
Abstract
Foxtail millet [Setaria italica (L.) P. Beauv.] is among the oldest cereal grains grown from time immemorial in the Himalayan regions of Nepal. However, Nepalese farmers do not have any improved variety officially released due to lack of substantial research in this crop. A total of 41 foxtail millet accessions were characterized using phenotypic or morphological markers at National Agriculture Genetic Resources Centre (Genebank), Khumaltar (1360 m a.s.l.), Lalitpur, Nepal during 2015 summer to enhance the utilization of foxtail millet genetic resources. Seven quantitative and nine qualitative traits were recorded using standard descriptors of foxtail millet to assess the intra-specific diversity. Significant diversity was observed among the accessions as revealed by Shannon–Weaver diversity indices (H′) for quantitative traits and qualitative traits. Six elite landraces in Cluster-4 and Cluster-5 (three from Lamjung district, and one each from Gorkha, Humla and Jumla districts) produced average grain yield of 3136 kg/ha at Khumaltar condition with earlier maturity (average of 89 days), taller height (average of 172 cm) and thicker panicles (average width of 27 mm). These landraces were selected for further evaluation in the farmers’ field of mountain region due to their early maturity, higher yield potential, disease resistance and attractive panicles which could be considered as important genetic resources to develop climate resilient varieties to cope with the adverse effects of climate change.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Foxtail millet [Setaria italica (L.) P. Beauv.], regarded as a native of China, is one of the world’s oldest cultivated crops. It was originated in China, where it’s domestication took place about 8000 YBP (Li and Wu 1996) as well as in south west Asia and central Europe (Vavilov 1926; Zohary et al. 2012) where it’s cultivation recorded about 5000 YBP (Hammer et al. 1999). The Hindukush Himalayan region is rich in foxtail millet diversity (Scheibe 1943; Hammer and Khosbhakht 2007). The crop has moved from China to Northern India and Nepal during 5th to 2nd Millennium BC (Stevens et al. 2016), thus the Nepalese foxtail millet, mostly is Setaria italica subsp. italica race maxima (de Wet et al. 1979). It is the second most widely planted species among millets in the world and the most important millet in East Asia (Kumari et al. 2011; Ning et al. 2015; Sheikh and Singh 2013; Xiaomei et al. 2016; Zhang et al. 2014). It is known for its better tolerance to abiotic stresses compared to other cereal crops. It is also known as Italian millet, German millet or Hay millet but locally known as Kaguno in Nepal. Foxtail millet grain contains 12.3% protein, 4.3% fat, 60.9% carbohydrates, 14.0% dietary fibre and 3.3% minerals, with 31 g calcium, 290 mg phosphorus, 5 mg iron and vitamins than major staple crops rice and wheat (Saha et al. 2016; Saud 2010). Foxtail millet is the third important crop among group of millets with wide range of utility in Nepal. The cooked grain is used as bhat (cooked like rice), dhindo (porridge) and kheer (like rice pudding). Foxtail millet is valued by mountain farmers for its nutritional content and health promoting properties, ability to grow under low external input conditions and tolerance to extreme environmental stress, particularly drought. It is also recently appreciated because of medicinal benefits such as reducing blood glucose levels and cholesterol control in normal as well as diabetic patients. It is the crop of future in the context of changing climate with great potentiality to cope with food insecurity in remote areas of the country (Goron and Raizada 2015).
In Nepal, this crop is considered as traditional climate resilient and nutritionally dense crop but trends of cultivation and use are shrinking fast due to globalization, land use change, out migration, social values, change in food habit, depleting traditional knowledge, lack of research and policy support such as crop improvement and formal seed distribution system (Gurung et al. 2016; Parajuli et al. 2017; Palikhey et al. 2016; Sheikh and Singh 2013; Bisht et al. 2006). Precise data of area and production under foxtail millet is not known because the production statistics of this crop had often been grouped with other millets. Major foxtail millet growing districts in Nepal are Mugu, Kalikot, Humla, Jumla, Bajhang, Bajura, Dolpa, Lamjung, Gorkha, Ramechhap, Kavre, etc. where crop is grown sole as well as mixed with finger millet, proso millet, beans, amaranths, maize etc. Nepal is one of the centres of diversity of foxtail millet (Nakayama et al. 1999) and has high genetic diversity of this crop (Mo FSC 2002), however, very little research has been conducted and its status is still unexplored in Nepal thus called neglected and underutilized crop (Amgai et al. 2011). Plant genetic resources are conserved so that they can be used to improve crop production and in other ways. However, Hodgkin et al. (2003) asserted that use of ex situ conserved germplasm is inadequate globally and that genetic diversity maintained in genebanks is underutilized.
Poor utilization of local genetic resources conserved in genebank for foxtail millet improvement and development program is evident in Nepal due to (1) absence of public and private sector breeding priority, and (2) unavailability of characterization and evaluation information for wider use. GEF-supported Local Crop Project (www.himalayancrops.org), executed jointly by Nepal Agricultural Research Council, Department of Agriculture, LI-BIRD and Bioversity International, therefore aims to fill this gap and re-introduce genebank materials of foxtail millet landraces in traditional farming system for improving farmers access to diversity and increasing on-farm diversity. The objective of this study is to characterize diverse farmer’s varieties of foxtail millet using agro-morphological data and use information to deploy useful diversity by participatory methods, and also for the efficient use of these accessions in future crop improvement programme.
Materials and methods
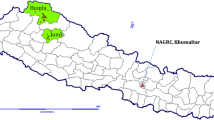
A total of 41 foxtail millet accessions were collected from six districts of Nepal (Fig. 1). Among them, 28 accessions were from Humla through diversity fair, six accessions were from Jumla, four accessions were from Lamjung and one accession each from Bajhang, Gorkha and Kavre districts. The detailed sources of accessions are presented in Table 1. These accessions were characterized ex situ at Genebank, Khumaltar (N27.4°, E085.2°, 1360 m a.s.l.), Nepal during summer 2015. Accessions were planted on 17th May, 2015 with an individual plot size of 6 m2 following row to row spacing of 25 cm and plant to plant spacing of 10 cm. Fertilizers were applied at the rate of 20:30:20 kg/ha N:P2O5:K2O as basal dose during land preparation and 20 kg/ha N was top-dressed 25 days after seeding. Different quantitative traits like days to 50% heading, days to 80% maturity, plant height (cm), panicle length (cm), panicle width (mm), panicle exertion (cm) and grain yield (kg/ha) as well as qualitative traits like growth habit, flag leaf angle, sheath pubescence, leaf pubescence, grain colour, grain yield potential, leaf blast susceptibility, stem borer susceptibility and overall phenotypic acceptability were recorded as per descriptors for Setaria italica (L.) P. Beauv. and S. pumila (Poir.) Roem. et Schult. (IBPGR 1985). Observations on days to heading, maturity and grain yield were based on whole plot data whereas observations on rest of the traits were from 10 randomly selected plants.

Collection map of experimental materials and characterization location
Shannon–Weaver diversity indices (Shannon and Weaver 1949) were calculated for each trait with Microsoft Excel using the formula: \(H^{\prime} = \left[ {\sum \left( {{n}/{N}} \right)^*\left\{ {log2\left( {{n}/{N}} \right)^*\left( { - 1} \right)} \right\}} \right]/log2k\); where, H′ is the standardized Shannon–Weaver diversity index, k is the number of phenotypic classes for a character, n is the frequency of a phenotypic class of that character and N is the total number of observations for that character. For the quantitative traits, accessions were divided into 10 phenotypic classes as < x − 2sd, x − 2sd, x − 1.5sd, x − sd, x − 0.5sd, x, x + 0.5sd, x + sd, x + 1.5sd, x + 2sd and > x+2sd are as the margins of the classes, where x is average and sd is standard deviation. The descriptive statistics, clustering, Principal Component Analysis (PCA) and correlation analysis is done by using Minitab-14.
Results and discussion
Descriptive statistics and Shannon–Weaver diversity
Nepalese foxtail millet germplasm found diverse in shape, size and colour of panicles as well as grains which has been illustrated in Fig. 2.

Diversity in panicle and grain types of Nepalese foxtail millet
Range and average of observations as well as Shannon–Weaver diversity indices for different quantitative traits has been presented in Table 2. Flowering of accessions ranged from 45 to 71 days with the mean of 60 days from seeding. This range was wider than the flowering range of Nepalese accessions (45–60 days) but much narrower than the global collections (32–135 days) in ICRISAT, India (545 m) reported by Reddy et al. (2006). According to that report, the earliest maturity accession was from Russia and the longest maturity was from Sri Lanka. This is very important adaptive trait for mountain farmers in rainfed dryland conditions, and also to escape for extreme cold temperature. Women farmers prefer earliness because the food is available during lean period. Plant height ranged from 108 to 232 cm with an average of 159 cm which is quite taller than the Nepalese (70–150 cm) as well as global (20–215 cm) accessions (Reddy et al. 2006). Our Nepalese accessions showed narrower range of panicle exertion (7–30 cm) as compared to the range in global collection of 1–36 cm (Reddy et al. 2006). Panicle length ranged from 13 to 30 cm which is similar to the range in Nepalese collection (13–25 cm) but narrower than the range in Indian collection (2–39) as reported by Reddy et al. (2006). Panicle width in our result showed narrower range (10–33 mm) wider than the range in Nepalese (10–25 mm) and very much narrower than the global (5–120 mm) collection (Reddy et al. 2006). Grain yield in our study ranged from 89 to 3483 kg/ha with the average of 1247 kg/ha. This is the preliminary yield from small plot (6 m2) non-replicated trial in high fertility condition of Khumaltar. Some accessions were severely damaged by leaf blast (Pyricularia setariae Nisikado.) and false smut (Ustilago crameri Korn.) diseases and gave very poor yield.
Shannon–Weaver diversity index (H′) considers both richness and evenness of the phenotypic classes of the traits. H′ for all quantitative traits is ranged from 0.689 to 0.899 (Table 2) that for qualitative traits ranged from 0.443 to 0.989 (Table 3), suggested that characterized 41 foxtail millet accessions were diverse for recorded traits.
Clustering observations
UPGMA clustering divided 41 accessions of foxtail millet into five clusters (Fig. 3). Two clusters, Cluster-1 and Cluster-2, comprised of 15 accessions in each cluster mainly from Humla (Table 4). Cluster-1 has accessions that were low to intermediate yielder, medium maturity and height. Accessions from Cluster-2 were very low yielder (89–500 kg/ha), shortest panicle width (11–19 mm) and mainly late in maturity (81–113 days). Cluster-4 and Cluster-5 jointly comprised of 6 accessions (3 from Lamjung, 1 each from Gorkha, Humla and Jumla districts) were of higher grain yield (2767–3473 kg/ha), taller plants (158–194 cm), greater panicle width (19–33 mm) and early maturity (86–99 days). Accessions from these two clusters can be further evaluated on-farm for their validation.

UPGMA clustering of 41 foxtail millet landraces based on Euclidean distance
Principal component analysis
Table 5 shows the principal component analysis with eigen vectors, eigen values and variances due to first five principal components. In the first principal component, grain yield, panicle width and plant height with negative loading of − 0.581, − 0.486 and – 0.435, respectively were the most important traits. Similarly in the second principal component, days to heading with negative loading of − 0.639 and panicle length with positive loading of 0.496 were the most important traits. Panicle exertion was the most important trait in the third principal component followed by days to maturity with positive loadings of 0.871 and 0.374, respectively. Eigen analysis of the correlation matrix revealed that five principal components explained 95% of the total variance among the accessions. The first five principal components accounted for 32, 28, 15, 11 and 9%, respectively of the total phenotypic variance among accessions based on 7 quantitative traits. Scatter plot of the first two principal components, accounting 60% of cumulative variance, suggested that PCA is in support with the result of cluster analysis (Fig. 4).

Scatter plots of foxtail millet landraces based on first two principal components
Correlation analysis
Pearson’s correlation coefficient between quantitative traits has been presented in Table 6. There was highly significant positive correlation between days to heading and days to maturity. Significant but negative correlation was observed between days to heading and panicle length. Similarly days to maturity was negatively correlated with grain yield suggested that early genotypes gave higher yield as an adaptive trait under semi-arid and cold environment, thus early maturing accessions can be selected for dry areas which can escape from terminal drought and cold stresses. Plant height showed highly significant positive correlations with grain yield and panicle width, respectively suggesting that taller height can be considered as a desirable character of high yielding accession. The strong association between grain yield and panicle width suggested that accessions with thicker panicles can be selected as a high yielding trait.
Crop genetic diversity of traditional mountain crops such as foxtail millet has disappeared in many parts of Himalayan mountains (Sheikh and Singh 2013). Community richness of foxtail millet diversity in Lamjung, Humla and Jumla has shown declining trend in recent decades. Recent surveys indicated area under foxtail millet and the number of households growing the crop is shrinking in mountains of Nepal (Gurung et al. 2016; Parajuli et al. 2017; Palikhey et al. 2016). Local crop genetic diversity from these areas exists in National Genebank or in farmers’ fields but they are not characterized and evaluated. They are not easily accessible to majority of farmers. This is not surprising as globally the use of ex situ materials is limited by lack of characterization data and information (Hodgkin et al. 2003) and direct use of genebank materials by re-introduction by farmers may benefits farming community immediately amongst less researched crops. With increased awareness on nutritional and health benefits and climate resilient properties of the crop, foxtail millet research for development in low HDI index districts of Nepal is recently recognized. Farmers of these areas of Nepal suffer from food insecurity, malnutrition and hidden hunger and therefore, importance of traditional crops like foxtail millet remains important. PRA surveys suggest that farmers prefer varieties have set of traits such as higher yield, earliness, easy threshing and taste. Participatory methods proposed by Joshi and Sthapit (1990) and Witcombe et al. (2016) will be employed for further evaluation in diverse geographic environments. The application of this study will be used to distribute a set of IRD kits i.e. half kg seed per HH of six landraces (namely C5644, H252, C5647, C5808, C5643 and C4581) in a range of 500–1000 kits for deploying new diversity and provide farmers for participatory selection for their needs. The pre-breeding or grassroots breeding of the crop will help to provide better options than available in local market (Sthapit and Ramanatha Rao 2009) and the best way in which genebank diversity can be used as a strategy for such nutritionally dense and climate resilient neglected crops, will have considerable positive effect on use of under-utilized ex situ genetic resources (Goron and Raizada 2015).
Conclusion
Among the 41 accessions of foxtail millet collected from six districts of the country, high level of diversity was observed for different agro-morphological markers like growth habit, phenology, plant stature, length and width of inflorescence, flag leaf angle, leaf and sheath pubescence, leaf blast and stem borer susceptibility, grain yield and grain colour, etc. High grain yield was associated with earliness, tallness and thicker panicles. Accessions C5644, H252, C5647, C5808, C5643 and C4581 are the important landraces (three from Lamjung and one each from Gorkha, Humla and Jumla) needs further evaluation in farmers’ field of multiple locations to identify candidate genotype for direct cultivation and also for future use in crop improvement programmes. The project aims to test these materials in large number (500–1000 kits) in wide geographic region of Nepal using participatory diversity kits for deploying new diversity and make traditional seed system more resilient. Farmers now have a greater range of genetic diversity available from genebank to them than ever before and have flexibility for local selection for target environment.
References
Amgai RB, Pantha S, Chhetri TB, Budhathoki SK, Khatiwada SP, Mudwari A (2011) Variation on agro-morphological traits in Nepalese foxtail millet (Setaria italica (L.) P. Beauv.). Agron J Nepal 2:2011
Bisht IS, Rao KS, Bhandari DC, Nautiyal S, Maikhuri RK (2006) A suitable site for in situ (on-farm) management of plant diversity in traditional agroecosystems of western Himalaya in Uttaranchal state: a case study. Genet Resour Crop Evol 53:1333–1350. https://doi.org/10.1007/s1022-005-4432-5
de Wet JME, Oestry-Stidd LL, Cubero JI (1979) Origins and evolution of foxtail millets (Setaria italica). Journ d’Agric Trad et de Bota Appl XXVI(1):53–64
Goron TL, Raizada MN (2015) Genetic diversity and genomic resources available for the small millet crops to accelerate a new green revolution. Rev Front Plant Sci 6:1–17. https://doi.org/10.3389/fpls.2015.00157
Gurung R, Sthapit SR, Gauchan D, Joshi BK, Sthapit BR (2016) Baseline Survey Report: II. Ghanpokhara, Lamjung. Integrating traditional crop genetic diversity into technology: using a biodiversity portfolio approach to buffer against unpredictable environmental change in the Nepal Himalayas. LI-BIRD, NARC and Bioversity International, Pokhara, Nepal
Hammer K, Khosbhakht K (2007) Foxtail millet (Setaria italica (L.) P. Beauv.) in Mazandaran/Northern Iran. Genet Resour Crop Evol 54:907–911. https://doi.org/10.1007/s10722-007-9211-z
Hammer K, Knüpffer H, Laghetti G, Perrino P (1999) Seeds from the past. A catalogue of crop germplasm in the Central and North Italy, IdG, Bari
Hodgkin T, Ramanatha Rao V, Cibrian-Jaramillo A, Gaiji S (2003) The use of ex situ conserved plant genetic resources. Plant Genet Resour 1(1):19–29. https://doi.org/10.1079/PGR200313
IBPGR (1985) Descriptors for Setaria italica and S. pumila. International Board for Plant Genetic Resources, Rome, Italy
Joshi KD, Sthapit BR (1990) Informal research and development (IRD): a new approach to research and extension. LARC Discussion Paper 1990/4, Lumle, Pokhara, Nepal
Kumari R, Dikshit N, Sharma D, Bhat KV (2011) Analysis of molecular genetic diversity in a representative collection of foxtail millet [Setaria italica (L.) P. Beauv.] from different agro-ecological regions of India. Physiol Mol Biol Plants 17(4):363–374. https://doi.org/10.1007/s12298-011-0085-3
Li J, Wu SZ (1996) Traditional maintenance and multiplication of foxtail millet (Setaria italica (L.) P. Beauv.) in China. Euphytica 87:33–38
Mo FSC (2002) Nepal biodiversity strategy. Ministry of Forests and Soil Conservation, HMG, Nepal
Nakayama H, Namai H, Okuno K (1999) Geographical variation of the alleles at the two prolamin loci, Pro1 and Pro2, in foxtail millet, Setaria italica (L.) P. Beauv. Genes Genet Syst 74:293–297
Ning N, Yuan X, Dong S, Wen Y, Gao Z, Guo M, Guo P (2015) Grain yield and quality of foxtail millet (Setaria italica L.) in response to tribenuron-methyl. PLoS ONE 10(11):e0142557. https://doi.org/10.1371/journal.pone.0142557
Palikhey E, Sthapit SR, Gautam, S, Gauchan D, Joshi BK, Sthapit BR (2016) Baseline Survey Report: III. Haku, Jumla. Integrating traditional crop genetic diversity into technology: using a biodiversity portfolio approach to buffer against unpredictable environmental change in the Nepal Himalayas. LI-BIRD, NARC and Bioversity International, Pokhara, Nepal
Parajuli A, Subedi A, Sthapit SR, Gauchan D, Adhikari A, Joshi BK, Sthapit BR (2017) Baseline Survey Report: IV. Chhipra, Humla. Integrating traditional crop genetic diversity into technology: using a biodiversity portfolio approach to buffer against unpredictable environmental change in the Nepal Himalayas. LI-BIRD, NARC, DOA and Bioversity International, Pokhara, Nepal
Reddy VG, Upadhyaya HD, Gowda CLL (2006) Characterization of world’s foxtail millet germplasm collections for morphological traits. ISMN 47:107–109
Saha D, Channabyre Gowda MV, Arya L, Verma M, Bansal KC (2016) Genetic and genomic resources of small millets. Crit Rev Plant Sci 35:56–79. https://doi.org/10.1080/07352689.2016.1147907
Saud NB (2010) Crops of Nepal and their sustainable farming (in Nepali, Nepalka balinali ra tinko digo kheti). Sajha Prakashan, Pulchok, Lalitpur, Nepal, pp 223–227
Scheibe A (1943) Die Hirsen im Hindukush. Eine Beitrag zur Kenntnis von Kulturpflanzen in geographischer Rückzugsposition. Z Pflanzenzüchtg 25:392–436
Shannon CE, Weaver W (1949) The mathematical theory of communication. The University of Illinois, Chicago, pp 3–24
Sheikh SM, Singh O (2013) Pseudocereals and millets: the lost crops of Kashmir. Genet Resour Crop Evol 60:1191–1199
Stevens CJ, Murphy C, Roberts R, Lucas L, Silva F, Fuller DQ (2016) Between China and South Asia: a middle Asian corridor of crop dispersal and agricultural innovation in the Bronze Age. The Holocene 26(10):1541–1555. https://doi.org/10.1177/0959683616650268
Sthapit BR, Ramanatha Rao V (2009) Consolidating community’s role in local crop development by promoting farmer innovation to maximise the use of local crop diversity for the well-being of people. Acta Hort 806:669–676
Vavilov NI (ed) (1926) Centers of origin of cultivated plants. In Origin and geography of cultivated plants. Cambridge University Press
Witcombe JR, Khadka K, Puri RR, Khanal N, Sapkota A, Joshi KD (2016) Adoption of rice varieties. 2. Accelerating uptake. Exp Agric. https://doi.org/10.1017/s0014479716000624
Xiaomei F, Dong K, Wang X, Liu T, He J, Ren R, Zhang L, Liu R, Liu X, Li M, Huang M, Zhang Z, Yang Y (2016) A high density genetic map and QTL for agronomic and yield traits in foxtail millet [Setaria italica (L.) P. Beauv.]. BMC Genom 17:336. https://doi.org/10.1186/s12864-016-2628-z
Zhang S, Tang C, Zhao Q, Li J, Yang L, Qie L, Fan X, Li L, Zhang N, Zhao M, Liu X, Chai Y, Zhang X, Wang H, Li Y, Li W, Zhi H, Jia G, Diao X (2014) Development of highly polymorphic simple sequence repeat markers using genome-wide microsatellite variant analysis in foxtail millet [Setaria italica (L.) P. Beauv.]. BMC Genom 15:78. https://doi.org/10.1186/1471-2164-15-78
Zohary D, Hopf M, Weiss E (2012) Domestication of plants in the Old World. Oxford University Press, Oxford
Acknowledgements
This paper is the output of the GEF/UNEP supported project, “Integrating Traditional Crop Genetic Diversity into Technology: Using a Biodiversity Portfolio Approach to Buffer against Unpredictable Environmental Change in the Nepal Himalayas” implemented in Nepal. The project is coordinated by the Bioversity International in collaboration with Nepal Agricultural Research Council (NARC), Department of Agriculture (DoA) and Local Initiatives for Biodiversity Research and Development (LI-BIRD). Field and technical support of Mr. M.R. Bhatta, Mr. M. Bhattarai and Dr. M.N. Paudel (NAGRC), Dr. D. Jarvis and Dr. D. Gauchan (Bioversity International) are gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
There are no conflicts of interests to be declared.
Rights and permissions
About this article

Cite this article
Ghimire, K.H., Joshi, B.K., Gurung, R. et al. Nepalese foxtail millet [Setaria italica (L.) P. Beauv.] genetic diversity revealed by morphological markers. Genet Resour Crop Evol 65, 1147–1157 (2018). https://doi.org/10.1007/s10722-017-0602-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-017-0602-5