
Abstract
The present study aimed to understand the pollen morphological variation among and within the Rosa L. species of Western Himalaya, India, and to investigate whether the pollen characters may help to distinguish these species of roses. The pollen morphology of nine Rosa species viz. R. canina L., R. centifolia L., R. damascena Mill., R. foetida var. persiana (Lem.) Rehder, R. moschata Herrm, R. multiflora Thunb, R. nanothamnus Boulenger, R. webbiana Wall. ex Royle, and R. macrophylla Lindl were studied. A total of 31 accessions of roses were collected from Jammu and Kashmir, Ladakh, and Himachal Pradesh. Ten quantitative characters, i.e., length of the polar axis (P), length of equatorial axis (E), exine thickness (Ex), length of colpi (LeC), width of striae (Sr), width of groove (Gr), number of striae (Ns), Polar area index (PAI) and P/E ratio were studied. The measurements and observations were carried out with both light microscopy and scanning electron microscopy. Pollen grains of all the studied species were tricolporate. Prolate and subprolate shapes were the dominant pollen shape types. The highest and lowest polar axis length was observed for R. canina (32.73 µm) and R. moschata (25.42 µm), respectively. The average colpi length in the studied species was 22.19 µm, with R. macrophylla having a minimum length of 13.42 µm and R. canina with a maximum length of 34.03 µm. Results of the principal component analysis indicated that the principal component (PC1) attributed 42.10% of the total variation, and positively associated with P, E, LeC, and Ns. Exine ornamentation almost distinguished all the species of Rosa in this study.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The genus Rosa L. comprises about 200 species distributed globally (Wissemann 2003), although in different studies depending on the taxonomic approach embraced, the number of species reported may vary up to 250 (Hutchinson 1964; Nilsson 1997; Henker 2000; Kalkman 2004). Most of the Rosa species are distributed in the subtropical and temperate habitats of the Northern hemisphere (Gu and Robertson 2003). Asia harbors the majority of the wild rose species (Broertjes and van Harten 1978). Eleven Rosa species grow wild in India (Chopra and Singh 2013), out of which ten Rosa species are found in the Indian Himalayan region (Kaul et al. 1999) between an altitude of 500 and 4700 m a.s.l (Hooker 1879; Ambasta 1986; Duthie 1971; Pal 1991). Wild roses are an important source of valuable germplasm for creating variability and improvement of roses as per the enormous future needs (Dhyani and Singh 2014). Some rose species produce essential oil, which is in high demand in the aroma industry. The rose hips of some rose species are often used in making jams, fruit juice, marmalade (Uggla et al. 2003). In different Indian states, the fruits of Rosa webbiana Wall. ex Royle, Rosa macrophylla Lindl., and Rosa moschata Herrm. have been used for curing various ailments such as lung problems, liver disorders, skin ailments, body inflammation, heart, and gastrointestinal disorders (Devi et al. 2013; Nand and Naithani 2018). Many recent reports have found the fruits (hips) of R. moschata, R. webbiana, and R. canina L. as a rich source of biologically active compounds viz., total phenolics, flavonoids and Vitamin C (Shameh et al. 2019) indicating their potential of having food and food additive value.
The uniformity in the morphological characters, along with the occurrence of hybridization, makes species identification tenacious in this genus Rosa (Fougere-Danezan et al. 2015). There is enormous taxonomic confusion in this genus due to the convoluted evolutionary history of wild species together with the long cultivation history and interbreeding of preferred genotypes (Koopman et al. 2008). Several morphological characters have been suggested by different researchers for species discrimination in wild roses from time to time (Riaz et al. 2011; Cheikh-Affene et al. 2015; Singh et al. 2020). In addition to the morphological characters, various molecular markers such as simple sequence repeats (SSRs), species-specific sequence-characterized amplified region (SCAR), sequence-tagged molecular sites markers (STMS), inter simple sequence repeats (ISSRs), random amplified polymorphic DNAs (RAPDs), and amplified fragment length polymorphisms (AFLPs) have been employed for distinguishing rose cultivars and also to determine genetic relationships (Debener et al. 1996; Besse et al. 2000; Esselink et al. 2003; Mohapatra and Rout 2006; Bashir et al. 2014; Nadeem et al. 2014; Panwar et al. 2015).
Pollen grains have peculiar biological attributes and portray strong genetic conservancy; thus, they can be put to use for species identification (Schori and Fumess 2014; Song et al. 2017; Almeida et al. 2018). The importance of pollen characters such as length of equatorial and polar axes, pollen shape, length of colpi, operculum structure, as well as the presence or lack of costae colpi on Rosa taxonomy is on record (Reitsma 1966; Fogle 1977; Eide 1981; Marcucci et al. 1984; Hebda and Chinnappa 1990, 1994; Ueda 1992; Ueda and Okada 1994; Popek 1996; Shinwari and Khan 2004; Wronska-Pilarek 2011). Perforations on the exine surface in Rosa are numerous and frequently stretch onto tectal striae (Reitsma 1966; Eide 1981; Hebda and Chinnappa 1990, 1994; Wronska-Pilarek 2011; Bai et al. 2011). Our insight concerning Rosa pollen morphology is by no means outright, because the available depictions are generally concise and sometimes restrained to mean dimensions.
The palynological characters of the Rosa species growing in the Western Himalayan region have not been thoroughly investigated yet. Currently, only a single report on the pollen morphology of Rosa sericea Wall. ex Lindl. and two unknown taxa from this critical genus has been reported from India (Ghosh and Saha 2017). The present study aimed to understand the pollen morphological variation among and within the Rosa species of Western Himalaya, India, and to investigate whether the pollen characters may help to distinguish these species of roses.
Material and methods
Extensive field trips were undertaken during spring and later summer seasons between 2015 and 2018 in Western Himalaya covering one Indian state of Himachal Pradesh, and two Union territories of India: Jammu and Kashmir (J&K), and Ladakh (Fig. 1). The total geographical area of the study area is 277,909 km2, which is almost double of the nearby Himalayan country of Nepal (147,181 km2), and comparable to the geographical areas of the countries like United Kingdom (242,495 km2) and New Zealand (270,467 km2). Herbarium specimens of Rosa species were collected and processed following standard taxonomic procedures (Rao and Sharma 1990). Taxonomic identities of the collected Rosa specimens were established using regional floras, eFloras, and monographs of genus Rosa (Andrews 1805; Baker 1869; Lewis 1957; Singh and Kachroo 1976; Kachroo et al. 1977; Sharma and Kachroo 1981; Chowdhery and Wadhwa 1984; eFlora 2019) and regional herbaria (RRLH, CSIR-IIIM, Jammu, J&K; Herbarium of University of Jammu, J&K). Duly identified voucher specimens were deposited to the internationally recognized Janaki Ammal Herbarium (RRLH) at CSIR-IIIM, Jammu. The Plant list Version 1.1 (TPL 2019) was consulted to verify the currently accepted scientific names of the Rosa species.

Map of the study area showing locations of the studied accessions of Rosa from Western Himalaya
The study using light (LM) and scanning electron microscopy (SEM) was conducted on 31 accessions representing nine Rosa species from Western Himalaya (Table 1). Pollen grains, both from fresh (27) and dried herbarium specimens (4) available at RRLH, were studied. Pollen grains were analyzed for ten quantitative pollen characters (Table 2), and qualitative characters like shape, perforation pattern on exine and outline. Exine sculpture elements were measured on an area of 25 µm2 following the methods of Ueda and Tomita (1989). From each plant, several randomly selected flowers were collected. The measurements were based on 30 fully developed pollen grains per individual specimen, as recommended by Wronska-Pilarek et al. (2015), and the measured characters are provided in Table 2. Overall, a total of 930 pollen grains were scrutinized. Pollen grains were mounted in glycerin jelly after acetolysis procedure (Erdtman 1952) with modifications for LM, and the examined characters were measured by Leica light microscope, model DM 750 (Japan) with the aid of a 100X eyepiece. For SEM, after acetolysis, the specimens were mounted on stubs of 12.5 mm diameter and then coated in a sputter coater with approximately 25 nm of gold–palladium and using the SEM model JEOL JSM-IT 300 (JEOL, Ltd., Tokyo, Japan). Pollen shapes and sizes were classified as per the Erdtman classification (1952). The pollen terminology, in general, followed Erdtman (1952), Punt et al. (2007), Hesse et al. (2009) and Halbritter et al. (2018) for describing exine ornamentation.
One-way analysis of variance (ANOVA) was applied to find out the differences in mean values among the taxa for assessing the variability. The Tukey’s post hoc test was used to test differences among means when the F-test was significant (p ≤ 0.05). Principal Component Analysis (PCA) was conducted on quantitative variables, and for hierarchial clustering, Euclidean distance and UPGMA method were used employing the PAST version 3.22 program (Hammer et al. 2001).
Results
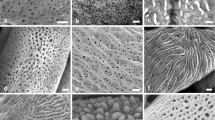
The quantitative pollen characters considered in the present study are summarized in Table 2. Ranges (min–max), mean values (± SE), and coefficient of variation (CV%) of pollen grains morphological characters of different Rosa species from Western Himalaya, India are given in Table 3. Figures 2, 3, 4, 5 and 6 depict LM and SEM micrographs of pollen grains showing variation in pollen shapes and exine sculpture in different accessions of R. moschata (A1a–c to A5a–c), R. webbiana (B1a–c to B12a–c), R. canina (C1a–c), R. foetida var. persiana (D1a–c), R. multiflora (E1a–c and E2a–c), R. centifolia (G1a–c), R. damascena (H1a–c), R. nanothamnus (I1a–c), and R. macrophylla (F1a–c to F7a–c).

LM (first column, A1a–A5a) and SEM micrographs of pollen grains in different R. moschata accessions displaying variation in pollen shape (second column, A1b–A5b) and exine sculpture (third column, A1c–A5c)

LM (first column, B1a–B6a) and SEM micrographs of pollen grains in different R. webbiana accessions displaying variation in pollen shape (second column, B1b–B6b) and exine sculpture (third column, B1c–B6c)

LM (first column, B7a–B12a) and SEM micrographs of pollen grains in different R. webbiana accessions displaying variation in pollen shape (second column, B7b–B12b) and exine sculpture (third column, B7c–B12c)

LM (First column) and SEM micrographs of pollen grains showing variation in pollen shape (second column) and exine sculpture (third column) in different Rosa species. R. canina (C1a–c), R. foetida var. persiana (D1a–c), R. multiflora (E1a–c and E2a–c), R. centifolia (G1a–c), R. damascena (H1a–c), R. nanothamnus (I1a–c)

LM (F1a–F7a) and SEM micrographs of pollen grains in different R. macrophylla accessions displaying variation in pollen shape (F1b–F7b) and exine sculpture (F1c–F7c)
General pollen morphological description
A total of 69.25% of the pollen grains of the studied Rosa species were of medium size and 30.75% of small size. The average length of the polar axis (P) and equatorial axis (E) in the studied species were 26.97 µm (19.26–42.39 µm) and 19.78 µm (11.59–36.90 µm), respectively. As far as the outline of the pollen grains were concerned, almost all had circular (in polar view) to elliptic outline (in equatorial view) (Figs. 2, 3, 4, 5 and 6). The mean P/E ratio of the pollen grains for all the studies species was 1.37 (0.95–2.09) (Table 3). The percentage of pollen shapes in the studied Rosa species in their decreasing order are prolate (56.88%), subprolate (40.65%), prolate spheroidal (2.04%), oblate spheroidal (0.22%) and perprolate (0.22%). The average distance between the apices of the two colpi (d) and polar area index (PAI) was 3.83 µm (2.08–7.51 µm) and 0.22 µm (0.08–0.50 µm), respectively (Table 3).
All the pollen grains possessed three colpi. The average colpi length was 22.14 µm, which ranged between 13.42 µm and 34.03 µm in R. macrophylla and R. canina, respectively (Table 3). The average exine thickness was 1.37 µm (0.68–1.95 µm). Among all the studied Rosa species, the lowest and highest coefficient of variation was recorded for the P/E ratio (2.11%) of R. foetida var. persiana, and groove width of R. webbiana (36.82%), respectively (Table 3).
The mean striae width ranged between 0.12 µm in R. canina and R. centifolia, and 0.22 µm in R. foetida var. persiana. On the other hand, the highest mean groove width was reported in R. foetida var. persiana (0.36 µm) and smallest in R. damascena (0.09 µm). Striae number per 25 µm2 was in the range of 10–27, with an average of 17. The grooves on the exine surface were prominent in most of the cases, with a mean width of 0.17 µm (0.06–0.47 µm). Exine ornamentation showed considerable variation at the species level (Figs. 2, 3, 4, 5 and 6). The arrangement of striae in the majority of the studied species was parallel to the colpi except in accession B–5 of R. webbiana. In this accession, striae did not follow the continuous parallel path and assumed a loop-shaped pattern in the upper region when viewed from the equator (Fig. 3).
Interspecific variability of pollen grains
All the analyzed characters of pollen grains of the studied Rosa species portrayed statistically significant differences (p < 0.001; Table 3). The quantitative details of all the pollen characters are given in Table 3. The maximum and minimum average polar axis length was observed for R. canina (32.73 µm) and R. moschata (25.42 µm), respectively. Whereas, equatorial axis length ranged between 18.51 µm and 24.23 µm for R. webbiana and R. canina, respectively. P/E ratio was highest (1.43) for R. nanothamnus and lowest (1.29) for R. centifolia. In all the studied thirty-one accessions belonging to the nine Rosa species, five types of pollen shapes were recorded.
For R. moschata accessions, four types of pollen shapes were documented viz., oblate spheroidal, subprolate, prolate and prolate spheroidal. Of these, oblate spheroidal (6.7%) and prolate spheroidal (36.7%) shaped pollen grains were present only in accession A-2. Also, prolate shaped pollen grains were absent in accession A-2. The majority of the pollen grains were of sub-prolate shape in A-3 (86.6%) and A-5 (76.6%), whereas most of the pollen grains were prolate shaped in A-1 (73.3%) and A-4 (80.0%).
In the studied accessions of R. webbiana, the four pollen grain shapes were observed viz., subprolate, perprolate, prolate, and prolate spheroidal shapes. Predominantly prolate shaped pollen grains were found in B-1 (83.3%), B-3 (93.3%), B-6 (90.0%), B-8 (86.7%), B-9 (90.0%), and B-10 (86.7%). Accession B-6 was the only accession of R. webbiana having perprolate shaped pollen grains (3.3%). Most of the pollen grains in accessions B-2 (60.0%), B-4 (93.3%), B-5 (80.0%), and B-7 (70.0%) were sub-prolate in shape. Prolate spheroidal shaped pollen grains were observed only in B-4 (3.3%) and B-12 (10.0%) accessions. Furthermore, the percentage of prolate (46.7%) and subprolate (43.3%) pollen grains shapes were comparable in the B-12 accession of R. webbiana.
In the accession of R. canina (C-1), R. foetida var. persiana (D-1), R. multiflora (E-1 and E-2), R. damascena (H-1) and R. nanothamnus (I-1) only sub-prolate and prolate shaped pollen grains were found, with prolate pollen shape being the dominant one in all except E-1 accession of R. multiflora, in which majority of the pollen grains (63.3%) were of sub-prolate shape.
In R. macrophylla accessions, four types of pollen grain shapes were found viz., subprolate, per-prolate, prolate and prolate spheroidal. In accession F-1, two pollen grain shapes were observed i.e., sub-prolate (90.0%) and perprolate (10.0%), while in accession F-2, prolate (70.0%) and sub-sub-prolate (30.0%) pollen grains were recorded. Similarly, the pollen grains of F-3 accession were prolate (33.3%) and sub-sub-prolate (66.7%). The only accession of R. macrophylla having three pollen grains shapes was F-4 having prolate (93.3%), sub-prolate (3.3%), and perprolate (3.3%) shapes. Most of the pollen grains were prolate shaped in F-5 (76.7%), followed by sub-prolate shape (23.3%), while the pollen grains in F-6 accession were predominantly sub-prolate (90.0%). The shapes of pollen grains in F-7 accession were also sub-prolate (13.3%) and prolate (86.7%).
In R. canina, the continuity of striae was often broken after certain intervals, whereas in the case of R. centifolia the striae were almost indistinguishable, and a unique pattern of granule like structures was noticed (Fig. 5). The striae of R. foetida were wave-like, which appeared at some places and disappeared after certain intervals (Fig. 5).
The average colpi length was lowest in R. webbiana (20.87 µm) and highest in R. canina (25.67 µm). In pollen grains of R. centifolia, the striae on exine surface were not prominent, and granules like structures were observed, which is a unique character for this species. The average number of striae per 25 µm2 area ranged between 12.23 in R. centifolia and 24.33 in R. canina. Almost similar numbers of striae per unit area were observed in R. foetida var. persiana, R. macrophylla, R. webbiana, and R. nanothamnus. Whereas, the number of striae and groove width was comparable in R. moschata and R. damascena (Table 3). The minimum coefficient of variation (CV = 10.18%) was obtained for the P/E ratio in the case of R. moschata and a maximum (CV = 40.41%) for groove width in the case of R. nanothamnus. All the details related to the average values of d, PAI, LeC, Sr, Ns, and Gr are given in Table 3. The pollen grains of R. canina, R. damascena, and R. centifolia were found to be of medium size. The coefficient of variation portrayed a broad range of variation in different pollen characters. The coefficient of variation of 7.98% and 11.29% was recorded for the pollen characters polar axis and equatorial axis, respectively. Statistically significant differences were reported in the mean values of P, E and Ns of R. canina as compared to the rest of the studied species, whereas the mean values of Ns in case of R. centifolia and R. damascena differ in a statistically significant manner from the rest of the species as well as from each other.
Results of principal component analysis (PCA) indicated that the first four principal components (PCs) with Eigenvalues higher than 1 encompassed 80.4% of the total variation (Fig. 7). Principal component first (PC1) attributed 42.1% of the total variability with P, E, LeC, Ns being the significant variables, whereas second component (PC2) covered 19.7% of the total variation and having P/E, Ex, Sr Gr as the significant variables. PC3 component encompassed 18.7% of the total variation and related to the pollen characters such as PAI and d. Three species, namely R. canina, R. webbiana, and R. foetida var. persiana, are distantly positioned from each other, affirming significant dissimilarity amongst them in PCA.

PCA of the first two components explaining 61.8% of the total variation using ten quantitative pollen characters
Using the UPGMA agglomeration method, a dendrogram was generated on a quantitative pollen character basis for the clustering of the species (Fig. 8). Two clusters were retrieved with a major cluster possessing all the Rosa species except R. canina. The main cluster was further divided into two groups with R. damascena and R. centifolia forming one group, and R. foetida var. persiana, R. centifolia, R. multiflora, R. nanothamnus, R. macrophylla, and R. webbiana forming another group.

Cluster groupings of 9 Rosa species based on quantitative morphological characters of pollen grains
Intraspecific variability of pollen grains
Rosa webbiana and R. macrophylla are the two most dominant wild Rosa species in the Western Himalaya. These two species are found spread across different ecological zones along the altitudinal gradient in the study area (Table 1). Keeping that in view, higher numbers of accessions of R. webbiana and R. macrophylla were evaluated in the present study. These two species were also considered for assessing intraspecific level diversity (Tables 4 and 5). The differences observed among the accessions collected from various localities with respect to all the pollen characters were statistically significant (p < 0.001).
The intraspecific variability of these two species revealed that the accession of R. webbiana collected from Nyoma locality showed the highest mean polar axis length (29.51 µm) while the accession collected from Dawar locality possessed lowest average polar axis length (22.16 µm). The number of striae per 25 µm2 ranged between 13.47–21.30 in R. webbiana accessions collected from Nyoma and Chutak locality, respectively (Table 4). Perforation on the exine surface was absent in the accession collected from BudhKahrbu. Rosa webbiana accession collected from Wassar locality had the highest prolate shaped pollen grains (93.33%), and Dawar accession had the lowest (3.33%). Pollen grains collected from Wassar, Dawar, Kundal, and Khardungla localities were small-sized (< 0.25 µm), while the rest were medium-sized (25.1–50.0 µm).
In the case of R. macrophylla, the highest polar axis length was documented for accession collected from Machail (32.28 µm) and lowest for Frislan (24.34 µm). 93.33% of prolate shaped pollen grains were recorded for accession from Machail locality while accession from Chorwan locality had no prolate pollens (Table 5). Accessions from Chattergala and Chorwan possessed 90% of subprolate shaped pollen. Exine perforation was absent in accessions collected from Chorwan, Khajjar, Machail, and Karit locality, while present in the rest of the accessions.
In some cases, intraspecific variation in striae and groove width was also observed. For example, R. webbiana accession of Kundal locality showed resemblance with the exine of R. nanothamnus. The exine surface in the case of R. macrophylla accession of Karit locality showed undulations, which are absent in other R. macrophylla accessions (Fig. 6). The perforations were present in the majority of the studied accessions of the Rosa species except for some accessions of R. webbiana (B-7), R. multiflora (E-2), R. macrophylla (F-5 and F-7), R. centifolia (G-1) and R. canina (C-1).
Discussion
Internationally few studies have been carried out on some of the Rosa species evaluated in the present study (Reitsma 1966; Eide 1981; Ueda and Tomita 1989; Ueda 1992; Wronska-Pilarek and Jagodzinski 2011; Wronska-Pilarek 2011; Fatemi et al. 2012; Wronska-Pilarek et al. 2015). However, the present is the first palynological study carried out on the Rosa species from the Western Himalayan Region of India. The present article has reported pollen grains characters of R. centifolia for the first time.
Earlier studies on pollen grains of family Rosaceae and Rosa genus suggested the use of exine structure as a primary diagnostic character (Eide 1981; Marcucci et al. 1984; Hebda et al. 1988; Ueda and Tomita 1989; Hebda and Chinnappa 1990; Popek 1996; Shinwari and Khan 2004), which is in agreement with our results. In the present study, it was observed that an exine structure could be used to separate R. canina from the rest of the species (Fig. 7). The size of grooves can be used as a diagnostic character for the separation of R. foetida var. persiana from the rest of the studied species. Similarly, in the case of genus Spiraea L. of family Rosaceae, Polyakovaa and Gataulina (2008) had earlier shown the importance of the size of grooves along with features of exine sculpture in species discrimination. Striae width and striae numbers were found useful in the delimitation of the studied species, which is in line with the observations of some of the earlier studies (Fogle 1977; Matsuta et al. 1982; Ueda and Okada 1994). In one more previous study, Menge (1985) found that perforations on the exine surface may help distinguish species. Similarly, in the present study, it was observed that perforations on the exine surface might help to segregate some of the studied species (Figs. 2, 3, 4, 5 and 6). However, in the case of species like R. webbiana, and R. macrophylla, this character may not be useful due to inconsistency in the presence of perforations in pollen grains of their accessions.
Some earlier researchers observed that pollen characters such as polar and equatorial axis, pollen shape, colpi length might be used for the Rosa species delineation (Eide 1981; Hebda and Chinnappa 1990; Shinwari and Khan 2004). However, our results suggest that this criterion cannot be universally applied for all the Rosa species. For example, in the case of R. damascena and R. multiflora, two pollen shape types were observed, whereas, in the case of R. webbiana and R. macrophylla, four pollen shape types were observed. The coefficients of variation for polar and equatorial axis obtained in the present study were found well within the range of the earlier reported values for other Rosa species (Wronska-Pilarek and Jagodzinski 2009). Similarly, polar axis length in R. damascena, R. centifolia, R. multiflora, and R. nanothamnus were comparable, while the colpi length was almost equal in R. macrophylla and R. damascena. In the same way, almost the same colpi length was recorded in R. centifolia, R. multiflora, R. nanothamnus, and R. foetida var. persiana.
The clustering analysis grouped some Rosa species belonging to different sections into a single clade (Fig. 8). Interestingly, R. foetida var. persiana of section Pimpenillifoliae, R. multiflora of section Synstylae, and R. nanothamnus of section Cinnamommae were grouped. However, in a recent study based on molecular data, Fougere-Danezan et al. (2015) had proved that they are not monophyletic sections. The possible reason for this ambiguous grouping pattern obtained in the present study could be polyploidy in most of the studied species. Jacob and Pierret (2000) observed a positive correlation between chromosome number and pollen size, particularly in diploid and tetraploid roses. The results of the present study are in partial agreement with the morphological studies carried out by Cheikh-Affene et al. (2015) on wild rose population in Tunisia in which sectional level, as well as species-level delineation, was attained. In the present study, species were discriminated, but species from separate sections were grouped into one cluster (Fig. 8). A recent study by Singh et al. (2020) on Western Himalayan wild roses based on 59 morphological characters supports the results of the current study to some extent in a manner that species were delineated.
RAPD and SSR markers have been stated to be useful in identifying the hybrids in scented roses (Kaul et al. 2009). A recent study carried out on 29 rose cultivars accessions by Agarwal et al. (2019) using 32 primers found SCoT markers very useful in fingerprinting as well as diversity analysis in roses. Kiani et al. (2010) found a high level of genetic variability in R. damascena accessions using SSR and RAPD. Also, they obtained a high correlation range between SSR, RAPD, and morphological similarity matrices. Samiei et al. (2010) showed the consistent nature of the phenetic tree with that of the traditional classification using SSR markers at the sectional level, which is not in agreement with our study. Since the grouping in the present study is entirely based on pollen characters alone, it is not as per the results of the studies done earlier based on the morphological (Henker 2000) and molecular characters (Fougere-Danezan et al. 2015). However, it will give us a general idea about the possible use of pollen characters in distinguishing the studied Rosa species from each other. Based on the quantitative characters, both PCA and clustering analysis confirmed the separation of studied Rosa species.
Conclusions
Based on the results obtained in the present study, the authors found that the pollen grain characters such as length of polar and equatorial axis, P/E ratio, exine thickness, number of striae, the width of grooves and exine surface ornamentation may be useful in species discrimination. However, at an intraspecific level, the perforation on exine and arrangement of striae did not show consistency in R. webbiana and R. macrophylla; thus, these characters may not be useful in discriminating these two species.
References
Agarwal A, Gupta V, Haq SU, Jatav PK, Kothari SL, Kachhwaha S (2019) Assessment of genetic diversity in 29 rose germplasms using SCoT marker. J King Saud Univ Sci 31:780–788
Almeida GS, Mezzonato-Pires AC, Mendonca CBF, Goncalves-Esteves V (2018) Pollen morphology of selected species of Piriqueta Aubl. (Passifloraceae sensu lato). Palynology 43:43–52
Ambasta SP (1986) The useful plants of India. In: CSIR New Delhi, publication and information directorate, India
Andrews HC (1805) Roses, or, a monograph of the genus Rosa: containing colored figures of all known species and beautiful varieties. Taylor R. and Co., London
Bai JR, Zhang QX, Luo L, Pan HT, Yu C (2011) Pollen morphology of some Chinese traditional roses. Bull Bot Res 31:15–23
Baker JG (1869) A monograph of the British roses. Bot J Linn Soc 11:197–243
Bashir KMI, Awan FS, Khan IA, Khan AI, Usman M (2014) Identification and authentication of Rosa species through development of species-specific SCAR marker(s). Genet Mol Res 13:4130–4139
Besse C, Cao MQ, Gandelin MH, Zhang D (2000) Evaluation of AFLPs for variety identification in modern rose (Rosa hybrida L.). Int Symp Mol Markers Charact Genotypes Identif Cultiv Hortic 546:351–357
Broertjes C, van Harten AM (1978) Application of mutation breeding methods in the improvement of vegetatively propagated crops. Elsevier, Amsterdam
Cheikh-Affene ZB, Haouala F, Harzallah-Skhiri F (2015) Morphometric variation and taxonomic identification of thirteen wild rose populations from Tunisia. Acta Bot Croat 74:1–7
Chopra VL, Singh M (2013) Ornamental plants for gardening. Scientific Publishers, Jodhpur
Chowdhery HJ, Wadhwa BM (1984) Flora of Himachal Pradesh, vol 1. Botanical Survey of India Publication, Delhi
Debener T, Bartels C, Mattiesch L (1996) RAPD analysis of genetic variation between a group of rose cultivars and selected wild rose species. Mol Breed 2:321–327
Devi U, Seth MK, Sharma P, Rana JC (2013) Study on ethnomedicinal plants of Kibber Wildlife Sanctuary: a cold desert in trans Himalaya, India. J Med Plant Res 7:3400–3419
Dhyani D, Singh S (2014) Potential wild rose germplasm of western Himalayas-conservation, evaluation, and registration. Indian J Agric Sci 84:229–235
Duthie JF (1971) Flora of the Upper gangetic plain and of the adjacent Siwalik and sub-Himalayan tracts. International Book Distributors, Dehradun
eFloras (2019) Published on the Internet. Missouri Botanical Garden, St. Louis, MO and Harvard University Herbaria, Cambridge, MA. https://www.efloras.org. Accessed 02 December 2019
Eide F (1981) Key for northwest European Rosaceae pollen. Grana 20:101–118
Erdtman G (1952) Pollen morphology and plant taxonomy of angiosperms. Almqvist and Wiksell, Stockholm
Esselink GD, Smulders MJ, Vosman B (2003) Identification of cut rose (Rosa hybrida) and rootstock varieties using robust sequence tagged microsatellite site markers. Theor Appl Genet 106:277–286
Fatemi N, Attar F, Assareh MH, Hamzehee B (2012) Pollen morphology of genus Rosa L. (Rosaceae) in Iran. Iran J Bot 18:284–293
Fogle HW (1977) Identification of clones within four tree fruit species by pollen exine pattern. J Am Soc Hortic Sci 102:552–560
Fougere-Danezan M, Joly S, Bruneau A, Gao XF, Zhang LB (2015) Phylogeny and biogeography of wild roses with specific attention to polyploids. Ann Bot 115:275–291
Ghosh A, Saha I (2017) Pollen morphological study of some selected Indian taxa of Rosaceae. Indian J Pure Appl Biol 32:121–130
Gu C, Robertson KR (2003) Rosa L. In: Team FoCe (eds) Flora of China, Missouri Botanical Garden Press, St. Louis, MO
Halbritter H, Ulrich S, Grimsson F, Weber M, Zetter R, Hesse M, Buchner R, Svojtka M, Frosch-Radivo A (2018) Illustrated pollen terminology. Springer, Berlin
Hammer O, Harper DAT, Ryan PD (2001) PAST: Paleontological statistics software package for education and data analysis. Palaeontol Electron 4:9
Hebda RJ, Chinnappa CC (1990) Studies on pollen morphology of Rosaceae in Canada. Rev Palaeobot Palynol 64:103–108
Hebda RJ, Chinnappa CC (1994) Studies on pollen morphology of Rosaceae. Acta Bot Gall 141:183–193
Hebda RJ, Chinnappa CC, Smith BM (1988) Pollen morphology of the Rosaceae of Western Canada I. Agrimonia to Crataegus Grana 27:93–113
Henker H (2000) Rosa. In: Hegi G (ed) Illustrierte Flora von Mitteleuropa. Band 4/2c. Parey Buchverlag, Berlin, pp 1–108
Hesse M, Halbritter H, Zetter R, Weber M, Buchner R, Frosch-Radivo A, Ulrich S (2009) Pollen terminology: an illustrated handbook. Springer, Vienna
Hooker JD (1879) Flora of British India. Reeve & Co., London
Hutchinson J (1964) The genera of flowering plants. Oxford University Press, London
Jacob Y, Pierret V (2000) Pollen size and ploidy level in the genus Rosa. Acta Hortic 508:289–292
Kachroo P, Bansilal S, Dhar U (1977) Flora of Ladakh: an ecological and taxonomical appraisal. International Book Distributors, Dehradun
Kalkman C (2004) Rosaceae. In: Kubitzki K (ed) The families and genera of vascular plants VI Flowering plants—dicotyledons. Springer, Berlin, pp 386–443
Kaul K, Karthigeyan S, Dhyani D, Kaur N, Sharma RK, Ahuja PS (2009) Morphological and molecular analyses of Rosa damascena× R. bourboniana interspecific hybrids. Sci Hortic 122:258–263
Kaul VK, Gujral RK, Singh B (1999) Volatile constituents of the essential oil of flowers of Rosa brunonii Lindl. Flavour Frag J 14:9–11
Kiani M, Zamani Z, Khalighi A, Fatahi R, Byrne DH (2010) Microsatellite analysis of Iranian Damask rose (Rosa damascena Mill.) germplasm. Plant Breed 129:551–557
Koopman WJM, Wissemann V, de Cock K, Huylenbroeck JV, de Riek J, Sabatino GJH, Visser D, Vosman B, Ritz CM, Maes B, Werlemark G, Nybom H, Debener T, Linde M, Smulders MJM (2008) AFLP markers as a tool to reconstruct complex relationships: a case study in Rosa (Rosaceae). Am J Bot 95:353–366
Lewis WH (1957) A monograph of the genus Rosa in North America east of the Rocky Mountains. Ph.D. Thesis submitted to University of Virginia, Virginia
Marcucci MC, Sansavini S, Ciampolini F, Cresti M (1984) Distinguishing apple clones and cultivars by surface morphology and pollen physiology. J Am Soc Hortic Sci 109:10–19
Matsuta N, Omura M, Akihama T (1982) Difference in micromorphological pattern on pollen surface of Japanese Pear cultivars. Jpn J Breer 32:123–128
Menge U (1985) Identification of rose varieties using pollen surface patterns/identification of Rose cultivars by patterns of pollen grain surfaces. Hort Sci 501:1–9
Mohapatra A, Rout GR (2006) Optimization of primer screening for evaluation of genetic relationship in rose cultivars. Biol Plantarum 50:295–299
Nadeem M, Wang X, Akond M, Awan FS, Riaz A, Younis A (2014) Hybrid identification, morphological evaluation and genetic diversity analysis of Rosa hybrida by SSR markers. Aust J Crop Sci 8:183
Nand K, Naithani S (2018) Ethnobotanical uses of wild medicinal plants by the local community in the Asi Ganga sub-basin, Western Himalaya. J Complement Med Res 9:34–46
Nilsson O (1997) Rosa. In: Davis PH (ed) Flora of Turkey and the East Aegean Islands. Edinburgh University Press, Edinburgh, pp 106–128
Pal BP (1991) The rose in India. Publication and Information Division, ICAR, New Delhi
Panwar S, Singh KP, Sonah H, Deshmukh RK, Prasad KV, Sharma TR (2015) Molecular fingerprinting and assessment of genetic diversity in rose (Rosa × hybrida). Indian J Biotechnol 14:518–524
Polyakovaa TA, Gataulina GN (2008) Morphology and variability of pollen of the genus Spiraea L. (Rosaceae) in Siberia and the far east. Contemp Prob Ecol 1:420–424
Popek R (1996) Biosystematic studies on the type of Rosa L. in Poland and neighboring countries. In: Scientific Publisher of the College of Pedagogy, Krakow. Prace Monograficzne, vol 218
Punt W, Hoen PP, Blackmore S, Nilsson S, Le Thomas A (2007) Glossary of pollen and spore terminology. Rev Palaeobot Palyno 143:1–81
Rao RR, Sharma BD (1990) A manual for herbarium collections. Botanical Survey of India, Kolkata
Reitsma TJ (1966) Pollen morphology of some European rosaceae. Acta Bot Neerl 15:290–307
Riaz A, Hameed M, Khan AI, Younis A, Awan FS (2011) Assessment of biodiversity based on morphological characteristics and RAPD markers among genotypes of wild rose species. Afr J Biotechnol 10:12520–21252
Samiei L, Naderi R, Khalighi A, Shahnejat-Bushehri AA, Mozaffarian V, Esselink GD, Osaloo K, Smulders MJ (2010) Genetic diversity and genetic similarities between Iranian rose species. J Hortic Sci Biotech 85:231–237
Schori M, Fumess CA (2014) Pollen diversity in Aquifoliales. Bot J Linn Soc 175:169–190
Shameh S, Alirezalu A, Hosseini B, Maleki R (2019) Fruit phytochemical composition and color parameters of 21 accessions of five Rosa species grown in North West Iran. J Sci Food Agric. https://doi.org/10.1002/jsfa.9842
Sharma BM, Kachroo P (1981) Flora of Jammu and Plants of neighborhood, vol 1. Bishen Singh Mahendra Pal Singh, Dehradun
Shinwari MI, Khan MA (2004) Pollen morphology of wild roses from Pakistan. Hamdard Med 47:5–13
Singh G, Kachroo P (1976) Forest flora of Srinagar and plants of neighborhood. Bishen Singh Mahendra Pal Singh, Dehradun
Singh K, Sharma YP, Gairola S (2020) Morphological characterization of wild Rosa L. germplasm from the Western Himalaya, India. Euphytica 216:41
Song JH, Oak MK, Roh HS, Hong SP (2017) Morphology of pollen and orbicules in the tribe Spiraeeae (Rosaceae) and its systematic implications. Grana 56:351–367
TPL (2019) The plant list, version 1.1. https://www.theplantlist.org. Accessed 02 December 2019
Ueda Y (1992) pollen surface morphology in genus Rosa and related genera. Jpn J Palynol 38:94–105
Ueda Y, Okada Y (1994) Discrimination of rose cultivar groups by pollen surface structure. J Jpn Soc Hortic Sci 694:601–607
Ueda Y, Tomita H (1989) Morphometric analysis of pollen exine pattern in Roses. J Jpn Soc Hortic Sci 58:211–220
Uggla M, Gao X, Werlemark G (2003) Variation Among and Within Dogrose Taxa (Rosa sect. caninae) in fruit weight, percentages of fruit flesh and dry matter, and vitamin C content. Acta Agric Scand Sect B Plant Soil Sci 53:147–155
Wissemann V (2003) Conventional taxonomy of wild roses. In: Roberts A, Debener T, Gudin S (eds) Encyclopedia of rose sciences. Elsevier Science, Oxford, pp 111–117
Wronska-Pilarek D (2011) Pollen morphology of Polish native species of the Rosa genus (Rosaceae) and its relation to systematics. Acta Soc Bot Pol 80:221–232
Wronska-Pilarek D, Jagodzinski AM (2009) Pollen morphology variability of Polish native species of Rosa L. (Rosaceae). Dendrobiology 62:71–82
Wronska-Pilarek D, Jagodzinski AM (2011) Systematic importance of pollen morphological features of selected species from the genus Rosa (Rosaceae). Plant Syst Evol 295:55–72
Wronska-Pilarek D, Jagodzinski AM, Bocianowski J, Janyszek M (2015) The optimal sample size in pollen morphological studies using the example of Rosa canina L. (Rosaceae). Palynology 39:56–75
Acknowledgements
Authors thank Director, IIIM Jammu, for providing necessary facilities to carry out the work. Authors are thankful to SERB DST for financial assistance under Start-Up Research Grant (Young Scientist), Grant No YSS/2015/000442. KS acknowledges the financial support provided by CSIR in the form of a JRF/SRF fellowship. CSIR-IIIM Publication No. CSIR-IIIM/IPR/00178.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Singh, K., Sharma, Y.P., Sharma, P.R. et al. Pollen morphology and variability of the Rosa L. species of Western Himalaya in India. Genet Resour Crop Evol 67, 2129–2148 (2020). https://doi.org/10.1007/s10722-020-00967-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-020-00967-8