
Abstract
Dry-raising rice seedlings in nurseries is a key technique in high-yield rice cultivation. The present study of morphological and physiological indexes showed dry-raised seedlings (DRS) had a shorter stature, more developed root systems, and significantly higher soluble sugar, starch, and N content than moist-raised seedlings (MRS), resulting in significantly increased grain yield. Compared to the MRS techniques, the dry-raised measures induced higher levels of abscisic acid (ABA), gibberellins (GA3), and indole-3-acetic acid (IAA) in leaves and roots of seedlings. We then utilized tandem mass tags (TMT) quantitative proteomics technology to analyze the mechanism by which rice exposed to the appropriate drought stress (dry-raised measures) during the seedling stage develop differently. Through mass spectrometry, we identified 281 significantly expressed proteins in roots and 268 in leaves. The differentially expressed proteins were then divided into 23 categories based on MapMan ontology. In addition, the hormonal-related protein expression patterns of DRS were confirmed with RT-PCR at the transcript level. On the basis of these findings, we proposed that appropriate drought stress during the rice seedling stage can change the expression of key proteins involved in nitrogen uptake and translocation, hormone synthesis, photosynthesis, and CHO metabolic processes, thus regulating rice seedling growth. In this process, the differentially expressed key proteins, such as the 14-3-3 protein, GTP-binding protein, and calcium, play important roles in transduction of signals regarding soil drought, and the upregulated heat shock protein, glutathione S-transferases, and peroxidases function in enhancing the stress tolerance of the seedlings under dry-raising nursery conditions. This study established the high yielding mechanism of dry-raised cultivates methods during seedling stage at the protein expression level.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Background
Strengthening seedlings by dry-raising is a key technique for high-yielding rice cultivation (Wang and Lin 1999). Controlling seedling growth by appropriate watering, promoting root growth by fertilizing, and enhanced tillering by root strengthening, have been strictly employed for controlling soil water during the seedling stage and keeping the seedlings in an appropriate drought environment (Zhang et al. 1999). In this way, the conditions, including soil ventilation, nursery bed light transmission, and temperature, are superior to those when other nursery methods are used (Lin et al. 1997, 1998). In recent years, because of the environmental changes, transplanted rice seedlings often encounter various extreme environmental stresses, which retard rice growth after transplanting and seriously affect rice yield (Deryng et al. 2014; Redona and Mackill 1996). However, because of their of drought hardening, the dry-raised seedlings (DRS) possess well-developed root systems and enriched tissues; therefore, they display strong stress tolerance to low or high temperatures, drought, and other adverse environmental stresses (Zhao et al. 2001; Lu et al. 1995). The dry-raising nursery technique can effectively control the leggy growth of seedlings, promote dwarfing in the above-ground parts, and produce high-quality seedlings with short stems and more tillers (Lu et al. 1996; Mishra and Salokhe 2008; Yang et al. 2000). After transplanting, the DRS can take root quickly, turn green early, and produce a high yield. Thus, to enhance the potential rice yield, it is important to further understand the high-yielding mechanisms of DRS.
There have been many studies on the morphology, anatomical structure, and physiological and biochemical characteristics of DRS. These studies have shown that the high photosynthetic potential, carbon and nitrogen metabolism, dry matter accumulation, rooting, and oxidizing ability of DRS are the physiological basis of their high yield (Ding et al. 2001; Lu et al. 1996; Solaiman and Hirata 1997; Zhou et al. 2004). DRS had significantly higher contents of soluble sugar, starch, total nitrogen, protein nitrogen, and non-protein nitrogen than moist-raised seedlings (MRS), which increased leaf chlorophyll contents and photosynthetic capacity, resulting in higher accumulation of dry matter (Lu et al. 1996; Chen et al. 2007). Further studies showed that the soluble sugar and nitrate nitrogen contents of DRS were several times greater than those of the MRS, and the large amounts of accumulated carbon and nitrogen provided sufficient nutrients for the rapid development of new root and the robust growth of the root system after the DRS were transplanted (Ding et al. 2001; Lin et al. 1998). In this way, the well-developed root system and high root activity were the main reasons for the robust growth of DRS (Ding et al. 2001; Wen et al. 2000; Mishra and Salokhe 2008). DRS had stronger stress tolerance and were able to rapidly resume growth after transplantation, resulting in significant green-turning and early tillering (Yang et al. 2000), which is considered as an important physiological basis of high-yield in rice at the early stage.
The technique of raising rice seedlings in dry-raising nurseries is widespread, and it has been greatly improved via combination with rice population quality optimization control techniques. However, so far, studies of dry-raising nursery methods have been limited to the physiological level. Some physiological results have been inconsistent, which might result from different experimental methods and cultivation environments. As we know, in different cultivation environments, the spatiotemporal variation in crop gene expression is the key mechanism that regulates the processes of cell life as they adapt to the external environment. In recent years, the study of the molecular ecological relationship between the crop and environment has drawn some interest, and molecular regulation has become an important topic in contemporary crop cultivation research (Doebley et al. 2006; Burger et al. 2008). To further enrich the theoretical basis for dry-raising nursery techniques, we based the current study on a quantitative proteomics strategy, using MRS controls to investigate the molecular response of DRS to dry-raising nursery and to establish the molecular ecological physiological mechanism underlying seedling strengthening as mediated by appropriate drought stress conditions at the rice seedling stage.
Methods
Seedling growth and treatment conditions
The experiments were carried out at Fujian Agriculture and Forestry University, Fuzhou, Fujian, China (119.280 E, 26.080 N) in 2014. The rice cultivar Jinhui No. 809 (Oryza sativa L. spp. Indica) was used in this study. The entire experimental field was fertilized on May 18, 2014. For each square meter of the area, 50 g urea, 100 g calcium superphosphate, 40 g potassium chloride, and 2 kg organic fertilizer were evenly mixed into the 0–15-cm soil layers. The seeding rate was 120 g/m2. Among the fertilizers used, the urea contains 46.4% nitrogen, the calcium superphosphate contains 12% P2O5, the potassium chloride contains 60% K2O, and the organic fertilizer contains ≥ 45.0% organic matter. The seeds were sown on May 26, and the seeding rate was 120 g per square meter. Except for water control, all cultivation conditions were established as in conventional management. The average temperature during the seedling stage (from 18 May to June 23) was 26.75 °C.
Two seedling cultivation scenarios were used in the experiment: (1) Dry-raising seedlings (DRS) in the nursery bed, where the water potential in soil with a depth of 20 cm was maintained at 15 ± 2 kPa; (2) Moist raising seedlings (MRS) in the bed, where a 1–2-cm shallow water layer was maintained. Three soil water sensors were installed in each DRS plot to monitor soil water potential. Field planting patterns were used, and the two treatments were established in triplicate in a total of six plots 2.5 m2 in size in a split plot design. Ridges were built between the plots, and plastic sheets were used for waterproofing. A waterproof greenhouse was also built to cover the experimental area to prevent the impact of rainfall on the experiments.
Physiological measurements of rice seedlings
The physiological characteristics of rice seedling were measured at 29 days seedling age (before the seedlings were transplanted). The SPAD values of leaves, plant height, root length, root activity, and white roots were recorded for five seedlings per plot and averaged. The SPAD value was measured using a SPAD-502 chlorophyll meter. Root activity was determined using the triphenyl tetrazolium chloride (TTC) method (Islam et al. 2007). Five hills per plot were sampled for the dry weight. Shoot and root dry weights of seedlings were measured after oven drying at 70 °C for 1 week. The leaves and roots were collected from ten seedlings per plot and assessed for soluble sugar content, starch content, and N content. The soluble sugar and starch content were determined by Anthrone colorimetry (Zhang and Qu 2003). Total nitrogen was determined using the Micro-Kjeldahl Method (Zhang and Qu 2003). Three replicate pots for each treatment were sampled at each stage.
Quantitative analysis of plant hormone
The determination of IAA, ABA, GA3, and ZT content was performed on the leaf and root samples at 29 days seedling age. Briefly, a 2.0-g sample was ground in liquid nitrogen, homogenized, and extracted for 24 h with 20 mL 80% cold aqueous methanol in the dark at 4 °C. The extract was centrifuged at 5000 rpm and 4 °C for 15 min, and the supernatant was collected. Then, fresh, cold methanol was poured into the residue, which was extracted three times using techniques described by Chen and Yang (2005). The IAA, ABA, GA3, and ZT were quantified using high-performance liquid chromatography (HPLC)-mass spectrometry with an external standard. The sample was filtered with a 0.22 μm membrane syringe filter and then injected onto an HPLC Eurospher-100 C18 column (250 × 4 mm I.D.; 5 μm particle size, Knauer, Germany). The column was eluted isocratically with mobile phase 100% methanol and 0.2% acetic acid (50:50 (v/v) at a flow rate of 0.8 mL·min− 1. The ABA peak was measured at 254 nm and 35 °C and quantified using a known standard solution. The experiment was performed three times.
Determination of grain yield and yield components
The 29-day seedlings of two raising seedling treatments were transplanted with a spacing of 19.8 cm × 16.5 cm and one seedling per hill. The experiment was laid out in a split-plot design with three replications. Each plot was 5 × 4 m and received a total dose of fertilizer of 225 kg N ha− 1, 112.5 kg P2O5 ha− 1, and 180 kg K2O ha− 1. The phosphorus fertilizer was used for the basal fertilization, and the potassium for the top dressing. The nitrogen fertilization was applied in three sessions, i.e., 60% before transplanting, 30% at tillering, and 10% at panicle initiation. At the ripened stage, ten rice plants randomly selected from each plot were used for the measurement of grain yield and yield components. Yield components included effective panicle number per m2, spikelets per panicle, seed setting rate, and 1000-grain weight.
Protein extraction and preparation
Total protein was extracted using the cold-acetone method. The three biological replicates of the frozen leaves and roots from 29-day-old seedlings were pooled for TMT analysis. Leaf and root samples were separately ground into a fine powder in a mortar and pestle under liquid nitrogen. Approximately 1 g of the ground sample material was combined with 10 ml of resuspension buffer (10% trichloroacetic acid, 0.07% v/v 2-mercaptoethanol in acetone), and the samples were kept at − 20 °C overnight. The thawed sample was then centrifuged at 15,000 rpm for 30 min at 0–4 °C. After decanting the supernatant, the precipitate was washed with pre-cooled, 100% acetone containing 0.07%-mercaptoethanol followed by centrifugation at 15,000 rpm for 30 min at 0–4 °C. These procedures were repeated every 8 h until the supernatant was achromatic. Then, the precipitate was dried under vacuum to yield a sample pellet, which was then dissolved in a TMT-compatible buffer containing 6 M urea, 50 mM triethylammonium bicarbonate (TEAB), pH 8.5, and 2% CHAPS. Protein concentration was determined by the Bradford assay using BSA as a standard.
Digestion and TMT labeling
Here, 45 μL of 100 mM TEAB was added to 100 μg samples, and samples were adjusted to a final volume of 100 μL with ultrapure water. Protein digestion was performed according to our previous report (Zhang et al. 2017). The tryptic peptides were incubated with 2-plex TMTs label reagent (Frankfurt am Main, Germany). The TMT reagents were equilibrated at room temperature and each aliquot was resuspended in 41 μL of anhydrous acetonitrile. Samples were divided into two groups and labeled as follows: Group 1: 127 for Leaf-DRS, 126 for Leaf-MRS; Group 2: 127 for Root-DRS, 126 for Root-MRS. After labeling, individual TMT 2-plex samples were mixed and diluted into 0.1% trifluoricacetic acid, followed by loading on a MacroSpin Vydac C18 reverse phase mini-column (The Nest Group Inc., Southborough, MA, US).
LC-MS/MS separation
TMT-labeled samples were analyzed using a Q Exactive™ mass spectrometer (Thermo Fisher Scientific, US) coupled to an EASY-nLC 1000 (Thermo Fisher Scientific). Each sample (2.5 μL) was injected into the LC-MS/MS system, and the peptide separation was carried out on a C18 column (Easy-spray C18, 100 Å, 75 μm × 50 cm × 2 μm, Thermo Scientific™ Dionex™) at a flow rate of 0.25 μL/min. Peptides were separated using linear gradient ranging from 3% solvent B (mobile phase A, 0.1% formic acid in water; mobile phase B, 0.1% formic acid in ACN) to 30% solvent B for 3 h. The mass spectrometer (MS) was operated in the positive ionization mode. The MS survey scan was performed using Fourier transform equipment from a mass range of 300–1800 m/z. The resolution was set to 70,000 at 200 m/z, and the automatic gain control was set to 1,000,000 ions. The 20 most intense precursor ions were selected for collision-induced fragmentation in the linear ion trap at a higher-energy collisional dissociation energy of 35%.
Protein database search and functional classification
The raw mass data were processed with ProteomeDiscover 1.4 (Thermo Fisher Scientific) and searched against the Rice Genome Annotation Project database (RAP-DB). In the database search, full tryptic specificity was required with tolerance set at one missed cleavage. Carbamidomethylation of cysteine and TMT-2-plex modification of the N terminus and K were set as fixed modifications. Gln→pyro-Glu of the N terminus, deamination of the N terminus, and oxidation of methionine were set as variable modifications. The initial precursor mass tolerance was set to 10 ppm, and the fragment ion level was set to 0.02 Da. The protein was designated as a hit only when two or more unique peptides with high confidence scores (FDR < 1%) were identified and their corresponding MS/MS spectra were manually inspected.
Proteins with significant changes in DRS were selected on the basis of the previous report (Cox and Mann 2008). In brief, the mean ratio and SD from the log2 ratios of 2675 leaf proteins and 2918 root proteins identified with RAP-DB were calculated. Next, 95% confidence intervals were used to select those proteins whose distribution was removed from the main distribution. For the up-regulated leaf proteins, the confidence interval was the mean ratio of 2675 leaf proteins + Z score (95%) × SD, corresponding to a protein ratio of 1.36. Similarly, for the down-regulated leaf proteins, the confidence interval was calculated (− mean ratio − Z score × SD), corresponding to a protein ratio of 0.62. Protein ratios outside this range were defined as being significantly different at P = 0.05. The cutoff value for the down-regulated leaf proteins was 0.62-fold, and for the up-regulated proteins it was 1.36-fold. Based on this rule, for the 2918 root proteins, the cutoff value for the down-regulated proteins was 0.78-fold, and for the up-regulated proteins it was 1.23-fold. Functional annotation was obtained as gene ontology annotations from the RAP-DB and matched against the identified proteins. Protein functional classification and assignment were performed according to the bincodes of MapMan (Thimm et al. 2004).
Quantitative RT-PCR analysis
Total RNA was extracted from leaves and roots at 29 days seedling age, similar to those used for proteome analysis in three independent extractions using Trizol reagent (Invitrogen, Life Technologies) and cDNA was reverse transcribed from 1 μg of total RNA using M-MLV reverse transcriptase (TaKaRa). The primer pairs were designed using Beacon Designer 7.0 software (Additional file 1, Table S1). Gene expression was assayed using a Mastercycler EP Realplex RT-PCR system (Eppendorf) and Bestar™ Real-time PCR Master Mix SYBR Green (DBI Bioscience, Germany). The reaction mixture contained 7.5 μL Bestar SybrGreen qPCR mastermix (DBI Bioscience), 0.5 μL cDNA, and 5 μm of each gene-specific primer in a final volume of 15 μL. The PCR program was 2 min at 95 °C, (10 s at 95 °C, 20 s at 54 °C, and 30 s at 72 °C) 40×, and 1 min at 72 °C followed by recording the melting curve. The specificity of the PCR amplification was checked with a melting curve analysis (from 55 to 98 °C, 10 s/cycle, 0.5 °C/cycle) after the final PCR cycle. To determine relative fold differences for each sample in each experiment, we studied Ct value β-actin as normalized and calculated relative to a calibrator using the formula 2− ΔΔCt. The calibrator is the sample that exhibited the lowest level of transcripts in this experiment (El-Sharkawy et al. 2010).
Results
Physiological and morphological characteristics of the dry-raised seedlings
Before the seedlings were transplanted (29-day-old seedlings), the physiological and morphological characteristics were examined. As shown in Fig. 1 and Table 1, the DRS were shorter, but had larger values for root length, root activity, number of white roots, and thickness of root hairs than MRS, while the MRS had mostly yellow roots and few root hairs. The dry weights of both the aboveground and underground parts were higher in DRS than in MRS at the 29-day-old seedling stage. The leaf SPAD values were also higher in DRS than in MRS. These results indicated that the synthesis and accumulation of photosynthetic substances are higher under dry-raising nursery conditions than its counterparts. The root-to-shoot ratios of DRS were 18.8% higher than MRS, indicating that the dry-raising nursery conditions had a greater impact on root than on leaf. Table 2 showed that the soluble sugar, starch, and N content of DRS leaves were 21.8, 18.0, and 32.29% higher than those of MRS leaves, respectively. However, the C/N ratio of DRS leaves was slightly lower than that of MRS leaves. The changes in all of the above C and N metabolism-related indexes in DRS root showed the same pattern as those in DRS leaf, that is, the soluble sugar, starch, and N content were higher, but the C/N ratio was lower (Table 2). In addition, Table S2 (Additional file 2) shows that the effective panicles per m2, spikelets per panicle, filled grains percent, and grain yield were 7.6, 2.5, 3.9, and 15.7% higher, respectively, in DRS than in MRS.

Morphological and physiological changes in rice seedlings under dry-raising nursery management. a Morphological observation. b Plant height. c Root length. d Root volume. e Number of white roots. DRS dry-raising seedling, MRS moist-raising seedling. Vertical bars represent ± standard error of the mean (n = 3). Statistically significant differences to the control were calculated using independent Student’s t tests: *P < 0.001
Changes in hormone content of the dry-raised seedlings
Table 3 showed that the ZT, GA3, ABA, and IAA content in leaves at the 29-day-old seedling stage significantly increased wth DRS treatments. Among of them, the GA3 and ABA were 7.3-fold and 3.4-fold higher than in MRS. For roots treated with the DRS, the GA3, ABA, and IAA content increased by 5.4-fold, 1.7-fold, and 1.5-fold, respectively, while the ZT level was reduced by 1.2-fold relative to MRS root.
Quantitative proteomics analysis of dry-raised seedlings
To further understand the molecular mechanism underlying the stronger seedlings of DRS, we selected 29-day-old rice seedlings and analyzed the changes in their protein expression using the TMT isotope labeling method. We detected 2918 proteins in the root system (Additional file 3) and 2675 in the leaves (Additional file 4). At a confidence level of 0.05, a total of 281 and 268 differentially expressed proteins were identified in the root system and leaves, respectively. By analyzing the expression levels of the proteins, we found 134 leaf proteins and 154 root proteins to be up-regulated in DRS (Fig. 2a), while 134 leaf proteins and 127 root proteins were down-regulated in DRS (Fig. 2b). Only 22 proteins did not show different expression patterns in both leaves and roots, among which 10 were up-regulated and 12 were down-regulated. MAPMAN software was used to perform functional analysis on the identified differentially expressed proteins, which were then divided into 23 categories, while 87 leaf and 59 root proteins were defined as being of unknown function (Additional file 5; Additional file 6). Proteins related to tricarboxylic acid cycle (TCA), N-metabolism, secondary metabolism, and hormone metabolism were up-regulated in both leaves and roots, while the proteins related to cell transport were down-regulated. Leaf proteins related to photosynthesis and root proteins related to stress showed significant upregulation, while the proteins related to RNA and DNA showed significant downregulation in both leaves and roots (Fig. 2c).

Qualitative comparison of differentially expressed proteins. a Venn diagram of the up-regulated protein identified in rice leaf and root under dry-raising nursery management. b Venn diagram of the down-regulated protein identified in rice leaves and roots under dry-raising nursery management. The diagram shows the overlap between the results from the leaf and root protein. c Functional characterization of leaf and root proteins under dry-raising nursery management
Validation of protein expression changes as assessed by quantitative RT-PCR
We further confirmed the changes in protein levels observed in DRS by evaluating the changes in transcript levels of hormone-related genes. Abscisic aldehyde oxidase (AAO) catalyzes the final step in the ABA biosynthesis pathway, which is one of the more important steps in the pathway. The RT-PCR results showed that the AAO1 in leaves and AAO in roots were increased by 2.2-fold and 3.7-fold, which corresponded well to the changes in their protein levels in DRS (Fig. 3). As expected, a significance increase was observed in the expression level of most of the hormonal-related genes, such as carotenoid cleavage dioxygenase 1 (CCD1), IAA-amino acid hydrolase ILR1 (ILL1), gibberellin 20 oxidase 2 (GA20OX2), lipoxygenase 2.3 (LOX2.3), allene oxide cyclase 4 (AOC 4) in leaves and ILL1, 1-aminocyclopropane-1-carboxylate oxidase (ACCO), and 12-oxophytodienoate reductase 2 (OPR2) in roots, which, at the mRNA level, confirmed the proteomic results (Fig. 3).

Hormone-related genes expression profile in DRS. Total RNA was extracted from leaves and roots of 29-day-old seedlings and subjected to qRT-PCR. The qRT-PCR amplification experiments were performed using two biological replicates with three technical replicates each, and the data were averaged. Relative expression levels were calculated and normalized with respect to β-actin mRNA, and error bars represent the standard error of the mean
Discussion
Dry-raising can increase the porosity and oxygen content in a nursery seedbed, benefitting seedling growth and development. Physiological analysis showed that drought stress during the seedling stage could significantly change the morphology and physiological characteristics of the seedlings. The DRS had shorter bodies, better-developed root systems, higher leaf SPAD value, significantly greater root activity, and significantly more white roots. The soluble sugar, starch, and N levels in DRS leaves were higher than in MRS, but the C/N ratio was slightly lower than that of MRS, indicating the vigorous carbon and nitrogen metabolism in DRS. This suggested that the DRS would adapt better to dry soil ecological environments, which explains why the DRS turn green faster and have better tillering after transplantation than MRS (Lu et al. 1997; Mishra and Salokhe 2008; Yang et al. 2000). The DRS in this study produced higher yields (15.7%) than the MRS, indicating the higher quality of DRS. After clarifying the physiological responses of seedlings to dry-raising nursery cultivation, we further utilized quantitative proteomics techniques and identified 268 leaf and 281 root proteins that were differentially expressed in response to drought stress. Six categories of functional proteins related to hormones, N-metabolism, photosynthesis, CHO metabolism, stress responses, and signaling were analyzed for their effects on the physiological characteristics of DRS.
Endogenous hormones play an important role in plant responses to environmental stress. Seven proteins in leaves and five proteins in roots were identified as hormone related. Results showed that three key proteins of ABA synthesis, the aldehyde oxidase 1 (Seo et al. 2000) and carotenoid cleavage dioxygenase 1 (Schwartz et al. 2003) in the leaves and abscisic-aldehyde oxidase in the root system, showed cumulative expression under dry cultivation conditions. The HPLC experiment also showed that the ABA content in DRS was significantly increased in both leaves and roots, which verified the proteomic results. The elevated ABA contents in the leaves can reduce the leaf transpiration rate and maintain photosynthetic activity during photosynthesis in DRS (Ramachandra et al. 2004). ABA accumulation in the root is also beneficial for increasing the water permeability of roots and the ion penetration of the xylem, thus enhancing the ability of the root system to take up and absorb adequate water and nutrients under drought conditions (Zhang et al. 2006; Sharp and LeNoble 2002). At the same time, results indicated high levels of expression of 1-aminocyclopropane-1-carboxylate oxidase (Dong et al. 1992), a rate-limiting enzyme involved in ethylene synthesis, and an IAA-amino acid hydrolase ILR1 precursor that releases iodoacetamide (IAA) (LeClere et al. 2002). Studies in Arabidopsis showed that ethylene in the root system can promote the formation of root hairs and increase the root width by increasing the synthesis of IAA (Van de Poel et al. 2015; Pitts et al. 1998). The present study also showed that the IAA content in DRS roots was increased 1.5-fold over that of the MRS roots. Thus, the higher synthesis of ethylene and the release rate of IAA in DRS root systems favor the formation of branching roots and the extension of roots in depth. Moreover, three key jasmonate (JA) synthesis proteins, lipoxygenase 2.3 (Chauvin et al. 2013), allene oxide cyclase 4 in the leaves (Stenzel et al. 2003), and 12-oxophytodienoate reductase 2 (Schaller et al. 2000) in the roots, also showed up-regulated expression in DRS. Many studies have shown that JA in plants can promote the expression of stress-related genes, thereby enhancing tolerance to drought stress (Creelman and Mullet 1995; Koda 1992). In addition, although none of the proteins related to GA3 synthesis have been identified, we also found the content of GA3 in DRS leaves and roots to be higher than that in MRS. Previous studies have found that improvements in plant architecture, such as semidwarfism, increased root systems, and higher tiller numbers could be induced by the high GA3 content (Lo et al. 2008). Based on these results, we presumed that the higher tiller numbers, shorter bodies, and better-developed root systems in DRS were closely related to the changes in GA3 content.
In this study, the N content of leaves and roots were 32.29 and 37.86% higher in DRS than in MRS, respectively, indicating stronger nitrogen metabolism. Enhancement of nitrogen metabolism favors rapid growth of rice seedlings, which has been confirmed by the physiological performance of DRS described above. The present study found that four key nitrogen metabolism proteins, nitrate reductase 1 (NR1), ferredoxin-nitrite reductase (NiR), glutamine synthetase (GS), and glutamine synthetase root isozyme 2 (GSRI2), were up-regulated under the dry cultivation environment (Fig. 4), which is consistent with previous studies (Chen et al. 2007; Wang et al. 2003), showing that the nitrase activity and the total nitrogen content of the plant were significantly higher in DRS than in MRS. These results suggested that DRS might absorb more nitrate from the dry-raising nursery bed soil, then degrade them by NR to nitrite, and hence to NH4 + by NiR. Subsequently, GS converts glutamic acid and NH4 + into glutamine, which is transported to the aboveground parts of the rice seedling, as shown in Table 1, with high nitrogen contents in DRS. Moreover, NiR and glutamate synthase (GOGAT) were upregulated in DRS leaves, which enables the nitrogen to be transported from roots to further transform into amino acids and participate in the protein synthesis of rice plants. The 12 amino acid metabolism proteins found to be differentially expressed in DRS leaves tended to be upregulated in this study. Vigorous amino acid metabolism in the leaves depends on the availability of a sufficient nitrogen supply from roots; this suggested that dry-raising nursery is beneficial for nitrogen assimilation and transportation and amino acid metabolism to support strong and healthy growth of DRS.

Changes in abundance of N-metabolism enzymes in root and leaf. The number refers to the fold change in dry-raised seedlings relative to moist-raised seedlings. NR1 nitrate reductase 1, NiR nitrite reductase, GS glutamine synthetase, GSRI2 glutamine synthetase root isozyme 2, GOGAT glutamate synthase
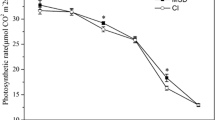
This study also demonstrated that the chlorophyll SPAD value of the DRS was greater than that of MRS, because chlorophyll content can indicate the efficiency of light absorption and utilization of crops (Sinclair and Horie 1989; Netto et al. 2005). The highest dry matter-weights were also detected in DRS rather than in its counterpart, which was consistent with the work reported by Lu et al. ( 1996), showing that the photosynthetic rate of DRS to be higher than that of MRS. Further analyses identified 31 photosynthesis-related proteins in the leaves of rice seedlings by quantitative proteomics methods are involved in photoreaction, Calvin cycle, and photorespiration (Additional file 7, Fig. S1). Among the 17 identified photoreaction-related proteins, 15 were up-regulated in DRS, including seven photosystem II proteins, one photosystem I protein, two ferredoxin-NADP reductase subunits, and three ATP synthase subunits, which covered the whole process of photoreaction. Furthermore, seven key proteins related to Calvin cycle were also identified, and all of them were upregulated in the leaves of DRS. Thus, these results further explained how the DRS could efficiently utilize the energy generated from the photoreaction process to fix more CO2. Also, the expression levels of the eight photorespiration proteins detected in this study demonstrated that the leaf respiration intensity was stronger in DRS than in MRS. Recent research has shown that photorespiration is a metabolic pathway formed during long-term evolution to adapt to environmental changes and enhance stress tolerance (Walker et al. 2016; Voss et al. 2013). The greater photorespiration rate of leaves in DRS can enhance resistance to excessive light energy in leaves and ensure normal photosynthetic reactions, thereby increasing carbohydrate assimilation and supplying adequate materials for the growth of seedlings under the dry raising regime. These results indicate an important molecular mechanism by which there comes to be more dry matter in DRS than in MRS, which is considered now as a physiological basis of high-yielding ability in DRS.
This study also determined the upregulation of several leaf proteins in DRS, including that of fructose-1,6-bisphosphatase, which is involved in sucrose synthesis, and glucose-1-phosphate adenylyltransferase, whose small subunit is related to starch synthesis. The present study also showed that the contents of soluble sugar and starch in DRS were significantly higher than those in MRS, which enhanced the stress resistance of DRS and favored rapid recovery and turning green after seedling transplantation (Lu et al. 1996; Wang et al. 2003). The ADP and ATP carrier protein (which are involved in starch transport), beta-amylase and alpha-glucan phosphorylase H isozyme (which are involved in starch degradation), and two alpha-amylase isozymes in the roots were upregulated in DRS, suggesting that DRS, which have high levels of soluble sugar and starch, have a stronger ability to undergo photosynthesis and substance translocation. This study also revealed that the expressions of most proteins involved in carbon metabolism pathways in leaves and roots were greater under dry cultivation conditions. These proteins included phosphoglucomutase, which is involved in glycolysis; 3-isopropylmalate dehydrogenase and carbonic anhydrase, which are involved in the TCA cycle; aldehyde dehydrogenase, which is involved in fermentation in the leaves; fructose-bisphosphate aldolase cytoplasmic isozyme, glyceraldehyde-3-phosphate dehydrogenase, glyceraldehyde-3-phosphate dehydrogenase A, and phosphoenolpyruvate carboxylase 2, which are involved in glycolysis; and citrate synthase 4, NADP-dependent malic enzyme, and carbonic anhydrase, which are involved in TCA cycle in the roots. The changes in carbon metabolism-related protein expression indicated that the carbon metabolism of DRS was greater than that of MRS, which further confirmed that DRS produced more energy for its growth. Moreover, the changes in protein expression resulted in the accumulation of large amounts of carbon metabolism intermediates and laid a material foundation for rice seedlings to recover rapidly, turn green, and grow quickly after transplantation.
Throughout our studies, we suggested that the higher stress tolerance of the DRS is one of the physiological mechanisms for the stronger resistant ability and rapidly greening properties after transplantation (Zhou et al. 2004). This study demonstrated the expression changes of 16 heat shock protein (HSP)-related proteins in roots and 8 HSP-related proteins in leaves, of which 23 showed high expression levels in DRS. Previous research has shown that seedlings subjected to drought stress hardening displayed higher resistance not only to drought but also to hot and cold temperatures (Zhao et al. 2001; Lu et al. 1995). Our results showed that drought stress could promote the expression of HSP proteins in seedlings and increase tolerance to different kinds of environmental stresses. This study also uncovered changes in expression of several glutathione S transferases (GST, three in leaves and six in roots) and peroxidase subunits (four in leaves and 11 in roots) under drought stress. GST can cause substantial glutathione production in plants, thus promoting plant growth and enhancing plant stress tolerance (Marrs 1996). Overexpression of GST in tobacco significantly improved its tolerance to draught and salt stress (Ji et al. 2010). Peroxidases can promote the lignification of plant tissue and enhance the ability of plants to resist dehydration. This study demonstrated that, under drought stress conditions, the HSP protein, GST, and peroxidases in the seedlings interact with each other and form a molecular defense system, enhancing the stress tolerance of the DRS, which allows the DRS to turn green faster than MRS after transplantation.
Further analysis showed that some signal molecules participated in the physiochemical process in DRS, suggesting that in dry-raising nurseries with little soil water and high oxygen content, the root system of the DRS sensed the signals in soil water stress and changes in oxygen concentration, and then controlled the growth of seedlings. Through proteomics, this study demonstrated that five calcium-signaling-related proteins in the root system first responded to the soil environmental signals and displayed changes in expression, the signals were transmitted to two receptor kinases and led to further changes in the expression of the 14-3-3 protein (Fig. 5). This eventually exerted regulatory effects on the functional proteins in the root system. The signals from the roots were further sensed by the four GTP-binding proteins in the leaves and transformed to calcium signaling systems, which led to changes in 14-3-3 protein expression exerting a regulatory effect on the functional proteins in leaves. The 14-3-3 protein family is reported to be involved in drought stress response in maize (Campo et al. 2012), rice (Chen et al. 2006), and Arabidopsis (He et al. 2015). Many studies have shown that by interacting with other proteins, the 14-3-3 protein regulates physiological and biochemical processes including ABA (del Viso et al. 2007) and ethylene (Yao et al. 2007) signaling pathways, carbon metabolism (Comparot et al. 2003), and starch synthesis (Sehnke et al. 2001). Our results demonstrated that these physiological and biochemical processes are important for raising strong seedlings in dry-raising nurseries. Moreover, previous research has shown that the 14-3-3 protein inhibits the activity of NR, a key enzyme of nitrogen metabolism (Lima et al. 2006). In this study, the 14-3-3 protein in root showed a down-regulation in DRS, while the expression of NR protein showed an up-regulation. We suggest that the 14-3-3 protein senses the soil environmental signal, then alters the activity of NR, and further regulates the absorption of soil nitrogen by the root system. In this way, 14-3-3 protein plays a major role in sensing the soil environmental signal and regulating the seedling growth, as shown in Fig. 5.

Changes in abundance of signaling-related proteins in roots and leaves. The number refers to the fold change in dry-raised seedlings relative to moist-raised seedlings
Conclusions
In summary, the present study showed that the dry-raising nursery technique can effectively improve rice seedling quality, as indicated by root activity, white root number, nitrogen content, and leaf SPAD value, leading to increased grain yield. We then used proteomics to identify a large number of proteins in the leaves and roots of the seedlings. These proteins showed changes in expression in response to dry cultivation. Moreover, we constructed a protein function network regulating rice seedling growth in a dry-raising nursery (Fig. 6) and produced further support for the dry-raising nursery high-yield cultivation techniques of rice. Proteins found here to be key to seedling growth, such as 14-3-3, may be used in the future to study molecular manipulation of rice traits and promotion of rice yield.

Model showing protein categories that respond differently to dry-raising nursery management. Up-and downward arrows represent increases and decreases in protein abundance
Abbreviations
- TMT:
-
Tandem mass tags
- HPLC:
-
High-performance liquid chromatography
- DRS:
-
Dry-raising seedlings
- MRS:
-
Moist-raising seedlings
- TCA:
-
Tricarboxylic acid cycle
- NR1:
-
Nitrate reductase 1
- NiR:
-
Nitrite reductase
- GS:
-
Glutamine synthetase
- GSRI2:
-
Glutamine synthetase root isozyme 2
- ABA:
-
Abscisic acid
- GA3:
-
Gibberellins
- IAA:
-
Indole-3-acetic acid
- JA:
-
Jasmonate
- HSP:
-
Heat shock proteins
- GST:
-
Glutathione S transferases
- TEAB:
-
Triethylammonium bicarbonate
- RT:
-
Room temperature
- MS:
-
Mass spectrometer
- RAP-DB:
-
Rice Genome Annotation Project database
- GOGAT:
-
Glutamate synthase
- AAO:
-
Abscisic aldehyde oxidase
- CCD1:
-
Carotenoid cleavage dioxygenase 1
- ILL1:
-
IAA-Amino acid hydrolase ILR1
- GA20OX2:
-
Gibberellin 20 oxidase 2
- LOX2.3:
-
Lipoxygenase 2.3
- AOC 4:
-
Allene oxide cyclase 4
- ACCO:
-
1-Aminocyclopropane-1-carboxylate oxidase
- OPR2:
-
12-Oxophytodienoate reductase 2
References
Burger JC, Chapman MA, Burke JM (2008) Molecular insights into the evolution of crop plants. Am J Bot 95(2):113–122
Campo S, Peris-Peris C, Montesinos L, Peñas G, Messeguer J, San Segundo B (2012) Expression of the maize ZmGF14-6 gene in rice confers tolerance to drought stress while enhancing susceptibility to pathogen infection. J Exp Bot 63(2):983–999
Chauvin A, Caldelari D, Wolfender JL, Farmer EE (2013) Four 13-lipoxygenases contribute to rapid jasmonate synthesis in wounded Arabidopsis thaliana leaves: a role for lipoxygenase 6 in responses to long-distance wound signals. New Phytol 197(2):566–575
Chen YP, Yang WY (2005) Determination of GA3, IAA, ABA and ZT in dormant buds of Allium ovalifolium by HPLC. J Sichuan Agric Univ 23:498–500
Chen F, Li Q, Sun L, He Z (2006) The rice 14-3-3 gene family and its involvement in responses to biotic and abiotic stress. DNA Res 13(16766513):53–63
Chen H, Liang Y, Lin W, Zheng L, Liang K (2007) Quality and physiobiochemical characteristics of the first rice crop seedlings under different raising seedling patterns for early rice and its ratoonal crop (I)—studies on super high-yield ecophysiology and its regulation technology in hybridize rice. Chin Agric Sci Bull 23(2):247–250
Comparot S, Lingiah G, Martin T (2003) Function and specificity of 14-3-3 proteins in the regulation of carbohydrate and nitrogen metabolism. J Exp Bot 54(382):595–604
Cox J, Mann M (2008) MaxQuant enables high peptide identification rates, individualized ppb-range mass accuracies and proteome-wide protein quantification. Nat Biotechnol 26(12):1367–1372
Creelman RA, Mullet JE (1995) Jasmonic acid distribution and action in plants: regulation during development and response to biotic and abiotic stress. Proc Natl Acad Sci USA 92(10):4114–4119
Del Viso F, Casaretto JA, Quatrano RS (2007) 14-3-3 Proteins are components of the transcription complex of the ATEM1 promoter in Arabidopsis. Planta 227(1):167–175
Deryng D, Conway D, Ramankutty N, Price J, Warren R (2014) Global crop yield response to extreme heat stress under multiple climate change futures. Environ Res Lett 9(3):034011
Ding Y, Wang Q, Wang S, Huang P (2001) Comparison studies of roots physiology activity between rice dry seedbed seedlings and wet seedbed seedlings. J Nangjing Agric Univ 24(3):1–5
Doebley JF, Gaut BS, Smith BD (2006) The molecular genetics of crop domestication. Cell 127(7):1309–1321
Dong JG, Fernandez-Maculet JC, Yang SF (1992) Purification and characterization of 1-aminocyclopropane-1-carboxylate oxidase from apple fruit. Proc Natl Acad Sci USA 89(20):9789–9793
El-Sharkawy I, Mila I, Bouzayen M, Jayasankar S (2010) Regulation of two germinlike protein genes during plum fruit development. J Exp Bot 61(6):1761–1770
He Y, Wu J, Lv B, Li J, Gao Z, Xu W, Baluska F, Shi W, Shaw PC, Zhang J (2015) Involvement of 14-3-3 protein GRF9 in root growth and response under polyethylene glycol-induced water stress. J Exp Bot 66(8):2271–2281
Islam E, Yang X, Li T, Liu D, Jin X, Meng F (2007) Effect of Pb toxicity on root morphology, physiology and ultrastructure in the two ecotypes of Elsholtzia argyi. J Hazard Mater 147(3):806–816
Ji W, Zhu Y, Li Y, Yang L, Zhao X, Cai H, Bai X (2010) Over-expression of a glutathione S-transferase gene, GsGST, from wild soybean (Glycine soja) enhances drought and salt tolerance in transgenic tobacco. Biotechnol Lett 32(8):1173–1179
Koda Y (1992) The role of jasmonic acid and related compounds in the regulation of plant development. Int Rev Cytol 135:155–199
LeClere S, Tellez R, Rampey RA, Matsuda SP, Bartel B (2002) Characterization of a family of IAA-amino acid conjugate hydrolases from Arabidopsis. J Biol Chem 277(23):20446–20452
Lima L, Seabra A, Melo P, Cullimore J, Carvalho H (2006) Post-translational regulation of cytosolic glutamine synthetase of Medicago truncatula. J Exp Bot 57(11):2751–2761
Lin W, Wang S, Liang Y, Guo Y, He S, Hong L, Zheng L, Weng D, Pang Z (1997) Physioecological study on highyielding cultivation of rice by dryraising seedling and thinspacing transplanting techniques I. Quality and ecophysiological characteristics of rice seedlings grown on dryfertile nursery. Chin J Appl Ecol 8(6):566–570
Lin W, Wang S, Liang Y, Guo Y, He S, Zheng F, Weng D, Hong L, Pan Z (1998) Physio-ecological study on high yielding cultivation of rice by dry raising seedling and thin spacing transplanting techniques II. High yielding formation and its physiobiochemical properties of early rice. Chin J Appl Ecol 9(4):395–399
Lo S-F, Yang SY, Chen KT, Hsing YI, Zeevaart JA, Chen LJ, Yu SM (2008) A novel class of gibberellin 2-oxidases control semidwarfism, tillering, and root development in rice. Plant Cell 20(10):2603–2618
Lu X, Peng L, Tang X, Liu X, Luo Z (1995) Studies on physiological reason for cold resistance of early rice seedlings raised in dry nursery. J Hunan Agric Univ 22(3):225–230
Lu X, Tang X, Peng L, Liu X, Zheng X, Luo Z (1996) The characteristics of morphology, physiology and biochemistry of late rice plants cultivated by raising seedlings with dry nursery management. J Hunan Agric Univ 23(4):307–315
Lu X, Peng L, Tang X, Liu X, Luo Z (1997) Studies on the morphology, tissue structure and physiological characteristics of early rice (Oryza sativa L.) seedlings raised in dry nursery. Acta Agron Sin 23(3):360–369
Marrs KA (1996) The functions and regulation of glutathione S-transferases in plants. Annu Rev Plant Biol 47:127–158
Mishra A, Salokhe V (2008) Seedling characteristics and the early growth of transplanted rice under different water regimes. Exp Agric 44(03):365–383
Netto AT, Campostrini E, de Oliveira JG, Bressan-Smith RE (2005) Photosynthetic pigments, nitrogen, chlorophyll a fluorescence and SPAD-502 readings in coffee leaves. Sci Hortic 104(2):199–209
Pitts RJ, Cernac A, Estelle M (1998) Auxin and ethylene promote root hair elongation in Arabidopsis. Plant J 16(5):553–560
Ramachandra RA, Chaitanya KV, Vivekanandan M (2004) Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J Plant Physiol 161(11):1189–1202
Redona E, Mackill D (1996) Genetic variation for seedling vigor traits in rice. Crop Sci 36(2):285–290
Schaller F, Biesgen C, Mussig C, Altmann T, Weiler EW (2000) 12-Oxophytodienoate reductase 3 (OPR3) is the isoenzyme involved in jasmonate biosynthesis. Planta 210(6):979–984
Schwartz SH, Tan BC, McCarty DR, Welch W, Zeevaart JA (2003) Substrate specificity and kinetics for VP14, a carotenoid cleavage dioxygenase in the ABA biosynthetic pathway. BBA-Biomembr 1619(1):9–14
Sehnke PC, Chung HJ, Wu K, Ferl RJ (2001) Regulation of starch accumulation by granule-associated plant 14-3-3 proteins. Proc Natl Acad Sci USA 98(2):765–770
Seo M, Peeters AJ, Koiwai H, Oritani T, Marion-Poll A, Zeevaart JA, Koornneef M, Kamiya Y, Koshiba T (2000) The Arabidopsis aldehyde oxidase 3 (AAO3) gene product catalyzes the final step in abscisic acid biosynthesis in leaves. Proc Natl Acad Sci USA 97(23):12908–12913
Sharp RE, LeNoble ME (2002) ABA, ethylene and the control of shoot and root growth under water stress. J Exp Bot 53(366):33–37
Sinclair T, Horie T (1989) Leaf nitrogen, photosynthesis, and crop radiation use efficiency: a review. Crop sci 29(1):90–98
Solaiman M, Hirata H (1997) Effect of arbuscular mycorrhizal fungi inoculation of rice seedlings at the nursery stage upon performance in the paddy field and greenhouse. Plant Soil 191(1):1–12
Stenzel I, Hause B, Miersch O, Kurz T, Maucher H, Weichert H, Ziegler J, Feussner I, Wasternack C (2003) Jasmonate biosynthesis and the allene oxide cyclase family of Arabidopsis thaliana. Plant Mol Biol 51(6):895–911
Thimm O, Bläsing O, Gibon Y, Nagel A, Meyer S, Krüger P, Selbig J, Müller LA, Rhee SY, Stitt M (2004) mapman: a user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J 37(6):914–939
Van de Poel B, Smet D, Van Der Straeten D (2015) Ethylene and hormonal cross talk in vegetative growth and development. Plant Physiol 169(1):61–72
Voss I, Sunil B, Scheibe R, Raghavendra A (2013) Emerging concept for the role of photorespiration as an important part of abiotic stress response. Plant Biol 15(4):713–722
Walker BJ, VanLoocke A, Bernacchi CJ, Ort DR (2016) The costs of photorespiration to food production now and in the future. Annu Rev Plant Biol 67:107–129
Wang S, Lin W (1999) The mechanism of high-yielding in rice under dry-raising and thin-planting and its regulating technique I. Advances in the mechanism of high-yielding in rice under dry-raising and thin-planting and its prospects. J Fujian Agric Univ 28(1):12–17
Wang Q, Ding Y, Wang S, Huang P, Miao B (2003) Physiological effects of sustainable water saturation in seedbed on rice dry nursery seedlings. Acta Agron Sin 30(3):210–214
Wen H, Zhao J, Zhao W, Mao G (2000) Rooting advantage of rice seedlings nursed by dry-nursing and its impact on characteristics of growth and development of aerial part. J Zhejiang Agric Sci 1:1–5
Yang D, Duang Z, Hang J, Liu C, Wu S (2000) Study on the growth and development characters of dry nursery seedlings and their regularities of yield formation in hybrid early rice III. Characteristics of tillering and earing of dry nursery seedlings. Hubei Agric Sci 6:13–14
Yao Y, Du Y, Jiang L, Liu JY (2007) I Interaction between ACC synthase 1 and 14–3-3 proteins in rice: a new insight. Biochemistry 72(9):1003–1007
Zhang Z, Qu W (2003) Guidance of plant physiology experiments, 3rd edn. Higher Education Press, Beijing. pp 127–132
Zhang Y, Wu H, Wang Z, Xiong F, Xie Y, Li A (1999) Effect of rice seedling raising conditions on rice seedling growth. Chinese J Rice Sci 13(2):86–90
Zhang J, Jia W, Yang J, Ismail AM (2006) Role of ABA in integrating plant responses to drought and salt stresses. Field Crop Res 97(1):111–119
Zhang Z, Zhang Y, Liu X, Li Z, Lin W (2017) The use of comparative quantitative proteomics analysis in rice grain-filling in determining response to moderate soil drying stress. Plant Growth Regul 82(2):219–232
Zhao Y, Ding Y, Chen L, Huang P (2001) Physiological characteristics of drought resistance of rice dry nursery seedlings. Sci Agric Sin 34(3):289–291
Zhou Q, Chen G, Chen L, Wang J, Zhang C, Lu C (2004) Study on antioxidation system in high yield hybrid rice Langyoupeijiu seedlings under dry raising conditions. Bull Bot Res 25(1):80–88
Acknowledgements
This work was sponsored by the National Natural Science Foundation of China (No. 31401306), the Fujian-Taiwan Joint Innovative Centre for Germplasm Resources and cultivation of crop (Fujian 2011 Program, No. 2015-75), the National Key Research and Development Program of China (2016YFD0300508) and the Natural Foundation of Fujian Higher Education Institutions for Young Scientists (Key Project) (JZ160435).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
10725_2017_347_MOESM3_ESM.xlsx
The detailed information of the 2918 root proteins from the Rice Genome Annotation Project database (RAP-DB). (XLSX 302 KB)
10725_2017_347_MOESM4_ESM.xlsx
The detailed information of the 2674 leaf proteins from the Rice Genome Annotation Project database (RAP-DB). (XLSX 308 KB)
10725_2017_347_MOESM7_ESM.docx
Figure S1: MapMan overview of leaf photosynthesis proteins with significant differences in abundance in dry-raised seedlings. (DOCX 1502 KB)
Rights and permissions
About this article

Cite this article
Zhang, Z., Huang, F., Shao, C. et al. Differential proteomic analysis of rice seedlings reveals the advantage of dry-raising nursery practices. Plant Growth Regul 84, 359–371 (2018). https://doi.org/10.1007/s10725-017-0347-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-017-0347-3