
Abstract
The relationship between host density and parasitism depends on a parasite’s life history. The abundance of a directly transmitted contagious parasite should increase with host density, whereas the abundance of a directly transmitted parasite that seeks its host might decrease due to the encounter-dilution effect. For parasites with complex life cycles, previous studies have found no association between parasite abundance and host density. We tested the relationship between host density and metacercarial abundance of a trematode parasite (Posthodiplostomum minimum) in two species of centrarchid fishes (Lepomis macrochirus and L. auritus) from eight small creeks. We found that host density was negatively associated with parasite abundance. Thus, our study represents the first evidence of the encounter-dilution effect for a parasite with complex life cycle in a natural system. We also report a positive association between total P. minimum population abundance and Lepomis spp. density, indicating that at low host density, cercarial mortality could moderate the encounter-dilution effect.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
How should parasitism change with host density? Epidemiological theory predicts that the number of individuals of a particular parasite on a host individual (infrapopulation abundance; Bush et al., 1997) should increase asymptotically with host density, because contact rate between hosts and transmission stages increases with host density (Anderson & May, 1978; May & Anderson, 1978; Dobson, 1990). This prediction works well for directly transmitted contagious parasites (i.e., those that are transmitted via close proximity among hosts or via infected feces; Côté & Poulin, 1995; Arneberg, 2001; Rifkin et al., 2012). For example, in mammals, the average number of strongylid nematodes per host increases with host population density (Arneberg et al., 1998). Conversely, the encounter-dilution effect, defined as diminishing risk of parasitism with increasing host group size or population density (Mooring & Hart, 1992; Côté & Poulin, 1995; Rifkin et al., 2012), predicts that the number of parasites per host should be negatively correlated with host density. Under this scenario, the total number of transmission stages is divided among all hosts in an area, reducing the number of parasites infecting each individual (Turner & Pitcher, 1986; Mooring & Hart, 1992). The encounter-dilution effect applies well to directly transmitted mobile parasites that seek their hosts (Côté & Poulin, 1995; Rifkin et al., 2012), because for mobile parasites, transmission stage production is spatially decoupled from infection risk. Much evidence for the encounter-dilution effect comes from studies of social animals and their mobile parasites. For example, Fauchald et al. (2007) found that the number of larval warble flies per reindeer calf decreased with herd size.
In contrast to the above predictions, for parasites with complex life cycles, previous studies have demonstrated or postulated a weak association or no association between parasites per host and host density (Moore et al., 1988; Côté & Poulin, 1995; Arneberg, 2001). In this case, the production of transmission stages occurs in an upstream host, and is therefore largely independent of the density of the host that they infect, except at very large spatial scales. It has been argued that time lags and population dynamics of intermediate host(s) preclude simple relationships between parasitism and host density (Moore et al., 1988). However, many parasites with complex life cycles have mobile transmission stages that actively seek hosts. Production of these transmission stages in one host is necessarily decoupled from transmission to the next host in the life cycle. For a given number of these transmission stages in the environment, additional host individuals could dilute infection risk, because transmission stages are divided among more hosts when hosts exist at high density (Turner & Pitcher, 1986; Mooring & Hart, 1992). However, evidence for the encounter-dilution effect for parasites with complex life cycles is limited (Côté & Poulin, 1995; but see Johnson et al., 2012; Stumbo et al., 2012), especially in natural systems.
Although the encounter-dilution effect might be predicted to apply to parasites with complex life cycles and mobile transmission stages, mortality of free living stages at low host density could diminish or even reverse a negative correlation between host density and parasite abundance. Without transmission stage mortality, theory predicts no association between host density and the total parasite population in an area (component population; Bush et al., 1997), because every transmission stage successfully infects a host. However, most parasites produce far more transmission stages than will ever reach hosts. If transmission stages that do not find a host die, then the success rate of transmission stages should increase with host density, leading to a positive relationship between host density and the total parasite population.
In this study, we tested the association between host density and parasitism. Because the parasite we investigated has a complex life cycle and a mobile transmission stage, we predicted that parasite abundance would be negatively related to host density due to an encounter-dilution effect. Furthermore, we predicted that the encounter-dilution effect would be moderated by mortality of transmission stages at low host densities, as indicated by a positive correlation between host density and total parasite population abundance.
Materials and methods
Study species
Posthodiplostomum minimum (MacCallum, 1921) is a trematode from the family Diplostomatidae with a 3-host life cycle (Miller, 1954). Miracidia hatch from eggs and penetrate first intermediate hosts, snails of the genus Physa, in which sporocysts develop. Once mature, cercariae leave the snail and penetrate the second intermediate host, fishes in the families Cyprinidae and Centrarchidae. Although P. minimum might actually represent several physiological strains or subspecies, or a species complex (Hoffman, 1958; Bedinger & Meade, 1967), currently recognized subspecies are P. minimum minimum (Hoffman, 1958), which is specific to cyprinids, and P. minimum centrarchi (Hoffman, 1958), which is specific to centrarchids (Hunter & Hunter, 1940; Klak, 1940; Ferguson, 1943; Miller, 1954; Hoffman, 1958; Avault Jr. & Smitherman, 1965; Bedinger & Meade, 1967). Based on experimental and observational studies, the latter might specialize on members of the genus Lepomis (Avault Jr. & Smitherman, 1965; Lane et al., 2015). After cercariae penetrate the fish, they encyst as metacercariae in the heart, liver, kidneys, spleen, gonads, and mesenteries (Miller, 1954). Upon consumption of the infected fish by the final host (piscivorous birds, commonly of the family Ardeidae), metacercariae excyst and mature in the bird intestine (Miller, 1954). Eggs are passed into the environment in the feces of the final host.
Bluegill (Lepomis macrochirus; Rafinesque, 1819) and redbreast sunfish (Lepomis auritus; Linnaeus, 1758) are centrarchid fishes native to eastern North America that inhabit diverse water bodies. Both species are second intermediate hosts of many trematode parasites including P. minimum (Hoffman, 1967). Infection prevalence in Lepomis spp. is often 100%, and mean intensity can exceed 100 metacercariae per fish (Lane et al., 2015). Infection intensity can negatively impact physiological performance (Lutterschmidt et al., 2007).
Fish sampling
We use a dataset from a published study, Martin and Lutterschmidt (2013), which investigated fish biodiversity in eight small southward-flowing creeks (Lindsey, Cooper, Flatrock, Bull, Dozier, Randall, Kendall, and Baker) within the Bull and Upatoi creeks watershed (Apalachicola-Chattahoochee-Flint river basin), Muscogee County, Georgia. To quantify fish density, Martin and Lutterschmidt (2013) seined three pools within each creek during June 2001 and August 2002. Five consecutive seine (4.6 m × 1.2 m with 5 mm mesh) hauls across each pool (average reach length = 21.5 m) standardized sampling for density estimates. Stream runs and riffles were not sampled to avoid collection of certain darter species (Percidae) that are state and/or federally listed species of concern. All fishes collected were preserved in a 10% formalin solution, washed, and transferred to 70% ethanol, identified to species (Mettee et al., 1996; Pflieger, 1997; Ross, 2002), and deposited in the Sam Houston State Vertebrate Museum (SHSVM) for archival.
For our study, we quantified relative fish density in four ways: A) “Fish density” is the total number of fishes captured divided by the creek reach length seined. Next, we used Parasite Niche Modeler (PaNic) (Strona & Lafferty, 2012) to identify known hosts of P. minimum from our fish sample. B) “Host density” is the total number of potential P. minimum hosts (as identified by PaNic) divided by the creek reach length seined. The subspecies of P. minimum that infects centrarchid fishes (P. minimum centrarchi) does not infect cyprinids (Hunter & Hunter, 1940; Klak, 1940; Ferguson, 1943; Miller, 1954; Hoffman, 1958; Avault Jr. & Smitherman, 1965; Bedinger & Meade, 1967). Therefore, we next quantified C) “centrarchid density” by dividing the total number of centrarchid fishes captured by the creek reach length seined. Lastly, P. minimum centrarchi might specialize on Lepomis (Avault Jr. & Smitherman, 1965; Lane et al., 2015). Therefore, we quantified D) “Lepomis density” by dividing the total number of Lepomis captured by the creek reach length seined. For each category, we automatically considered all species in the next most specific category to be members.
Parasite sampling
We use a dataset from a published study, Anderson et al. (2015), which investigated parasite biodiversity in Lepomis fishes in the same eight creeks. To quantify P. minimum in hosts across creeks, Anderson et al. (2015) collected 222 bluegill (Lepomis macrochirus) and 205 redbreast sunfish (Lepomis auritus) during February (winter sample) and August (summer sample) 2002. Fish were collected using a backpack electrofisher (Smith-Root® Inc.) and were transported live to a field laboratory and euthanized. Each fish was identified to species and measured for standard length (mm) prior to dissection. Fish that were not dissected immediately were frozen for transport and later dissection at Sam Houston State University, Huntsville, Texas. Anderson et al. (2015) quantified the numerically dominant trematode, P. minimum, recovered from the visceral organs (heart, liver, kidneys, spleen, gonads) and mesenteries. A full description of dissection methods and other parasites recovered from these fishes are presented in Anderson et al. (2015). Parasites per host consisted of the number of P. minimum metacercariae recovered from all tissues of fishes (n = 427). To obtain estimates of total parasite population abundances of P. minimum within each creek (n = 8), we multiplied mean P. minimum per host measured for a creek by the relative Lepomis density estimated for that creek.
Statistics
First, we investigated associations between fish density and parasite abundance. Using generalized linear mixed-effects models with creek as a random effect, we tested for an association between P. minimum metacercariae per host and four measures of potential host density: (A) fish density, (B) host density, (C) centrarchid density, and (D) Lepomis density. To account for overdispersion in the response variable, which is common for parasite abundance data (e.g., Crofton, 1971), we used a negative binomial distribution and statistically controlled for season, host species, and fish standardized length. Although we tested multiple hypotheses for this analysis, we did not employ a correction for multiple comparisons because the hypotheses were not independent (i.e., the categories were nested). However, we determined that the most conservative approach, a Bonferroni correction, would not have changed the results.
Next, we investigated the association between fish density and parasite success (transmission). Using a linear model, we tested for an association between Lepomis spp. density and total P. minimum population abundance. For this analysis, each creek was treated as a replicate (n = 8). All statistical analyses were conducted in R (version 3.2.2), and we used the lme4 package to construct generalized linear mixed-effects models.
Results
Martin and Lutterschmidt (2013) reported 2407 fishes of 34 species from the eight study creeks (Table 1). Variance in fish density among creeks was ~2.5 times higher than average variance among pools within a creek. Parasite Niche Modeler (PaNic) identified 861 individuals of 13 species (36% of original sample) as known hosts of P. minimum. Of these, 776 individuals of 10 species (32% of original sample) were centrarchids, and 659 individuals of 7 species (27% of original sample) were Lepomis spp. Mean standardized length (±SE) of captured Lepomis macrochirus was 46.81 ± 0.65 mm and mean standardized length of captured Lepomis auritus was 63.36 ± 1.60 mm.
Anderson et al. (2015) collected and dissected 222 L. macrochirus and 205 L. auritus from the eight creeks over the two sampling occasions (Table 2). Mean standardized length (±SE) of dissected L. macrochirus was 84.6 ± 1.4 mm and mean standardized length (±SE) of dissected L. auritus was 88.4 ± 1.6 mm. Anderson et al. (2015) recovered 26,742 P. minimum metacercariae (mean ± SE: 62.6 ± 4.85) from the 427 individual hosts dissected.
First, we sequentially restricted estimates of host density and investigated its relationship with parasite abundance. Our generalized linear mixed-effects models revealed no association between P. minimum per host and fish density (P = 0.267; Table 3A; Fig. 1A). However, we found negative associations between P. minimum per host and host density (P < 0.001; Table 3B; Fig. 1B), centrarchid density (P < 0.001; Table 3C; Fig. 1C), and Lepomis density (P < 0.001; Table 3D; Fig. 1D). Estimates of parasites per host at the lowest Lepomis densities measured were approximately six times higher than estimates of parasites per host at the highest Lepomis densities measured. In each of these four models, all covariates were significant predictors of P. minimum per host (Table 3). On average, fishes captured during the summer sample were infected with more than twice as many P. minimum metacercariae as fishes captured during the winter sample (P < 0.034 in all analyses; mean ± SE: summer = 89.6 ± 7.9, winter = 41.8 ± 5.7). On average, L. macrochirus were infected with 33% more P. minimum metacercariae than L. auritus (P < 0.001 in all analyses; mean ± SE: L. macrochirus = 71.2 ± 7.35, L. auritus = 53.3 ± 6.18). Standardized length was positively associated with P. minimum abundance (P < 0.001 in all analyses), with an average of an additional 2.2 metacercariae per millimeter of fish.

Partial residual plots showing the association between P. minimum per host and (A) fish density, (B) host density, (C) centrarchid density, and (D) Lepomis density
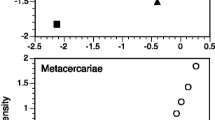
Next, we tested for an association between total P. minimum population abundance and Lepomis spp. density. Our linear model revealed a positive relationship between estimates of total P. minimum population abundance and Lepomis density (P = 0.017; Table 4; Fig. 2). Estimates of total parasite population abundance were approximately four times higher at the highest Lepomis densities measured than at the lowest Lepomis densities measured.

The association between total P. minimum population abundance and Lepomis density
Discussion
Whereas previous studies have demonstrated or postulated a weak association or no association between parasite abundance and host density for parasites with complex life cycles (Moore et al., 1988; Côté & Poulin, 1995; Arneberg, 2001), we found that parasite abundance was negatively associated with host density, presumably due to an encounter-dilution effect. Although we agree that a positive association between host density and parasite abundance is unlikely for parasites with complex life cycles (Moore et al., 1988; Côté & Poulin, 1995; Arneberg, 2001), we posit that a negative association is a logical outcome for certain stages in the life cycle. Mathematical models and empirical evidence support the encounter-dilution effect for snails infected by trematode miracidia (Anderson, 1978; Johnson et al., 2012), and for fish infected by trematode cercariae (Stumbo et al., 2012). To our knowledge, however, our study represents the first evidence of the encounter-dilution effect for a parasite with complex life cycle in a natural system.
We stress that the encounter-dilution effect results from decoupling of transmission stage production and infection risk. For directly transmitted parasites, decoupling can be achieved via a mobile transmission stage (e.g., warble flies). For parasites with complex life cycles, the existence of multiple hosts results in decoupling; the supply of transmission stages to an area depends more heavily on density of the upstream host than on density of the downstream host. Furthermore, these transmission stages are often mobile (e.g., cercariae), and could also be transported passively (e.g., via stream flow), augmenting decoupling. Moreover, if a host in the life cycle is particularly mobile (e.g., birds), then decoupling might be further enhanced. Decoupling also depends on spatial scale. At a scale relevant to infection risk for an individual host, decoupling predicts the encounter-dilution effect. At spatial scales larger than the range of transmission stages or the hosts that transport them, however, the supply of transmission stages might depend on the density of hosts that they infect. Thus, the encounter-dilution effect depends on scale.
Despite our evidence for the encounter-dilution effect, a positive relationship between Lepomis density and total parasite population abundance suggests that at low host densities, the encounter-dilution effect could be moderated by cercarial mortality. At high host densities, we found relatively high total parasite population abundance, presumably because the chances of each infective stage successfully contacting a host were improved. That our results for parasites per host and total parasite population oppose one another highlights the importance of considering parasitism from both the perspective of the host and the parasite (Anderson, 1978; Sonnenholzner et al., 2011). High host density represents a win–win situation for both the parasite and the host; the host benefits through encounter-dilution and the parasite benefits through reduced infective stage mortality. However, increased parasite transmission at high host density could lead to increased production of transmission stages in future generations, making the encounter-dilution effect temporally transient.
Although the encounter-dilution effect provides a plausible proximate explanation for our results, it does not explain why fish density varied between the eight creeks in our study area. Using the same dataset we used to quantify fish density in these creeks, Martin and Lutterschmidt (2013) found that fish abundance was negatively related to urban land use. Thus, urbanization might provide an ultimate explanation for our results. However, the correlational nature of our study makes possible several alternative explanations for the observed patterns. For example, urban land use could have increased infected snail density, thus increasing cercarial production in urban creeks. We did not measure density of snails or cercariae, and therefore cannot rule out this possibility. However, under this scenario, we would predict a positive association between land use and parasite abundance in Lepomis fishes. Using land use data from Martin and Lutterschmidt (2013), we tested this hypothesis and found no evidence to support it (analyses not shown). Another possibility is that fish density and infected snail density were controlled by an unmeasured factor. However, whereas several factors (e.g., productivity) might be predicted to drive a positive association between these variables, the observed pattern would require a negative association.
Predictions about the relationship between parasite abundance and host density depend on host specificity. Nonsuitable hosts could reduce parasitism by acting as decoys, diverting some proportion of transmission stages away from suitable hosts (Christensen et al., 1976). Alternatively, if transmission stages are able to avoid nonsuitable hosts, then density of nonsuitable hosts is unlikely to alter parasite abundance. Because we sequentially restricted host density estimates and tested for associations with parasite abundance, our results can be used to examine the host specificity of P. minimum centrarchi. Lane et al. (2015) suggest that P. minimum centrarchi probably specializes on Lepomis fishes. We found that P. minimum per host in two Lepomis species was negatively correlated with host density, centrarchid density, and Lepomis density, but not with the density of all fishes. Thus, our results support and extend their conclusion, suggesting that non-Lepomis fishes infected by P. minimum centrarchi and P. minimum minimum might serve as decoy hosts of P. minimum centrarchi, but that fishes not infected by P. minimum do not.
Conclusion
We found that the abundance of P. minimum metacercariae in two species of Lepomis fishes decreased with host density, presumably due to the encounter-dilution effect. We also report a positive association between host density and total P. minimum population abundance, indicating that the parasite also benefits from elevated host density. Our study represents the first evidence of the encounter-dilution effect for a parasite with a complex life cycle in a natural system.
References
Anderson, R. M., 1978. Population dynamics of snail infection by miracidia. Parasitology 77: 201–224.
Anderson, R. M. & R. M. May, 1978. Regulation and stability of host-parasite population interactions: I. Regulatory processes. Journal of Animal Ecology 47: 219–247.
Anderson, S. M., R. A. Fiorillo, T. J. Cook & W. I. Lutterschmidt, 2015. Helminth parasites of two species of Lepomis (Osteichthyes: Centrarchidae) from an urban watershed and their potential use in environmental monitoring. Georgia Journal of Science 73: 123–135.
Arneberg, P., 2001. An ecological law and its macroecological consequences as revealed by studies of relationships between host densities and parasite prevalence. Ecography 24: 352–358.
Arneberg, P., A. Skorping, B. Grenfell & A. F. Read, 1998. Host densities as determinants of abundance in parasite communities. Proceedings of the Royal Society B: Biological Sciences 265: 1283–1289.
Avault Jr., J. W. & R. O. Smitherman, 1965. Experimental host specificity of Posthodiplostomum minimum. Experimental Parasitology 17: 268–270.
Bedinger Jr., C. A. & T. G. Meade, 1967. Biology of a new cercaria for Posthodiplostomum minimum (Trematoda: Diplostomidae). The Journal of Parasitology 53: 985–988.
Bush, A. O., K. D. Lafferty, J. M. Lotz & A. W. Shostak, 1997. Parasitology meets ecology on its own terms: Margolis et al. revisited. The Journal of Parasitology 83: 575–583.
Christensen, N. O., P. Nansen & F. Frandsen, 1976. Molluscs interfering with the capacity of Fasciola hepatica miracidia to infect Lymnaea truncatula. Parasitology 73: 161–167.
Côté, I. M. & R. Poulin, 1995. Parasitism and group-size in social animals: a metaanalysis. Behavioral Ecology 6: 159–165.
Crofton, H. D., 1971. A quantitative approach to parasitism. Parasitology 62: 179–193.
Dobson, A. P., 1990. Models for multi-species parasite-host communities. In Esch, G. W., A. O. Bush & J. M. Aho (eds), Parasite communities: patterns and processes. Chapman and Hall, London: 261–288.
Fauchald, P., R. Rodven, B.-J. Bardsen, K. Langeland, T. Tveraa, N. G. Yoccoz & R. A. Ims, 2007. Escaping parasitism in the selfish herd: age, size and density-dependent warble fly infestation in reindeer. Oikos 116: 491–499.
Ferguson, M. S., 1943. Experimental studies on the fish hosts of Posthodiplostomum minimum (Trematoda: Strigeida). The Journal of Parasitology 29: 350–353.
Hoffman, G. L., 1958. Experimental studies on the cercaria and metacercaria of a strigeoid trematode, Posthodiplostomum minimum. Experimental Parasitology 7: 23–50.
Hoffman, G. L., 1967. Parasites of North American Freshwater Fishes. University of California Press, Berkeley.
Hunter III, G. W. & W. S. Hunter, 1940. Studies on the development of the metacercaria and the nature of the cyst of Posthodiplostomum minimum (MacCallum 1921) (Trematoda; Strigeata). Transactions of the American Microscopical Society 59: 52–63.
Johnson, P. T. J., D. L. Preston, J. T. Hoverman, J. S. Henderson, S. H. Paull, K. L. D. Richgels & M. D. Redmond, 2012. Species diversity reduces parasite infection through cross-generational effects on host abundance. Ecology 93: 56–64.
Klak, G. E., 1940. Neascus infestation of black-head, blunt-nosed, and other forage minnows. Transactions of the American Fisheries Society 69: 273–278.
Lane, B., T. Spier, J. Wiederholt & S. Meagher, 2015. Host specificity of a parasitic fluke: is Posthodiplostomum minimum a centrarchid-infecting generalist or specialist? The Journal of Parasitology 101: 6–17.
Lutterschmidt, W. I., J. F. Schaefer & R. A. Fiorillo, 2007. The ecological significance of helminth endoparasites on the physiological performance of two sympatric fishes. Comparative Parasitology 74: 194–203.
Martin, S. L. & W. I. Lutterschmidt, 2013. A checklist to the common cyprinid and centrarchid fishes of the Bull and Upatoi creeks watershed of Georgia with a brief glimpse of correlative urban influences and land use. Southeastern Naturalist 12: 769–780.
May, R. M. & R. M. Anderson, 1978. Regulation and stability of host-parasite population interactions: II. Destabilizing processes. Journal of Animal Ecology 47: 249–267.
Mettee, M. F., P. E. O’Neil & J. M. Pierson, 1996. Fishes of Alabama and the Mobile Basin. Oxmoor House, Birmingham, Ala.
Miller, J. H., 1954. Studies on the life history of Posthodiplostomum minimum (MacCallum 1921). The Journal of Parasitology 40: 255–270.
Moore, J., D. Simberloff & M. Freehling, 1988. Relationships between bobwhite quail social-group size and intestinal helminth parasitism. American Naturalist 131: 22–32.
Mooring, M. S. & B. L. Hart, 1992. Animal grouping for protection from parasites: selfish herd and encounter-dilution effects. Behaviour 123: 173–193.
Page, L. M. & B. M. Burr, 1991. A Field Guide to Freshwater Fishes of North America North of Mexico. Houghton Mifflin Company, Boston, Massachusetts.
Pflieger, W. L., 1997. The Fishes of Missouri. Missouri Dept. of Conservation, Jefferson City.
Rifkin, J. L., C. L. Nunn & L. Z. Garamszegi, 2012. Do animals living in larger groups experience greater parasitism? A meta-analysis. American Naturalist 180: 70–82.
Ross, S. T., 2002. Inland Fishes of Mississippi. University Press of Mississippi, Jackson.
Sonnenholzner, J. I., K. D. Lafferty & L. B. Ladah, 2011. Food webs and fishing affect parasitism of the sea urchin Eucidaris galapagensis in the Galapagos. Ecology 92: 2276–2284.
Strona, G. & K. D. Lafferty, 2012. How to catch a parasite: Parasite Niche Modeler (PaNic) meets Fishbase. Ecography 35: 481–486.
Stumbo, A. D., C. T. James, C. P. Goater & B. D. Wisenden, 2012. Shoaling as an antiparasite defence in minnows (Pimephales promelas) exposed to trematode cercariae. Journal of Animal Ecology 81: 1319–1326.
Turner, G. F. & T. J. Pitcher, 1986. Attack abatement: a model for group protection by combined avoidance and dilution. The American Naturalist 128: 228–240.
Acknowledgments
We thank S.M. Anderson, R.A. Fiorillo L. Dent, D.S. Millican, and D.K. Wasco for assistance related to this research. We thank K.D. Lafferty, A.M. Kuris, A. Smith-Herron, T.J. Cook, and two anonymous reviewers for useful discussions and comments on the manuscript. This work was supported by a grant from the Engineer Research and Development Center (ERDC) and the Civil Engineering Research Lab (CERL) of the US Army Corps of Engineers (ERDC-CERL Contract #DACA 42-00-C-0047) to W.I.L. We acknowledge the assistance provided by Gordon A. Plishker and all personnel of the Texas Research Institute for Environmental Studies (TRIES) at Sam Houston State University. We also thank Hugh Westbury of the Strategic Environmental Research and Development Program (SERDP), Ecosystem Management Project (SEMP), and Harold (Hal) Balbach of the US Army ERDC and CERL for their sincere interest in and support of this project.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Lee B. Kats.
Rights and permissions
About this article

Cite this article
Buck, J.C., Lutterschmidt, W.I. Parasite abundance decreases with host density: evidence of the encounter-dilution effect for a parasite with a complex life cycle. Hydrobiologia 784, 201–210 (2017). https://doi.org/10.1007/s10750-016-2874-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-016-2874-8