
Abstract
The aim of this study was used a basic hypothetical structural model with latent variables to analyze the interconnections between the pools of stable P (inorganic P (Pi) and organic P (Po)), labile P (Pi and Po) and available P (Mehlich-1 P) and the pools of organic matter (OM) content and physicochemical properties in tropical soils of differing pedogenesis. We used structural equation modeling for designing models for two groups of soil: (1) mineral soils with low to medium organic matter content and (2) mineral soils with high organic matter content and organic soils. The proposed structural models were consistent with the hypothesis of dependence between the pools of P and organic matter as well as physicochemical properties in tropical soils. In general, stable and labile P pools acted as P sources for the available P pool; furthermore, the strength of these structural relationships was strongly associated with soil organic matter content. Yet the pool of physicochemical properties behaved as a sink of P for the labile P pool, however with a beneficial effect in maintaining the stable P pool. The pools of P and OM are strongly bonded in tropical soils under different pedogenesis. All structural models evidenced that various forms of P in different levels of lability could contribute in keeping the supply of bioavailable P, yet its magnitude would be regulated by P buffer capacity of each soil.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Phosphorus (P) availability is one of the most limiting factors for productivity of forestry systems in highly weathered tropical soils (Johnson et al. 2003; Vitousek et al. 2010; Costa et al. 2016). This shortcoming is due to a low total P content in the different types of soil, aside from a high adsorption of inorganic P (Pi) on the surface of Al and Fe oxides, which are dominant in the soil mineral phase (Yang and Post 2011). This means tropical soils are predominantly Pi sinks, in which most of the Pi is fixed, remaining thus inaccessible to plants (Condron and Tiessen 2005). As a result, production costs increases since larger amounts of phosphate fertilizers are needed to meet P demands of crops (Grierson et al. 2004). Against this background, the understanding of soil P transformations is relevant, particularly in non-fertilized soils wherein P uptake by plants would be regulated prevailingly by organic P (Po) turnover (Gama-Rodrigues et al. 2014).
Bowman’s sequential extraction (Bowman 1989) has been considered to be suitable for comparisons of total Po contents across different tropical soils types (Condron et al. 1990). Several studies have been carried out on Po dynamics in soils under forests and pastures (Guerra et al. 1996; Cunha et al. 2007; Zaia et al. 2008a, 2012; Rita et al. 2013; Oliveira et al. 2014). Additionally, these studies have also assessed the fraction of labile Po (rapid mineralization compounds) extracted with bicarbonate (Bowman and Cole 1978). The findings showed that soils in advanced weathering stage had a higher amount of total Po, which ranged from 29 to 42% of the total P, while labile Po varied from 35 to 75% in soil surface layers (Guerra et al. 1996; Cunha et al. 2007; Zaia et al. 2008a, 2012; Oliveira et al. 2014). Conversely, the same studies have shown a wide variation regarding the influence of organic matter and other physico-chemical properties on Po and Pi fractions among various soil types, and seemingly contradictory results among the case studies. For instance, Zaia et al. (2012) reported a negative relation between Po (total and labile) with organic C and clay in Oxisols and Inceptisols under cocoa agroforestry; however, Cunha et al. (2007) found a positive relation among these attributes in Oxisols and Inceptisols under forests and pastures.
Therefore, the results of these studies demonstrate that some statistical tools as Pearson’s correlation, multiple regression and principal component analysis (PCA) are limited in a simultaneous understanding of all relations between P fractions (Pi and Po) and soil physical and chemical properties. As a result, structural equation modeling (SEM) has been considered appropriate to analyze multiple relationships within P cycle in tropical soils (Gama-Rodrigues et al. 2014; Sales et al. 2015; Costa et al. 2016).
SEM allows analyzing and validating relational theories through strong causal connections when studying complex phenomena with empirical data and theoretical concepts using non-observed latent variables (Grace 2006). Latent variables (constructs) are conceptual variables that form and explain a set of observed variables and are incorporated into structural equation models to be theoretically most consistent with reduced measurement errors, as well as testing hypotheses of multiple processes operating within a system (Sales et al. 2015). Furthermore, the SEM can also be used for exploring several possible multivariate alternative hypotheses (Eisenhauer et al. 2015). Such assumptions are simultaneously analyzed by imposed relationships between two (or more) model variables. This way, a set of interrelated structural equations is formed, being represented by composite structural models of latent variables and measurement models. Thus, the relationships between each latent variable and the group of measurable variables are imposed on these models (Byrne 1994; Bollen e Noble 2011).
The cause-effect aspects among variables within a model are understood by means of SEM; in this model, it is possible to measure how changes of one variable can influence other variables within a model. That way, an understanding of the causes is critical not only to support theories, but also these models are a rigorous method to predict potential impacts related to natural sources or even human activity (Grace et al. 2015). This feature differentiates the SEM from other conventional statistical techniques, which study cause-effect assumptions individually.
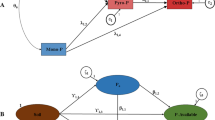
In this study, we elaborated hypothetical structural equation models to analyze interconnections between the P cycle and soil properties based on the Bowman’s extraction method (Bowman 1989). The latent variables of the basic model were composed of pools of stable P (Pi and Po), labile P (Pi and Po) and available P (Mehlich-1 P) associated with pools of organic matter (OM) and physicochemical properties of soil (PCP) (Fig. 1). The theoretical concept consisted of evaluating how much the pool of available P is controlled by different P pools in association to different pools of attributes in soils of differing pedogenesis in natural and managed ecosystems.

The hypothesized structural model for the soil P cycle. (OM = Organic matter pool, PCP = Physicochemical properties). The ζi (i = 1–4) represent the variables of random error variance of the endogenous latent variables. The parameters δj,i represent the regression coefficients between the endogenous latent variables of position i and the endogenous latent variables of position j (with i and j integers ranging from 1 to 4). The parameters γj,i represent the regression coefficients between the exogenous latent variable of position i and the endogenous latent variables of position j, (i = 1, j = 2 to 4). The symbols (+) and (−) represent, respectively, the positive and negative effects of the structural assumptions
Materials and methods
Table 1a shows the phosphorus (P) data gathered from literature corresponding to studies performed using the sequential extraction methods of Bowman and Cole (1978) and Bowman (1989). The database (n = 68) covers a wide range of soil types, encompassing non-fertilized soils with low to medium organic matter content (from Entisols to Oxisols) under various tropical land uses (natural forest, planted forest, cocoa agroforest, pasture and savannah woodland). The fractions of P used were Mehlich-1 P (extractable P), bicarbonate P (Pi and Po) and stable P (Pi and Po) fractions. The first is associated with the fraction readily available for plant absorption (available P). The second is the amount released by ligand exchange with bicarbonate ions (labile P or reversibly available) (Sato and Comerford 2006). Yet the third represents non-labile P fraction, i.e. a non-readily available form (sparingly available). Stable Pi and Po were estimated by the differences between total and labile Pi and Po, respectively. In addition to that, the soil properties such as the contents of organic carbon (C), total nitrogen (N), aluminum oxide (Al2O3), iron oxide (Fe2O3), remaining P (Prem), clay and the soil pH were also used to assess their effect on soil P availability. The fraction of Prem represents the buffer capacity of P in the soil and it was measured in equilibrium solution after homogenizing a 5-cm3 soil sample with 50 mL of a 10 mmol L−1 CaCl2 solution containing 60 mg L−1 P for 1 h (Oliveira et al. 2014).
Figure 1 shows a hypothesized basic structural model designed for estimates of the soil P cycle. Based on that, we could identify direct and indirect influences of the latent variables [stable P, labile P, organic matter (OM) and physicochemical properties of soil (PCP)] on the endogenous latent variable (available P). We assumed that the functional P pools (unmeasurable variables = latent) derived from the various P fractions are categorized as a function of their degree of availability to plants: Mehlich-1 P was the indicator variable of the available P pool, bicarbonate P (Pi and Po) were the indicator variables of the labile P pool, and stable P (Pi and Po) were the indicator variables of the stable P pool. In addition, C and N were the indicator variables of the soil OM pool. The indicator variables C, N and Po were not put in the same pool based on the McGill and Cole (1981) model, in which C and N are stabilized together and Po is stabilized independently of the main organic moiety. Clay, Al2O3, Fe2O3, Prem and pH were the indicator variables of the soil PCP pool. These indicator variables of the PCP pool make part of the abiotic processes of stabilization of P (Pi and Po) (Celi and Barberis 2005) and organic matter (C and N) (Wiseman and Püttmann 2006) in soils. Thus, the basic hypothetical model had the PCP pool as exogenous latent variable and all P pools and OM pool as endogenous latent variables.
Figure 1 also displays the following specific hypotheses tested on the basic hypothetical model:
- H1 :
-
The stable P pool acts directly as a sink of P on the available P pool;
- H2 :
-
The stable P pool acts directly as a source of P for the labile P pool;
- H3 :
-
The stable P pool acts positively on the soil OM pool;
- H4 :
-
The OM pool acts positively on the labile P pool;
- H5 :
-
The soil PCP pool acts positively on the OM pool;
- H6 :
-
The soil PCP pool acts positively on the stable P pool;
- H7 :
-
The soil PCP pool acts negatively on the labile P pool;
- H8 :
-
The labile P pool acts directly as source of P on the available P pool
The hypothesized basic structural model was analyzed in five steps as proposed by Bollen and Noble (2011): (1) model specification, (2) model identification, (3) estimation of parameters, (4) model fit test, and (5) model respecification. The fitted model were set through comparisons with database collected by Oliveira et al. (2014), which consists of 38 observations covering a wide range of soil surface layers of high OM content, such as histic H and O, chernozemic A and humic A (Table 1b).
Both univariate and multivariate normality were tested for all observed variables (skewness and kurtosis), followed by proper transformations, using particularly Log 10. Then, descriptive statistics were applied to the databases (Table 2), and the Pearson’s correlation analysis was made for all the variables which assist in understanding the relationships established in the hypothetical models. In addition, the collinearity between independent variables highly correlated with values ≥0.90 was tested (Hair et al. 2009). We also assumed the correlation structure to be obtained from a single random sample as being equal to that observed through literature data.
The error variance and factor loading were set to build the hypothesized structural model, according criteria referred by Sales et al. (2015) in order to change an unidentified model into an identified one. Thus, it was possible to estimate all other model free parameters (Grace 2006; Byrne 2009). Assuming the absence of measurement errors, the variance error for Mehlich-1 P was set to zero; therefore, its factor loading was then set to one. As for the latent variables (also constructs) stable and labile P and OM, values of 0.1 (10%) were set. All presented models had these specifications.
Once the model was identified, the standardized parameters were estimated and convergent validity of the constructs determined. The convergence of a construct indicates how well it was developed. For this analysis, it is essential the use of standardized factor loadings (the data were standardized by applying a mean of zero and standard deviation of (1), once high loadings (ideally ≥0.7) lead to a common conversion point (Hair et al. 2009). The convergence was calculated using variance extracted (VE) and construct reliability (CR).
The VE indicates how much of an indicator variance can be explained by the respective constructs, being calculated as follows:
where \(\lambda_{i}\) represents standardized factor loadings and \(\varepsilon_{i}\) represents the terms for error variance of each construct. The VE is considered suitable for values above 0.5, implying in proper convergence of its indicators (Fornell and Larcker 1981; Hair et al. 2009).
The CR is also an indicator of convergent validity and may be calculated through the following expression:
where reliability values above 0.7 are ideal and those between 0.6 and 0.7 are acceptable. However, the reliability values of other constructs should be suitable for the model to be considered adequate for the analysis (Fornell and Larcker 1981; Hair et al. 2009).
The maximum likelihood method (ML) was used to estimate model parameters. For overall fit, the Chi-squared (X 2), degrees of freedom (df) and probability level (P) associated with the model were used. In the χ2 test, the hypothetical model has good fit when P value is >0.05, and so the null hypothesis is not rejected. Given the χ2 test’s sensitivity to sample sizes, the goodness fit index (GFI), the comparative fit index (CFI), the root mean square error of approximation (RMSEA) and X 2/df ratio were considered as alternatives to measure the model fitting. GFI and CFI values ≥0.95 (Hair et al. 2009) and RMSEA ≤ 0.08 (Byrne 2009) show proper model fitting. According to Iacobucci (2010), the ratio X 2/df should be less than three for an acceptable model fit. The overall effect (direct + indirect) of exogenous construct on the endogenous construct corresponds to the regression coefficient of structural equations (β).
For the respecified models, we calculated the χ2 statistical difference (∆X 2). Whether this difference is above 3.84 and ∆df is one, we can conclude that at a 0.05 probability level, the alternative model was better fitted (Hair et al. 2009). The Akaike’s Information Criterion (AIC) was also used as comparative fit index of models (Kenny 2014), which the lower, the better the model is fit. The SEM analyses were carried through AMOS software, version 22 (IBM—SPSS Inc., Chicago, IL, USA).
Results
The soil samples used here showed a wide range in concentrations of the different P fractions and physicochemical properties of soil (Table 2). Therefore, through correlation analysis, it was possible to evaluate how these P fractions varied for each soil property and the interrelationships among them. Thus, we could identify a close relationship among all P fractions in soils of low to medium OM content (Table 3). In addition, C and N contents had significant relationships with all P fractions. The Mehlich-1 P also had a significant positive relationship with pH, but negative with clay and Fe2O3. Whereas stable Pi had a significant positive relation to clay, and stable Po to pH. As expected, C and N correlation was positively significant; and both of these elements were positively related to clay and negatively to Fe2O3 and Al2O3. In contrast, there were no significant correlations between all P fractions in soils with high OM content (Table 4). There was not significant positive relationship of the Mehlich-1 P fraction only with stable Po, we also observed significant correlation between Po–HCO3 and Pi–HCO3, as well as between both stable P fractions. Regarding the significant correlations between P fractions and soil properties, we noted negative relationships of stable P and Prem, as well as between pH against Mehlich-1 P and HCO3 P fractions; however, these P fractions had positive correlations with C and N. In addition, only stable Pi had positive correlations to Fe2O3 and Al2O3.
By identifying the hypothetical structural model (Model A—Fig. 2), the fit indexes were χ 2 = 190.375, df = 40, P < 0.0001. These adjustment indexes indicate that the proposed structural model has not yet been fitted to the data. Other indices for overall fit are shown in Table 5, confirming the lack of fit of this model. However, the directions (positive or negative) of all structural relationships corresponding to the model specific hypotheses were confirmed, except for H4, H5 and H6.

Model A, hypothesized structural model for the P cycle in soils with low to medium organic matter content. All measured variables (in boxes) are represented as effect indicators associated with latent variables (in circles). The numbers correspond to the standardized parameters estimated (P < 0.001) and the R2 values (numbers in bold); “ns” no significant. Error variables (ε1–ε7 and δ1–δ4) are standardized values. Model χ 2 = 190.375, df = 40, P < 0.0001. (n = 68)
In model A, stable and labile P, OM, and PCP pools showed an VE of 71, 67, 81, and 45%, respectively. Moreover, the same constructs presented respective CR values of 83, 80, 90, and 58%. These values point out that PCP was the only exogenous latent variable mathematically non-adapted to literature specifications. Furthermore, the explained variance (R 2) of the available P pool (endogenous latent variable) was of 58%.
As model A did not show proper fit, we carried its respecification by removing the PCP pool (exogenous latent variable), once its VE and CR were unsatisfactory. This way, model B is a simplified version of the complete structural model, having the stable P pool as exogenous construct (Fig. 3). This model showed a highly satisfactory overall adjustment in all tests (Table 5), and stable and labile P, and OM pools explained 57% of the variance (R 2) of the available P pool.

Model B, structural model respecified for the P cycle in soils with low to medium organic matter content. All measured variables (in boxes) are represented as effect indicators associated with latent variables (in circles). The numbers correspond to the standardized parameters estimated (P < 0.001) and the R2 values (numbers in bold); “ns” no significant. Error variables (ε1, ε4– ε7, δ5, δ6) are standardized values. Model χ 2 = 14.374, df = 11, P = 0.213. (n = 68)
In order to confirm model B, we used data from 38 soil samples with high organic matter content (Table 2). Nonetheless, while trying to estimating the parameters, we could not identify the model because of the high correlation between C and N of the OM pool causing collinearity problem (Table 4). As a result, the database was reduced to 28 samples (Table 2), removing samples of Histosols (histic H and O horizons), decreasing thus the correlation between the variables C and N (r = 0.738, P < 0.001). Thus, model B1, which is a confirmation of the model B, was prepared and identified with highly satisfactory overall fit indexes (Table 5) and increase in the explained variance (R 2 = 0.60) of the endogenous variable available P pool (Fig. 4).

Model B1, structural model for the P cycle in soils with high organic matter content. All measured variables (in boxes) are represented as effect indicators associated with latent variables (in circles). The numbers correspond to the standardized parameters estimated (P < 0.001) and the R2 values (numbers in bold). †, * and **significant at P < 0.10, 0.05 and 0.01 respectively, and “ns” no significant. Error variables (ε1, ε4–ε7, δ5, δ6) are standardized values. Model χ 2 = 6.004, df = 10, P = 0.815. (n = 28)
The available P pool was directly, positively and strongly dependent on the labile P one for soils of low to medium OM content (Model B—Fig. 3); while for soils of high content, such relationship was observed with the stable P pool (Model B1—Fig. 4). The stable P pool, as exogenous construct, had a distinct overall effect (direct + indirect) on the available P pool of both models. In model B1, the overall effect of stable P (β = 0.68 + [(0.35 × 0.14) + (0.49 × 0.69 × 0.14)] = 0.776) was 45% superior to that found in model B (β = −0.02 + [(0.71 × 0.770) + (0.54 × 0.02 × 0.77)] = 0.535). The model B show that the main effect of the stable P pool on the available P pool was indirect through labile P pool; while in B1, the relationship of those pools was direct and corresponded to 87.6% of the overall effect of the exogenous variable. Additionally, the overall effect of stable P pool on the labile P pool was similar for both models B (β = 0.71 + (0.54 × 0.02) = 0.721) and B1 (β = 0.35 + (0.49 × 0.69) = 0.688). Nonetheless, in model B, this effect corresponded to 98.5% of the overall effect of exogenous variable; while in B1, direct and indirect effects were similar.
On the other side, applying the same database as in B1 (n = 28), we could establish an alternative model using as exogenous construct the PCP pool, however, without the OM pool (Model C—Fig. 5). The variables pH and clay were excluded from the PCP pool, and the Prem was included. After these changes, the model C showed a satisfactory overall fit in all tests, except in GFI (Table 5). Furthermore, the validity of the constructs was also altered. In this model, the PCP pool yielded a VE = 52% and CR = 80%, VE = 59% and CR = 73% for stable P pool, and VE = 65% and CR = 78% for labile P pool. However, there were decreases in the explained variance (R 2 = 0.50) for the endogenous latent available P pool.

Model C, structural model for the P cycle in soils with high organic matter content. All measured variables (in boxes) are represented as effect indicators associated with latent variables (in circles). The numbers correspond to the standardized parameters estimated (P < 0.001) and the R2 values (numbers in bold). †, * and **significant at P < 0.10, 0.05 and 0.01 respectively, and “ns” no significant. Error variables (ε1–ε5, δ1–δ3) are standardized values. Model χ 2 = 15.308, df = 16, P = 0.502. (n = 28)
By including samples of Histosols, a full database was used (n = 38), setting thus the model C1, which showed proper fit and index values similar to model C (Table 5). However, a slight change in VE and CR values was observed for the constructs in this model. VE = 62% and CR = 76% for the stable P pool, VE = 56% and CR = 71% for the labile P pool, and VE = 57% and CR = 79% for the PCP pool. The models C and C1 differ from one another by changes in values and significance levels of the estimated parameters for structural relationships in C1. In addition, the R 2 = 0.83 value, for endogenous latent variable available P pool, indicates a good explained variation. There has also been distinction between models C and C1 for overall effect of PCP pool on the available P pool. In model C, overall effect was positive (β = (0.52 × 0.43) + (0.52 × 0.73 × 0.37) + [(−0.33) × 0.37] = 0.242); however, in model C1, it was negative and almost null (β = (0.58 × 0.37) + (0.58 × 0.54 × 0.77) + [(−0.66) × 0.77] = −0.052). In turn, the available P pool was direct, positive and strongly dependent on the stable P and labile P pools, for both models. In model C, overall effect of the stable P pool on the available P pool was positive (β = 0.44 + (0.72 × 0.36) = 0.699) and near its effect found in C1 (β = 0.37 + (0.54 × 0.77) = 0.786).
Discussion
The adjusted structural models were consistent with the theoretical plausibility of P availability to plants being affected by interconnections between the P cycle and the organic and physicochemical properties of soil. The structural relations of the adjusted model B (Fig. 3), for low to medium levels of soil organic matter, were also statistically confirmed in soils with high organic C content (Model B1—Fig. 4). Therefore, we can conclude that the pools of P and OM are strongly bonded in tropical soils under different pedogenesis. In this context, estimated parameter direction (positive or negative), magnitude and significance of the structural relationships within P cycle were affected by the OM pool.
In both models, the regression between stable P pool and OM pool explains 24–29% of the OM variability (Figs. 3, 4). The non-explained variability of this structural relationship stems from several factors such as management practice, soil type, climate, relief and land use. Organic C and total N, which are indicators of the OM pool, were positively correlated with fractions of stable P, mainly in soils of low OM content (Table 3). These results suggest that part of the P fractions of low biological availability would not be limiting for accumulation of OM in the soil. Furthermore, this close structural relationship should be considered when part of the stable Po is physically incorporated into soil OM structures (Celi and Barberis 2005). Based on Walker and Syers (1976) conceptual model, we can infer that as total P is a primary soil property, i.e. not being dependent on any other variable, the availability and transformations of soil P are a major control on accumulation and transformations of soil OM (Tiessen et al. 1984). Thus, we can make the assumption that varying the stable P pool would lead to changes in OM pool, but the variation of OM pool would not lead to changes in the stable P pool a long-term.
On the other hand, the OM pool together with the stable P pool explains 52—83% of the variability in the labile P pool (Figs. 3, 4). This effect was stronger in soils with high OM contents (Model B1—Fig. 4). Some processes or mechanisms may contribute to a positive relationship between those pools, among them: (1) several kinds of labile C could be substrate in transformations of solution Pi into labile Po (Grierson et al. 2004); (2) the competition of organic anions with Pi for sites of high adsorption can release this element, thereby reducing the fixation rate thereof (Guppy et al. 2005; Souza et al. 2014); (3) organic anions can form complexes with Fe and Al (P-Fe and P-Al), thus releasing P to the soil solution (Bais et al. 2006); (4) the OM pool with high N content can have a positive effect on the labile P pool, since an increased in N availability is essential for the production of enzymes rich in N (phosphatases) which mineralize the Po (Nasto et al. 2014), increasing the bioavailability of P. The production of such exoenzymes by plants and microorganisms has been demonstrated in a substantial increase responding to an increment of N in rainforest soils (Olander and Vitousek 2000; Treseder and Vitousek 2001; Marklein and Houlton 2012). In addition, part of the absorbed N by plants and microorganisms is also a constituent of various phosphate compounds, particularly various P-diesters (Turner et al. 2005), which return to soil through turnover of litter, root and microbial biomass, thus restoring the labile P pool. As all these soil processes/mechanisms occur simultaneously to the fractions of labile P (Pi and Po); they tend to be positively related to organic C and total N as shown in Tables 3 and 4.
In structural models consisting of interconnections between the P cycle and soil physicochemical properties for soils with high OM content (models C—Fig. 5, C1—Fig. 6), the PCP pool, as an exogenous variable, affected differently each P pool. This variable had a direct and positive influence on stable P pool, but negative on labile P pool. Therefore, the PCP pool has a P sink action on labile P pool, retaining strongly the Pi and stabilizing the Po and, consequently, maintaining the stable P fraction. However, the PCP pool overall effect on labile P pool was distinct for each model, being almost null in model C (β = −0.33 + (0.52 × 0.73) = 0.05), and negative in C1 (β = −0.66 + (0.58 × 0.54), assigning to this variable a P sink function on the labile P pool. The addition of Histosols into model C1 has stressed this sink function, since P immobilization in such soil be related to highly stable organic materials as lignin and organometallic complexes (Yang and Post 2011). Furthermore, this soil type has low contents of remaining P (Prem), thus signaling a high P buffer capacity (Oliveira et al. 2014). Both models imply that several P forms at various liability levels added to maintain bioavailable P supplies; however, it was regulated by the P buffer capacity of each soil.

Model C1, structural model for the P cycle in soils with high organic matter content. All measured variables (in boxes) are represented as effect indicators associated with latent variables (in circles). The numbers correspond to the standardized parameters estimated (P < 0.001) and the R2 values (numbers in bold). †, * and **significant at P < 0.10, 0.05 and 0.01 respectively. Error variables (ε1–ε5, δ1–δ3) are standardized values. Model χ 2 = 18.952, df = 16, P = 0.271. (n = 38)
In all adjusted models, both direct and indirect effects (via pools of labile P and OM), being the stable P pool a source of P to the available P pool, can be attributed to dissolution of stable forms of Pi and/or mineralization of recalcitrant forms of Po. In non-fertilized soils, P availability is highly dependent on mechanisms undertaken by plant roots and soil organisms for accessing the organic and inorganic sources of P (Gama-Rodrigues et al. 2014). The extrusion of organic anions may enhance the bioavailability of Pi by desorption of orthophosphate from sparingly-available sources in soil by ligand exchange reactions (Hinsinger 2001; Richardson et al. 2011). These organic anions can also increase Po solubility facilitating hydrolysis, once they raise the solubility of Po substrate (Reed et al. 2011). It is noteworthy mention that in soils poor in P sources, the proliferation of free-living microorganisms and symbiotic associations with mycorrhizal fungi may mobilize and mineralize Po from other unavailable forms, besides solubilizing Pi (Nash et al. 2014). Therefore, the structural models suggest that soil organic matter is a key factor for increasing the P availability in low fertility tropical soils. Thus, the development of soil organic matter conservation practices in agricultural systems such as no tillage practices, the incorporation of trees, and the continuous input of organic matter are important to improve organic pools of P and therefore reduce de phosphate fertilization in different production systems.
By comparing the models B1 and C, which were analyzed with the same database (Table 2, n = 28), we found that even the AIC as the χ 2/df indexes were lower in model B1; therefore, model B1 was statistically better adjusted to the observed data (Table 5). Meanwhile, the unexplained variance (17–50%) of available P pool in all proposed models indicates that non-measured additional factors might have influenced P availability (Figs. 3, 4, 5, 6), as well as it is still necessary to improve the fit of the regression coefficients in the structural relationship of these models. Further studies are still needed to improve fit models and to recover the full basic structural model (Model A—Fig. 1) for soils of differing pedogenesis. However, it must be considered the different plant mechanisms to extract various forms of P (Turner 2008; Steidinger et al. 2015). Thus, the P use efficiency may be maximized in low-input production systems or even natural ecosystems in the tropics.
Conclusions
The proposed structural models were consistent with the hypothesis of dependence between the pools of P and organic matter as well as physicochemical properties in tropical soils. The pool of labile P was the main source of P for the pool of available P in soils of low to medium organic matter content. In contrast, in soils with high levels of organic matter, the pool of available P was heavily dependent on the stable P pool. On the other hand, the structural models consisting of interconnection between the P cycle and soil physicochemical properties (PCP) were only adjusted in soils with high organic matter content. The pool of PCP acted as P sink for the labile P pool and, in turn, had a positive effect in stable P maintenance. Yet, both P pools acted as sources of P for the available P pool. All structural models evidenced that various forms of P in different levels of lability could contribute in keeping the supply of bioavailable P, yet its magnitude would be regulated by P buffer capacity of each soil.
References
Agbenin JO, Iwuafor ENO, Ayuba B (1999) A critical assessment of methods for determining organic phosphorus in savanna soils. Biol Fertl Soils 28:177–181. doi:10.1007/s003740050481
Bais HP, Weir TL, Perry LG, Gilroy S, Vivanco JM (2006) The role of root exudates in rhizosphere interactions with plants and other organisms. Annu Rev Plant Biol 57:233–266. doi:10.1146/annurev.arplant.57.032905.105159
Bollen KA, Noble MD (2011) Structural equation models and the quantification of behavior. Proc Natl Acad Sci 108:15639–15646. doi:10.1073/pnas.1010661108
Bowman RA (1989) A sequential extraction procedure with concentrated sulfuric acid and diluted base for soil organic phosphorus. Soil Sci Soc Am J 53:326–366. doi:10.2136/sssaj1989.03615995005300020008x
Bowman RA, Cole CV (1978) Transformations of organic phosphorus substrates in soils as evaluated by NaHCO3 extraction. Soil Sci 125:49–54. doi:10.1097/00010694-197801000-00008
Byrne BM (1994) Structural equation modeling with EQS and EQS/windows: basic concepts, applications and programming. Sage Publications, Thousand Oaks
Byrne BM (2009) Structural equation modeling with AMOS: basic concepts, applications and programming, 2nd edn. Taylor and Francis, Routledge
Celi L, Barberis E (2005) Abiotic stabilization of organic phosphorus in the environment. In: Turner BL, Frossard E, Baldwin D (eds) Organic phosphorus in the environment. CABI Publishing, Wallingford, pp 113–132. doi:10.1079/9780851998220.0000
Condron LM, Tiessen H (2005) Interactions of organic phosphorus in terrestrial ecosystems. In: Turner BL, Frossard E, Baldwin D (eds) Organic phosphorus in the environment. CABI Publishing, Wallingford, pp 295–307. doi:10.1079/9780851998220.0000
Condron LM, Moir JO, Tiessen H, Stewart JWB (1990) Critical-evaluation of methods for determining total organic phosphorus in tropical soils. Soil Sci Soc Am J 54:1261–1266. doi:10.2136/sssaj1990.03615995005400050010x
Costa MG, Gama-Rodrigues AC, Gonçalves JLM, Gama-Rodrigues EF, Sales MVS, Aleixo S (2016) Labile and non-labile fractions of phosphorus and its transformations in soil under eucalyptus plantations, Brazil. Forests 7:1–15. doi:10.3390/f7010015
Cunha GM, Gama-Rodrigues AC, Costa GS, Velloso ACX (2007) Organic phosphorus in soils under montane forest, pasture and eucalypt in the North of Rio de Janeiro State, Brazil. Rev Bras Ciênc Solo 31:667–672. doi:10.1590/S0100-06832007000400007
Duda GP (2000) Conteúdo de fósforo microbiano, orgânico e biodisponível em diferentes classes de solos. Tese (Thesis), Universidade Federal Rural do Rio de Janeiro, Itaguaí
Eisenhauer N, Bowker MA, Grace JB, Powell JR (2015) From patterns to causal understanding: structural equation modeling (SEM) in soil ecology. Pedobiologia 58:65–72. doi:10.1016/j.pedobi.2015.03.002
Fornell C, Larcker DF (1981) Evaluating structural equation with unobservable variables and measurement error. J Mark Res 18:39–50. doi:10.2307/3151312
Gama-Rodrigues AC, Sales MVS, Silva PSD, Comerford NB, Cropper WP, Gama-Rodrigues EF (2014) An exploratory analysis of phosphorus transformations in tropical soils using structural equation modeling. Biogeochemistry 118:453–469. doi:10.1007/s10533-013-9946-x
Grace JB (2006) Structural equation modeling and natural systems. Cambridge University Press, New York
Grace JB, Scheiner SM, Schoolmaster DR Jr (2015) Structural equation modeling: building and evaluating causal models. In: Fox GA, Negrete-Yankelevich S, Sosa VJ (eds) Ecological statistics: contemporary theory and application. Oxford University Press, Oxford, pp 168–197. doi:10.1093/acprof:oso/9780199672547.001.0001
Grierson PF, Smithson P, Nziguheba G, Radersma S, Comerford NB (2004) Phosphorus dynamics and mobilization by plants. In: Noordwisk M, Cadisch G, Ong CK (eds) Below-ground interactions in tropical agroecosystems: concepts and models with multiple plant components. CABI International, Wallingford, pp 127–142. doi:10.1079/9780851996738.0000
Guerra JGM, Almeida DLD, Santos GDA, Fernandes MS (1996) Conteúdo de fósforo orgânico em amostras de solo. Pesq Agropec Bras 31:291–299
Guppy CN, Menzies NW, Moody PW, Blamey FPC (2005) Competitive sorption reactions between phosphorus and organic matter in soil: a review. Aust J Soil 43:189–202. doi:10.1071/SR04049
Hair JF, Anderson RE, Tatham RL, Black WC (2009) Multivariate data analysis. Prentice-Hall Inc=, Upper Saddle River
Hinsinger P (2001) Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant Soil 237:173–195. doi:10.1023/A:1013351617532
Iacobucci D (2010) Structural equations modeling: fit Indices, sample size, and advanced topics. J Consum Psychol 20:90–98. doi:10.1016/j.jcps.2009.09.003
Johnson AH, Frizano J, Vann DR (2003) Biogeochemical implications of labile phosphorus in forest soils determined by the Hedley fractionation procedure. Oecologia 135:487–499. doi:10.1007/s00442-002-1164-5
Kenny DA (2014) Measuring model fit. http://davidakenny.net/cm/fit.htm
Marklein AR, Houlton BZ (2012) Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol 193:696–704. doi:10.1111/j.1469-8137.2011.03967.x
McGill WB, Cole CV (1981) Comparative aspects of cycling of organic C, N, S and P through soil organic matter. Geoderma 26:267–286
Nash DM, Haygarth PM, Turner BL, Condron LM, McDowell RW, Richardson AE, Watkins M, Heaven MW (2014) Using organic phosphorus to sustain pasture productivity: a perspective. Geoderma 221–222:11–19. doi:10.1016/j.geoderma.2013.12.004
Nasto MK, Alvarez-Clare S, Lekberg Y, Sullivan BW, Townsend AR, Cleveland CC (2014) Interactions among nitrogen fixation and phosphorus acquisition strategies in lowland tropical rain forests. Ecol Lett 17:1282–1289. doi:10.1111/ele.12335
Olander L, Vitousek P (2000) Regulation of soil phosphatase and chitinase activity by N and P availability. Biogeochemistry 49:175–191. doi:10.1023/A:1006316117817
Oliveira RI, Gama-Rodrigues AC, Gama-Rodrigues EF, Zaia FC, Pereira MG, Fontana A (2014) Organic phosphorus in diagnostic surface horizons of different Brazilian soil orders. Rev Bras Ciênc Solo 38:1411–1420. doi:10.1590/S0100-06832014000500006
Reed SC, Townsend AR, Taylor PG, Cleveland CC (2011) Phosphorus cycling in tropical forests growing on highly weathered soils. In: Bünemann EK, Oberson A, Frossard E (eds) Phosphorus in action: Biological processes in soil phosphorus cycling. Springer, Berlin, pp 339–369. doi:10.1007/s978-3-642-15271-9
Richardson AE, Lynch JP, Ryan PR, Delhaize E, Smith FA, Smith SE, Harvey PR, Ryan MH, Veneklaas EJ, Lambers H, Oberson A, Culvenor RA, Simpson RJ (2011) Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil 349:121–156. doi:10.1007/s11104-011-0950-4
Rita JCDO, Gama-Rodrigues AC, Gama-Rodrigues EF, Zaia FC, Nunes DAD (2013) Mineralization of organic phosphorus in soil size fractions under different vegetation covers in the north of Rio de Janeiro. Rev Bras Ciênc Solo 37:1207–1215. doi:10.1590/S0100-06832013000500010
Sales MVS, Gama-Rodrigues AC, Comerford NB, Cropper WP, Gama-Rodrigues EF, Oliveira PHG (2015) Respecification of structural equation models for the P cycle in tropical soils. Nutr Cycl Agroecosyst 102:1–16. doi:10.1007/s10705-015-9706-5
Sato S, Comerford NB (2006) Organic anions and phosphorus desorption and bioavailabilty in a humid Brazilian Ultisol. Soil Sci 171:695–705. doi:10.1097/01.ss.0000228043.10765.79
Souza MF, Soares EMB, Silva IR, Novais RF, Silva MFO (2014) Competitive sorption and desorption of phosphate and citrate in clayey and sandy loam soils. Rev Bras Ciênc Solo 38:1153–1161. doi:10.1590/S0100-06832014000400011
Steidinger BS, Turner BL, Corrales A, Dalling JW (2015) Variability in potential to exploit different soil organic phosphorus compounds among tropical montane tree species. Func Ecol 29:121–130. doi:10.1111/1365-2435.12325
Tiessen H, Stewart JWB, Cole CV (1984) Pathways of phosphorus transformations in soils of differing pedogenesis. Soil Sci Soc Am J 48:853–858. doi:10.2136/sssaj1984.03615995004800040031x
Treseder K, Vitousek P (2001) Effects of soil nutrient availability on investment in acquisition of N and P in Hawaiian rain forests. Ecology 82:946–954. doi:10.1890/0012-9658(2001)082[0946:EOSNAO]2.0.CO;2
Turner BL (2008) Resource partitioning for soil phosphorus: a hypothesis. J Ecol 96:698–702. doi:10.1111/j.1365-2745.2008.01384.x
Turner BL, Frossard E, Baldwin DS (2005) Organic phosphorus in the environment. CABI Publishing, Wallingford. doi:10.1079/9780851998220.0000
Vitousek PM, Porder S, Houlton BZ, Chadwick OA (2010) Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen–phosphorus interactions. Ecol Appl 20:5–15. doi:10.1890/08-0127.1
Walker TW, Syers JK (1976) The fate of phosphorus during pedogenesis. Geoderma 15:1–19. doi:10.1016/0016-7061(76)90066-5
Wiseman CLS, Pütmann W (2006) Interactions between mineral phases in the preservation of soil organic matter. Geoderma 13:109–118
Yang X, Post WM (2011) Phosphorus transformation as a function of pedogenesis: a synthesis of soil phosphorus data using Hedley fractionation method. Biogeosciences 8:2907–2916. doi:10.5194/bg-8-2907-2011
Zaia FC, Gama-Rodrigues AC, Gama-Rodrigues EF (2008a) Soil phosphorus forms under leguminous tree species, secondary forest and pasture in Northern Rio de Janeiro State, Brazil. Rev Bras Ciênc Solo 32:1191–1197. doi:10.1590/S0100-06832008000300027
Zaia FC, Gama-Rodrigues AC, Gama-Rodrigues EF, Machado RCR (2008b) Organic phosphorus in soils under cocoa agrosystems. Rev Bras Ciênc Solo 32:1987–1995. doi:10.1590/S0100-06832008000500020
Zaia FC, Gama-Rodrigues AC, Gama-Rodrigues EF, Moço MKS, Fontes AG, Machado RCR, Baligar VC (2012) Carbon, nitrogen, organic phosphorus, microbial biomass and N mineralization in soils under cacao agroforestry systems in Bahia, Brazil. Agrofor Syst 86:197–212. doi:10.1007/s10457-012-9550-4
Acknowledgements
We thank the Rio de Janeiro Research Foundation (FAPERJ) for the award of a research fellowship to Antonio Carlos Gama-Rodrigues. We also thank the Brazilian National Council for Scientific and Technological Development (CNPq)—Grant 2013/475222-0 (Universal Project) for its financial support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Sales, M.V.S., Aleixo, S., Gama-Rodrigues, A.C. et al. Structural equation modeling for the estimation of interconnections between the P cycle and soil properties. Nutr Cycl Agroecosyst 109, 193–207 (2017). https://doi.org/10.1007/s10705-017-9879-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10705-017-9879-1