
Abstract
Wild palms contribute significantly to food security and local economy in tropical areas, and particularly in sub-Saharan Africa. In light of this importance, eight palm species were explored [Borassus aethiopum (L.) Mart, Eremospatha macrocarpa (G. Mann et H. Wendl.) H. Wendl., Laccosperma opacum (G. Mann et H. Wendl.) Drude, Hyphaene thebaica (L.) Mart, Phoenix reclinata Jacq., Raphia hookeri G. Mann et H. Wendl., R. sudanica A. Chev., and R. vinifera P. Beauv.] as targets for conservation, domestication, and cultivation in Benin. Cultivation potential was evaluated in a coarse-resolution, first-pass effort using ecological niche models to relate known occurrences of each species to vegetation indices (VEG), gross primary productivity (GPP), and soil characteristics (SOIL), and model outputs were related to human distribution and land-use patterns. Results showed that wild palms responded differentially to different suites of environmental factors: some species showed best model performance with VEG + GPP + SOIL, others with GPP + SOIL or VEG + GPP, or with a single factor. Two species had broad potential distributions across the country; others had potential areas in the north (2 species) or the south (4 species). Raphia hookeri and R. vinifera showed greatest overlap in terms of ecology and distribution, whereas L. opacum and R. sudanica had the lowest similarity. These models constitute initial steps toward a sustainable scheme for planning exploration of the possibility of cultivation of these species.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Palms represent an important plant family distributed in the tropics and subtropics globally. Wild palms have long been valued as natural resources by many local people for food supply, building material, medicines, fiber, oil, fruit, nuts, and more, for both local uses and commercial purposes, and especially in West Africa (Bekele-Tesemma 2007). In this region, wild palms are among the plants that have the greatest use, since almost all of the organs are used (Bi and Kouakou 2004). Many wild palm species exist, and most of them are considered open-access resources with no law regulating their exploitation.
In Benin, almost all parts of socioeconomically important palms are used (Akoègninou et al. 2006). In Mali, the wood is used for poles, shafts, and harpoons, and also as fuel and for making charcoal (Sidiyene 1996). A socioeconomic study in Côte d’Ivoire showed that the rattan trade is profitable at both rural and urban levels (Bi and Kouakou 2004). In Uganda, firewood and charcoal are obtained from Borassus aethiopum L. Mart and the strong trunks are used as posts and for construction of bridges (Tabuti 2007). The conjunction of these many uses and pressures could put at risk the natural populations of the species. To this end, appropriate solutions that would allow local people to benefit from these resources must be found without damaging or compromising wild populations and natural systems.
Protected areas have long been considered the most appropriate modality in conserving biodiversity (Naughton-Treves et al. 2005). However, it has become apparent that protected areas by themselves are insufficient to preserve biodiversity, especially in tropical areas (Furze et al. 1996). Small-scale agroforestry systems offer an intriguing alternative for threatened trees such as palms, and may make significant contributions to national economies and in maintaining some degree of forest cover in heavily-exploited systems.
To meet the increasing demands for palm-derived products without compromising their availability in the future, domestication and cultivation remain a key solution. Cultivation can reduce production time, while preserving desired markets and availability of the products (Mack 2000). Cultivation also may help to increase recognition of the importance of the wild species, and encourage preservation and restoration of actual wild populations (Cuni Sanchez et al. 2011). Even when not formally cultivated, some palm species are currently maintained in home gardens or peri-domestic agroforestry systems. To this end, assessing and identifying potential areas for cultivation of palm species using quantitative methods becomes an important research goal.
Cultivation potential of species such as Adansonia digitata L. (Cuni Sanchez et al. 2011) and Tamarindus indica L. (Fandohan et al. 2013) has been assessed in climatic dimensions; the relevant climatic variables are generally derived via spatial interpolation among weather station data (Hijmans et al. 2005). However, incorporation of local environmental features would give better and more relevant insights into suitability of areas (Idohou et al. 2015; Pearson and Dawson 2003), as climate-based models will frequently miss fine details owing to their broad spatial autocorrelation structure and the interpolation that is necessary for their derivation. Satellite imagery provides finer spatial resolution, and also the advantage of reflecting more dimensions of the environmental landscape, including dimensions of land use and land cover, soil characteristics, and topography. Development of effective models of cultivation potential for wild palm species requires detailed, multi-dimensional assessment that can extend to finer spatial resolutions thanks to such data resources. Such a fine-scale assessment is the aim of this study.
Materials and methods
Species occurrence records
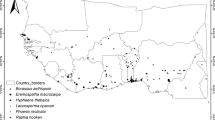
All socioeconomically important wild palm species occurring in Benin were examined, based on occurrence data from across their geographic distributions in the country (Idohou et al. 2015). The species were Borassus aethiopum, Eremospatha macrocarpa (G. Mann et H. Wendl.) H. Wendl., Laccosperma opacum (G. Mann et H. Wendl.) Drude, Hyphaene thebaica (L.) Mart, Phoenix reclinata Jacq., Raphia hookeri G.Mann et H. Wendl., R. sudanica A. Chev., and R. vinifera P. Beauv. An intensive field survey during 2013–2014 served to collect occurrence data for the eight species at fine spatial resolutions (Fig. 1), as all records were documented with good spatial precision via GPS readings. Duplicate records and records with obvious errors were discarded prior to analysis using utilities in ENMtools (www.ENMTools.com) and visual inspections.

Map of the study area showing occurrence data of each of the eight palm species
Environmental data
Environmental data included two major sources: remotely sensed data on vegetation, and interpolated data on soil characteristics. Vegetation data were obtained in the form of imagery and data obtained from the Moderate Resolution Imaging Spectroradiometer (MODIS) satellite at 1 km spatial resolution, using the NASA Reverb Echo data portal (https://reverb.echo.nasa.gov/reverb/). All 16-day composite data layers for 2013 and 2014 were obtained, in terms of Normalized Difference Vegetation Index (NDVI), Enhanced Vegetation Index (EVI) and Gross Primary Production (GPP). Soil data were also obtained at 1 km resolution, from the Africa Soil Profiles Database (https://www.isric.org). Soil characteristics included soil organic carbon (g/kg), pH in H2O, sand content (%), silt content (%), clay content (%), cation exchange capacity (cmol/kg), and bulk density (t/m3), for six depth horizons (0–5, 5–15, 15–30, 30–60, 60–100, and 100–200 cm). These variables have excellent potential to be highly informative for this project as they meld land use and vegetation characteristics with indirect reflections of climate, and because they offer a reasonably fine spatial resolution (1 km) with no interpolation in the case of remotely sensed data.
Data analysis and interpretation
Occurrence data for each species were split into training (50 %) and testing (50 %) subsets five times at random, to permit assessment of how robust and predictive the models are. To reduce dimensionality of environmental spaces and create composite environmental variables with minimal collinearity, principal component analyses (PCA) was performed on each of the environmental data sets (NDVI and EVI, being highly correlated, were combined into a single data set, termed VEG). All combinations of the three sets of environmental layers (GPP, GPP + SOIL, GPP + SOIL + VEG, GPP + VEG, etc.) were considered.
The maximum entropy algorithm MaxEnt version 3.3.3 (Phillips et al. 2006) was used to link environmental conditions to species’ occurrence patterns, as this approach is considered to be robust and predictive (Elith and Graham 2009; García-Callejas and Araújo 2015). Initially, 10 bootstrap replicate models were developed for each of the 5 random subsamples for the 8 species with 7 environmental data combinations, for 2800 total model runs. Results of each of the models were evaluated using a partial ROC procedure (Peterson et al. 2008), a threshold-independent approach that considers ability to predict distributions of independent occurrence data in a relatively small predicted area; the procedure requires specification of an omission error tolerance (in this case, given that the occurrence data were carefully collected and geo-referenced by the first author, this error parameter E was set to 0). In all, 100 random 50 % sub-sampling iterations of the partial ROC were run to permit assessment of whether the AUC ratios that result were significantly elevated above the AUC ratio of 1.0, which is the performance of a random classifier (Peterson et al. 2008). Via these model evaluation steps, we were able to select optimal combinations of environmental data sets for each species, at which point we produced final models for each species using all data available.
Resulting maps were thresholded to create binary maps to avoid effects of overfitting common in MaxEnt models (Peterson et al. 2007) by seeking the highest (i.e., most restrictive) threshold that included 95 % of the occurrence data used in model calibration, which is a modification of the least training presence thresholding approach (Pearson et al. 2007), and which appropriately weights omission error more heavily than commission error (Fielding and Bell 1997; Peterson 2014).
To identify potential cultivation areas for each species, binary maps were overlaid with water data (http://glcf.umd.edu/data/watercover/) human demographic patterns (2013 census; http://wms.cartographic.com.www2.lib.ku.edu/LandScan2013/) and the existing protected areas network (http://www.wdpa.org). These post-model-calibration modifications of the potential cultivation area were applied under the idea that heavily populated or protected areas will not constitute appropriate cultivation regions for palm species, and that water will be necessary for the species first initiation.
The degree of range overlap between final binary models for pairs of palm species was assessed to understand similarity of range and ecology. To this end, the D and I similarity measures implemented in ENMTools (version 1.4.4) were used as measures of range similarity across Benin (Warren et al. 2010). Both indices range from 0, indicating complete dissimilarity, to 1, which indicates precise range overlap.
Results
Exploration of the 7 combinations of environmental variables showed different suites of environmental drivers for distributions of each of the species (Table 1). For B. aethiopum, all three environmental drivers (GPP, VEG, SOIL) were necessary to achieve best model performance. For E. macrocarpa, L. opacum, R. hookeri, and R. vinifera, highest partial ROC AUC ratios were obtained with data on soils only. Vegetation and soils data were necessary to obtain best performance for H. thebaica and R. sudanica; P. reclinata required data on GPP and VEG. Partial ROC evaluations indicated that all of our models were robust, yielding predictions statistically significantly better than random (P < 0.01).
Geographically, the results suggested high potential for cultivation of palm species across Benin. Suitable areas for B. aethiopum and P. reclinata (Fig. 2) were broad, and crossed the three biogeographic zones of the country. For E. macrocarpa, L. opacum, R. hookeri, and R. vinifera (Fig. 3), suitable areas were concentrated in the southern part of the country (Guineo-Congolean zone) only. For H. thebaica and R. sudanica, models identified the northern (Sudanian) areas, with some extension southward towards the Guineo-Sudanian zone (Fig. 4).

Suitable areas for cultivation of Borassus aethiopum and Phoenix reclinata across Benin

Suitable areas for cultivation of Eremospatha macrocarpa, Laccosperma opacum, Raphia hookeri, and Raphia vinifera across Benin

Suitable areas for cultivation of Hyphaene thebaica and Raphia sudanica across Benin
Overlay of potential distributional areas with other environmental features (protected areas network, human demography) showed that, for B. aethiopum, H. thebaica, P. reclinata, and R. sudanica, areas around some protected areas were suitable for the species; these areas held only sparse human populations (Figs. 5, 6, 7). For other species with more restricted distributions (E. macrocarpa, L. opacum, R. hookeri and R. vinifera; Figs. 2, 3, 4), water was also generally in close proximity to potential distributional areas, but human populations were high and protected areas were few.

Maps of the potentially suitable areas for Borassus aethiopum and Phoenix reclinata with consideration of human population distribution and existing protected areas

Maps of the suitable areas for Eremospatha macrocarpa, Laccosperma opacum, Raphia hookeri, and Raphia vinifera cultivation with consideration of human population distribution and existing protected areas

Maps of the suitable areas for Hyphaene thebaica and Raphia sudanica cultivation with consideration of human population distribution and existing protected areas
Assessing range overlap in both geographic and environmental dimensions, overlap was highest between R. hookeri and R. vinifera (Tables 2, 3). Overlap between L. opacum and R. vinifera was also high (Tables 2, 3). At the opposite end of the spectrum, L. opacum and R. sudanica showed lowest overlap (Tables 2, 3), suggesting low similarity between potential areas for these two species.
Discussion
Wild palm species showed diverse responses to environmental drivers in terms of their distributional characteristics across Benin. Whereas, for some species, the ecological niche was influenced by single factors, others showed effects of 2 or 3 suites of environmental variables. For B. aethiopum, all three environmental dimensions were needed to characterize its potential areas for cultivation: although the species is distributed countrywide and is found in different habitats, its presence appears to require specific combinations of environmental conditions. Indeed, vegetation indices (providing important information on canopy mass) were very important for this species, which makes sense, as it is known to be heliophilous (Giffard 1967; Lamotte 2012). These requirements may differ among systems and soil types, as B. aethiopum is found on diverse soil types, but may be replaced by other palm species on the heavier sodic/saline soils in drainage lines and more arid areas (Katende et al. 1995). For species like H. thebaica and R. sudanica, their cultivation potential is more related to vegetation and soil characteristics; E. macrocarpa, L. opacum, R. hookeri, and R. vinifera were affected only by soil characteristics, probably because they occur along river banks and marshy areas. Whereas R. hookeri, and R. vinifera occur in the understory, E. macrocarpa is extremely light-demanding (Ebanyenle and Oteng-Amoako 2003), and so occurs along the margins of forest vegetation.
Almost all of the species analyzed in this study showed potential for cultivation across their distributions, although palms often require a shallow ground water table for population establishment and reproduction. This point is important, as cultivation of these palms could reduce pressures on existing wild populations (Blach-Overgaard et al. 2010). Promotion of wild palm cultivation can also help to raise awareness of the multiple uses for the species and their historical and cultural significance. Opportunities exist now for their incorporation in the current country-wide reforestation program based on native species, which has been announced via the slogan, “10 million hearts, 10 million trees.” Palms could also be introduced in home gardens, as a recent study revealed low representation of these species in home gardening (Idohou et al. 2014), but the success of such ideas will depend on cultural acceptance.
Local and regional markets exist presently for palm-derived products (Belcher and Schreckenberg 2007; Bi and Kouakou 2004); incentives could be offered to increase demand and create new markets, locally and regionally, for palm-derived products. Care should be taken with regard to agroforestry development for some palms (B. aethiopum and H. thebaica), as they occur in areas presently under cotton production, with intensive pesticide use. In such areas, intensive agriculture may degrade soils or lead to erosion.
Cultivation of palm species in areas not currently hosting the species (e.g., H. thebaica in the Guineo-Sudanian zone) should be undertaken with care, as success will depend on whether the species is able to adapt to the new environment—such variation may manifest in dimensions not assessed in this study, and to avoid creation of invasive-species problems. As such, for species with widespread potential cultivation areas, further assessments would be needed, particularly in genetic dimensions; such studies could reveal ecotypes or differentiated populations. Regardless of the species and sites, sensitization and training of local people will also be needed, so that they are aware of best practices for cultivation and potential benefits from wild palm products.
This study is however limited in several ways. In particular, we emphasize that it is intended as a first-pass assessment of the potential for domestication and cultivation of these palm species. If a particular species were to become the object of more intensive efforts, many additional considerations should be taken into account, such as details of soil composition, which are clearly both immediately relevant and difficult to summarize using current remote-sensing data products. We performed the analyses with only currently available data, once again as a first-pass assessment of cultivation potential; these models can be improved as more data—both in term of occurrence information and in terms of environmental information—but this result is a summary of knowledge and inference available at present.
Conclusion
Wild palms showed great potential to domestication although the species responded differently to environmental conditions. While some species showed best performance for the combination of the three variables, other performed well with two or just one variable. Some species could be cultivated across the country while others have the potential areas for cultivation in one or two zones. For both indices and for almost all species some concordance has been noticed in the species overlapping. Experiments are highly needed and could help to validate the current models for a sustainable cultivation of the species.
References
Akoègninou A, Van der Burg WJ, L.J.G. VdM (2006) Flore analytique du Bénin. Blackhuys Publishers, Cotonou & Wageningen
Bekele-Tesemma A (2007) Useful trees and shrubs for Ethiopia. Identification, propagation and management for 17 agroclimatic zones. RELMA in ICRAF Project, World Agroforestry Centre, Eastern Africa Region
Belcher B, Schreckenberg K (2007) Commercialisation of non-timber forest products: a reality check. Dev Policy Rev 25:355–377. doi:10.1111/j.1467-7679.2007.00374.x
Bi IZ, Kouakou KL (2004) Etude de la filière rotin dans le district d’Abidjan (Sud Côte d’Ivoire). Biotechnol Agron Soc 8:199–209
Blach-Overgaard A, Svenning J-C, Dransfield J, Greve M, Balslev H (2010) Determinants of palm species distributions across Africa: the relative roles of climate, non-climatic environmental factors, and spatial constraints. Ecography 33:380–391
Cuni Sanchez A, Osborne PE, Haq N (2011) Climate change and the African baobab (Adansonia digitata L.): the need for better conservation strategies. Afr J Ecol 49:234–245. doi:10.1111/j.1365-2028.2011.01257.x
Ebanyenle E, Oteng-Amoako AA (2003) Anatomy and identification of five indigenous rattan species of Ghana. Ghana J For 11:77–90
Elith J, Graham CH (2009) Do they? How do they? WHY do they differ? on finding reasons for differing performances of species distribution models. Ecography 32:66–77
Fandohan B, Gouwakinnou GN, Fonton NH, Sinsin B, Liu J (2013) Impact des changements climatiques sur la répartition géographique des aires favorables à la culture et à la conservation des fruitiers sous-utilisés: cas du tamarinier au Bénin. Biotechnol Agron Soc 17:450–462
Fielding AH, Bell JF (1997) A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ Conserv 24:38–49
Furze B, De Lacy T, Birckhead J (1996) Culture, conservation and biodiversity: the social dimension of linking local level development and conservation through protected areas. Wiley, New York
García-Callejas D, Araújo MB (2015) The effects of model and data complexity on predictions from species distributions models. Ecol Model 326:4–12. doi:10.1016/j.ecolmodel.2015.06.002
Giffard PL (1967) Le palmier rônier Borassus aethiopum Mart. Bois For Trop 116:3–12
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25:1965–1978. doi:10.1002/joc.1276
Idohou R et al (2014) Biodiversity conservation in home gardens: traditional knowledge, use patterns and implications for management. Int J Biodivers Sci Manag 10:89–100
Idohou R, Ariño AH, Assogbadjo AE, Glele R (2015) Knowledge of diversity of wild palms (Arecaceae) in the Republic of Benin: finding gaps in the national inventory by combining field and digital accessible knowledge. Biodivers Inform 10:45–55
Katende AB, Birnie A, Tengnäs B (1995) Useful trees and shrubs for Uganda: identification, propagation and management for agricultural and pastoral communities. Technical Handbook 10. Regional Soil Conservation Unit, Nairobi
Lamotte M (2012) Trop Savannah Ecosyst Trop Ecol Syst Trends Terr Aquat Res 11:179
Mack R (2000) Cultivation fosters plant naturalization by reducing environmental stochasticity. Biol Invasions 2:111–122. doi:10.1023/A:1010088422771
Naughton-Treves L, Buck Holland M, Brandon K (2005) The role of protected areas in conserving biodiversity and sustaining local livelihoods. Annu Rev Env Resour 30:219–252. doi:10.1146/annurev.energy.30.050504.164507
Pearson RG, Dawson TP (2003) Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob Ecol Biogeogr 12:361–371. doi:10.1046/j.1466-822X.2003.00042.x
Pearson RG, Raxworthy CJ, Nakamura M, Peterson AT (2007) Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J Biogeogr 34:102–117. doi:10.1111/j.1365-2699.2006.01594.x
Peterson AT (2014) Mapping disease transmission risk: enriching models using biogeography and ecology. JHU Press, Baltimore
Peterson AT, Papes M, Eaton M (2007) Transferability and model evaluation in ecological niche modeling: a comparison of GARP and Maxent. Ecography 30:550–560
Peterson AT, Papeş M, Soberón J (2008) Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol Model 213:63–72. doi:10.1016/j.ecolmodel.2007.11.008
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190:231–259. doi:10.1016/j.ecolmodel.2005.03.026
Sidiyene EA (1996) Trees and spontaneous shrubs of the Adrar des Iforas (Mali). Orstom
Tabuti JRS (2007) The uses, local perceptions and ecological status of 16 woody species of Gadumire Sub-county, Uganda. Biodivers Conserv 16:1901–1915. doi:10.1007/s10531-006-9097-7
Warren DL, Glor RE, Turelli M (2010) ENMTools: a toolbox for comparative studies of environmental niche models. Ecography 33:607–611
Acknowledgments
This work was fully funded by University of Abomey-Calavi through Wild Palm project and the World Bank via a scholarship (RSM McNamara fellowship) provided to Rodrigue Idohou for a 6 months stay at the University of Kansas. We are grateful to the entire Biogeography and Macroecology lab group for fruitful exchange and support.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests
The author(s) declare that they have no conflict of interests.
Rights and permissions
About this article

Cite this article
Idohou, R., Townsend Peterson, A., Assogbadjo, A.E. et al. Identification of potential areas for wild palm cultivation in the Republic of Benin through remote sensing and ecological niche modeling. Genet Resour Crop Evol 64, 1383–1393 (2017). https://doi.org/10.1007/s10722-016-0443-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-016-0443-7