327.
Jing, S. R. & Y. F. Lin, 2004. Seasonal effect on ammonia nitrogen removal by constructed wetlands treating polluted river water in southern Taiwan. Environmental Pollution 127: 291–301.
Kadlec, R. H., 1999. The limits of phosphorus removal in wetlands. Wetlands Ecology and Management 7: 165–175.
Kadlec, R. H., 2006. Free surface wetlands for phosphorus removal: the position of the Everglades Nutrient Removal Project. Ecological Engineering 27: 361–379.
Kadlec, R. H. & R. L. Knight, 1996. Treatment Wetlands. CRC/Lewis Publishers, Boca Raton FL: 893.
Kadlec, R. H. & K. R. Reddy, 2001. Temperature effects in treatment wetlands. Water Environment Research 73: 543–557.
Kadlec, R. H. & S. D. Wallace, 2009. Treatment Wetlands, 2nd ed. CRC Press, Boca Raton, FL.
Kadlec, R. H., S. B. Roy, R. K. Munson, S. Charlton & W. Brownlie, 2010. Water quality performance of treatment wetlands in the Imperial Valley, California. Ecological Engineering 36: 1093–1107.
Kadlec, R. H., J. S. Bays, L. E. Mokry, S. Andrews & M. R. Ernst, 2011. Performance analysis of the Richland-Chambers treatment wetlands. Ecological Engineering 37: 176–190.
Li, L., Y. Li, D. K. Biswas, Y. Nian & G. Jiang, 2008. Potential of constructed wetlands in treating the eutrophic water: evidence from Taihu Lake of China. Bioresource Technology 99: 1656–1663.
Mander, Ü., K. Lõhmus, S. Teiter, T. Mauring, K. Nurk & J. Augustin, 2008. Gaseous fluxes in the nitrogen and carbon budgets of subsurface flow constructed wetlands. Science of the Total Environment 404(2): 343–353.
Martín, M., N. Oliver, C. Hernández-Crespo, S. Gargallo & M. C. Regidor, 2013. The use of free water surface constructed wetland to treat the eutrophicated waters of lake L’Albufera de Valencia (Spain). Ecological Engineering 50: 52–61.
Mitsch, W. J. & J. G. Gosselink, 2000. Wetlands, 3rd ed. Wiley, New York.
Mitsch, W. J., B. Bernal, A. M. Nahlik, Ü. Mander, L. Zhang, C. J. Anderson, S. E. Jørgensen & H. Brix, 2013. Wetlands, carbon, and climate change. Landscape Ecology 28(4): 583–597.
Moustafa, M. Z., T. D. Fontaine, M. Guardo & R. T. James, 1998. The response of a freshwater wetland to long-term ‘low level’ nutrients loads: nutrients and water budget. Hydrobiologia 264: 41–53.
Nairn, R. W. & W. M. Mitsch, 2000. Phosphorus removal in created wetland ponds receiving river overflow. Ecological Engineering 14: 107–126.
Nichols, D. S., 1983. Capacity of natural wetlands to remove nutrients from wastewater. Water Pollution Control Federation 55(5): 495–505.
Phipps, R. G. & W. G. Crumpton, 1994. Factors affecting nitrogen loss in experimental wetlands with different hydrologic loads. Ecological Engineering 3: 399–408.
Phillips, G., A. Bramwell, J. Pitt, J. Stansfield & M. Perrow, 1999. Practical application of 25 years’ research into the management of shallow lakes. Hydrobiologia 395(396): 61–76.
Picot, B., S. Moersidik, C. Casellas & J. Bontoux, 1993. Using diurnal variations in a high rate algal pond for management pattern. Water Science and Technology 28(10): 209–215.
Pomogyi, P., 1993. Nutrient retention by the Kis-Balaton Water Protection System. Hydrobiologia 251: 309–320.
Reddy, K. R., R. H. Kadlec, E. Flaig & P. M. Gale, 1999. Phosphorus retention in streams and wetlands: a review. Critical Reviews in Environmental Science and Technology 29(1): 83–146.
Reilly, J. F., A. J. Horne & C. D. Miller, 2000. Nitrate removal from a drinking water supply with large free-surface constructed wetlands prior to groundwater recharge. Ecological Engineering 14: 33–47.
Richardson, C. J. & S. S. Quian, 1999. Long-term phosphorus assimilative capacity in freshwater wetlands: a new paradigm for sustaining ecosystem structure and function. Environmental Science and Technology 33(10): 1545–1551.
Rodrigo, M. A., M. Martín, C. Rojo, S. Gargallo, M. Segura & N. Oliver, 2013. The role of eutrophication reduction of two small man-made Mediterranean lagoons in the context of a broader remediation system: effects on water quality and plankton contribution. Ecological Engineering 61: 371–382.
Smith, V. H. & D. W. Schindler, 2009. Eutrophication science: where do we go from here? Trends in Ecology and Evolution 24: 201–207.
Smith, V. H., G. D. Tilman & J. C. Nekola, 1999. Eutrophication: impacts of excess nutrient inputs on freshwater, marine, and terrestrial ecosystems. Environmental Pollution 100: 179–196.
Søndergaard, M., J. P. Jensen & E. Jeppesen, 2003. Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia 506–507: 135–145.
Sollie, S., H. Coops & J. T. A. Verhoeven, 2008. Natural and constructed littoral zones as nutrient traps in eutrophicated shallow lakes. Hydrobiologia 605: 219–233.
Spieles, D. J. & W. J. Mitsch, 2000. The effects of season and hydrologic and chemical loading on nitrate retention in constructed wetlands: a comparison of low- and high-nutrient riverine systems. Ecological Engineering 14: 77–91.
Tang, X., S. Huang, M. Scholz & L. Jinzhong, 2009. Nutrient removal in pilot-scale constructed wetlands treating eutrophic river water: assessment of plants, intermittent artificial aeration and polyhedron hollow polypropylene balls. Water air Soil Pollution 197: 61–73.
Tanner, C. C., J. S. Clayton & M. P. Upsdell, 1995. Effect of loading rate and planting on treatment of dairy farm wastewaters in constructed wetlands – II. Removal of nitrogen and phosphorus. Water Research 29: 27–34.
Vymazal, J., 2007. Removal of nutrients in various types of constructed wetlands. Science of the Total Environment 380: 48–65.
Zamparas, M. & I. Zacharias, 2014. Restoration of eutrophic freshwater by managing internal nutrient loads. A review. Science of the Total Environmental 96: 551–562.
Acknowledgements
Núria Oliver acknowledges the scholarship provided by the Generalitat Valenciana, Spain (VALi + D PhD Program). The authors are also grateful to the Confederación Hidrográfica del Jucar (CHJ, MMARM) for the financial support of the project.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Ethical approval
This manuscript has been prepared according to the ethical rules of Hydrobiologia and it has not been submitted to other journals.
Additional information
Handling editor: Pierluigi Viaroli
Rights and permissions
About this article

Cite this article
Oliver, N., Martín, M., Gargallo, S. et al. Influence of operational parameters on nutrient removal from eutrophic water in a constructed wetland. Hydrobiologia 792, 105–120 (2017). https://doi.org/10.1007/s10750-016-3048-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-016-3048-4