
Abstract
In New Zealand, dogs appear in the earliest archaeological contexts and were a common sight in the Māori villages described by European explorers in the late eighteenth century. In this review, we argue that the fundamental nature of dog–human relationships remained stable throughout New Zealand’s early history, despite major environmental change and transformations in Māori settlement patterns, subsistence systems, and social organisation. Although there are variations in dog abundance levels in the archaeological record through time, these are likely due to differences in site function rather than dogs’ diminishing importance or changing status. Dogs are unique in early New Zealand as the only animal that shared living space with humans. They did so as free-ranging populations; travelling with people, interacting in village life, and playing diverse roles in Māori social, technological, and ritual systems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The colonisation of the Southwest Pacific by the ancestors of the Polynesian peoples took place over a period of 3,000 years and involved movement east into increasingly biogeographically and geologically impoverished environments (Kirch, 2017). For this reason, the migrants needed to transport most of their subsistence system with them, including their horticultural plants and domestic animals. Dogs (Canis familiaris) accompanied people in these oceanic migrations and eventually reached the limits of human expansion in the Pacific: the Hawaiian Islands to the northeast and New Zealand and some of its sub-Antarctic islands to the south. Throughout this range, dog bones in archaeological sites reflect their widespread use as food– a practice that continues in parts of Polynesia to this day (Davidson, 1984:129). In New Zealand, dog bones appear in the earliest archaeological contexts, and were a common sight in the Māori villages described by the European explorers in the late eighteenth century.
In this paper we draw on the literature in archaeology, traditional and historical sources, molecular biology, and ecology to develop an archaeology of the kurī (the indigenous Māori dog, Canis familiaris). After briefly describing the environmental setting, we look at the genetic evidence for the founding population of dogs and review the archaeological evidence for dogs in pre-contact New Zealand. The pre-contact period refers to the period of New Zealand history (circa 1300 to 1770 AD) prior to sustained interactions between Māori and Europeans. We argue that the fundamental nature of dog–human relationships has remained stable for the duration of New Zealand’s early history despite major environmental change, and transformations in Māori settlement patterns, subsistence systems and social organisation. This contrasts with a common interpretation of the archaeological record that there was a decline in the abundance and significance of dogs in Māori communities through time. We also show that while dogs have performed a wide range of roles in Māori society, they have always lived as free-ranging populations.
Environmental Setting
Aotearoa New Zealand has the distinction of being the last major land mass to be colonised by humans when migrants from a central East Polynesian homeland arrived around the turn of the fourteenth century AD (Walter et al., 2017). They introduced two commensal animal species, the dog and the rat (Rattus exulans), as well as a variety of plant crops. New Zealand provided a land of opportunity and challenge for the first humans. In the absence of mammalian predators, birds had evolved to inhabit a wide range of environmental niches. Naïve, easily hunted taxa included megafauna such as the giant flightless moa (Dinornithiformes), plus strandline sea mammals (seals and sea lions). In addition, there were waterfowl, many species of parrot, pigeons and penguins, and other flightless birds, and colonial ground-nesting seabirds. In total 85% to 90% of the country was covered in forest, and continental geological processes had resulted in the formation of a diversity of stone resources ideal for tool manufacture. The arrival of the Polynesian settlers with their commensal plants and animals kicked off a cascade of environmental changes. These included extinctions, extirpations, and range restrictions of terrestrial and marine avifauna and marine mammals, plus changes in forest cover.
Kurī bones are found in virtually all archaeological sites throughout the country where midden has been recovered (Davidson, 1984:129). Usually disarticulated and sometimes with cut marks, they are normally interpreted as food waste. However, kurī bones and teeth were also used for industrial purposes, and traditional histories and later European observations show that kurī pelts were used to make clothing (Greig et al., 2015). In addition to their role as an economic resource, kurī bones have been found buried under house posts and as complete burials suggesting their occasional role in ritual contexts (Clark, 1996). Dogs also featured in Māori art forms including rock drawings and wood carving (Anderson, 1981:17).
Although it is no longer extant in New Zealand, descriptions of kurī by the first European visitors paint a consistent picture of a small fox-like animal with straight ears and a thick tail (Colenso, 1877). Analysis of kurī skeletons suggests an average height at shoulder of 390 mm and a body weight of 14.5 kg (Clark, 1997). The dominant colours were black and white, either singly or in patchy combinations, although yellow coats were also recorded (Clark & Greig, 2021:279–280). Accounts are also consistent about their use as food (often as a ceremonial food) and sometimes as a hunting animal (Anderson, 1981; Davidson, 1984:129).
Founding Dog Population
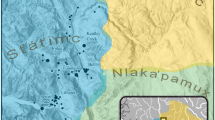
The apparent abundance of dog bones in pre-contact New Zealand archaeological sites has long been noted as a characteristic of colonisation era settlements. For example, sites such as Wairau Bar, Houhora, and Shag River Mouth (Fig. 1) all have relatively high numbers of dog bones and were all occupied within a generation or so of colonisation (Anderson et al., 1996; Furey, 2002; Greig et al., 2018b). Based on this observation it could be expected that commensurately high numbers of dogs were brought to New Zealand. In fact, we might expect the structure of the founding dog population to resemble the founding human population, which can be characterised as numerous and genetically diverse. For example, variability in modern Māori mitochondrial DNA (mtDNA) indicates that the founding colony included at least 200 women (Whyte et al., 2005). Furthermore, the DNA results from four first- or second-generation settlers buried at Wairau Bar suggest that the first colonists were not from a single community but hailed from different populations (probably different islands) within an East Polynesian homeland zone encompassing several archipelagos (Knapp et al., 2012; Walter et al., 2017). In contrast to the findings for people, the results of mitochondrial genome studies of kurī from colonisation era sites show that their mtDNA diversity is extremely limited. Thirty-four complete, or nearly complete, ancient mitogenomes from dog teeth sampled from four colonisation era archaeological sites show a striking lack of mitochondrial genetic diversity, with no observable geographic patterning (Greig et al., 2018a). In contrast, the only other commensal animal brought to New Zealand, the Polynesian rat, shows the same levels of wide genetic diversity as the people (Matisoo-Smith & Robins, 2004).
Molecular methods offer other insights into the pre-contact dog population. Stable isotope analysis of five dog specimens from Wairau Bar reveal strontium values consistent with the local environment (Kinaston et al., 2013), showing that they were born, lived, and died in the region. Strontium studies of human remains from Wairau Bar indicate far greater environmental diversity (Kinaston et al., 2013).
In summary, the molecular data show that a small number of dogs were successfully introduced into New Zealand from tropical East Polynesia in the early fourteenth century and were very quickly distributed around the country, either accompanying the dispersal of people or via exchange practices. This is entirely consistent with the evidence of human mobility patterns in the first decades of settlement (Walter et al., 2017). If the Wairau Bar isotope data are consistent with the broader patterns, it is likely that dog mobility became more restricted once they were regionally established.

Map of New Zealand showing the location of sites mentioned in the text and the Greater Hauraki and Otago-Catlins study areas (after Smith & James-Lee, 2010)
Dogs in New Zealand Archaeology
Avocational archaeology in New Zealand commenced in the late nineteenth century, with professional archaeology following the appointment of the first qualified archaeologists in the early 1960s (Campbell, 2004). More than a century of archaeological investigation has produced a large archaeofaunal data set scattered across three minimally overlapping bodies of literature. Many of the country’s most diverse, midden-rich sites were excavated before 1960 using unsystematic or undescribed methodologies. These efforts produced a vast quantity of material, but little analysis or record keeping. A second body of data contained within the unpublished reports of cultural resource management archaeology is probably the largest set available. We have drawn on this literature where we can, and on a comprehensive attempt at synthesis by Smith and James-Lee (2010). But the collection and analysis methods employed by many of the cultural resource management teams were either undescribed, unreported, or so varied that the value of a comparative analysis is limited. By far the best source of data is the work of university-based researchers. This includes some good descriptions of dog assemblages (e.g., Irwin, 2004; Smith, 1981, 1996; Taylor & Irwin, 2008) and some focussed dissertations on the topic (Allo, 1970; Clark, 1995; Greig, 2017; Pillay, 2020). Unfortunately, this is by far the smallest body of data on the archaeology of dogs. The following summary draws on all three sources of information but suffers the limitations implied above, and especially in the availability of reliable quantitative data.
Several archaeological review articles have suggested that there was a decline in the frequency and relative abundance of kurī through time (Allo Bay-Petersen, 1979; Allo, 1970; Bay-Petersen, 1983:126; Davidson, 1984:129; Irwin, 2008:203). Oddly, this coincides with a dramatic decline in the availability of meat resources in general. The first settlers relied heavily on the hunting of sea mammals and avifauna but there was a significant wave of terrestrial extinctions within a century of human arrival and sea mammals were extirpated over much of their range around the same time (Holdaway & Jacomb, 2000; Smith, 2011). Given this loss of protein resources, it seems counter-intuitive for domestic animals to have become less important too, and several explanations have been extended to explain the apparent contradiction. For example, the decline in wild meat sources may have increased competition between people and dogs making the cost of supporting dogs too high (Allo Bay-Petersen, 1979). Such competition might be associated with an observed decrease in kurī limb bone dimensions (Clark, 1997). However, there is no indication that the decline in wild meat resources resulted in changes in kurī health. Pillay’s (2020) analysis of kurī dentition from North Island sites spanning the pre-contact period indicated that overall kurī maintained good health, with very few dental pathologies. Allo (1970) also found few instances of caries, trauma, or abscesses in kurī dentition, although she did note the differing incidence of periodontal disease and associated tooth wear and attrition in dogs from North versus South Island sites. This could be related to diet and the amount of softer vegetable foods consumed by North Island kurī. Clark (1997) and Pillay (2020) also observed an increase in kurī tooth wear in later sites, most likely derived from the shift to a marine diet containing a high proportion of sand and grit. This mirrors the changing diet of humans, which also moved towards a grittier diet because of an increasing reliance on sandy-shore shellfish and fern-root through time (Houghton, 1980:124).
Evaluating the claim for a decrease in the importance of dogs is problematic; there are few sites in New Zealand with long occupation sequences so investigating change through time usually relies on building sequences at a regional level. There are two regions of New Zealand where there appears to be sufficient dog data to undertake such a study: Greater Hauraki and Otago-Catlins (Smith & James-Lee, 2010. The Greater Hauraki study area lies on the upper east coast of the North Island, while the Otago-Catlins study area extends along the east coast of the lower part of the South Island (Fig. 1). Smith and James-Lee (2010:2) assigned sites in these regions to three broad chronological periods based on radiocarbon chronology: Early (c. 1250–1450 AD), Middle (1450–1650 AD), and Late (1650–1800 AD).Footnote 1 In cases where the radiocarbon data overlapped these period boundaries, assemblages were classified as either Early/Middle, or Middle/Late (Smith & James-Lee, 2010:2). In the regional reviews below we draw on the summary information provided in Smith and James-Lee (2010: Appendix 9) of that report. This review uses MNI (Minimum Number of Individuals) because raw data and NISP (Number of Identified Specimens) were rarely reported in the unpublished reports relied on by Smith and James-Lee (2010). Many unpublished reports also fail to comment on matters such as cut marks or other taphonomic indicators.
Consistent with claims of earlier studies (above) in both study areas the average number of dogs in sites appears to be highest in the Early Period and falls off through time (Smith & James-Lee, 2010:Appendix 9, Fig. 2).

Graphs showing dog MNI for each of the study regions organised by time period. Number of assemblages shown in brackets on right-hand graphs
Otago-Catlins
The reported archaeological contexts of the assemblages in the Otago-Catlins region were extremely varied and included diverse features such as structures, hearths, cooking areas and middens. In total, dog bone was present in 31 of the 32 assemblages analysed (Fig. 2). The very high number of dogs in the Early Period assemblages is due in part to the inclusion of the Shag River Mouth site (J43/2) where an MNI value of 76 dogs was calculated (Anderson et al., 1996). This site is from the first century of settlement, spans 50 to 100 years, and is one of a few sites that extends into the period of faunal extinctions and subsistence change (Anderson et al., 1996). Analysis of meat weights for the major classes of fauna shows the impact of the depletion of moa and marine mammal resources, and a commensurate rise in importance of fish (Smith, 2005). There is a slight decline in the amount of dog meat, but overall the contribution remains low in comparison to other major meat sources.
The very small number of dogs in the Otago-Catlins data from the Early/Middle to Middle/Late period may simply reflect a hiatus in the occupation of southern New Zealand that occurred between the 1500s and mid-1700s (Davidson, 1984; Jacomb et al., 2010).
Greater Hauraki
Kurī bone is present in 29% of the analysed assemblages in the Greater Hauraki region, which contrasts with 97% of assemblages with dog bone in the Otago-Catlins region. Only 22 of the 75 assemblages in the Greater Hauraki region contain dog bone, with 15 of these coming from sites containing evidence of houses, storage pits, middens, and artefact working areas (Fig. 2). Seven are smaller short-stay camps, either solely middens, or middens and ovens, containing mainly shellfish. Most of the midden and midden and oven assemblages in the Greater Hauraki region do not contain any dog bone. Although this does appear to show a decline in dog numbers through time, we offer an alternative interpretation below.
Dogs and Villages
Although the regional studies cited above seem to show dog abundance levels decreasing with time, the evidence for a decline is equivocal; dogs were plentiful and highly regarded in early nineteenth century villages (Davidson, 1984:197) and Clark (1995) suggests that the late pre-contact archaeological record supports this. We suggest that the relevant variable in the correlation is site function not chronology. The earliest assemblages, with high relative abundance levels for dogs, are mainly from village sites occupied by sedentary communities of various sizes. The late pre-contact sites generally have lesser evidence for dog use, and smaller short-stay camps or single feature middens are more numerous (Fig. 3). While many large moa hunting settlements have been the focus of archaeological investigations, large scale systematic excavations of village sites dating to later periods are less common. There are several exceptions – late pre-contact sites with higher dog abundance level.

Graph showing dog MNI by time period for each of the study regions, by site type
Kohika (V15/80)
Kohika was occupied in the mid-seventeenth century and was situated on a small swampy island in a lake in the Bay of Plenty (Irwin, 2004) (Fig. 1). The excavations revealed evidence of dwelling structures, raised storehouses, and miscellaneous small shelters along with a diverse array of domestic tools and textile fragments. Lines of palisades were exposed along the shoreline, and canoe landing places were located in gaps in the palisading. Kohika is interpreted as a permanently occupied and lightly defended lakeside village. Excavations produced a combined MNI of 32 dogs from inside and outside the palisades (Taylor & Irwin, 2008). The bones from outside the palisades would have been deposited in the former lake and, likely due to the anaerobic environment, were well-preserved and often intact. The bones from within the village were in a dry, sandy environment, and showed evidence of weathering, human modification, and prolonged dog and rat gnawing.
Masonic Tavern Site (R11/2517)
The Masonic Tavern (Fig. 1) site was also a village settlement. Excavations revealed a diversity of domestic activities including a stone-working and tool-manufacturing zone, and another area associated with food preparation and cooking including a stone-lined hearth and scattered fire features. There were dense deposits of midden remains across the site and dog bones made up over 16% MNI of the total faunal assemblage (NISP = 2529, MNI = 289). Microscopic and aDNA analyses of coprolites showed the dogs had an omnivorous diet that included fish, shell-fish, and cultivated plants, both Pacific introductions and native species (Wood et al., 2016). The frequency of charcoal in the coprolites suggests that food had been scavenged from fire pits. Radiocarbon dating placed occupation after 1500 AD (Wood et al., 2016).
NRD Site (R11/859)
The NRD site was a small village occupied in the seventeenth century situated on the edge of Manukau Harbour in Auckland (Fig. 1). It was excavated in 2008 and 2009 to enable an extension of Auckland International Airport. The settlement included two burial grounds containing the remains of 88 people and at least four complete dog burials (Campbell & Hudson, 2011). One of the dogs was buried in a disused food storage pit, a burial method also used for some of the people. Some of the human burials also contained dog remains. Dog bone was also found within structures, midden dumps, and in contexts that suggest dog meat was consumed as part of a mortuary ritual (Campbell & Hudson, 2011:147). No MNI value is available but with a NISP value greater than 1700, the abundance levels of dog are broadly comparable with the colonisation era Shag River Mouth site (NISP = 1990, MNI = 76) (Smith, 1996:189).
Interactions Between Kurī and People
There is good evidence in the midden data that dogs were regularly eaten over the course of early New Zealand history, but both archaeological and historical records show that they fulfilled multiple roles in Māori society. In our conclusion, we draw on modern ethological studies of human–dog relationships to understand the New Zealand record. Ethological studies demonstrate the flexibility of dogs and their ability to survive in a diverse range of human communities and environments (e.g. Gompper, 2013; Miklosi, 2015; Vanak & Gompper, 2009). They also serve to highlight the ways in which dogs may co-habit space with people, often not as pets in the contemporary Western sense, but as a group sharing living spaces and resources in a domestic setting. These studies describe three general modes of interaction:
Managed Dog Populations: Dogs are actively managed by people throughout their life. Their range is artificially circumscribed, their food is supplied by people and controls are placed over reproduction. The circumstances of death may also be artificially controlled.
Free-Ranging Dog Populations: Dogs live in human environments but are not actively managed. Their food supply is mostly human derived and is obtained by foraging, although dogs may occasionally be fed. Dog populations may be culled from time to time, but otherwise their life cycles are not under direct control. Even within such a free-ranging lifestyle, some dogs may be treated preferentially to the group as a whole, being kept as companion animals or for a particular use and having a different relationship with people at some points in their lives (Hughes & Macdonald, 2013).
Feral Dog Populations: Dogs live entirely independently from humans. They scavenge or hunt in the wild and sometimes around the fringes of human settlements.
These modes of interaction can be modelled by four intersecting primary variables: diet, habitat, human constraints on mobility, and human control over demographic structure, including reproduction (Table 1).
The evidence from archaeological and historical records and molecular studies shows that kurī interactions with people are a best fit with the free-ranging mode. Molecular and microscopic analysis of dog coprolites indicate that kurī diet was made up of foods derived from human environments and activities. It comprised a wide range of plants and terrestrial and marine vertebrates, most of which were also part of the human diet (Clark, 1995; Wood et al., 2016). Dog gnawing on bones from many archaeological sites shows that kurī were free to roam in and around village, and this is confirmed in European contact period depictions and observations on village life (Dieffenbach, 1843; Earle, 1832). Villages all contained food and other types of storage structures on poles raised above the ground to protect contents from dogs (Polack, 1838:34). Oral tradition and historical accounts describe dogs travelling with people in canoes and on foot (Colenso, 1877).
On the other hand, dog remains have never been reported in palaeofaunal sites, or in cave sites or rockshelters where animal bones accumulate naturally. There are no known traditional accounts of feral dog populations or descriptions of feral dogs at the time of first European contact. This demonstrates that the distribution of kurī overlapped that of human communities for the full duration of history prior to European colonisation. By the mid-1800s however, there are reports of feral or ‘wild’ dog populations indicating changes in dog–human relationships (e.g., Barrington et al., 1864). These populations were of European dogs or possibly European-kurī crossbreds and are associated with the establishment of pastoralism (Clark, 1995).
Conclusions
The archaeological record documents the persistence of a close relationship between dogs and people in New Zealand from the colonisation period onwards. Once present, given the plentiful amounts of food available, the dog population increased rapidly, as evidenced by the numerous dog bones in colonisation era sites. Studies of free-ranging dog populations have demonstrated the close association between dog numbers and the associated human population, resource availability, and the carrying capacity of the domestic niche, and highlight the potential for dog numbers to rapidly increase under the appropriate conditions (Wandeler et al., 1988). This is what we see in colonisation era New Zealand sites. Interestingly, there is no archaeological evidence for the other two domestic animals (pigs and chickens) in use in tropical East Polynesia, ever reaching New Zealand. This lack, and the limited genetic diversity of dogs, may indicate challenges in the transportation of domesticated animals to New Zealand.
We have shown that any apparent decrease in abundance of dogs through time is more likely due to dogs being mainly associated with villages sites, where dogs are consistently present in higher numbers than in temporary camp sites. However, it is possible that the value of dogs changed through time. Ethnographic accounts suggest that dog skin clothing and adornment was restricted to those of the chiefly class by the early nineteenth century (Parkinson, 1984; Sparrman et al., 1944) and white dog hair was especially used to adorn chiefly weapons and poi (equipment used to accompany musical performance) (Colenso, 1877). The circumstances around the consumption of dogs may have changed too. The evidence from Wairau Bar demonstrates that dogs were consumed in large-scale public feasting events (Walter et al., 2017). By the early nineteenth century, however, some ethnographic accounts suggest that the consumption of dogs was more socially restricted; as a special food for chiefs and tohunga (religious or ritual specialists) (Colenso, 1877:150). Oral traditions and historical accounts also record conflicts arising from the loss of favoured dogs (ibid.).
Although dog is ubiquitous in pre-contact New Zealand and other Polynesian sites associated with food preparation and consumption, and we know many historical Polynesian communities ate dogs, treating archaeological dog bone as food remains alone will always be problematic. As Bogucki (1993:492) comments “faunal remains are archaeological data and prehistoric living animals are abstract concepts.” Dogs are unique in pre-contact New Zealand as the only animal that shared living space with humans. They did so as free-ranging populations; travelling with people, sharing food, interacting in village life, and playing roles in Māori social, technological, and ritual systems. Studies from other parts of the world that investigate the roles of dogs in human communities, both in life and at death (e.g., Hill, 2018; Kerber, 1997; Losey et al., 2018; Morey & Wiant, 1992), demonstrate the potential to develop a more nuanced understanding of the place of dogs in New Zealand's early history. To do so requires the detailed examination and reporting of dog bones in archaeological sites, and we would encourage archaeologists to take this into account in future research designs.
Notes
The archaeological record in New Zealand is short and lies within a particularly ‘wiggly’ portion of the radiocarbon calibration curve so archaeologists can only rarely generate tight and reliable radiocarbon determinations for single stratigraphic events (Hogg et al., 2019). Consequently, in regional reviews, simple periodisation like this is often necessary.
References
Allo Bay-Petersen, J. (1979). The role of the dog in the economy of the New Zealand Maori. In, Anderson, A. (ed.) Birds of a feather. Osteological and archaeological papers from the South Pacific in honour of R.J. Scarlett. BAR International Series 62, NZAA Monograph 11, pp 165–181.
Allo, J. (1970). The Maori dog - a study of the Polynesian dog in New Zealand. MA Thesis, University of Auckland.
Anderson, A. (1981). Pre-European hunting dogs in the South Island, New Zealand. New Zealand Journal of Archaeology, 3, 15–20.
Anderson, A., Allingham, B., & Smith, I. W. G. (Eds.). (1996). Shag River Mouth: The Archaeology of an Early Southern Maori Village. ANH Occasional Series Monograph
Barrington, A. J., Farrell, J., & Simonin, A. (1864). Abstract of a Prospecting Tour to the West Coast, from March 1st to June 11th. Otago Daily Times, 30 June 1864, p.6.
Bay-Petersen, J. (1983). Competition for resources: The role of pig and dog in the Polynesian agricultural economy. Journal De La Société Des Océanistes, 77, 121–129.
Bogucki, P. (1993). Animal traction and household economies in Neolithic Europe. Antiquity, 67, 492–503.
Campbell, M. (ed.). (2004). Digging into History: 50 years of the New Zealand Archaeological Association. Archaeology in New Zealand Special Issue.
Campbell, M., & Hudson, B. (2011). The NRD Site Community Report. Unpublished report.
Clark, G. (1996). Animal burials from Polynesia. Archaeology in New Zealand, 39, 30–38.
Clark, G. (1997). Osteology of the Kuri Maori: The prehistoric dog of New Zealand. Journal of Archaeological Science, 24, 113–126.
Clark, G. R. (1995). The Kuri in Prehistory: A skeletal analysis of the extinct Maori dog. MA Thesis, University of Otago.
Clark, G. R., & Greig, K. (2021). Canis familiaris. In C. M. King & D. M. Forsyth (Eds.), The handbook of New Zealand mammals (3rd ed., pp. 279–284). CSIRO Publishing.
Colenso, W. (1877). Notes, chiefly historical, on the ancient dog of the New Zealanders. Transactions and Proceedings of the Royal Society of New Zealand, 10, 135–155.
Davidson, J. M. (1984). The Prehistory of New Zealand. Longman Paul.
Dieffenbach, E. (1843). Travels in New Zealand. John Murray.
Earle, A. (1832). A Narrative of Nine Months’ Residence in New Zealand in 1827. Longman.
Furey, L. (2002). Houhora: A fourteenth century Maori village in Northland. Bulletin of the Auckland Museum No. 19, Auckland War Memorial Museum.
Gompper, M. (Ed.). (2013). Free-Ranging Dogs and Wildlife Conservation. Oxford University Press.
Greig, K. (2017). Tracking dogs across the Pacific. An archaeological and ancient DNA study. PhD thesis, University of Otago.
Greig, K., Boocock, J., Allen, M., Walter, R., & Matisoo-Smith, E. (2018a). Ancient DNA evidence for the introduction and dispersal of dogs in New Zealand. Journal of Pacific Archaeology, 9, 1–10.
Greig, K., Boocock, J., Prost, S., Horsburgh, K. A., Jacomb, C., Walter, R. & Matisoo-Smith, E. (2015). Complete Mitochondrial Genomes of New Zealand's First Dogs. PloS one, 10: e0138536.
Greig, K., Gosling, A., Collins, C. J., Boocock, J., McDonald, K., Addison, D. J., Allen, M. S., David, B., Gibbs, M., Higham, C. F. W., Liu, F., Mcniven, I. J., O’Connor, S., Tsang, C. H., Walter, R., & Matisoo-Smith, E. (2018b). Complex history of dog (Canis familiaris) origins and translocations in the Pacific revealed by ancient mitogenomes. Scientific Reports, 8, 9130.
Hill, E. (2018). The archaeology of human–dog relations in Northwest Alaska. In Losey, R.J., Wishart, R.P. and Loovers, J.P.L. (eds). Dogs in the North: stories of cooperation and co-domestication. Routledge, pp. 99–116.
Hogg, A. G., Heaton, T. J., Ramsey, C. B., Boswijk, G., Palmer, J. G., Turney, C. S. M., Southon, J., & Gumbley, W. (2019). The Influence of Calibration Curve Construction and Composition on the Accuracy and Precision of Radiocarbon Wiggle-Matching of Tree Rings, Illustrated by Southern Hemisphere Atmospheric Data Sets from AD 1500–1950. Radiocarbon, 61, 1265–1291.
Holdaway, R. N., & Jacomb, C. (2000). Rapid extinction of the moas (Aves : Dinorinthiformes): Model, test, and implications. Science, 287, 2250–2254.
Houghton, P. (1980). The First New Zealanders. Hodder & Stoughton.
Hughes, J., & Macdonald, D. W. (2013). A review of the interactions between free-roaming domestic dogs and wildlife. Biological Conservation, 157, 341–351.
Irwin, G. J. (Ed.). (2004). Kohika: The archaeology of a late Maori lake village in the Ngati Awa rohe, Bay of Plenty, New Zealand. Auckland University Press.
Irwin, G. J. (2008). A summary of archaeological investigations at Kohika, V15/80, 2004–2008. Archaeology in New Zealand, 51: 36–45.
Jacomb, C., Walter, R., & Jennings, C. (2010). A Review of the Archaeology of Foveaux Strait, New Zealand. Journal of the Polynesian Society, 119, 25–59.
Kerber, J. E. (1997). Native American treatment of dogs in Northeastern North America: Archaeological and ethnohistorical perspectives. Archaeology of Eastern North America, 25, 81–95.
Kinaston, R. L., Walter, R., Jacomb, C., Brooks, E., Tayles, N., Halcrow, S. E., Stirling, C., Reid, M., Gray, A. R., & Spinks, J. (2013). The First New Zealanders: Patterns of Diet and Mobility Revealed through Isotope Analysis. PloS one, 8: e64580.
Kirch, P. V. (2017). On the road of the winds : An archaeological history of the Pacific islands before European contact. University of California Press.
Knapp, M., Horsburgh, K. A., Prost, S., Stanton, J. A., Buckley, H. R., Walter, R. K., & Matisoo-Smith, E. A. (2012). Complete mitochondrial DNA genome sequences from the first New Zealanders. Proceedings of the National Academy of Sciences of the United States of America, 109, 18350–18354.
Losey, R. J., Nomokonova, T., Fleming, L. S., Kharinskii, A. V., Kovychev, E. V., Konstantinov, M. V., Diatchina, N. G., Sablin, M. V., & Iaroslavtseva, L. G. (2018). Buried, eaten, sacrificed: Archaeological dog remains from Trans-Baikal, Siberia. Archaeological Research in Asia, 16, 58–65.
Matisoo-Smith, E., & Robins, J. H. (2004). Origins and dispersals of Pacific peoples: Evidence from mtDNA phylogenies of the Pacific rat. Proceedings of the National Academy of Sciences, 101, 9167–9172.
Miklosi, A. (2015). Intraspecific social organization in dogs and related forms. In Dog Behaviour, Evolution, and Cognition (2cnd ed.). Oxford University Press. pp. 172–173.
Morey, D., & Wiant, M. (1992). Early Holocene Domestic Dog Burials From the North American Midwest. Current Anthropology, 33, 224–229.
Parkinson, S. (1984). Journal of a voyage to the South Seas. Caliban Books.
Pillay, P. (2020). Investigating Diet and Oral Health in the Māori Dog (Kurī) using Archaeological Assemblages from the North Island. MA Thesis, University of Auckland.
Polack, J. S. (1838). New Zealand; being a narrative of travels and adventures during a residence in that country between the years 1831 and 1837. Vol 2. R. Bentley.
Smith, I. (2005). Retreat and resilience: Fur seals and human settlement in New Zealand. In G. G. Monks (Ed.), Exploitation and Cultural Importance of Sea Mammals (pp. 6–18). Oxbow Books.
Smith, I., & James-Lee, T. (2010). Data for an Archaeozoological Analysis of Marine Resource Use in Two New Zealand Study Areas (Revised edition). Otago Archaeological Laboratory Reports No. 7. Department of Anthropology & Archaeology; University of Otago. Retrieved from http://hdl.handle.net/10523/5945
Smith, I. W. G. (1981). Mammalian fauna from an archaic site on Motutapu Island, New Zealand. Records of the Auckland Institute and Museum, 18, 95–105.
Smith, I. W. G. (1996). The Mammal Remains. In A. Anderson, I. W. G. Smith, & B. Allingham (Eds.), Shag River Mouth: The Archaeology of an Early Southern Maori Village (pp. 185–199). ANH Publications.
Smith, I. W. G. (2011). Pre-European Maori exploitation of marine resources in two New Zealand case study areas: Species range and temporal change. Journal of the Royal Society of New Zealand, 2011, 1–37.
Sparrman, A., Rutter, O., Mackenzie-Grieve, A., Beamish, H. V., & Barker-Mill, P. (1944). A voyage round the world with Captain James Cook in H.M.S. Resolution. The Golden Cockerel Press.
Taylor, G., & Irwin, G. J. (2008). The dry and the wet: The variable effects of taphonomy on the dog remains from Kohika Lake Village, Bay of Plenty, New Zealand. In G. Clark, B. F. Leach, & S. O’Connor (Eds.), Islands of Inquiry: Colonisation, Seafaring and the Archaeology of Maritime Landscapes (pp. 453–474). ANU EPress.
Vanak, A. T., & Gompper, M. E. (2009). Dogs (Canis familiaris) as carnivores: Their role and function in intraguild competition. Mammal Review, 39, 265–283.
Walter, R., Buckley, H., Jacomb, C., & Matisoo-Smith, E. (2017). Mass Migration and the Polynesian Settlement of New Zealand. Journal of World Prehistory, 30, 351–376.
Wandeler, A. I., Budde, A., Capt, S., Kappeler, A., & Matter, H. (1988). Dog ecology and dog rabies control. Reviews of Infectious Diseases, 10(S4), S684–S688.
Whyte, A. L. H., Marshall, S. J., & Chambers, G. K. (2005). Human evolution in Polynesia. Human Biology, 77, 157–177.
Wood, J. R., Crown, A., Cole, T. L., & Wilmshurst, J. M. (2016). Microscopic and ancient DNA profiling of Polynesian dog (kurī) coprolites from northern New Zealand. Journal of Archaeological Science: Reports, 6, 496–505.
Acknowledgements
We thank Luis Pacheco-Cobos and Bruce Winterhalder for the opportunity to contribute this article. We would like to acknowledge the pioneering studies of kurī carried out by Jan Bay-Petersen (née Allo) and Geoffrey Clark. Les O’Neill produced the illustrations. We would also like to thank the two anonymous reviewers for their constructive comments which greatly improved the clarity of this review.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical Approval
This study did not involve any human or animal subjects.
Conflicts of Interest/Competing Interests
The authors have no relevant financial or non-financial interests to disclose.
Informed Consent
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Greig, K., Walter, R. A Re-Assessment of the Role of Dogs (Canis familiaris) in Early Aotearoa New Zealand. Hum Ecol 49, 755–764 (2021). https://doi.org/10.1007/s10745-021-00285-2
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10745-021-00285-2