
Abstract
There are evidences that plant morphology is shaped by genotype, but local environment mainly climate influences morphology as well. In this study the morphological variability of Vitex doniana, a multipurpose tree species was characterised in relation with climatic parameters in order to provide insights to the species possible responses to future climate change. Morphological data were collected on 102 trees randomly selected along unfixed transects in the three climatic zones of Benin. Data were collected on fruiting trees at three levels: tree (trunk and canopy), leaves and fruits. Variance components were estimated for identification of variability sources regarding leaves and fruits characteristics. The most important discriminant descriptors regarding climatic zones were selected through a stepwise discriminant analysis. Relationship between those discriminant morphological traits and bioclimatic variables were assessed through a redundancy analysis. Our findings confirmed that there is an important variability of morphological traits of the species and climate, mainly some of its extremes parameters plays a non-negligible role. Trees in the Sudanian region are the biggest with fruits producing little pulp while individuals in the more humid Guinean region present a higher amount of pulp whereas Sudano-Guinean trees are the tallest with larger leaves. Although the climate-induced variability of the species is relatively low, the study gives insights in probable effects of climate variability on its morphology. Population genetic studies are required for a better understanding of climatic impacts on V. doniana in order to develop selection and domestication schemes which could contribute to its conservation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Morphological variability reflecting general intraspecific genetic variability occurs in most plant species as response to local environmental fluctuations (Ellison et al. 2004; Miner et al. 2005). This variability is an essential component of plant fitness and a precondition to adapt to various environments and is also the basis for selection by humans for domestication purposes (Cornelius 1994; Ellison et al. 2004; Ewédjè et al. 2012). There are evidences that plant morphology is shaped by genotype, but local environment mainly climate can play an important role too (Schlichting and Pigliucci 1998; Ellison et al. 2004; Guerin et al. 2012). It is therefore important to evaluate this variability in order to sustainably manage useful plant species and give insights into probable effects of unpredictable hazards like climate change.
Our study focused on Vitex doniana Sweet, an important agroforestry tree species widespread in tropical Africa. Different parts of the plant are widely used for several purposes among which are young leaves for sauces preparation and ripened fruits for direct consumption being most common among people in its geographical range (Ky 2008). The mature leaves, the bark and the roots are also used to treat several diseases (Kilani 2006; Padmalatha et al. 2009; Dadjo et al. 2012). Moreover, it presents some agronomic potentialities regarding soil fertility improvement through litter production (Mapongmetsem et al. 2005). In Benin, V. doniana is known as a particular agroforestry tree with high socio-economic values (Oumorou et al. 2010; Achigan-Dako et al. 2011; Dadjo et al. 2012).
Some studies have addressed its uses in pharmacology (Kilani 2006; Padmalatha et al. 2009) and in food processing (Okigbo 2003; Agbédé and Ibitoyé 2007), its ethnobotanical values (Dadjo et al. 2012), domestication status and germinative abilities (Sanoussi et al. 2012; Achigan-Dako et al. 2014; N’Danikou et al. 2014). However, little is known on its morphological variability and the relationship between this variability and climatic conditions.
Following the concept of phenotypic plasticity—capacity of a given genotype to produce physiological or morphological phenotypes in response to variations of environmental conditions (Pigliucci 2006)—we expect that V. doniana develops specific morphological traits according to climatic zones in Benin. More specifically, since the three climatic zones of Benin reflect a south-north climate drying gradient (wet–dry) coupled with an opening of the vegetation (forest–savannah), we hypothesize that (1) due to the availability of water and competition for light, trees of the Guinean zone are higher, thinner, with the best fruiting performance and larger leaves (lower evapotranspiration); (2) in contrast, trees of the Sudanian zone are shorter, thicker, with wider canopy, smaller leaves (adaptation to higher evapotranspiration) and lower fruiting performance; (3) trees in the Sudano-Guinean zone are assumed to have intermediate characteristics.
Therefore, we characterized the morphological variability of the species in relation with climatic conditions in order to provide relevant information on its probable responses to future climate change.
Materials and methods
Target species and study area
The black plum (Vitex doniana Sweet, Lamiaceae formerly Verbenaceae) is a deciduous plant species widespread in tropical Africa, from Senegal to Somalia and to South Africa, and also in Comoros and Seychelles (Ky 2008; Orwa et al. 2009). It is occasionally cultivated in Mauritius. The species occurs in various habitats from forests to savannahs, often in wet localities and along rivers, and on termite mounds, up to 2000 m altitude. Regions with mean annual rainfall between 750–2000 mm and temperatures ranging from 10 to 30 °C are suitable for the species (Ky 2008). The variability of V. doniana is remarkable regarding its morphology as well as its habitat choice. It is a deciduous small to medium sized tree species up to 25 m, bole branchless for up to 11 m with a diameter which can reach 160 cm. It has opposite and digitate compound leaves. The flowers are bisexual and zygomorphic and the fruits obovoid to oblong-ellipsoid drupes (2–3 cm long), purplish black, fleshy, with woody 4-celled stone, up to 4-seeded (Arbonnier 2002; Ky 2008) .
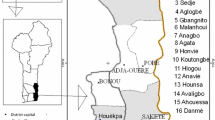
The study was carried out in the three climatic zones of Benin Republic in West Africa (Fig. 1). The Guinean zone located between 6°25′ and 7°30′N is characterised by a subequatorial climate with four seasons (two rainy and two dry). The rainfall is bimodal (averagely 1200 mm per year), the temperature varies between 25 and 29 °C, and relative humidity is between 69 and 97 %. The Sudano-Guinean zone, from 7°30′ to 9°45′N, is a transitional zone with two rainy seasons merging in a unimodal regime. The annual rainfall varies between 900 and 1110 mm, the temperature is between 25 and 29 °C and relative humidity ranges from 31 to 98 %. The Sudanian zone, between 9°45′ to 12°25′N, has a tropical dry climate with two equally long seasons (rainy and dry). The mean annual rainfall in this zone is often <1000 mm; the relative humidity varies from 18 to 99 % and temperature from 24 to 31 °C (Adomou et al. 2006; Assogbadjo et al. 2012; Gnanglè et al. 2012).

Location of sampled trees in Benin Republic. Climatic zones (from South to North: Guinean, Sudano-Guinean and Sudanian zone) are adapted from Adomou et al. (2006)
Data collection
Morphological data were collected from July to October 2014 in the three climatic zones following unfixed transects (length varying from 1 to 6 km). Seven, six and nine transects were followed respectively in the Guinean, Sudano-Guinean and Sudanian zone. A distance of at least 20 m was left between fruiting trees in order to avoid genetically close individuals. A total of 102 fruiting individuals of V. doniana were assessed (35, 31, and 36 respectively in the Guinean, Sudano-Guinean and Sudanian zone). Ten leaves and ten fruits were randomly selected per tree (Cornelissen et al. 2003) for morphological description. Moreover, data were collected on the trunk, the canopy, the leaves and the fruits in the field (Fig. 2). On the trunk, circumference at 1.3 m (circ, cm), total height (htree, m) and bole height (hbole, m) i.e. height from ground to the first big branch were measured. Concerning the canopy, four radii (r i ) were measured from the projection of the crown on the ground (Fig. 2a). For the leaves, leaf length (Llgth, mm), length and width of the main leaflet (llgth and lwdth, mm), petiole length and diameter (petlgth and petdiam, mm) and number of leaflets (lnumb) were noted (Fig. 2b). For the fruits, length and width of fruit (flgth and fwdth, mm) and fruit fresh mass (ffmass, g) were measured (Fig. 2c).

Morphological descriptors measured on V. doniana (adapted from Ky (2008)). a Tree: circumference at 1.3 m (circ, cm); total height (htree, m); bole height (hbole, m) and crown height (hcrown, m) and crown radii (r i , m). b Leaf: leaf length (Llgth, mm), length and width of the main leaflet (llgth and lwdth, mm), petiole length (petlgth, mm). c Fruit: length and width of fruit (flgth and fwdth, mm)
These parameters were measured with an electronic digital calliper (0.01 mm resolution) for length and width and a 0.01 g-precision scale for weight.
After measuring the above described parameters, fruits were oven dried at 65 °C for 48 h for determination of their dry mass (fdmass, g) following Fandohan et al. (2011). The pulp and fibres have been removed from the dry fruits in order to have the endocarp for which length (endolgth, mm), width (endowdth, mm) and mass (endomass, g) were measured.
Finally, the GPS coordinates of each tree were recorded and used in QGIS 1.8.0 (Quantum Development Team 2016) to extract bioclimatic data for each tree from WorldClim 2.5′ × 2.5′ database (Hijmans et al. 2005).
Data processing and analysis
From the measured parameters, the following variables were computed:
-
Mean diameter of canopy (dcrown, m): It was computed from the four measured radii (r i , i from 1 to 4) using the following formula (Glèlè Kakaï et al. 2011):
$$ dcrown = 2\sqrt {\sum \frac{{r_{i}^{2} }}{4}} $$(1) -
Height of canopy (hcrown, m): It is the difference between total height and bole height
$$ hcrown = htree - hbole $$(2) -
Canopy shape (shcrown): It is the ratio of height of canopy over mean diameter of canopy
$$ shcrown = \frac{hcrown}{dcrown} $$(3) -
Pulp Thickness (pulpthick, mm): It is the difference between fruit width and endocarp width divided by 2.
$$ pulpthick = \frac{{\left( {fwdth - endowdth} \right)}}{2} $$(4) -
Pulp mass (pulpmass, g): It is the difference between dry mass of fruit and endocarp mass
$$ pulpmass = fdwght - endowght $$(5) -
Fruit shape (fshape): It is the ratio of fruit length over fruit width
$$ fshape = \frac{flgth}{fwdth} $$(6) -
Endocarp shape (endshape): It is the ratio of endocarp length over endocarp width
$$ endshape = \frac{endolgth}{endowdth} $$(7)
Mean values, standard error of mean and coefficient of variation were computed for trunk and canopy traits per climatic zone. The same descriptive statistics were calculated for each morphological parameter of leaves and fruits per climatic zone taking into account variation in fruits/leaves by using the two-stage sampling formulas of Cochran (1977). The skewness was also calculated for each morphological trait to appreciate normality of its distribution within climatic zone. One way-ANOVA was used to test for differences in trunk/canopy morphological traits between climatic zones. The analysis was performed on log transformed data except for crown diameter in order to meet normality and homogeneity of variance assumptions. Student-Newman-Keuls’ posthoc test was used to classify mean values of those morphological traits. Meanwhile, linear mixed effects models using the restricted maximum likelihood (REML) from lme4 (Bates et al. 2015) and lmerTest packages (Kuznetsova et al. 2014) were performed on leaves and fruits morphological traits for the same comparison purpose and to estimate the part of their variability explained by between-climatic zone effect and within-climatic zone effect (tree-within climatic zone, tree-within transect and within-tree effects). Since sample sizes (number of transects per climatic zone, number of trees considered per climatic zone and per transect) were unequal, Satterthwaite approximation was used as correction formula for the degrees of freedom in order to have accurate P values (Mason et al. 2003). Tukey’s contrasts for multiple comparisons of means from multcomp package (Hothorn et al. 2008) was used for means classification (Mangiafico 2015). For these linear mixed effects models, climatic zone with its three modalities (Guinean, Sudano-Guinean and Sudanian) was considered as fixed factor while transect (7, 6 and 9 respectively in the Guinean, Sudano-Guinean and Sudanian zones) and fruiting tree (from which ten leaves/fruits were measured) were considered as nested factors.
A stepwise linear discriminant analysis was performed in order to find out the most important morphological traits that discriminate trees regarding climatic zones. For this analysis, morphological traits at trunk and canopy levels and mean values per tree for leaves and fruits characteristics were used.
Finally, a redundancy analysis (RDA) was performed to assess relationships between morphological descriptors and bioclimatic variables. The analysis was run on the standardized means values of the previously selected discriminant morphological traits and all the 19 bioclimatic variables extracted from WorldClim database. The best model was selected by removing sequentially bioclimatic variables with the step function in Vegan package (Oksanen et al. 2013). The model selection was based on the Akaike’s Information Criterion (AIC) which measures the goodness of fit, with the best model showing the small AIC (Burnham and Anderson 2003). For the selected model, significance of canonical axes was tested using the marginal permutation test from Vegan package. Moreover, the significance of each bioclimatic variables in the model was tested using the permutation ANOVA test. All these analyses were run with the vegan package (Oksanen et al. 2013).
R 3.2.2 software (R Core Team 2015) was used for the one-way ANOVA, linear mixed effects models and RDA while SPSS 20 (SPSS IBM. 2011) was used for the stepwise discriminant analysis.
Results
Morphological variability of Vitex doniana
Morphological descriptors of trunk and canopy show high amplitude of variation in all climatic zones with coefficients of variation above 25 % (Table 1). However, these morphological descriptors do not vary in the same way from one climatic zone to another. For instance, circumference at 1.3 m and crown diameter showed the greatest variability in the Guinean zone (CV = 48.37 %), while bole height was the most fluctuating trait in the Sudano-Guinean zone (CV = 51.21 %). The other descriptors (tree height, crown height and crown shape) were most dispersed in the Sudanian zone (Table 1). The Skewness coefficient of all these descriptors (except tree height in the Sudano-Guinean zone) was positive suggesting right asymmetrical distribution. Apart from the crown diameter, all the descriptors of trunk and canopy varied significantly between climatic zones. Trees from the Sudanian zone were the thickest while trees from the Sudano-Guinean zone were the tallest with relatively rounded crown (crown shape i.e. ratio hcrown/dcrown = 1, Table 1).
Most of the morphological descriptors of leaves and fruits were relatively less dispersed (CV <15 %) around mean values in all climatic zones (Table 2). Leaves and fruits descriptors showed the highest amplitude of variation in respectively the Guinean and Sudanian climatic zone. The Skewness coefficient was close to zero for most of the leaves and fruits features in all climatic zones suggesting a normal distribution. All the leaves and fruits descriptors varied significantly between climatic zones and the lowest values were mostly recorded in the Sudanian zone. Trees in the Sudano-Guinean zone presented slightly higher leaves characteristics, while the greatest features for fruits were mostly recorded in the Guinean zone (Table 2). In general, fruits were relatively oblong (length/width ratio was slightly above 1), but fruits from the Sudano-Guinean zone were longer than wide.
The variance components estimation revealed that the within-climatic zone effect represented the most important source of variability of leaves and fruits traits, with variation within tree being the highest (Table 3). The between climatic zones variability of leaves and fruits traits was relatively weak (<30 % of total variation). Its lowest and highest values were recorded respectively for number of leaflets per leaf (2.19 %) and endocarp mass (26.93 %). Variability in morphological traits due to climatic zone was higher for fruits characteristics than for leaves (Table 3).
Discriminant morphologic descriptors of Vitex doniana according to climatic zones
The stepwise discriminant analysis indicated that two significant discriminant axes were needed to separate V. doniana trees according to climatic zones (P = 0.000). The model accuracy rate revealed that 84.3 % of trees were perfectly assigned to their respective climatic zone. The Wilks’Lambda statistic ranged from 0.757 to 0.195 (first to last step) and P value was 0.000. Only 7 of the 23 descriptors were the most discriminant for climatic zone. The first axis explaining 85.5 % of the total variance discriminated trees based on tree height, canopy height, leaf length and number of leaflets. This axis separated Sudano-Guinean trees from the others. The second axis accounting for 14.5 % of total variance performed trees discrimination based on circumference, petiole diameter and pulp mass. It discriminated Guinean trees from Sudanian trees (Fig. 3).

Projection of V. doniana trees regarding morphological traits in the system defined by the two canonical functions. Function 1 and 2 represent respectively 85.5 and 14.5 % of the total variance of 23 descriptors on 102 individuals. Symbols correspond to trees from each climatic zone of Benin (Circles Sudanian trees, Lozenges Sudano-Guinean trees, Crosses Guinean trees, Squares centroid of each group)
Trees from the Sudano-Guinean zone were taller with higher canopy. They exhibited longer leaves with often more than 5 leaflets per leaf whereas those from the Sudanian zone were thicker (high circumference values) with relatively small petiole diameter for leaves and produced fruits with reduced pulp mass. Guinean trees thus diverged from the Sudanian counterparts.
Relationships between discriminant morphological descriptors and bioclimatic variables
From the redundancy analysis, the best model explaining the relationships between the discriminant morphological descriptors of V. doniana and bioclimatic variables took into account 8 of the 19 bioclimatic variables (F = 7.095, P value = 0.001, AIC = 842.4, adjusted R2 = 0.326). Only the first two axes were significant (P value = 0.001) and they explained 37.9 % of the total variation in morphological traits due to the bioclimatic variables. The first axis (RDA1) explaining 32.05 % of total variance, was a linear combination of annual mean diurnal range of temperature (bio2), isothermality (bio3), temperature seasonality (bio4), precipitation seasonality (bio15), and mean temperature of the coldest quarter (bio11). The second axis (RDA2, 0.06 % of total variation) on the other hand, was a linear combination of annual precipitation (bio12), precipitation of the warmest quarter (bio18) and mean temperature of the warmest quarter (bio10). However some of these bioclimatic variables such as annual precipitation and mean temperature of the coldest quarter were not statistically significant in the model (Table 4).
Regarding the discriminant morphological variables, circumference at 1.3 m, tree height, crown height, leaf length, petiole diameter and pulp mass were loaded on RDA1, while number of leaflets was loaded on RDA2 (Table 5).
Based on scores of morphological traits and bioclimatic variables on RDA axes (Tables 4, 5), it was noted that circumference at 1.3 m, tree height, crown height, leaf length and petiole diameter were positively influenced by annual mean diurnal range of temperature (bio2), temperature seasonality (bio4) and precipitation seasonality (bio15). Meanwhile, pulp mass and number of leaflets were negatively affected respectively by isothermality (bio3) and mean temperature of the warmest quarter (bio10). The projection of V. doniana trees in the RDA axes system showed that Guinean trees were mainly located on the negative side of RDA1 and RDA2 while Sudanian trees were mainly in the positive side of RDA2 (Fig. 4). The morphology of trees from these zones was likely affected by bioclimatic parameters in diverse way.

Projection of V. doniana trees in the RDA correlation triplot (scaling = 2) defined by the two first axes. Trees are represented by triangles (green, blue and black for respectively Guinean, Sudano-Guinean and Sudanian trees). Bioclimatic variables are represented by arrows (bio 2 = Annual mean diurnal range of temperature; bio 3 = Isothermality; bio 4 = Temperature seasonality; bio 10 = Mean temperature of the warmest quarter; bio 11 = mean temperature of the coldest quarter; bio 12 = Annual precipitation; bio 15 = Precipitation seasonality; bio 18 = Precipitation of the warmest quarter). Morphological variables (circ circumference, htree total height of tree, hcrown crown height, Llgth leaf length, petdiam petiole diameter, lnumb number of leaflets, pulpmass pulp mass)
There were no direct significant effects of the commonly used climatic parameters such as annual mean temperature and precipitation on morphological traits of V. doniana. However, several bioclimatic variables based on their extremes such as temperature/precipitation of the warmest quarter and other related parameters showed significant influences on the morphological descriptors (Table 4).
Discussion
In the present study, we investigated the morphological diversity of V. doniana in relation to climate in order to highlight probable effects of climate change on the species. The morphological description was performed at three levels: tree (trunk and canopy), leaf and fruit. Findings from the study confirmed that there is an important variability of morphological traits in the species and climate plays a non-negligible role.
The relatively important dispersion of morphological descriptors around mean values (Tables 1, 2) gives evidences of the existence of heterogeneity in V. doniana populations as mentioned by Ky (2008). This heterogeneity can be seen as a result of fitness of the species in various habitats and can increase its survival chance under climate change (Visser 2008). Moreover, the existence of this variability strengthens the potential of the species in cultivars selection for a domestication process (Leakey et al. 2008; Fandohan et al. 2011). Indeed, genetic diversity improves fitness of populations by providing options to respond differently to environmental changes (Whitham et al. 2003; Hughes and Stachowicz 2004).
Significant differences were observed between climatic zones regarding the considered morphological traits, but part of variability due to climatic zone was relatively low and the most important variability was generally recorded at individual tree level (Table 3). These findings are in agreement with previous studies on native and introduced plant species in Benin such as Adansonia digitata L. (Assogbadjo et al. 2011), Jatropha curcas L. (Gbèmavo et al. 2015), Sclerocarya birrea subsp. birrea (A. Rich) Hochst. (Gouwakinnou et al. 2011). Since morphology is under the control of genotype, environment and their interactions, with genotype leading the control (Schlichting and Pigliucci 1998), our findings show that most of the variability in V. doniana was probably genetically-induced. Therefore, population genetic studies about this species are required to deepen knowledge on its gene pool in relation with climatic parameters.
Although the projection of V. doniana trees in the axes system defined by the two canonical functions from the discriminant analysis did not show a clear separation of trees (Fig. 3), the major differences between trees were revealed regarding position of group centroids. Based on contrasting features of Guinean and Sudanian trees (heavy vs. light pulp mass for example), climate may be considered as playing an important role in the morphological variability of the species. Indeed, the Guinean zone known to be wetter and more forested than the Sudanian zone, hosted thinner and taller trees with globally larger leaves and higher fruiting parameters mainly regarding fruits and pulp masses. These results confirmed our expectations and may suggest that the species undergoes adaptive strategies in regard to climatic factors. According to trees in the Sudano-Guinean zone, our findings showed some deviation from the expected trends. Trees are generally taller with larger leaves in this zone than elsewhere. These trends might suggest that this climatic zone is very close to the humid climatic conditions. Previous studies have already highlighted the role of climate for the morphological variation of several useful species. This was the case for instance for Anogeissus leiocarpa (DC.) Guill. et Perr. in Burkina Faso (Ouédraogo et al. 2013).
The redundancy analysis revealed significant relationships between bioclimatic parameters and discriminant morphological descriptors of V. doniana. Although the bioclimatic parameters explained a relatively low part of variation in morphological traits, impacts of climate on the species’ morphology are obvious even if there might not be strong. This relatively weak influence of the bioclimatic variables on morphological descriptors emphasized findings on low variability due to climate. Also, the low influence of bioclimatic parameters might be due to the fact that climate is not the only factor affecting the morphology of the species. Indeed, environmental factors like topography, soil properties (type, moisture, nutrients content) have been reported as potential predictors (Sankaran et al. 2005). Since soil types and properties varied significantly across climatic zones of Benin (Adomou et al. 2006), soil may have contributed to the morphological variation of V. doniana in Benin. The study also revealed that mean annual temperature and rainfall do not have direct impacts on the discriminant descriptors, but rather their extremes (temperature and precipitation of the warmest quarter) and other related parameters such as annual diurnal range of temperature, temperature and precipitation seasonality that drive the plant’s morphological diversity.
Conclusion
In the present study, we investigated the morphological diversity of V. doniana in relation with climate in order to highlight probable effects of climate change on the species. The morphological description was performed at three levels: tree (trunk and canopy), leaves and fruits. The study revealed that V. doniana shows an important morphological variability and that climate contributes somehow to its morphological diversity in Benin. Moreover, our findings suggest that extreme parameters of a changing climate in the future could affect the morphology of the species. However, population genetic studies should be undertaken on the species in order to improve knowledge on its variability in relation with climate and help in developing selection and domestication schemes for the species. Finally, conservation programs should be envisaged for the species by taking into account this morphological variability.
References
Achigan-Dako EG, N’Danikou S, Assogba-Komlan F, Ambrose-Oji B, Ahanchédé A, Pasquini MW (2011) Diversity, geographical, and consumption patterns of traditional vegetables in sociolinguistic communities in Benin: implications for domestication and utilization. Econ Bot 65(2):129–145
Achigan-Dako EG, N’Danikou S, Tchokponhoué DA, Komlan FA, Larwanou M, Vodouhè RS, Ahanchédé A (2014) Sustainable use and conservation of Vitex doniana Sweet: unlocking the propagation ability using stem cuttings. J Agric Environ Int Dev 108(1):43–62
Adomou AC, Sinsin B, Van der Maesen LJG (2006) Phytosociological and chorological approaches to phytogeography: a meso-scale study in Benin. Syst Geogr Plants 76(2):155–178
Agbédé JO, Ibitoyé AA (2007) Chemical composition of black plum (Vitex doniana): an under-utilized fruit. J Food Agric Environ 5(2):95–96
Arbonnier M (2002) Arbres, arbustes et lianes des zones sèches d’Afrique de l’Ouest, 2nd edn. Editions Quae, France
Assogbadjo AE, Glèlè Kakaï R, Edon S, Kyndt T, Sinsin B (2011) Natural variation in fruit characteristics, seed germination and seedling growth of Adansonia digitata L. Benin. New For 41(1):113–125. doi:10.1007/s11056-010-9214-z
Assogbadjo AE, Chadaré FJ, Glèlè Kakaï R, Fandohan B, Baidu-Forson JJ (2012) Variation in biochemical composition of baobab (Adansonia digitata) pulp, leaves and seeds in relation to soil types and tree provenances. Agric Ecosyst Environ 157:94–99. doi:10.1016/j.agee.2012.01.021
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67(1):1–48. doi:10.18637/jss.v067.i01
Burnham KP, Anderson DR (2003) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Cochran WG (1977) Sampling techniques, 3rd edn. Wiley, New York
Cornelissen JHC, Lavorel S, Garnier E, Diaz S, Buchmann N, Gurvich DE, Reich PB, Ter Steege H, Morgan HD, Van Der Heijden MGA et al (2003) A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust J Bot 51(4):335–380
Cornelius J (1994) The effectiveness of plus-tree selection for yield. For Ecol Manag 67(1):23–34
Dadjo C, Assogbadjo AE, Fandohan B, Glèlè Kakaï R, Chakeredza S, Houehanou TD, Damme PV, Sinsin B (2012) Uses and management of black plum (Vitex doniana Sweet) in Southern Benin. Fruits 67(4):239–248. doi:10.1051/fruits/2012017
Ellison AM, Buckley HL, Miller TE, Gotelli NJ (2004) Morphological variation in Sarracenia purpurea (Sarraceniaceae): geographic, environmental and taxonomic correlates. Am J Bot 91(11):1930–1935
Ewédjè E-EBK, Parmentier I, Natta A, Ahanchédé A, Hardy OJ (2012) Morphological variability of the tallow tree, Pentadesma butyracea Sabine (Clusiaceae), in Benin. Genet Resour Crop Evol 59:625–633. doi:10.1007/s10722-012-9802-1
Fandohan B, Assogbadjo AE, Glèlè Kakaï R, Kyndt T, Sinsin B (2011) Quantitative morphological descriptors confirm traditionally classified morphotypes of Tamarindus indica L. fruits. Genet Resour Crop Evol 58(2):299–309. doi:10.1007/s10722-010-9575-3
Gbèmavo CJ, Gandji K, Gnanglè CP, Assogbadjo AE, Glèlè Kakaï RL (2015) Variabilité morphologiques et conservation des morphotypes de Jatropha curcas Linn. (Euphoriaceae) au Benin. J Agric Environ Int Dev 109(1):55–69. doi:10.12895/jaeid.20151.234
Glèlè Kakaï R, Akpona T, Assogbadjo AE, Gaoué OG, Chakeredza S, Gnanglè PC, Mensah GA, Sinsin B (2011) Ecological adaptation of the shea butter tree (Vitellaria paradoxa CF Gaertn.) along climatic gradient in Bénin, West Africa. Afr J Ecol 49(4):440–449
Gnanglè PC, Egah J, Baco MN, Gbèmavo CDSJ, Glèlè Kakaï R, Sokpon N (2012) Perceptions locales du changement climatique et mesures d’adaptation dans la gestion des parcs à karité au Nord-Bénin. Int J Biol Chem Sci. doi:10.4314/ijbcs.v6i1.13
Gouwakinnou GN, Assogbadjo AE, Lykke AM, Sinsin B (2011) Phenotypic variations in fruits and selection potential in Sclerocarya birrea subsp. birrea. Sci Hortic 129(4):777–783. doi:10.1016/j.scienta.2011.05.041
Guerin GR, Wen H, Lowe AJ (2012) Leaf morphology shift linked to climate change. Biol Lett 8(5):882–886. doi:10.1098/rsbl.2012.0458
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A et al (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25(15):1965–1978
Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biom J 50(3):346–363. doi:10.1002/bimj.200810425
Hughes AR, Stachowicz JJ (2004) Genetic diversity enhances the resistance of a seagrass ecosystem to disturbance. Proc Natl Acad Sci USA 101(24):8998–9002
Kilani AM (2006) Antibacterial assessment of whole stem bark of Vitex doniana against some enterobactriaceae. Afr J Biotechnol 5(10):958–959
Kuznetsova A, Brockhoff PB, Christensen RHB (2014) lmerTest: tests in linear mixed effects models. R package Version 2.0-20. https://cran.r-project.org/web/packages/lmertest/lmertest.pdf
Ky KJM (2008) Vitex doniana sweet. In: Louppe D, Oteng-Amoako AA, Brink M (eds) Prota: Timbers/Bois d’oeuvre, vol 7(1). Backhuys Publishers, Leiden, pp 578–581
Leakey R, Fuller S, Treloar T, Stevenson L, Hunter D, Nevenimo T, Binifa J, Moxon J (2008) Characterization of tree-to-tree variation in morphological, nutritional and medicinal properties of Canarium indicum nuts. Agrofor Syst 73(1):77–87
Mangiafico SS (2015) An R companion for the handbook of biological statistics, version 1.09i. Rutgers Cooperative Extension, New Brunswick
Mapongmetsem PM, Benoit LB, Nkongmeneck BA, Ngassoum MB, Gübbük H, Baye-Niwah C, Longmou J (2005) Litterfall, decomposition and nutrients release in Vitex doniana Sweet. and Vitex madiensis Oliv. in the Sudano–Guinea Savannah. Akdeniz Üniv Ziraat Fak Derg 18(1):63–75
Mason RL, Gunst RF, Hess JL (2003) Statistical design and analysis of experiments: with applications to engineering and science, vol 474, 2nd edn. Wiley, Hoboken
Miner BG, Sultan SE, Morgan SG, Padilla DK, Relyea RA (2005) Ecological consequences of phenotypic plasticity. Trends Ecol Evol 20(12):685–692. doi:10.1016/j.tree.2005.08.002
N’Danikou S, Achigan-Dako EG, Tchokponhoué DA, Assogba Komlan F, Gebauer J, Vodouhè RS, Ahanchédé A (2014) Enhancing germination and seedling growth in Vitex doniana Sweet for horticultural prospects and conservation of genetic resources. Fruits 69(4):279–291. doi:10.1051/fruits/2014017
Okigbo RN (2003) Fermentation of black plum (Vitex doniana Sweet) juice for production of wine. Fruits 58(06):363–369
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2013) Vegan: community ecology package. R package version 2.0-10. http://CRAN.R-project.org/package=vegan
Orwa C, Mutua A, Kindt R, Jamnadass R, Simons A (2009) Agroforestree Database: a tree reference and selection guide version 4.0. http://www.worldagroforestry.org/af/treedb/
Ouédraogo A, Glèlè Kakaï R, Thiombiano A (2013) Population structure of the widespread species, Anogeissus leiocarpa (DC.) Guill. & Perr. across the climatic gradient in West Africa semi-arid area. S Afr J Bot 88:286–295. doi:10.1016/j.sajb.2013.07.029
Oumorou M, Sinadouwirou T, Kiki M, Glèlè Kakaï R, Mensah GA, Sinsin B (2010) Disturbance and population structure of Vitex doniana Sw. in northern Benin, West Africa. Int J Biol Chem Sci 4(3):624–632
Padmalatha K, Jayaram K, Raju NL, Prasad MNV, Arora R (2009) Ethnopharmacological and biotechnological significance of Vitex. Bioremediat Biodivers Bioavailab 3(1):6–14
Pigliucci M (2006) Phenotypic plasticity and evolution by genetic assimilation. J Exp Biol 209(12):2362–2367. doi:10.1242/jeb.02070
Quantum Development Team (2016) Quantum GIS geographic information system. 2.12.3-Lyon. Free Software Foundation, India
R Core Team (2015) R: a language and environment for statistical computing. Version 3.2.2. R Foundation for Statistical Computing, Vienna
Sankaran M, Hanan NP, Scholes RJ, Ratnam J, Augustine DJ, Cade BS, Gignoux J, Higgins SI, Le Roux X, Ludwig F et al (2005) Determinants of woody cover in African savannas. Nature 438(7069):846–849
Sanoussi A, Ahoton LE, Odjo T (2012) Propagation of black plum (Vitex donania Sweet) using stem and root cuttings in the ecological conditions of South Benin. Tropicultura 30(2):107–112
Schlichting C, Pigliucci M (1998) Phenotypic evolution: a reaction norm perspective. Sinauer Associates Incorporated, Sunderland
SPSS IBM (2011) Statistical package for social science. Version 20. SPSS IBM, Inc., Chicago
Visser ME (2008) Keeping up with a warming world; assessing the rate of adaptation to climate change. Proc R Soc B 275:649–659. doi:10.1098/rspb.2007.0997
Whitham TG, Young WP, Martinsen GD, Gehring CA, Schweitzer JA, Shuster SM, Wimp GM, Fischer DG, Bailey JK, Lindroth RL et al (2003) Community and ecosystem genetics: a consequence of the extended phenotype. Ecology 84(3):559–573
Acknowledgments
Authors thank the German Ministry of Education and Research (BMBF) for its financial support to AH through the West African Service Centre on Climate Change and Adapted Land use (WASCAL). They are also grateful to field guides for their courage and determination during data collection and to Enoch Bessah for language editing.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
We declare that there is no conflict of interest.
Rights and permissions
About this article

Cite this article
Hounkpèvi, A., Azihou, A.F., Kouassi, É.K. et al. Climate-induced morphological variation of black plum (Vitex doniana Sw.) in Benin, West Africa. Genet Resour Crop Evol 63, 1073–1084 (2016). https://doi.org/10.1007/s10722-016-0409-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-016-0409-9