
Abstract
We investigated the main structuring forces driving epiphytic algae dynamics with wind disturbances and zooplankton grazing acting as potential stressors by inducing a natural 60-day epiphyton summer succession. We cleaned macrophytes in the subtropical shallow Mangueira Lake, southern Brazil, using soft sponges and sampled them randomly at short-term intervals. Simultaneously, we sampled and classified zooplankton from the littoral zone according to their particle ingestion size. Disturbance by wind was the main factor driving the epiphyton succession. Tightly attached diatoms were well adapted to the system’s high mean wind velocity (15 m s−1), whereas low wind velocity (<7 m s−1) reduced community diversity. Summer storms (46.7 mm of precipitation; 29.5 m s−1 wind velocity) caused phosphorus input and favored prostrate diatoms. Epiphyton was very productive (8028.8 mg C m−2 h−1), consistent with a heterogeneous community. Small rotifers and ciliates were abundant in the water column during the study. Our data indicate that rotifers as well as other zooplankton grazed on epiphyton, due to the positive correlation between algal diversity and rotifer abundance as well as between diatom and zooplankton biomasses, whereas Copepods and Cladocerans followed large motile diatoms in abundance. We conclude that epiphyton is an important food source for zooplankton in Mangueira Lake.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Attachment strategies and algal growth forms have been used to assess ecological traits in running waters, and correspond strongly to adaptations to resources (Passy, 2007; Lange et al., 2011; Berthon et al., 2011) and to physical disturbances (Schneck & Mello 2012). During succession, temporal changes in dominant growth forms are often related to changes in community resources (Passy, 2007; Kelly et al., 2009), and the use of growth forms can supply valuable information about the structure of the biofilm (Rimet & Bouchez, 2011). However, the use of growth forms for understanding ecological adaptations of benthic algae is still poorly realized in lentic environments (DeNicola & Kelly, 2014).
Colonization of clean substrata involves the formation of an organic biofilm followed by the passive settlement of cells with subsequent increases in abundance arising via reproduction as r-strategists (Szlauer-Łukaszewska, 2007). Species growth occurs by a sequence defined in time and space leading to the development of a complex community in a mature state (Hoagland et al., 1982; Biggs, 1996; Stevenson, 1996). The succession is determined by complex interactions between abiotic and biotic factors such as light, grazing, temperature, and water chemistry (Patrick & Reimer, 1966; Stevenson, 1996); habitat, substrate availability, and reserve of propagules of species (Hutchinson, 1975; Peterson & Stevenson, 1992); and species-specific traits (e.g. resource competitiveness; Ferragut & Bicudo, 2010). Successional stages vary with environmental factors, trophic state, substrata exposure times, and disturbances (Szlauer-Łukaszewska, 2007; Rodrigues dos Santos & Ferragut 2013). Thus, the availability of resources, such as light and nutrients, are regulatory factors that can redirect the successional trajectory on a local scale (Ferragut & Bicudo, 2010).
Shallow lakes have high potential for development of attached algae (Wetzel, 1990), as they possess large and well-lit areas and a diverse variety of submerged substrates for algal growth (Lowe, 1996; Liboriussen & Jeppesen, 2006), such as sediment (epipelon), stones (epilithon), and macrophytes (epiphyton). The size, shape (Wehr & Sheath, 2003), and hydrodynamics of coastal shallow lakes directly affect their physical and chemical properties (Scheffer, 1998). In subtropical shallow lakes with wind-driven hydrodynamics, which cause intense water circulation, the water column is not stratified (Cardoso et al., 2012). Furthermore, Cardoso & Motta Marques (2004) noted particular zooplankton species associated with the effects of fetch in the lake. Besides the effects of wind action, littoral zones of shallow depths are exposed to wave action and high irradiances (Cantonati & Lowe, 2014). Moreover, the littoral community suffers intense changes in response to nutrient inflow and water level due to rain, watershed, and river flow (Rimet et al., 2015). Disturbance events affect community structure (Peterson & Stevenson, 1992), and the presence, frequency, and intensity of such disturbance events are more important than their origin (Sommer et al., 1993). Disturbance intensity affects community diversity (Ács & Kiss, 1993) and also emphasizes the importance of refuges for stream benthic algae (Schneck & Mello, 2012), protecting organisms from disturbance, which act as areas with reduced flow (Lancaster & Hildrew). Refuge and propagule availability will determine the community’s ability to recover (Lake, 2000). In a subtropical shallow lake, Cardoso & Motta Marques (2004, 2009) showed that phytoplankton and zooplankton community changes were strictly related to wind action in the pelagic zone, and the littoral zone may act as a refuge for benthic organisms reducing wind action. On the other hand, wind action in the littoral zone was also shown to model the attached community, selecting them by their adaptations (Faria et al., 2015; Rimet et al., 2015, 2016). However, disturbance effects still remain poorly understood, particularly in terms of establishment of equilibrium states and non-equilibrium theories (Lengyel et al., 2014).
Benthic algae are an important source for whole-lake primary production in shallow lakes (Vadeboncoeur et al., 2008) and depend not only on light but also on the morphometry of the lake (Ask et al., 2009). In general, benthic algae are a quality food resource for many consumers (McCormick et al., 2001). In Mangueira Lake, the food web is sustained by autochthonous carbon and in addition to macrophytes, epiphyton plays an important role in nutrient cycling and storage, with consequences for zooplankton and fish biomasses, as observed by Rodrigues et al. (2014) using isotopic signatures and stomach contents. In summer, high temperatures and low nutrient supplies as well as high grazing rates limit phytoplankton growth (Fragoso et al., 2011). Zooplankton biomass in the pelagic zone of the southern part of Mangueira Lake showed temporal variability following the phytoplankton successional rates, but large-bodied zooplankton were not related to phytoplankton composition and abundances (Rosa et al., 2016), hence we assumed that epiphyton could act as the principal carbon supply for the community.
In this study, we investigated epiphytic algae development and their autogenic processes, such as algal nutrient content and primary production, in their natural environment in order to understand the main structuring forces driving epiphytic algae dynamics. During summer, we monitored a 60-day epiphytic algal succession on natural macrophytes in Mangueira Lake. We examined the following: (1) whether nutrients and/or disturbances are the main factors driving natural epiphyton succession; (2) whether a positive connection between primary production and grazer abundance as well as between zooplankton and epiphytic algae growth forms exists, indicating that the epiphyton community is structured by zooplankton grazing; and (3) whether high primary production is connected with high zooplankton abundance, which would suggest that epiphyton is an important food source for zooplankton.
Materials and methods
Study area
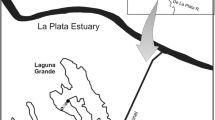
Mangueira Lake is located in southern Brazil on a narrow strip of land between the Atlantic Ocean and Mirim Lake (Crossetti et al., 2013). This coastal lake ranges from oligo- to mesotrophic conditions and covers approximately 820 km2. It is 90 km long and 3–10 km wide and the average depth is 3 m (Z max = 6 m) (Cardoso et al., 2012). The lake is under continuous wind influence, with predominantly NE–SW winds during the summer.
Field and laboratory procedures
Wind (velocity and direction) and precipitation data were obtained from Santa Vitoria do Palmar Meteorological Station (maintained by INMET, the National Meteorological Institute) from 13 January to 18 March of 2012, three times per day (12 am, 12 pm, and 6 pm), to describe the summer wind and precipitation patterns as well as to evaluate their disturbance potential.
We conducted the experiment in situ in the southern part of the lake (33°30′36.94′′S; 53°7′0.64′′W) over 60 days in the summer of 2012 (from January 18 to March 18). We chose two banks, 50 meters apart, of the emergent macrophyte Schoenoplectus californicus L. in the littoral zone for study. The macrophytes (n = 240; with 120 stems in each bank) were randomly chosen and carefully cleaned with a soft sponge to remove the preexisting biofilm. The tops of the stems were marked with adhesive tape for identification, which established ‘day one’ of the experiment. Two samples, one in each bank, in which stems were randomly sampled and stored, were taken on days 3, 6, 9, 12, 15, 20, 30, 45, and 60 of succession. Immediately after sampling, the biofilms were removed from the substrate with toothbrushes, fixed in Lugol 5% for quantitative analysis (n = 2, replicates), and stored frozen for nutrient and pigment analyses (compound sample, n = 2). On each sampling day, zooplankton samples were taken in the littoral zone using a suction pump, filtering 100 l through a nylon net with 25-µm mesh which was then concentrated into 250-mL polyethylene bottles and fixed with 4% formaldehyde (Wetzel & Likens, 2000). Water was sampled with plastic bottles (n = 2) that were stored frozen for nutrient analysis (nitrogen and phosphorus series) according to the method below.
Water depth and transparency were measured using a metered cable and Secchi disk, respectively. Temperature, dissolved oxygen, pH, and conductivity were measured in situ with a YSI 6920 probe. Water samples were collected in the littoral zone. The following nutrients were measured in the water column and in the biofilm (see sampling above): total nitrogen (TN-w for water and TN-b for biofilm), nitrate (NO3), ammonia (NH4 +), total phosphorus (TP-w for water and TP-b for biofilm), and orthophosphate (PO4) (Mackereth et al., 1989). Chlorophyll (Chla-w for water and Chla-b for biofilm) and pheophytin (Pheo-w for water and Pheo-b for biofilm) were extracted from GF/F filters in 90% ethanol (Jespersen & Christoffersen, 1987), and measured by spectrophotometry (APHA, 2005). In addition to these measurements, TN and TP molar ratios were calculated for water and biofilm. We used the data of Kahlert (1998) for the optimum ratios for benthic algae (18:1).
Primary production
Immediately after sampling, epiphyton was incubated for estimation of carbon uptake. Attached algae were removed and incubated with filtered lake water. The surface area of each plant was measured before scraping its biofilm. Algae primary production was estimated using the radiolabeled sodium bicarbonate (Na H14CO3) method (Wetzel & Likens, 2000). For each sample, 500 µl of radiolabeled bicarbonate (8.0 µCi ml−1 final concentration, PerkinElmer) was added to glass bottles (50 mL) which were then filled with the water containing the sampled algae (1 dark, 2 light, 1 initial). The covered bottles were incubated for 4 h (12–4 pm) inside a PVC box. A Photosynthetically Active Radiation (PAR) sensor (Hobo weather station) was used to detect and record the incidence of PAR during the experiment. The data, registered with the logger aid, provided a daily profile of light radiation. Interruption of incubation activity in the initial and incubated bottles was achieved by adding 1 mL of formaldehyde (final concentration, 0.4%, 1:100). The bottles were refrigerated and stored in the dark until processing. Beta-particle emission counts were performed in a Rackbeta Liquid Scintillation Counter (LKB Wallac 1209). The results were expressed in g C m−2 h−1.
Biotic data
Algae were identified and counted according to Utermöhl’s (1958) method, which yields 90% counting efficiency (Pappas & Stoermer, 1996). The data were converted into density (ind m−2 according to APHA, 2005) and biomass (105 µm−3 m−2 according to Hillebrand et al., 1999). Species diversity (H) was calculated using the Shannon–Wiener index. We classified taxa (total density and biomass >1%, according to Lavoie et al., 2009) into six categories of growth forms (modified from Schneck & Mello, 2012; Rimet & Bouchez, 2012; and Rimet et al., 2015) presented in Table 3. Growth forms were sub-classified based on the largest linear dimension axis (GALD) (Lewis, 1976) into four categories: class I (<10 μm), class II (11–20 μm), class III (21–50 μm), and class IV (>50 μm).
For Zooplankton, an aliquot of each sample unit was quantified in a Sedgwick-Rafter chamber (APHA, 2005) with a minimum of 80% efficiency (Pappas & Stoermer, 1996), and the values expressed as density (ind m−3) and biomass (mg l−1) (adapted from Wetzel & Likens, 2000). Species diversity (H) was calculated using the Shannon–Wiener index. The cell volume was calculated using geometric formulas for the specific forms or length–weight regression (Bottrell et al., 1976; Dumont et al., 1975; Ruttner-Kolisko, 1977; Malley et al., 1989), in which 10–20 individuals of each taxon were measured. Zooplankton were identified (Koste, 1978; Elmoor-Loureiro, 1997; Ogden & Hedley, 1980) and species and groups were also classified according to the size of particles that they are able to ingest and cross-referenced with algae GALD with the aim of understanding the relationship between zooplankton feeding behavior and epiphyton.
Data analyses
The succession rate of the community composition (σ) was calculated using the sum-of-differences method (Lewis, 1978). We conducted descriptive analyses for the biotic data and T-tests of dependent samples between days (in successional sequence) to investigate the responses of the categories to disturbances using Statistica 7.1 software (StatSoft, Inc., 2005). We also performed correlation analyses (r-Pearson P < 0.05) between the epiphyton community and environmental data (meteorological data and water data), biofilm nutritional states, primary production, and zooplankton biomass to identify both relationships among them as well as explanatory variables for inclusion in the subsequent multivariate analyses. Ordination analyses were performed using the software PC-ORD version 6.08 for Windows (McCune & Mefford, 2011). PCA of water attributes was performed to reveal environmental changes over the course of the experiment in the littoral zone. CCA was used to identify relationships among the algal growth form and zooplankton groups (both with biomass data) with the associated water measurements—TP-w, TN-w, Chla-w, Secchi depth, and temperature—as well as disturbances (precipitation and wind). The data were log x + 1 transformed prior to multivariate analysis.
Results
Environmental data and water attributes
Wind direction was constant (N–NE), with a high mean velocity (15 ± 6 m s−1) throughout the summer. Wind velocity peaked on day 20 (29.5 m s−1, with a mean of 28 m s−1 3 days before sampling).Wind velocity remained high until day 30 (7 m s−1) and then stabilized at 16 m s−1 until the end of experiment. The daily precipitation range was low (3–11 mm), with two peaks, on days 20 (47 mm) and 36 (33 mm); subsequent precipitation was constant and moderate until day 60 (2.5 mm). Therefore, we considered the summer storm (day 20) a disturbance, the mean wind velocity for the system (15 m s−1) as typical, and winds lower than that a quiescence, which drove the epiphyton succession process.
Ordination analysis (PCA) of the environmental data revealed the significance of the first axis (80.73%, P < 0.01), identifying the influence of the disturbance on the nutrients’ homogeneity in the water column as a result of the high precipitation (r = 0.99) and wind velocity (r = 0.82). This, in turn, increased TP-w (r = 0.63) and PO4 (r = 0.55) while reducing pH (r = −0.90) and conductivity (r = −0.95) (Fig. 1). Littoral zone water was under N-limiting conditions (N:P molar ratio <16 of Redfield ratio) and presented a homogeneous distribution of nutrient concentrations (CV < 25%) during summer 2012 (Table 1). The pH was alkaline (mean 8.7) and oligohaline (mean 0.42 mS cm−1).

Ordination analysis of environmental data on the littoral zone during study time in summer 2012. For the legend, see Table 1
Epiphyton nutrient levels and primary production
Epiphyton was N-limited (4 ± 1) (Table 2). TP-b increased with time, whereas TN-b varied during the succession, being correlated with Pheo-b (r = 0.80, P = 0.017). No correlations were observed between nutrient content and water chemistry.
Chla-b versus Pheo-b values revealed that community degradation occurred after day 12 (Fig. 2). Maximum carbon uptake was on day 15 (8028.8 mg C m−2 h−1). Even with low Chla-b (2.5 mg m−2), high assimilation was detected (7917.0 mg C m−2 h−1) on day 6. The PP:Chla ratio was negatively correlated with Pheo-b (r = - 0.73; P = 0.041), evidence of epiphyton productivity. No correlations were observed between nutrients and carbon uptake. Lower solar radiation favored production (PAR ranged from 243 to 1806 μmol m−2 s−1); however, production also occurred under high light incidence (PAR > 1365 μmol m−2 s−1). Aside from a high value identified on day 6, it was observed that primary production followed epiphyton biomass variation (Fig. 2).

Variation of chlorophyll a (Chla), phaeopigments (Pheo), PP:Chl a, solar radiation (PAR), and biomass of epiphytic algae succession during study time in summer 2012
Epiphyton and zooplankton community structures and succession rates
During the succession, 55 epiphytic taxa were counted (>1%), and the peaks of richness and total biomass (0.09 mg l−1) were registered on day 15, characterizing the end of the early successional stage and coinciding with the zooplankton’s first total density peak (Table 2, Fig. 3). No correlations were observed among community attributes and water chemistry. No significant changes of richness were observed in epiphyton afterwards (P > 0.01; T test for dependent samples), and density substantially decreased from day 15 to 20 (P = 0.048). Some important relationships between producers and grazers were observed. Epiphyton density was inversely correlated with zooplankton richness (r = −0.80, P = 0.017) and diversity (r = −0.81, P = 0.014). On the other hand, algal diversity intensified with zooplankton density (r = 0.74, P = 0.037).

Epiphyton and zooplankton data during study time in summer 2012. Biomass, total density (TD), diversity (H′), and Richness (S) as taxa numbers. BAC Bacillariophyceae, CYA Cyanobacteria, ZYG Zygnemaphyceae, CHL Chlorophyceae, PROTI Protist, ROTI Rotifera, CLADO Cladoceran, COPE Copepod
Epiphyton succession (Fig. 3) began with Chlorophyceae (low biomass [0.01 105 μm−3 cm−2], but with high density [96 ± 68 ind m−2]), which were responsible for the peak in Chla-b (r = 0.94, P = 0.001). Day 6 was marked by low biomass and the peak of phytoflagellates. Zygnemaphyceae (Cosmarium spp.) dominated the community biomass until day 12. Diatoms dominated the advanced stage of succession and were correlated with increases in biomass (r = 0.91, P = 0.001), being responsible for more than 70% of total biomass after day 20. The summer storm on day 20 favored diatoms attached parallel to substrata (mainly Epithemia spp.). The succession rate was high until day 9 due to the small-sized taxa that were registered as being able to colonize substrates faster compared to other species; the rate reduced after days 12–15 due to diatom stabilization. Biofilm NH4 + concentration increased with the rate stabilization (r = −0.73, P = 0.039). At the same time, Pheo-b was correlated with biomass (r = 0.90, P = 0.002). The zooplankton community was dominated by Rotifera and Protist (Tecamoeba and Ciliate) during the study. Therefore, potential grazers of small-sized class were found, and thus zooplankton density and biomass were strongly correlated (r = 0.96, P < 0.001) (Fig. 3). Zooplankton biomass was correlated with diatom biomass (r = 0.95, P < 0.001). The disturbance substantially decreased zooplankton in density and biomass; biomass and density recovered only on day 45. Copepod and Cladoceran were more frequent when large motile diatoms appeared in the biofilm toward the end of the experiment. The succession rate showed a constant decline after day 9, and epiphyton and zooplankton succession rates were correlated (r = 0.97, P < 0.001). Zooplankton individuals were also found in the biofilm; Protist (mainly Ciliate) were found in epiphyton samples throughout the study period. The tecamoeba peak occurred on day 45 when Rotifera eggs were found on the biofilm, reinforcing the relations between producers and grazers.
For the algae taxa, six growth forms and four GALD (the largest linear dimension axis) were classified in this study, while the zooplankton taxa were organized into the zooplankton groups and the GALD of particles that they are able to ingest (Table 3). We did not consider day 6 in the ordination analysis because the monodominance of phytoflagellates made it an outlier. Gradient analysis (CCA) explained 73.0% of the variance for the first two axes (P = 0.02), clearly separating the early phase of succession (until day 15) from the advanced phase (Fig. 4). Nutrients available in the water formed a gradient, where TN-w was related to the early phase (r = 0.61, axis 1) and TP-w (r = −0.56, axis 1; r = −0.59, axis 2) to the storm (r = −0.40, axis 1). The community responded to this gradient: succession began with loosely attached algae (I-L, r = 0.70; axis 1 and III-L, r = −0.65; IV-L, r = −0.77 axis 2) and small filamentous growth form (II-F, r = 0.47 axis 1). After day 12, prostrate diatoms increased in biomass (II-P, r = 0.46 axis 1 and III-P, r = −0.81 axis 2). On day 12, small ciliates were more abundant (I-CIL, r = −0.83 axis 2). Large prostrate diatoms (IV-P, r = −0.79 axis 1, r = −0.55 axis 2) were resistant and resilient to the disturbance and were strongly correlated with precipitation (r = 0.90, P = 0.002) and wind (r = 0.83, P = 0.011). Small erect growth form (II-E, r = −0.80 axis 1) and Tecamoeba (II-TECA, r = −0.54 axis 2) were also favored by the disturbance. Day 30 was characterized by dry weather, high temperature, and moderate wind (<7 ms−1), causing a loss of biomass favoring small filamentous algae (trichomes of Cyanobacteria) and the dominance of phytoflagellates. The advanced phase of succession (wind, 15 ms−1, and low precipitation, 1.7 mm) was driven by high transparency (r = −0.77) and Chla-w (r = −0.41) on axis 1. Motile diatoms increased in biomass (III-M, r = −0.84) on days 45 and 60 (IV-M, r = −0.46), and erect diatoms (IV-E, r = −0.80) and filamentous growth form (IV-F, r = −0.86) were related to the biggest zooplankton size class, for example, Cladoceran (IV-CLADO, r = 0.52), Copepods (IV-COPE, r = 0.47), and Rotifera (IV-ROTI, r = 0.41).

Large filamentous growth form and small motile diatoms favored greater Secchi depth (r = 0.77, P = 0.024; r = 0.085, P = 0.007; respectively). Large motile diatoms were correlated with the availability of TP in the biofilm (r = 0.095, P < 0.001), which also favored zooplankton density (r = 0.84, P = 0.008). Moreover, Rotifera (III-ROTI) were associated with increases in filamentous Cyanobacteria (II-F; r = 0.78, P = 0.022) and small loosely attached Chlorophyceae (II-L; r = 0.82, P = 0.011). Ciliates (II-CIL) were also correlated with filamentous Cyanobacteria (II-F; r = 0.89, P = 0.003). Erect growth form did not vary in time. Precipitation and wind were responsible for TP-w inputs favoring adnate and/or prostrate growth form (r = 0.72, P = 0.040) which rapidly increased in biomass (r = 0.72, P = 0.043). The community composition subsequently changed, experiencing an increase and persistence of diatoms in the advanced phase, which was dominated by tightly attached and motile diatoms, and also experiencing increases in loosely attached and filamentous algae.
Discussion
Disturbance is the main factor driving epiphyton succession and stabilization
In Mangueira Lake, the early stage of the succession was observed during the first two weeks, and loosely attached algae (unicellular green algae, desmids, and flagellate algae) were shown to be efficient pioneering colonizers, which are commonly reported in tropical reservoirs during early colonization (Ferragut & Bicudo, 2012; Pellegrini & Ferragut, 2012; Rodrigues dos Santos & Ferragut, 2013). However, in temperate environments, small adnate or erect diatoms are known to start the colonization (Hoagland et al., 1982; Stevenson, 1996; Ács et al., 2007, in laboratory conditions). Surface colonization always begins with a nutrient ‘conditioning film’ (Ács et al., 2007) due to organic matter and bacterial activities (Carrias et al., 2002) and our explanatory hypothesis is that mucilaginous biofilm could favor the adhesion of loosely attached algae after cell settlement. The stabilization of the waves in the water column due to the macrophyte bank (Jeppesen et al., 1998), it guarantees the success of this growth form as a pioneer under the intense wind action at Mangueira Lake. The community composition subsequently changed, with an increase in and persistence of diatoms and filamentous algae in the advanced phase, as expected (Hoagland et al., 1982; Biggs 1996; Szlauer-Łukaszewska, 2007).
In the present study, we recognized that summer storms, such as on day 20, are natural disturbances that drive epiphyton succession, whereas tightly attached growth forms, such as adnate and/or prostrate (Epithemia spp) and some erect diatoms, were resistant to such disturbances. Schneck & Mello (2012), in evaluating the resistance and resilience of organisms in a lotic environment, also found that adnate and/or prostrate taxa (including Epithemia) were more resistant to disturbance. These groups, primarily formed by diatoms, have specialized structures for tightly adhering to the substrate (Peterson & Stevenson, 1992) that are flexible enough to tolerate wave action (Lowe, 1996). The resistance to disturbances defines algal persistence (Stevenson, 1996). Community composition in Mangueira Lake was consistent with Lowe’s (1996) observation that epiphytic communities in the eulittoral zone are dominated by tightly attached algae in the presence of regular high-energy disturbances. As observed by Ács & Kiss (1993) for lotic environments, under stress, algae accommodated some disturbance levels. Disturbance effects were also observed in zooplankton, whose biomass was substantially reduced and was dominated by small forms such as tecamoebas—organisms suspended from sediment.
When wind disturbance was strongly reduced in frequency and intensity (<7 m s−1, based on the system mean of 15 m s−1) and coupled with high temperatures, as on days 6 and 30, community richness and diversity decreased. Since Mangueira Lake was under strong wind forces during the summer of the study, their reduction could have acted as an intermediate quiescence. According to Connell (1978), the absence of disturbance causes competitive exclusion and reduces diversity to minimal levels, which is accompanied by a decrease in phytoplankton biomass. Diversity and richness increased when wind velocities were under the mean for the system and the epiphytic community became resilient 15 days after the disturbance, when species tended to return to pre-disturbance patterns of heterogeneity (Fraterrigo & Rusak, 2008). Wind velocity, below and above the mean found for the system, should be considered a disturbance for the attached community during the succession process once it begins to influence community structure, richness, and diversity.
Mangueira Lake was under N-limiting conditions, and epiphyton was identified as an N-limited community. Nutrient limitation could be related to phytoplankton spring growth rates induced by warming, leading to nutrient limitation in the summer (Fragoso et al., 2011). This condition favored Epithemia, a known N-fixing diatom (DeYoe et al., 1992). Nitrogen-limited environments commonly present N-fixing algal (as well as Cyanobacteria) proliferations, such as Epithemia in temperate rivers; moreover, when consumed by grazers or while decomposing, they can also become an N source for the environment (Power et al., 2009). TP-w concentrations were highly consistent with a eutrophic state, contrasting with historical oligo-mesotrophic conditions. On the other hand, we found that TP increases in the water were strongly correlated with disturbances, as TP-w can rapidly increase during periods of strong wind in shallow lakes (Zhu et al., 2005). As a wind-exposed shallow lake, Mangueira Lake is under complete vertical mixture and seiches can transport materials (Fragoso et al., 2011), thus the nutrient pool is communal in coastal shallow lakes in southern Brazil due to sediment suspension driven by wind hydrodynamics (Cardoso & Motta Marques, 2009).
We found low correlations of littoral community with local water; however, TP-b was strongly related to TP-w inputs to the water due to the disturbance caused by the summer storm.TP-b also increased with succession time; the accumulation of TP in biofilm is well known (McCormick et al., 2001) and favors high diatom biomass which is known to increase with TP (Liboriussen & Jeppesen, 2006).
As observed by Rimet et al. (2015) in Lake Geneva, water chemistry in littoral zones changes faster compared to pelagic chemistry. Lake Geneva has a surface area similar to, but is deeper than, Mangueira Lake and is also influenced by northeast-dominated wind dynamics. As observed by them, littoral zones are exposed to wind and wave action where chemical and biological compartments are created; those zones which are not protected from disturbance are dominated by diatoms adapted to the turbulent environment (pioneer growth form) masking the effects of nutrient changes. In a previous study conducted over several summers in Mangueira Lake, we also found that wind dynamics are the main force regulating littoral zone dynamics and their attached communities, and diatoms in the southern area are dominated by pioneer growth forms that are related more to wind disturbance than to wind-driven nutrient gradients (Faria et al., 2015). In the present study, correlations showed that increases in biomass (adnate and/or prostrate growth form after a storm) were related both to wind and precipitation. We noted that during a succession process in a macrophyte bank, it was possible to better identify some growth form responding to nutrient inputs in water chemistry due to disturbances, than in a steady-state community which was dominated by a pioneer and tolerant growth form adapted to the disturbance condition (Faria et al., 2015).
Epiphyton was a productive community and zooplankton graze epiphytic algae.
Epiphyton was revealed to be a productive community and our results are in agreement with studies showing that the food web in Mangueira Lake is sustained by autochthonous carbon from macrophytes and epiphyton (Rodrigues et al., 2014), reinforcing the importance of epiphyton for lake production (Liboriussen & Jeppesen, 2003; Vadeboncoeur et al., 2008). This result was also observed in large ecosystems in the near-shore benthic zone, where benthic production can compensate for phytoplankton decline due to oligotrophication (Brothers et al., 2016). Indeed, macrophytes as non-inert substrates can be a significant carbon source for epiphyton, furnishing to the biofilm metabolites such as calcium carbonate (Burkholder & Wetzel, 1989). Besides that, living substrates could favor epiphyton colonization (Pip & Robinson, 1984) and greatly contribute to carbon production (e.g. 1770 mg C m−2 year−1, Villar et al., 1996). Moreover, high TP-w concentration occurs in the southern part of the lake where the water is clear, and production was superior to that found for eutrophic lakes, which usually present turbid waters (e.g. 1279 mg C m−2, Liboriussen & Jeppesen, 2003).
Autochthonous processes and light regulated primary production, whereas maximum production was coincident with a heterogeneous community. As we registered on day 6, phytoflagellates greatly contribute to primary production (Ilmavirta, 1988; Stoecker et al., 2000; Mariazzi et al., 1991). High production was also noted due to the abundance of green algae and desmids under high PAR (1806 μmol m−2 s−1). Studies have shown that green algae and diatoms are favored in high light regimes (Wall & Briand, 1979) and some diatoms present chromatic adaptation, responding to spectral distributions favoring algal growth (Wallen & Geen, 1971; Seiji & Gianesella-Galvão, 1991). In our controlled condition, summer radiation was high for the entire study (1326 ± 438 μmol m−2s−1). Liboriussen & Jeppesen (2003) showed that photosynthesis saturation happens between 100–400 μmol m−2 s−1. Radiation can negatively affect phytoplankton Chla (PAR 600–1000 μmol m−2 s−1), increasing at lower PAR (Collos et al., 1992). However, in Mangueira Lake, shading from macrophytes (Liboriussen & Jeppesen, 2003) could attenuate light levels, favoring epiphyton production in situ and yielding the increased results of the present study.
Small microzooplankton such as ciliate and small rotifers were dominant in the water column for the entire study of the epiphyton succession, and increased rotifer abundance was correlated with increasing algal diversity, suggesting a grazing-action relationship. As observed by Agasild et al. (2007), such groups could have a strong grazing impact on the small algae (5–15 μm) when those interactions are size-specific (Blukacz et al., 2010), as well as act as an important food resource for copepods (Bundy et al., 2005). Adnate/prostrate algae increased in the community with grazing, and this could be explained by the fact that grazers prefer loosely attached diatoms due to the ease of gathering (Peterson, 1987).
In Mangueira Lake, the significant correlations between the epiphyton and zooplankton communities could indicate not only top-down regulations of epiphyton by zooplankton, but also bottom-up regulation of zooplankton by epiphyton (Chen et al., 1997). Top-down theories, in which phytoplankton are controlled by zooplankton, are well studied (Sommer et al., 2003; Sommer & Sommer, 2006; Agasild et al. 2007; Agasild et al., 2013), whereas the epiphyton and zooplankton relationship is not. In Mangueira Lake, Fragoso et al. (2011) revealed that phytoplankton are limited by low N supplies in the summer and zooplankton grazing is limited by this light- and temperature-driven phytoplankton demand. Thus, epiphyton could act as a carbon resource for them. Kluijver et al. (2015) showed that carbon from macrophytes and periphyton greatly improves the cascading effects of planktonic food webs by benthic energy. The phytoplankton community in Mangueira Lake consists mainly of non-toxic cyanobacteria (Crossetti et al., 2014; Rosa et al., 2016) which are considered a poor food resource for zooplankton (DeMott et al., 2001), forcing them to look for other sources of food such as bacterioplankton or epiphyton. Because of that, macrozooplankton richness and abundance in Mangueira Lake were low and, as observed in historical data, larger cladocerans and Daphnia O.F. Müller, 1785 were never registered in this environment (Rosa et al., 2016). This explains why Copepods and Cladocerans (macrozooplankton) increased in the littoral zone when diatom biomass increased in the biofilm, as they provide good nutritional value for grazers (Cardoso & Motta Marques, 2004).
Biofilm removal by zooplankton grazing could favor the community by admitting more light, nutrient recycling, or favoring the settlement of other species in the epiphyton, thereby increasing its richness and diversity, as we observed. Our data reinforce the importance of the epiphytic community in subtropical shallow lake food webs. The community can supply substantial amounts of carbon during the summer, and periphytic diatoms can act as a quality food resource for zooplankton in an environment dominated by planktonic cyanobacteria.
References
Ács, E. & K. T. Kiss, 1993. Effects of the water discharge on periphyton abundance and diversity in a large river (River Danube, Hungary). Hydrobiologia 249: 125–133.
Ács, E., A. K. Borsodi, K. Kropfl, P. Vladar & G. Zarai, 2007. Changes in the algal composition, bacterial metabolic activity and element content of biofilms developed on artificial substrata in the early phase of colonization. Acta Bot Croat 66: 89–100.
Agasild, H., P. Zingel, I. Tõnno, J. Haberman & T. Nõges, 2007. Contribution of different zooplankton groups in grazing on phytoplankton in shallow eutrophic Lake Võrtsjäev (Estonia). Hydrobiologia 584: 167–177.
Agasild, H., P. Zingel, K. Karus, K. Kangro, J. Salujõe & T. Nõges, 2013. Does metazooplankton regulate the ciliate community in a shallow eutrophic lake? Freshwater Biology 58: 183–191.
American Public Health Association (APHA), 2005. Standard methods for examination of water and wastewater, Washington (DC).
Ask, J., J. Karlsson, L. Persson, P. Ask, P. Byström & M. Jansson, 2009. Terrestrial organic matter and light penetration: effects on bacterial and primary production in lakes. Limnology and Oceanography 54: 2034–2040.
Berthon, V., A. Bouchez & F. Rimet, 2011. Using diatom life-forms and ecological guilds to assess organic pollution and trophic level in rivers: a case of study of rivers in south-eastern France. Hydrobiologia 673: 259–271.
Biggs, B. J. F., 1996. Patterns in benthic algae of streams. In Stevenson, R. J., M. L. Bothwell & R. L. Lowe (eds), Algal Ecology: Freshwater Benthic Ecosystems. Elsevier, San Diego: 31–56.
Blukacz, E. A., W. G. Sprules, B. J. Shuter & J. P. Richards, 2010. Evaluating the effect of wind-driven patchiness on trophic interactions between zooplankton and phytoplankton. Limnology and Oceanography 55: 1590–1600.
Bottrell, H. H., A. Duncan, Z. M. Gliwicz, E. Grygierek, A. Herzig, A. Hillbricht-Ilkowska, H. Kurosawa, P. Larsson & T. Weglenska, 1976. A review of some problems in zooplankton production studies. Norwegian Journal of Zoology 24: 419–456.
Brothers, S., Y. Vadeboncoeur & P. Sibley, 2016. Benthic algae compensate for phytoplankton losses in large aquatic ecosystems. Global Change Biology. doi:10.1111/gcb.13306.
Bundy, M. H., H. A. Vanderploeg, P. J. Lavrentyev & P. A. Kovalcilk, 2005. The importance of microzooplankton versus phytoplankton to copepod populations during late winter and early spring in Lake Michigan. Canadian Journal of Fish and Aquatic Sciences 62: 2371–2385.
Burkholder, J. M. & R. G. Wetzel, 1989. Epiphytic microalgae on natural substrata in a hard water lake: seasonal dynamics of community structure, biomass and ATP content. Archives of Hydrobiology 83: 1–56.
Cantonati, M. & R. L. Lowe, 2014. Lake benthic algae: toward an understanding of their ecology. Freshwater Science 33: 475–486.
Cardoso, L. S. & D. Motta Marques, 2004. Structure of the zooplankton community in a subtropical shallow lake (Itapeva Lake – South of Brazil) and its relationship to hydrodynamic aspects. Hydrobiologia 518: 123–134.
Cardoso, L. S. & D. Motta Marques, 2009. Hydrodynamics-driven plankton community in a shallow lake. Aquatic Ecology 43: 73–84.
Cardoso, L. S., C. R. Fragoso Jr., R. S. Souza & D. Motta Marques, 2012. Hydrodynamic control of plankton spatial and temporal heterogeneity in subtropical shallow lakes. In Schulz, H. E., A. L. A. Simões & R. J. Lobosco (eds), Hydrodynamics-Natural Water Bodie. Intech Open Access Publisher, Rijeka: 27–48.
Carrias, J. F., J. P. Serre, T. S. Ngando & C. Amblard, 2002. Distribution, size, and bacterial colonization of pico- and nano-detrital organic particles (DOP) in two lakes of different trophic status. Limnology and Oceanography 47: 1202–1209.
Chen, C. C., J. E. Petersen & W. M. Kemp, 1997. Spatial and temporal scaling of periphyton growth on walls of estuarine mesocosms. Marine Ecology Pregress Series 155: 1–15.
Collos, Y., C. Descolas-Gros, M. Fontugne, A. Mortain-Bertand, M. J. Chrétiennot-Dinet & M. G. Frikha, 1992. Carbon and nitrogen dynamics during growth and degradation of phytoplankton under natural surface irradiance. Marine Biology 112: 491–496.
Connell, J. H., 1978. Diversity in tropical rain forests and coral reefs. Science 199: 1302–1310.
Crossetti, L. O., V. Becker, L. S. Cardoso, L. H. Rodrigues, L. S. Costa & D. Motta Marques, 2013. Is phytoplankton functional classification a suitable tool to investigate spatial heterogeneity in a subtropical shallow lake? Limnologica 43: 157–163.
Crossetti, L. O., F. Schneck, L. M. Freitas-Teixeira & D. Motta-Marques, 2014. The influence of environmental variables on spatial and temporal phytoplankton dissimilarity in a large shallow subtropical lake (Lake Mangueira, southern Brazil). Acta Limnologica Brasiliensia 26: 111–118.
DeMott, W. R., R. D. Gulati & E. Van Donk, 2001. Daphnia food limitation in three hypereutrophic Dutch lakes: evidence for exclusion of large-bodied species by interfering filaments of Cyanobacteria. Limnology and Oceanography 46: 2054–2060.
DeNicola, D. M. & M. Kelly, 2014. Role of periphyton in ecological assessment of lakes. Freshwater Science 33: 619–638.
DeYoe, H. R., R. L. Lowe & J. C. Marks, 1992. Effects of nitrogen phosphorus on the endosymbiont load of Rhopalodia gibba and Epithemia turgida (Bacillariophyta). Journal of Phycology 28: 773–777.
Dumont, H. J., I. van de Velde & S. Dumont, 1975. The dry weight estimate of biomass in a selection of Cladocera, Copepoda and Rotifera from the plankton, periphyton and benthos of continental waters. Oecologia 19: 75–97.
Elmoor-Loureiro, L. M. A. 1997. Manual de identificação de cladóceros límnicos do Brasil. Universa, Brasília. 156p.
Faria, D. M., L. S. Cardoso & D. Motta-Marques, 2015. Periphytic diatoms exhibit a longitudinal gradient in a large subtropical shallow lake. Inland Waters 5: 117–124.
Ferragut, C. & D. C. Bicudo, 2010. Periphytic algal community adaptative strategies in N and P enriched experiments in a tropical oligotrophic reservoir. Hydrobiologia 646: 295–309.
Ferragut, C. & D. C. Bicudo, 2012. Effect of N and P enrichment on periphytic algal community in a succession in a tropical oligotrophic reservoir. Limnology 13: 131–141.
Fragoso Jr., C. R., D. Motta Marques, T. F. Ferreira, J. H. Janse & E. H. van Nes, 2011. Potential effects of climate change and eutrophication on a large subtropical shallow lake. Environmental Modeling & Software 26: 1337–1348.
Fraterrigo, J. M. & J. A. Rusak, 2008. Disturbance-driven changes in the variability of ecological patterns and process. Ecology Letters 11: 756–770.
Hillebrand, H., C. D. Durselen, D. Kirschtel, U. Pollinger & T. Zohary, 1999. Biovolume calculation for pelagic and benthic microalgae. Journal of Phycology 35: 403–424.
Hoagland, K. D., S. C. Roemer & J. R. Rosowski, 1982. Colonization and community structure of two periphyton assemblages, with emphasis on the diatoms (Baccillariophyceae). American Journal of Botany 69: 188–213.
Hutchinson, G. E., 1975. A treatise on Limnology. Limnological Botany, Interscience, New York.
Ilmavirta, V., 1988. Phytoflagellates and their ecology in Finnish brown-water lakes. Hydrobiologia 161: 255–270.
Jespersen, A. M. & K. Christoffersen, 1987. Measurements of chlorophyll – a from phytoplankton using ethanol as extraction solvent. Archiv fur Hydrobiologie 109: 445–454.
Jeppesen, E., T. L. Lauridsen, T. Kairesalo & M. Perrow, 1998. Impact of submerged macrophytes on fish-zooplankton interactions in lakes. In Jeppesen, E., M. S. Søndergaard, M. Søndergaard & K. Christoffersen (eds), The Structuring Role of Submerged Macrophytes in Lakes. Springer, New York: 91–114.
Kahlert, M., 1998. C:N: P ratios of freshwater benthic algae. Archives of Hydrobiology 51: 105–114.
Kelly, M. G., L. King & B. NíChatháin, 2009. The conceptual basis of ecological-status assessments using diatoms. Biology and Environment: Proceedings of the Royal Irish Academy 109: 175–189
Kluijver, A., J. Ning, Z. Liu, E. Jeppensen, R. D. Gulati & J. J. Middelburg, 2015. Macrophytes and periphyton carbon subsidies to bacterioplankton and zooplankton in a shallow eutrophic lake in tropical China. Limnology and Oceanography 60: 375–385.
Koste, W. 1978. Rotatoria. II Tafelband. Gebrüder Borntraeger, Berlin, Stuttgart. 234p.
Lake, P. S., 2000. Disturbance, patchiness, and diversity in streams. Journal of National American Benthological Society 19: 573–592.
Lange, K., A. Liess, J. J. Piggott, C. R. Townsend & C. D. Matthaei, 2011. Light, nutrients and grazing interact to determine stream diatom community composition and functional group structure. Freshwater Biology 56: 164–278.
Lavoie, I., P. J. Dillon & S. Campeau, 2009. The effect of excluding diatom and reducing taxonomic resolution on multivariate analysis and strem bioassessment. Ecological Indicators 9: 213–225.
Lengyel, E., J. Padisák & C. Stenger-Kovács, 2015. Establishment of equilibrium states and effect of disturbances on benthic diatom assemblages of the Torna-stream, Hungary. Hydrobiologia 750(1): 43–56.
Lewis Jr., W. M., 1976. Surface/Volume ratio: implications for phytoplankton morphology. Science 192: 885–887.
Lewis Jr., W. M., 1978. Analysis of succession in a tropical phytoplankton community and a new measure of succession rate. The American Naturalist 122: 401–414.
Liboriussen, L. & E. Jeppesen, 2003. Temporal dynamics in epipelic, pelagic and epiphytic algal production in a clear and a turbid shallow lake. Freshwater Biology 48: 418–431.
Liboriussen, L. & E. Jeppesen, 2006. Structure, biomass, production and depth distribution of periphyton on artificial substratum in shallow lakes with contrasting nutrient concentration. Freshwater Biology 51: 95–109.
Lowe, R. L., 1996. Periphyton patterns in lakes. In Stevenson, R. J., M. L. Bothwell & R. L. Lowe (eds), Algal Ecology: Freshwater Benthic. Elsevier, San Diego: 57–76.
Mariazzi, A., V. Conzonno, R. Echenique & H. Labollita, 1991. Physical and chemical characters, phytoplankton and primary production of Ezequiel Ramos Mexíareresvoir (Argentina). Hydrobiologia 209: 107–116.
Mackereth, F. J. H., J. Heron & J. F. Talling, 1989. Water Analysis: Some Revised Methods for Limnologists. Freshwater Biological Association, Scientific Publication, Ambleside.
Malley, D. F., S. G. Lawrence, M. A. Maclver & W. J. Findlay, 1989. Range of variation in estimates of dry weight for planktonic Crustacea and Rotifera from temperate North American lakes. Canadian Technical Report of Fisheries and Aquatic Science 1666: 1–49.
McCormick, P. V., M. B. O’Dell, R. B. E. Shuford II, J. G. Backus & W. C. Kennedy, 2001. Periphyton responses to experimental phosphorus enrichment in a subtropical wetland. Aquatic Botany 71: 119–139.
McCune, B. & M. J. Mefford, 2011. PC-ORD Multivariate Analysis of Ecological Data.Version 6.0 MjM Software. Gleneden Beach, Oregon.
Ogden, C. G. & R. H. Hedley, 1980. An atlas of freshwater testate amoebae. British Museum (Natural History), Oxford University, Oxford. 222p.
Pappas, J. L. & E. F. Stoermer, 1996. Quantitative method for determining a representative algal sample count. Journal of Phycology 32: 693–696.
Passy, S. I., 2007. Diatom ecological guilds display distinct and predictable behavior along nutrient and disturbance gradients in running waters. Aquatic Botany 86: 171–178.
Patrick, R. & C. W. Reimer, 1966. The diatoms of United States. Academy of Natural Sciences, Philadelphia.
Pellegrini, B. G. & C. Ferragut, 2012. Seasonal and successional variation of a periphytic algal community on natural substrate in a tropical mesotrophic reservoir. Acta Botânica Brasilica 26: 807–818.
Peterson, C. G., 1987. Gut passage and insect grazer selectivity of lotic diatoms. Freshwater Biology 18: 455–460.
Peterson, C. G. & J. R. Stevenson, 1992. Resistance and resilience of lotic algal communities: importance of disturbance timing and current. Ecology 73: 1445–1461.
Pip, E. & G. G. C. Robinson, 1984. A comparison of algal periphyton composition on eleven species of submerged macrophytes. Hydrobiological Bulletin 18: 109–118.
Power, M., R. Lowe, P. Furey, J. Welter, M. Limm, J. Finlay, C. Bode, S. Chang, M. Goodrich & J. Sculley, 2009. Algal mats and insect emergence in rivers under Mediterranean climates: towards photogrammetric surveillance. Freshwater Biology. doi:10.1111/j.1365-2427.2008.02163.x.
Rimet, F. & A. Bouchez, 2011. Use of diatom life-form and ecological guilds to assess pesticide contamination in rivers: lotic mesocosm approaches. Ecological Indicators 11: 489–499.
Rimet, F. & A. Bouchez, 2012. Life-forms, cell-sizes and ecological guilds of diatoms in European Rivers. Knowledge and Management of Aquatic Ecosystems 406: 01.
Rimet, F., A. Bouchez & B. Montuelle, 2015. Benthic diatoms and phytoplankton to assess nutrients in a large lake: complementary of their use in Lake Geneva (France-Switzerland). Ecological Indicators 53: 231–239.
Rimet, F., A. Bouchez & K. Tapolczai, 2016. Spatial heterogeneity of littoral benthic in a large lake: monitoring implications. Hydrobiologia 771: 179–193.
Rodrigues, L. H. R., N. F. Fontoura & D. Motta Marques, 2014. Food web structure in a subtropical coastal lake: how phylogenetic constraints may affect species linkages. Marine & Freshwater Research 65: 453–465.
Rodrigues dos Santos, T. & C. Ferragut, 2013. The successional phases of a periphytic algal community in a shallow tropical reservoir during the dry and rainy seasons. Limnetica 32: 337–352.
Rosa, L. M., L. S. Cardoso, L. O. Crossetti & D. Motta-Marques, 2016. Spatial and temporal variability of zooplankton–phytoplankton interactions in a large subtropical shallow lake dominated by non-toxic cyanobacteria. Marine and Freswater Research. doi:10.1071/MF15356.
Ruttner-Kolisko, A., 1977. Suggestions for biomass calculation of plankton rotifers. Archiv für Hydrobiologie-Beiheft Ergebnisse der Limnologie 8: 71–76.
Scheffer, M., 1998. Ecology of Shallow Lakes. Chapman and Hall, London.
Schneck, F. & A. S. Mello, 2012. Hydrological disturbance overrides the effect of substratum roughness on the resistance and resilience of stream benhtic algae. Freshwater Biology 57: 1678–1688.
Seiji, D. & S. M. F. Gianesella-Galvão, 1991. Pigment chromatic adaptation in Cyclotella caspia Grunow (Bacillariophyta). Boletim do Instituto de Oceanografia 39: 123–130.
Sommer, U. & F. Sommer, 2006. Cladocerans versus copepods: the cause of contrasting top-down controls on freshwater and marine phytoplankton. Oecologia 147: 183–194.
Sommer, U., J. Padisák, C. S. Reynolds & P. Juhász-Nagy, 1993. Hutchinson’s heritage: the diversity-disturbance relationship in phytoplankton. Hydrobiologia 249: 1–7.
Sommer, U., F. Sommer, B. Santer, E. Zöllner, K. Jürgens, C. Jamieson, M. Boersma & K. Gocke, 2003. Daphinia versus copepod impact on summer phytoplankton: functional compensation at both trophic levels. Oecologia 135: 639–647.
Stevenson, R. J., 1996. An introduction to algal ecology in freshwater benthic habitats. In Stevenson, R. J., M. L. Bothwell & R. L. Lowe (eds), Algal Ecology: Freshwater Benthic Ecosystems. Elsevier, San Diego: 3–30.
Stoecker, D. K., D. E. Gustafson, C. T. Baier & M. M. D. Black, 2000. Primary production in the upper sea ice. Aquatic Microbial Ecology 21: 274–287.
Szlauer-Łukaszewska, A., 2007. Succession of periphyton developing on artificial substrate immersed in polysaprobic wastewater reservoir. Polish Journal of Envinronmental Studies 16: 753–762.
Utermöhl, H., 1958. Zur Vervollkommnung der quantitativen Phytoplankton Methodik. Mitteilungen des International Verein Limnologie 9: 1–38.
Vadeboncoeur, Y., G. Peterson, M. J. V. Zanden & J. Kalff, 2008. Benthic algal production across lake size gradients: interactions among morphometry, nutrients and light. Ecology 89: 2542–2552.
Villar, C., L. Cabot & C. A. Bonetto, 1996. Macrophytic primary production and nutrient concentration in a deltic floodplain marsh of the Lower Paraná River. Hydrobiologia 330: 59–66.
Wall, D. & F. Briand, 1979. Response of lake phytoplankton communities to in situ manipulations of light intensity and colour. Journal of Plankton Research 1: 103–111.
Wallen, D. G. & G. H. Geen, 1971. Light quality in relation to growth, photosynthetic rates and carbon metabolism in two species of marine plankton algae. Marine Biology 10: 34–43.
Wehr, J. D. & R. G. Sheath, 2003. Freshwater Algae of North America: Ecology and Classification. Elsevier, San Diego.
Wetzel, R. G., 1990. Land-water interfaces: metabolic and limnological regulators. Verhandlungen des Internationalen Verein Limnologie 24: 6–24.
Wetzel, R. G. & G. E. Likens, 2000. Limnological analysis. Springer, New York.
Zhu, G., B. Qin & G. GAO, 2005. Direct evidence of phosphorus outbreak release from sediment to overlying water in large shallow lake cause by strong wind wave disturbance. Chinese Science Bulletin 50: 577–582.
Acknowledgments
The authors thank the CAPES (Coordination of Improvement of Higher Education Personnel) for the doctoral grant awarded to the first author. We are grateful to CNPq and Dr. Lucia H.R. Rodrigues for logistical support; the IPH (Hydraulic Research Institute, at UFRGS) technicians for sampling support; Gustavo F. Hartmann for zooplankton counting; and Professors Carla Ferragut, Lezilda C. Torgan, Lucia H.R. Rodrigues, and Luciane de O. Crossetti, and the two unknown referees for constructive comments on an earlier version of the manuscript. The English language was reviewed by Cary Collett.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Handling editor: Luigi Naselli-Flores
Rights and permissions
About this article

Cite this article
de Faria, D.M., Cardoso, L.S. & da Motta Marques, D. Epiphyton dynamics during an induced succession in a large shallow lake: wind disturbance and zooplankton grazing act as main structuring forces. Hydrobiologia 788, 267–280 (2017). https://doi.org/10.1007/s10750-016-3002-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-016-3002-5