
Abstract
An ancient landrace of wheat Triticum sphaerococcum Perc. has been characterized for agronomic and physiological traits under timely sown-irrigated (TS-IR), timely sown restricted irrigation (TS-RI), and irrigated late sown conditions (IR-LS) in the winter season of 2021–22. A total of 116 accessions were collected from major gene banks across the world and examined, along with 20 bread wheat cultivars. The assessed genotypes revealed a considerable degree of variance for the trait under study. Average grain yields for the T. sphaerococcum accessions were 33.29 q/ha in TS-IR, 28.95 q/ha in TS-RI, and 26.15 q/ha in LS-IR. The extent of grain yield decline is lesser than that of bread wheat. This indicates that these species can withstand conditions of limited irrigation and extreme heat. T. sphaerococcum accessions had lower yield than bread wheat, although accessions like TS 49, TS5, TS6, TS17, TS27, and TS 61 have higher yield (> 35 q/ha) under stress conditions. Under the TS-RI trial, 25 accessions of T. sphaerococcum achieved significantly greater yields when compared to one of the well-known drought-tolerant cultivars, C 306 (29.2 q/ha). Grain yield under TS-RI and LS-IR trial exhibits substantial negative association with canopy temperature at physiological maturity (− 0.63*, and − 0.74***), and strong positive correlation with TGW (0.83*** and 0.82***), GRPS (0.67***, and 0.92***). This demonstrates that the greater grain yield can be attributed to the T. sphaerococcum accessions’ ability to keep their canopies cooler in the face of heat stress. Thus, the introduction of ancient wheat in wheat cross breeding programme will increase genetic diversity and diversify current cultivars.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The cornerstone of global food security is wheat (Triticum aestivum L.), which is the most widely cultivated crop in the world. For millions of people who rely on wheat-based diets, wheat offers over 40% of their daily requirements for critical micronutrients like zinc, iron, manganese, magnesium, and vitamins B and E. It also contributes 20% of human nutrition’s total protein and calories (Velu et al. 2017). A more effective strategy to increase agricultural output and resilience to various stresses can be achieved by determining the genetic variability of the crop. Because of this, the genetic development of crops, including wheat, depends on the availability of a wide range of superior genotypes. However, it has commonly been argued that the shrinking of agricultural genetic variability was caused by extensive cultivation of mega cultivars that have led to susceptibility to biotic and abiotic stresses. As a result, it is crucial for plant breeding programs to diversify the genetic background of their crop lines in order to maintain enough genetic diversity to allow for the developing new and diverse cultivars that are suitable for growing under a range of biotic and abiotic stress conditions and also have better nutritional quality (Zampieri et al. 2017). The genetic diversity of elite wheat breeding pools through the introduction of beneficial variation is essential to maximizing grain production potential. The identification and introduction of new/novel genetic variability from landraces or related species is one method for expanding the genetic base of contemporary wheat breeding lines (Gaikwad et al. 2023). Wheat landraces are made up of intricately varied, genetically dynamic, and diversified populations that are able to coexist with both biotic and abiotic environmental pressures. The genetic makeup of wheat landraces represents an evolutionary strategy for performance and survival, particularly in arid and semi-arid environments and with organic inputs. The combined effects of natural selection and farmer selection have produced genotype architectures that represent various combinations of traits, including growth habit, tolerance to abiotic stresses, early growth vigour, competition with weeds, disease tolerance, water and nutrient use-efficiency, and quality traits appropriate for a range of subsistence goals and regional food preferences.
A major portion of the genetic diversity that is present for a crop species is frequently seen in landraces. Wheat and rice have been successfully bred using landraces and related species (Swaminathan 2006; Gaikwad et al. 2014). Utilizing one of the free threshing hexaploid species, Triticum sphaerococcum (2n = 6x = 42) in breeding programmes is worth investigating. The cultivation of this landrace, which is native to north-western India and southern Pakistan, dates back to the Bronze Age (Gupta et al. 2021). Its cultivation dates back to ancient times in India, where it was grown in the area between the Hindukush and the Northwest Himalayas (Pal 1966). Around 3000 BC, Indian dwarf wheat T. sphaerococcum was being cultivated, according to archaeological data gathered from the Mohan-jo-Daro excavation (Gupta et al. 2021). Ancient Indian cultures grew it as one of their primary winter crops. However, it vanished from history in the early 20th century, particularly after modern wheat cultivars were introduced in India through the Green Revolution (Mori et al. 2013). The Indian Subcontinent’s hot and dry climate is home to natural populations of this species. The very earlier report suggests that this landrace is early flowering and resistant to yellow rust (Percival 1921), however, we believe that during the course of time this species has become susceptible to rust and this could be one of the reasons for their discontinued cultivation in the farmers’ field.
The unique characteristics of this species like Hemi-spherical glumes and grains, compact head, and short awns are controlled by the pleiotropic effects by a single locus Sphaerococcum 1 (S1), which is located on chromosome 3D (Sears 1947). The genetic identity of the locus and molecular mechanisms underlying the selection of this wheat type is recently revealed (Gupta et al. 2021). The Rht gene of the Green Revolution (Hedden 2003) does not contribute to the semi-dwarf phenotype in Indian dwarf wheat. This suggests that people in ancient India might have utilized their own dwarfing gene thousands of years before the Green Revolution (Mori et al. 2013). Being a landrace, sphaerococcum wheat is now almost extinct from cultivation. Few farmers are growing this wheat in extremely limited areas in states of Rajasthan and Madhya Pradesh. Therefore, it has not been studied in detail. Therefore, the future survival of this species is uncertain.
The Indian Subcontinent’s hot and dry climate is home to natural populations of this species. It enhanced drought tolerance and accumulation of nitrogen and phosphate because of the presence of Brassinosteroid Insensitive 2 (BIN2) gene that contributed to the adaptation of such a small-grain form of wheat to the dry climate of India and Pakistan (Gupta et al. 2021). The occurrence of heat waves in the month of march is now a regular kind of phenomenon. In 2022, the excessive temperatures in March, the steepest since the country’s temperature record-keeping began in 1901, impacted wheat production. Yields are 10–15% down, driving India’s forecasted market year (MY) 2022/23 (April-March) wheat production down from 110 million metric tons (MMT) to 99 MMT (FAS, USDA 2022). Under this scenario, the popular wheat cultivars in the farmers’ fields of North and Central India have failed to beat the heat. Thus, exploring heat and moisture stress tolerant germplasm and related species of wheat to tackle the problem of abiotic stresses is a need of an hour. Introgression of abiotic stress tolerance from T. sphaerococcum to bread wheat should be explored.
Very few accessions of T. sphaerococcum have been used in wheat improvement programmes. They do have advantageous qualities, but one drawback of this wheat is that it yields less than bread wheat. Being a landrace, the morpho-physiological and molecular characteristics of sphaerococcum wheat are little understood. Additionally, there are very few accessions available in the major gene banks around the world. For enhanced plant architecture, tolerance to abiotic challenges including heat and moisture stress, and most significantly, nutritional quality features, this species may be a valuable gene pool for wheat breeding. So, it’s crucial to research these resources’ genetic diversity in order to develop conservation strategies. In order to explore for genetic variation for traits that are distinctive and to identify ways to utilise them in the breeding programme, accessions of Indian dwarf wheat were evaluated in this study under optimal, terminal heat stress and deficient irrigation conditions in the field.
Materials and methods
Location, experimental site and environments
The field experiment was carried out in the Research fields of ICAR-Indian Agricultural research Institute, New Delhi which is situated at N 28° 38’ 24.0252”, E 77° 10’ 26.328” and altitude of 228.61 m (750 ft) above sea level in subtropical region of North Western Plain Zone of India. The experiment was set up in three different production environments: timely sown-irrigated conditions (TS-IR), timely sown restricted irrigation (TS-RI), and irrigated late sown conditions (IR-LS). The trials were planted on 30th October 2021 and 10th November 2021 for restricted irrigated and timely sown conditions respectively. The late sown trial was planted on 20th December 2021.
Plant material
The experimental materials consist of 116 accessions of T. sphaerococcum procured from three gene banks. Out of these 32 accessions were procured from IPK Germany, 27 accessions from Institute of Plant Genetic Resources “Konstantin Malkov”, Sadovo, Bulgaria, 31 accessions from National Small Grain Collections, USDA, 22 accessions from Punjab Agricultural University Ludhiana, India and 5 accessions were available from farmer’s field. Twenty bread wheat commercially cultivars were used as checks according to their production environments. Cultivars HD 3226, HD 3086, HD 2967, DBW 327, DBW 187, DBW 303, HDCSW 18 were used as checks in TS-IR trial, HD 3171, HD 3293, HI 1628, HI 1612, HI 1605, HI 1620, C 306 were used in TS-RI trial, and HD 3298, HD 3271, HI 1633, and HI 1634 were used in IR-LS trial. The details of the T. sphaerococcum accessions are given in Table S1.
Field experiment and data recording
The experiment was laid out in two replications using a randomized block design (RBD). When adequate moisture was available, genotypes were seeded in the field. In each replication, genotypes were grown in 2 m2 plot. Depending on the rainfall, the field was irrigated at regular intervals for TS-IR and LS-IR trials. For TS-RI trial, apart from the pre sowing irrigation, the first irrigation was given at 30 days after sowing and thereafter no irrigation was given to the trial. Rainfall in the month of January completed the requirement of second irrigation in TS-RI trial. Recommended standard cultural and agronomic practices were followed to raise a healthy crop. Agronomic traits including the day to flowering (DTF), days to physiological maturity (DPM), plant height (PH), tillers per mt row length (TPM), spike length (SL), grains per spike (GRPS), 1000 grain weight (TGW), and yield per hectare (YLD) were measured. In addition to these traits, the geometric parameters of grains, namely grain length (GL), grain breadth (GB), and grain surface area (GSA), were assessed using the software SmartGrain Ver.1.3 (Tanabata et al. 2012). In the present study two physiological traits were also recorded. These includes, Normalized Difference Vegetative Index (NDVI) and canopy temperature measurements taken at four different growth stages: active tillering (NDVI 1 and CT1), heading (NDVI 2 and CT2), grain filling (NDVI 3 and CT 3), and physiological maturity (NDVI 4 and CT 4) respectively.
Environmental evaluation
The weather parameters were collected from the Division of Agri. Physics, ICAR-IARI, New Delhi, for the year 2021-2022 during crop growing season (November to May) (Fig. 1a). The TS-RI and TS-IR conditions faced a temperature of around 29.4 °C and late sown conditions had temperature of 20.6 °C during the sowing. At the anthesis stage for TS-RI trial the mean maximum temperature was around 21 °C, relative humidity (RH) was 91% and sunshine hours was around 8.5; for TS-IR trial the mean maximum temperature was around 25.3 °C, RH was 91%, and sunshine hours was around 6.7; for IR-LS trial the mean maximum temperature was around 32.4 °C, relative humidity (RH) was 83% and sunshine hours was around 8.4. A major event of rainfall (95 mm) on 8th and 9th January 2022 completed the requirement of one irrigation. No rainfall was recorded after the anthesis stage in all the three trials. The TS-IR and TS-RI crop experienced effect of heat stress during grain filling stage whereas the late sown crop have experienced heat stress during anthesis to grain filling stage (Fig. 1b). When encountered anthesis and post-anthesis stage, high temperatures [> 28–30 °C] (Vignjevic et al. 2015) can dramatically shrink mature grain weight in wheat, lowering yields (Kino et al. 2020). From the last fortnight of March to first week of April, India experienced sudden rise in temperature due to which the wheat crop which was at anthesis and grain filling stage experienced terminal heat stress. This unusual heat of March 2022 led to lower wheat production by 4 to 5 million tonnes.
Statistical analysis
Statistical analysis was done using the mean values of the observations recorded on five randomly selected plants from each of the replication. Analysis of variance (ANOVA) was analyzed as a randomized block design with STAR Software developed by IRRI. The obtained data were statistically analyzed and least significant difference (LSD) values were performed at the 5% and 1% probability level. The phenotypic, genotypic coefficients of variation were calculated by “variability” pacakge in R studio. The correlation between the traits under the present study was analysed through the “corrplot” package (Wei et al. 2017) in R studio. The relation among environments, mean vs. stability, and ‘which-won-where’ were estimated and represented in the form of biplot graphs using PB Tools software developed by IRRI.

a Weekly weather parameter data of 2021-22 wheat growing season of IARI, New Delhi b Weather parameter data from second fortnight of march 2022 to first fortnight of April 2022
Results
Assessment of genetic variability
Analysis of variance revealed significant difference among T. sphaerococcum accessions for all the agronomic and physiological traits under all three production environments (Table 1).
The genotype × environment (G×E) interaction was also significant for all the traits studied (Table 1). These results indicated a high variation among genotypes that allow us to choose superior genotypes under terminal heat stress and deficient irrigation conditions. A quite noticeable range was observed for most of the traits in 116 accessions of T. sphaerococcum. The DTF had range of 100–141 days under TS-IR, 98–136 under TS-RI and 75–102 under IR-LS conditions, PH had range of 77.4 to 148.5 cm under IR-TS, 77.3–134.5 cm under TS-RI and 78.62–132.8 under IR-LS conditions. On of the grain geometric parameters like GSA had 0.58–2.11 under TS-IR, 1.07–2.22 under TS-RI, and 0.69–2.04 under LS-IR conditions. The grain yield observed good range of variation under all three production environments like 6.69–48.25 q/ha under TS-IR, 5.44–44.09 q/ha under TS-RI, and 3.23–42.44 q/ha under LS-IR conditions. The physiological parameters like NDVI4 observed range of 0.13–0.42 under TS-IR, 0.19–0.55 under TS-RI, and 0.20–0.40 under LS-IR conditions. The CT4 observed range of 31.60–42.4 °C under TS-IR, 35.5–41.7 °C under TS-RI, and 32.6–40 °C under LS-IR conditions. The box plots (Fig. 2) represented the genetic variation between the T. sphaerococcum accessions and bread wheat cultivars released as checks evaluated under three production environments. For DTF and DTM, the released cultivars were earlier than sphaerococcum accessions by at least15 days under TS-IR and TS-RI and 10 days under LS-IR conditions. The PH of T. sphaerococcum accessions is significantly lower than released cultivars (range 85–95 cm) under all production environments. The yield contributing traits like TPM, SL, GRPS were significantly higher in released cultivars. The TGW and grain yield of T. sphaerococcum accessions was significantly lower than released cultivars in all production environments. When T. sphaerococcum accessions were compared for average grain yield under stress environments like TS-RI (28.85 q/ha) and LS-IR (26.15 q/ha), the reduction in yield when compared to TS-IR (33.29 q/ha) is lower, it means these species has tolerance to terminal heat and deficit irrigation conditions. The mean GSA of T. sphaerococcum under stress environments were larger (1.56 in TS-RI and 1.52 in LS-IR) than the GSA under optimum environment. It shows that grain filling was better under stress environment. The mean trait value of physiological parameters like NDVI3 was higher in T. sphaerococcum accessions (0.7 in TS-IR and TS-RI, 0.53 in LS-IR) as compare to released varieties (0.6 in TS-IR and TS-RI, and 0.3 in LS-IR). The mean trait value of CT3 (31.3 °C in TS-IR, 31.8 °C in TS-RI and 31.9 °C in LS-IR) was lower than the released cultivars (37.2 °C in TS-IR, 36.5 °C in TS-RI, and 33.4 °C in LS-IR). The mean values of CT4 also shown similar trend. This shows that T. sphaerococcum accession could able to maintain cooler canopy with high greenness index under stress as well as optimum environments.

Boxplot showing genetic variation among accessions of T. sphaerococcum (SP) and check (C) cultivars for agronomic and physilogical traits under LS-IR, TS-IR, and TS-RI production environments
Components of genetic variance
High GCV values were seen for YLD (41.27%), SL (40.30%), GRPS (30.89%), TGW (24.99%), and NDVI4 (23.85%) (Table 2). GCV values for CT4 (3.19%), DTF (7.06%), and CT3 (7.56%) were found to be low. YLD (44.23%) had the highest magnitude of PCV, followed by SL (42.83%), GRPS (36.92%), TPM (31.69%), and NDVI4 (29.84%). Low PCV magnitudes were seen for CT3 (9.05) and CT4 (6.51%) (Table 2). With the exception of DTF (17%) and CT4 (24%), most of the traits under study have substantial heritability. PH (85%), TGW (82%), YLD (86%), and SL (88%), showed the highest heritability. High genetic advance was noted for TPM (47.15), PH (29.43), and YLD (23.32). The NDVI4 (0.11) and the geometrical parameters of the grains, GB (0.22), GL (0.41), and GSA (0.44), showed a low magnitude of genetic gain. For every character under study in this study, the phenotypic coefficient of variation (PCV) was slightly greater than the corresponding genotypic coefficient of variation (GCV), suggesting that the environment has effect on how a character expresses themselves. The expected genetic advance over mean (GAM) was in the range (3.22–78.98%) and highest expected genetic advance was observed for YLD followed by SL (78.12), GRPS (53.25), and TGW (46.85). Traits like CT4 (3.22), DTF (6) observed low GAM in the present study (Table 2).
Relationships among the traits
The correlation worked out among the different traits studied in all three production environments is presented in Fig. 3a-c. In TS-IR, the YLD ha significant correlation with TGW (0.82***), GRPS (0.74***), GL (0.54*), and GB (0.52*). YLD also has high correlation value GSA (0.45) however, it is non-significant. The YLD has significant negative correlation with DTF (− 0.82***) and DTM (-0.67**) (Fig. 3a). The key yield contributing traits like GRPS, TGW, GL, GB, GSA have significant negative correlation with DTF. The CT4 has negative significant correlation (− 0.55*) with YLD. It means cooler canopy is positively correlated with high grain yield. Under TS-RI conditions (Fig. 3b), YLD ha significant correlation with TGW (0.83***), GRPS (0.67***), GL (0.46*), and GB (0.49*). The YLD has significant negative correlation with DTF (− 0.85***) and DTM (-0.54**). The CT4 has negative significant correlation (− 0.63*) with YLD. The cooler canopy is responsible for high grain yield under moisture deficit conditions. The important yield contributing traits like GRPS, TGW, and GB have significant negative correlation with DTF. Under LS-IR conditions (Fig. 3c), the CT1 and CT4 have negative significant correlation (− 0.63**, and − 0.74***) with YLD. It shows that the T. sphaerococcum accessions could able to maintain cooler canopy under heat stress conditions and is responsible for high grain yield. The YLD has significant positive correlation with TGW (0.82***), and GRPS (0.92**), it also has positive correlation with TPM (0.31) however, it is non-significant. The GL and GSA have negative correlation viz., − 0.48*, and − 0.35 respectively with the YLD. This is apparent because under heat stress conditions the grain size decreases. The correlation of GSA with YLD is negative but non-significant, it shows that the effect of heat stress is not much on the grain size and ultimately over the yield. In all three productions environments, TGW, GRPS, GL, GB, and GSA showed positive correlation and DTF, DM and PH shows negative correlations with YLD.

a Correlation among traits under TS-IR conditions, b Correlation among traits under TS-RI conditions, and c Correlation among traits under LS-IR conditions
Principal component analysis
Biplot analysis was performed by comparing the eigenvalues of PC1 and PC2 of principal component analysis (PCA) for all the genotypes and the morpho-physiological traits in order to deeply ascertain the multivariate correlations between accessions and the production environments (Fig. 4). The results of the first two PC axes (PC1, 28.9% and PC2, 19.6%) accounted for almost 48.5% of the overall variability regarding the relationship between the traits and genotypes, illustrating the intricacy of the variation between the depicted components. The biplot displays a positive correlation between the vectors representing traits (variables) with acute angles, negative correlations for those with obtuse or straight angles, and no connection for those with right angles (Fig. 5). According to the PCA, over 73.19% of the gross variance could be described by the first five vectors combined that had eigen values greater than one (Table 3). The first PC explained 28.90% of the variation, whereas the values for the second through fifth PCs were 19.61%, 10.81%, 7.47%, and 6.39%, respectively. The first PC included traits pertaining to physiological traits, such as flowering, greenness index, and canopy temperature. The main contributors of the variation seen in the first PC were CT1, DTM, CT2, NDVI3, DTF, NDVI1, and GRPS.

Contribution of variables to principal components 1 (PC1) and 2 (PC2)

Principal component analysis (PCA) based biplot showing correlation among the traits under three production environments
Figure 5 characters having the biggest absolute values within the first principal component, closer to unity, are said to have a greater influence on the clustering than characters with lower absolute values, closer to zero. Grain-related traits like GSA, GL, and GB as well as yield-contributing traits like SL, TGW, CT3, and YLD were the primary causes of the variation in the second PC (Fig. 5). For the third PC, the traits NDVI4, CT4, and PH stand in for; for the fourth PC, SL and TPM; and for the fifth PC, NDVI2 and PH. (Table 3) The significance of the biggest contributor to the overall variance at each differentiation axis is shown in the principal component analysis. A biplot of genotype by trait showed the spatial variety of accessions in two dimensions, resembling a clustering pattern where distinct clusters were represented. Because both production environments—TS-IR and TS-RI—were timely seeded and had comparable sowing dates, they clustered together. But the IR-LS environment can be easily identified from the other two (Fig. 5). Larger vector lengths for traits like CT1, CT2, DTM, DTF, GRPS, NDVI3, YLD, GSA, TGW, and SL indicate high variability for these traits. There was a strong association found between significant trails such as YLD and TGW, GRPS, GL, GSA, SL, GB, and CT4. Traits such as CT1 and CT2 are highly correlated. In the same way, there was positive association between YLD and TGW and the geometric parameters of the grain in all three environments.
Mean yield performance among the environments
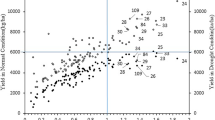
In three productions environments all 116 T. sphaerococcum accessions along with the 20 released cultivars as checks representing each production environment were evaluated for agronomic and physiological traits. As discussed earlier mean yield of all 116 T. sphaerococcum accessions in three trials was TS-IR (33.29 q/ha), TS-RI (28.95 q/ha) and LS-IR (26.15 q/ha). When compared to the bread wheat released cultivars these yield levels are very low, this is because the grains are smaller and weigh less. Under stress conditions, one or two accessions, however, were good yielders and comparable to the checks. Although it is illogical to compare T. sphaerococcum with T. aestivum, we are interested in knowing the yield potential of T. sphaerococcum accessions in relation to bread wheat cultivars. Among the 116 accessions, we have selected 45 accessions which resembles true plant type of T. sphaerococcum, their grain yield is presented in Fig. 6a, b, c. In TS-IR trial (Fig. 6a), all the checks yielded from 55 to 74.28 q/ha. The cultivar DBW 187 yield highest with 74.28 q/ha followed by HD 3226 with 69.33 q/ha. Among sphaerococcum accessions the yield range was from 20 q/ha to 47.2 q/ha. Few accessions like TS49 (47.25 q/ha), TS5, TS6 (44.7 q/ha), TS56 (42.8 q/ha) and Sugar free, TS61, TS27, TS17, PAUTS16 yielded more than 40 q/ha (Figure a). In TS-RI condition (Fig. 6b), TS49 has more yield (44.09 q/ha) as compared to bread wheat cultivars HI 1612 (43.35 q/ha), and HI 1628 (42.62 q/ha) (Figure b). Under TS-RI condition, the yield gap between T. sphaerococcum accessions and bread wheat cultivars is not much. It shows that, T. sphaerococcum have tolerance to moisture deficit conditions. The yield range among sphaerococcum accessions was from 13 q/ha to 44 q/ha. Accessions like TS27, TS6, TS5, PAUTS16, TS56, TS17 have yield more than 37 q/ha under moisture deficit conditions. Compare to check C 306 (29.2 q/ha) which is a drought tolerant cultivar, 25 accessions of T. sphaerococcum recorded significantly higher yield. Under, LS-IR conditions (Fig. 6c), the yield of T. sphaerococcum accessions ranged from 19.6 to 40.2 q/ha and checks ranged from 36.5 to 45.9 q/ha. TS49 (40.2 q/ha) and TS61(38.28 q/ha) have statistically higher yield than check HI 1621 (36.57 q/ha) and at par with the checks like HD 3271 (40.15 q/ha), HD 3298 (39.57 q/ha). Accessions like PAUTS16, Sugar free, PAUTS1, TS56, PAUTS17 and PAUTS19 had yield more than 35 q/ha under heat stress conditions.
GGE biplot analysis
For the visual examination of the relationships among the test environments, genotypes, and the genotype × environment (G×E) interactions GGE biplot analysis was performed on the 116 T. sphaerococcum accessions excluding the checks. Biplots were constructed for all the traits, however, two important traits TGW and YLD are discussed. In environment view plot for TGW, environment E1 (LS-IR) and E3 (TS-IR) are closer to the ideal environment and are highly correlated to each other (Fig. 7a). Environment E2 (TS-RI) is away from E1 an E3 and therefore distinct form these two. In genotype view biplot, genotype G70 (TS45) was ideal genotype however, due to its longer vector length from AEA, it is specific to TS-RI environment. Genotypes G78 (TS49), G28 (TS12), G27 (TS11-1), G91 (TS6), G43 (TS24), G110 (TS77) were stable in performance and having high mean value for the trait. In which-won-where biplot, all the accessions were distributed in 5 clusters (Fig. 7a). Environments E1 and E3 were in similar cluster where genotypes G70 (TS45), and G78 (TS49) were winning. Genotypes G107 (TS74), and G66 (TS41) were specific for environment E2. In environment view plot for YLD, environment E2 (TS-RI) and E3 (TS-IR) are closer to the ideal environment and are correlated to each other (Fig. 7b). Environment E1 (LS-IR) is away from E3 an E2 and therefore distinct form these two. In genotype view biplot, genotype G78 (TS45) and G24 (TS1) were having high mean performance and stability hence, an ideal genotype. In which-won-where biplot, all the accessions were distributed in 4 clusters (Fig. 7b). Although all the environments lie in the same cluster, LS-IR environment is different from the rest two. For all the environments G78 (TS49), and G24 (TS1), were winning genotypes.

a Mean grain yield (q/ha) of selected 45 accessions of T. sphaerococcum under TS-IR, b under TS-Ri, and c under LS-IR condition

a GGT biplot showing environment, genotype view and what-won-where plot for 1000 grain weight (TGW), and b GGT biplot showing environment, genotype view and what-won-where plot for grain yield (YLD)
Discussion
For millennia, traditional wheat farming communities have contributed to the evolution and enrichment of on-farm conservation of varied wheat landraces by establishing efficient seed exchange systems that support the landraces’ continued expansion and diversification. Wheat landraces are composed of genetically dynamic, highly diversified, and intricately differentiated populations that can withstand both biotic and abiotic environmental stresses. Wheat landraces’ genetic composition reflects an evolutionary strategy for adaptation and survival, especially in dry and semi-arid conditions and with organic inputs. Since high-yielding cultivars were introduced and wheat agricultural systems underwent structural changes, wheat landraces have lost genetic diversity. However, the fact that some wheat landraces are still farmed indicates either how important they will be to farmers in the future or how much better they are than current cultivars in terms of nutrition or agronomy. The present study aimed to characterize one of such ancient landraces of Indian sub-continent under optimum, terminal heat stress and deficit irrigation conditions. Significant differences were found by analysis of variance between T. sphaerococcum accessions for all agronomic and physiological parameters in each of the three production conditions. For every trait under investigation, the genotype × environment (G×E) interaction was also significant. These findings showed significant genotype diversity, which enables us to select better genotypes in situations of extreme heat stress and inadequate irrigation. Prior research has not documented the phenotyping of these many T. sphaerococcum accessions under optimum and abiotic stress conditions. Thus, the study’s results are examined in respect to reports in bread wheat.
In the current study, the YLD had the greatest GCV, followed by SL, GRPS, and TGW. For DTF, CT3, and CT4, the magnitude of PCV and GCV was minimal (Table 2). Similar findings in bread wheat have previously been reported (Bhushan et al. 2013; Yadav et al. 2014; Hossain et al. 2021). For the majority of the characters, the heritability estimates were relatively high, indicating a strong genetic component to all of the features. Given this genotype set’s increased heritability, selection for the majority of the traits may be successful. For YLD, PH, GRPS, and geometric grain parameters, significant heritability estimates were found in conjunction with strong genetic advance. These findings suggested that the aforementioned traits were controlled by additive gene action and would, therefore, be expected to show improvement by direct selection. Nukasani et al. (2013) also reported findings that were similar. There are no reports available on T. sphaerococcum, and all of these experiments have been conducted on bread wheat.
The Indian dwarf wheat accessions were having both generative and more vegetative tillers however, generative tillers were less as compared to bread wheat cultivars. Similar findings were also reported by Szczepanek et al. (2020) in this specie. This species was adapted to dry climates of the Indian sub continents, hence, the more biomass at the early growth period under deficit water conditions is helpful. In the present study, when T. sphaerococcum accessions were compared for average grain yield under optimum environment (33.29 q/ha) and under stress environments like TS-RI (28.85 q/ha) and LS-IR (26.15 q/ha). The reduction in yield under TS-RI condition is 13.33% and LS-IR is 21.44%. This magnitude of reduction in grain yield is lower when compare to bread wheat. Under terminal heat stress, significant yield reduction of 30–47.5% in bread wheat was accounted (Kumar et al. 2021; Poudel et al. 2021) it means these species has tolerance to terminal heat and deficit irrigation conditions. Heat waves during the month of March are becoming a common occurrence. Excessive temperatures in March 2022, the highest since the country’s temperature record-keeping began in 1901, hampered wheat production. Yields are 10–15% down, driving India’s forecasted market year (MY) 2022/23 (April-March) wheat production down from 110 million metric tons (MMT) to 99 MMT (FAS, USDA 2022). The high temperature (Fig. 1a and b) at IARI, New Delhi, coincided with the anthesis stage in the LS-IR experiment and the grain filling period in the TS-IR and TS-RI trials. In the LS-IR experiments, the sphaerococcum accessions performed best with the least amount of yield drop at such a high temperature. This illustrates the species’ ability to withstand extreme heat stress. Recently work has been done to elucidate the drought tolerance ability of this species. Gupta et al. (2021) investigated the genetic basis of drought tolerance in T. sphaerococcum. They discovered that a GSK3-like kinase similar to the Arabidopsis BIN2 protein, a negative regulator of brassinosteroid signalling, is encoded by the S1 locus. The S1 protein is insensitive to brassinosteroid, which upregulates brassinosteroid biosynthesis genes and dramatically improves drought tolerance. The improved drought tolerance and nitrogen and phosphate accumulation helped to the adaptation of such a small-grain wheat to the dry environment of India and Pakistan. Previously Bansal and Sinha (1991) tested accession of T. sphaerococcum for drought tolerance and yield stability. They found that on the basis of grain yield stability, T. sphaerococcum, T. vavilovii and T. aestivum cv. C 306 exhibited better yield stability and drought resistance than the other species tested.
Understanding relationships among evaluated agronomic and physiological traits could provide useful information for wheat breeding programs. The Canopy temperature is one of the key physiological factors that affects yield under stress. Due to its strong adverse correlations with wheat’s thousand-grain weight, grain weight per spike, and grain number per spike when exposed to terminal heat stress in the field, this trait has been suggested as a possible selection criterion for breeding programmes (Blum et al. 1989). In the current study, CT4 demonstrated a strong negative correlation with GRPS (-0.89***), TGW (-0.64**), and YLD (− 0.74**) under LS-IR conditions and TGW (− 0.45*) and YLD (− 0.63**) under TS-RI conditions. The physiological changes that a plant makes in response to abiotic stress—such as heat and drought—often involve trade-offs. Although not the only explanation, this trade-off helps to explain the strong correlation between the stresses of heat and drought (Reynolds et al. 2007). Our research revealed a comparable trade-off between YLD and canopy temperature (CT1 and CT 4); however, NDVI3, and NDVI4 was significantly negative under heat and moisture deficient conditions. These results are consistent with previous research conducted under late heat and drought stress conditions in a landrace T. compactum (Blum et al. 1989), durum wheat (Gautam et al. 2015) bread wheat (Rutkoski et al. 2016; Thapa et al. 2018; Yashavanthakumar et al. 2021; Thakur et al. 2022). There is currently no published research on T. sphaerococcum on this aspect.
The yield in both optimum and stress conditions was significantly positively correlated with the two major yield-contributing traits: grains per spike and thousand-grain weight. Therefore, breeding for stable genotypes appropriate for stress and non-stress conditions requires selection for these traits (Dwivedi et al. 2018; Gupta et al. 2020; Yashavanthakumar et al. 2021). Under both stress conditions in the current study, there was a strong positive correlation between YLD and TGW and GRPS. Under TS-RI conditions, the geometric parameters of the grain in this study demonstrated a significant positive association with YLD. Limited irrigations have enhanced the overall grain lustre and GSA under TS-RI conditions in T. sphaerococcum. Such observations are prevalent in bread wheat cultivated in central India, where moisture is scarce towards the conclusion of the harvest season. Sheng et al.‘s (2022) research on bread wheat has also shown that optimum water management can improve wheat’s end-use quality without reducing grain yield. The relationship between YLD, DH, and DTM in this study was inversely correlated with each stress scenario. Breeding for drought and high-temperature tolerance has acknowledged this indicated drought escape mechanism by earliness and short duration as an important trait (Gupta et al. 2020; Yashavanthakumar et al. 2021; Mondal et al. 2013). The principal component analysis inferred a significant relationship between yield and TGW, GSA, SL, and CT4, all of which are useful selection traits for use in breeding of wheat for abiotic stress tolerance. The importance of these traits in ideotype breeding suitable for heat and drought stress is indicated by the substantial positive connection seen between grain yield and the aforementioned traits. Our findings for T. sphaerococcum are consistent with those of Manu et al. (2020) in bread wheat.
Due to the strong environmental influence on quantitative traits, the genotype selection process form multi-environment trail, is frequently challenging. In order to detect and select stable genotypes, multi-environment trials utilising pertinent statistical approaches, such as GGE, are helpful. A GGE biplot makes it simple to see which genotypes perform best in each environment and across all habitats. The accessions TS49, TS45, TS1, TS6, TS24, and TS77 in our investigation performed consistently in each production conditions. Out of all the evaluated production conditions, TS49 had the highest level of stability and yield. These wheat genotypes have shown to be the most promising as possible donors in breeding initiatives creating drought-tolerant wheat cultivars to counteract climate change’s effects and could be used for developing mapping populations to identify quantitative trait loci. Under controlled circumstances, accurate responses to genotype-by-environment interaction can be evaluated, but it is frequently challenging to correlate these with real-world reactions (Talukder et al. 2014). Thus, it is necessary to establish robust, relevant, repeatable, and highly heritable phenotyping methods.
As indicated earlier, phenotyping of this large number (116) of T. sphaerococcum accessions under optimum and abiotic stress conditions has not been documented in prior research. However, Das et al. (2023) have recently looked at traits related to nitrogen use efficiency (NUE) and nitrogen deficiency tolerance (NDT) in 11 Indian dwarf wheat accessions in order to study this species’ ability to flourish in low nitrogen conditions. There have been reports of a small number of accessions being assessed under organic conditions. Szczepanek et al. (2020) assessed the Persian wheat cultivar “Persa” and T. sphaerococcum cultivar called “Trispa” in an organic environment. They reported that compared to Persian wheat (2.15 Mg ha-1), the grain yield of Indian dwarf wheat (2.83 Mg ha-1) was noticeably higher. Additionally, they have reported that T. sphaerococcum yields better (1 qt/ha more) than bread wheat when grown in organic environments. T. sphaerococcum accessions in our study likewise had comparable average grain yields in TS-IR (33.29 q/ha), TS-RI (28.95 q/ha), and LS-IR (26.15 q/ha) production environments. Nonetheless, as compared to bread wheat cultivars, a small number of accessions exhibit superior grain yielding under stress in our study. The sowing density of T. sphaerococcum has been standardized by Szczepanek et al. (2022) in order to increase grain production. They came to the conclusion that a sowing density of 600 pieces per millimetre squared, along with mild droughts throughout the growth season, results in a better yield of T. sphaerococcum in integrated low-input agriculture. These investigations demonstrate that this species does better in environments with lower moisture requirements and can be a viable donor for wheat that can withstand moisture stress.
Indian dwarf wheat has a number of desirable characteristics these include uniformly sized grains, sturdy but short culms, upright leaves, resilience to high temperatures, moisture stress, and high baking quality. T. sphaerococcum has, however, not been extensively used in breeding programmes to introduce desirable traits. One wheat cultivar developed by ICAR-IARI, New Delhi, namely HD 2888 (*C306/T. sphaerococcum/ HW 2004), which has one of the parent T. sphaerococcum, was released for cultivation in rainfed locations in the North Eastern plains of India (Singh et al. 2007). Using this species in their hybridization programme, Askhadullin et al. (2021) produced a better line, Sh-359-11-5, with a greater yield than its Indian dwarf wheat parent, k-45,738. The outcomes of these two initiatives show that it is possible to select for improvement while maintaining T. sphaerococcum’s advantageous traits.
Conclusion
Our research presents, for the first time, the characteristics of agronomical and physiological traits in ancient landrace Indian dwarf wheat under optimum, terminal heat stress and deficit irrigation conditions. The current study’s accessions showed significant degrees of variation, which were represented statistically via principal component and correlation analyses. Assessment of T. sphaerococcum accessions under conditions of terminal heat and moisture stress confirmed that they have the ability to function better under stress. This species may be of interest in the breeding of climate-resilient wheat cultivars due to the smaller magnitude of decline in grain production under stress conditions when compared to bread wheat. Under optimum, terminal heat stress, and deficit irrigation conditions, accessions like as TS49 emerged as the best performing genotype. Although landraces such as T. sphaerococcum cannot be directly compared to contemporary improved cultivars, their use in breeding programmes ought to be promoted. If farmers are to be encouraged to cultivate and manage wheat landraces on their farms, the landraces’ social worth needs to be elevated to the level of high-yielding wheat cultivars, or even higher. This will facilitate the continuation of the evolutionary processes that shape landraces, particularly in their centres of diversity.
References
Askhadullin DF, Askhadullin DF, Vasilova NZ, Lysenko NS (2021) Prospects of creating Indian dwarf wheat varieties Triticum sphaerococcum Perciv. based on samples endemic to the Hindustan Peninsula. Indian J Genet Plant Breed 81(03):383–389
Bansal KC, Sinha SK (1991) Assessment of drought resistance in 20 accessions of Triticum aestivum and related species I. Total dry matter and grain yield stability. Euphytica 56:7–14
Bhushan B, Bharti S, Ojha A, Pandey M, Gaurav SS, Tyagi BS, Singh G (2013) Genetic variability, correlation coefficient and path analysis of some quantitative traits in bread wheat. J Wheat Res 5(1):21–26
Blum A, Golan G, Mayer J, Sinmena B, Shpiler L, Burra J (1989) The drought response of landraces of wheat from the northern Negev Desert in Israel. Euphytica 43:87–96
Das S, Singh D, Meena HS, Jha SK, Kumari J, Chinnusamy V, Sathee L (2023) Long term nitrogen deficiency alters expression of miRNAs and alters nitrogen metabolism and root architecture in Indian dwarf wheat (Triticum sphaerococcum Perc.) Genotypes. Sci Rep 13(1):5002
Dwivedi SK, Arora A, Singh VP, Singh GP (2018) Induction of water deficit tolerance in wheat due to exogenous application of plant growth regulators: membrane stability, water relations, and photosynthesis. Photosynthetica 56:478–486
FAS, USDA (2022) https://fas.usda.gov/data/india-india-extreme-temperatures-scorch-indian-wheat-production
Gaikwad KB, Singh N, Bhatia D, Kaur R, Bains NS, Bharaj TS, Singh K (2014) Yield-enhancing heterotic QTL transferred from wild species to cultivated rice Oryzasativa L. PLoS ONE 9(6):e96939
Gaikwad KB, Yadav R, Kumar M, Kumar N, Babu P, Singh AG, Ansari R, Pandey R (2023) Synthetic hexaploid wheat as a source of variation for the traits specific to conservation agriculture. Indian J Genet Plant Breed 83(01):32–40
Gautam A, Prasad SV, Jajoo A, Ambati D (2015) Canopy temperature as a selection parameter for Grain Yield and its components in Durum Wheat under terminal heat stress in late sown conditions. Agric Res 4:238–244
Gupta PK, Balyan HS, Sharma S, Kumar R (2020) Genetics of yield, abiotic stress tolerance and biofortification in wheat (Triticum aestivum L). Theor Appl Genet 133:1569–1602
Gupta A, Hua L, Lin G, Molnár I, Doležel J, Liu S, Li W (2021) Multiple origins of Indian dwarf wheat by mutations targeting the TREE domain of a GSK3-like kinase for drought tolerance, phosphate uptake, and grain quality. Theor Appl Genet 134(2):633–645
Hedden P (2003) The genes of the green revolution. Trends Genet 19(1):5–9
Hossain MM, Azad MAK, Alam MS, Eaton TEJ (2021) Estimation of variability, heritability and genetic advance for phenological, physiological and yield contributing attributes in wheat genotypes under heat stress condition. Am J Plant Sc 12(04):586
Kino RI, Pellny TK, Mitchell RA, Gonzalez-Uriarte A, Tosi P (2020) High post-anthesis temperature effects on bread wheat (Triticum aestivum L.) grain transcriptome during early grain-filling. BMC Plant Biol 20(1):1–7
Kumar P, Gupta V, Singh G, Singh C, Tyagi BS, Singh GP (2021) Assessment of terminal heat tolerance based on agro-morphological and stress selection indices in wheat. Cereal Res Commun 49:217–226
Manu B, Kumara PS, Biradar S, Phuke R, Ambati D, Prasad SS et al (2020) Genetic gain and morpho-physiological characterisation of BILs (Backcross inbred lines) under different moisture regimes in wheat (Triticumaestivum L.). Indian J Genet Plant Breed 80(01):84–93
Mondal S, Singh RP, Crossa J, Huerta-Espino J, Sharma I, Chatrath R, Singh GP, Sohu VS, Mavi GS, Sukuru VSP, Kalappanavar IK, Mishra VK, Hussain M, Gautam NR, Uddin J, Barma NCD, Hakim A, Joshi AK (2013) Earliness in wheat: a key to adaptation under terminal and continual high temperature stress in South Asia. Field Crops Res 151:19–26
Mori N, Ohta S, Chiba H, Takagi T, Niimi Y, Shinde V, Osada T (2013) Rediscovery of Indian dwarf wheat (Triticum aestivum L. ssp. sphaerococcum (Perc.) MK.) an ancient crop of the Indian subcontinent. Genet Resour Crop Evol 60(6):1771–1775
Nukasani V, Potdukhe NR, Bharad S, Deshmukh S, Shinde SM (2013) Genetic variability, correlation and path analysis in wheat. J Wheat Res 5(2):48–51
Pal BP (1966) Wheat, ICAR Monograph. Indian Council of Agricultural Research, New Delhi
Percival J (1921) The wheat plant (a monograph). Duckworth and Co, London
Poudel PB, Poudel MR, Puri RR (2021) Evaluation of heat stress tolerance in spring wheat (Triticum aestivum L.) genotypes using stress tolerance indices in western region of Nepal. J Agric Food Res 5:100179
Reynolds MP, Pierre CS, Saad AS, Vargas M, Condon AG (2007) Evaluating potential genetic gains in wheat associated with stress-adaptive trait expression in elite genetic resources under drought and heat stress. Crop Sci 47:172–189
Rutkoski J, Poland J, Mondal S, Autrique E, Pérez LG, Crossa J, Reynolds M, Singh R (2016) Canopy temperature and vegetation indices from high-throughput phenotyping improve accuracy of pedigree and genomic selection for grain yield in wheat. G3 Genes Genomes Genet 6:2799–2808
Sears ER (1947) The sphaerococcum gene in wheat. Genetics 32:102–103
Sheng K, Xu L, Wang M, Lei H, Duan A (2022) The end-use quality of wheat can be enhanced by optimal water management without incurring yield loss. Front Plant Sci 13:1030763
Singh B, Tomar SMS, Vinod SM, Chowdhary S (2007) Notification of crop varieties and registration of germplasm. Indian J Genet Plant Breed 67(4):418–422
Swaminathan MS (2006) An evergreen revolution. Crop Sci 46(5):2293–2303
Szczepanek M, Lemańczyk G, Lamparski R, Wilczewski E, Graczyk R, Nowak R, Prus P (2020) Ancient wheat species (Triticum sphaerococcum Perc. and T. persicum Vav.) in organic farming: Influence of sowing density on agronomic traits, pests and diseases occurrence, and weed infestation. Agriculture 10(11):556
Szczepanek M, Lemańczyk G, Nowak R, Graczyk R (2022) Response of Indian dwarf wheat and persian wheat to sowing density and hydrothermal conditions of the growing seasons. Agriculture 12(2):205
Talukder ASMHM, McDonald GK, Gill GS (2014) Effect of short-term heat stress prior to flowering and early grain set on the grain yield of wheat. Field Crops Res 160:54–63
Tanabata T, Shibaya T, Hori K, Ebana K, Yano M (2012) SmartGrain: high-throughput phenotyping software for measuring seed shape through image analysis. Plant Physiol 160(4):1871–1880
Thakur V, Rane J, Nankar AN (2022) Comparative analysis of canopy cooling in wheat under high temperature and drought stress. Agronomy 12(4):978
Thapa S, Jessup KE, Pradhan GP, Rudd JC, Liu S, Mahan JR, Devkota RN, Baker JA, Xue Q (2018) Canopy temperature depression at grain filling correlates to winter wheat yield in the U.S. Southern High Plains. Field Crops Res 217:11–19
Velu G, Singh RP, Huerta J, Guzmán C (2017) Genetic impact of Rht dwarfing genes on grain micronutrients concentration in wheat. Field Crop Res 214:373–377
Vignjevic M, Wang X, Olesen JE, Wollenweber B (2015) Traits in spring wheat cultivars associated with yield loss caused by a heat stress episode after Anthesis. J Agron Crop Sci 201:32–48
Wei T, Simko V, Levy M, Xie Y, Jin Y, Zemla J (2017) Package ‘corrplot.’ Statistician 56(316):e24
Yadav SK, Singh AK, Baghel SS, Jarman M, Singh AK (2014) Assessment of genetic variability and diversity for yield and it’s contributing traits among CIMMYT based wheat germplasm. J Wheat Res 6(2):154–159
Yashavanthakumar KJ, Baviskar VS, Navathe S et al (2021) Impact of heat and drought stress on phenological development and yield in bread wheat. Plant Physiol Rep 26:357–367
Zampieri M, Ceglar A, Dentener F, Toreti A (2017) Wheat yield loss attributable to heat waves, drought and water excess at the global, national and subnational scales. Environ Res Lett 12(6):064008
Acknowledgements
The researchers would like to acknowledge the Head, Division of Genetics and Joint Director (Research) ICAR-IARI, for providing field support to conduct the trial.
Funding
No specific funds have been received to conduct present research.
Author information
Authors and Affiliations
Contributions
KBG: The collection of experimental material, guided the study, the writing of the manuscript, and the editing of the manuscript. AD, AS, RK: performed the field experiments. PB, MK, NK, AKP: formal analysis and editing of the manuscriptAKM, RA, and NS: review of the original draft. RY: Finalized the drafting.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Gaikwad, K.B., Dawar, A., Singh, A. et al. Trait phenotyping in an ancient Indian landrace of wheat Triticum sphaerococcum under optimum, terminal heat stress and deficit irrigation conditions. Genet Resour Crop Evol 71, 2779–2795 (2024). https://doi.org/10.1007/s10722-023-01817-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-023-01817-z