
Abstract
An experiment was conducted at Msekera Research Station from 1998 to 2002 to determine effect of planted fallows of tree species mixed with herbaceous legumes and those of single species fallows on biomass production, soil properties and subsequent maize grain yields. After 2 years of fallow the highest total above ground biomass of 9.2 t ha−1 was recorded in sole Gliricidia. Pre-season soil total inorganic N in 0–20 cm soil depth under tree fallows was highest in Sesbania + Macrotyloma axillare mixture. Sesbania + M. axillare mixture had the highest cumulative water intake at fallow clearing. Among the 7 mixtures and 3 sole species treatments, Sesbania + M. axillare mixture produced the highest maize yield of 2.7 and 1.9 t ha−1 in 2001 and 2002 season, respectively, after 2 years of fallow. The results of the study indicate the complementarity of herbaceous legumes and tree mixtures in improving soil quality. We conclude that mixed species have the potential to increase the amount and quantity of total N added to the soil and increase nutrient cycling. However, maize grain yields were not significantly improved as compared with single species due to low biomass production of the trees. The coiling nature and the added labour requirements for weed control would limit the use of these mixtures by smallholder farmers who are labour constrained. Agroforestry trees and non-twining legumes could be promoted instead.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Soil fertility depletion is a major constraint for the resource poor farmers in most sub-Sahara African countries. High population pressure has led to land shortages for traditional fallowing and eventually decline in soil fertility (Buresh et al. 1997). Resource poor farmers who previously used inorganic fertilizers can no longer afford to due to escalating costs (Gladwin 1992; Brussaard et al. 2007). The use of fast-growing nitrogen-fixing legumes such as sesbania (Sesbania sesban (L.) Merr.), and tephrosia (Tephrosia vogelii Hook. f.) in an improved fallow system has been reported to provide nutrient inputs that increases yields of subsequent maize and produce fuel wood (Kwesiga et al. 1994; Jama et al. 1998). However, the residual effect of these fallows on subsequent crop declines rapidly after 1 or 2 years of cropping. One of the key constraint to adoption of these technologies is the labour demand in weeding the fallows especially in the first year of establishment. Continuous cultivation has also been reported by several researchers (Lal 1985; Wilkinson 1975) as being responsible for structural degradation of soil leading to soil compaction. Soil compaction decreases the number of larger pores and hence affects soil moisture contents at higher potentials. Research on improved fallows in southern Africa has concentrated on single species of tree legumes such as sesbania, tephrosia and Gliricidia [Gliricidia sepium (Jacq.) Walp.]. The wide spread use of single species such as Sesbania in improved fallows in eastern Zambia has led to disease or pests outbreaks such as the defoliating beetle (Mesoplatys ochroptera), a serious pest of Sesbania (Kwesiga et al. 1999). Sileshi et al. (2000) reported that there is an ecological danger in relying on single species fallows in terms of pests and diseases. Sesbania is also susceptible to nematodes, particularly the root-knot nematodes (Meloidogyne incognita and Meloidogyne javanica), which cause severe stunting and death of trees. Single species fallows may also be inefficient in terms of resource capture (water, light and nutrients). This would result in productivity loss should the single species be affected by drought, for instance. In Western Kenya mixed fallows of species that differ in growth habit have been shown to be more efficient in resource capture, control of weeds, reduced incidence of nematodes (Dasaeger and Rao 2000) and lead to higher residual effects on maize yield (Gathumbi 2000). Short-lived residual effects and ineffective weed suppression in planted fallows of single species such as Sesbania sesban and Tephrosia vogelii have been raised as being among limiting factors for adoption under resource poor farming communities. However, there has been little work on mixed fallows using tree and herbaceous legumes in southern Africa. Therefore, the objective of our study was to determine the residual effect of tree fallow species when mixed with herbaceous legumes on biomass production, soil chemical and physical properties and maize grain yield.
Materials and methods
Site description
The study was conducted at Msekera Research Station from 1998 to 2002. The station lies at an altitude of 1030 m above sea level, longitude of 32°34′E and latitude 13°38′S (Commissaris 1975). The soils at Msekera are Ferric luvisols which are moderately deep, dark reddish brown, moderately structured, slightly eroded, well drained clayey soil which is sandy loam in the top 20 cm (FAO-UNESCO (1998). The surface soil (0–20 cm) consists of <1 % carbon content, pH (CaCl2) of 4.8, 14.4 % clay, 65.6 % sand. The average unimodal (during the cropping season) rainfall is 1092 mm. Twelve land-use systems (LUSs) that were compared were sole sesbania, sole Tephrosia, sole Gliricidia, sesbania + Velvet bean [Mucuna pruriens (L.) DC], Sesbania + archer dolichos [Macrotyloma axillare (E. Mey.) Verde], Tephrosia + velvet bean, Tephrosia + archer dolichos, gliricidia + velvet bean, Gliricidia + archer dolichos, Sesbania + Tephrosia, natural fallow and sole maize (Zea mays L.) with and without fertilizer. The treatments were arranged in randomized complete blocks design (RCBD) with three replicates. The gross plots size was 10 × 10 m. Sesbania (prov. Chipata dam) and Gliricidia (prov Ratalhuleu) were planted in the field from nursery raised bare rooted seedlings at the age of 5 and 8 weeks, respectively. The spacing between plants was 1.0 × 1.0 m (10,000 trees ha−1). While Tephrosia (prov. Ex-misamfu) seeds and herbaceous legumes of archer dolichos and velvet beans were planted directly in the field at the same time of transplanting the trees. They were planted at spacing of 1.0 × 0.50 m (20,000 trees ha−1) for tephrosia and 1.0 × 0.25 m (40,000 plants ha−1) for velvet beans and archer dolichos. At 24 months of fallow litter was assessed from three 2.0 × 1.0 m quadrants in the net plots. Trees were clear felled at ground level in November 2000 after 2 years of fallow. Stumps and root systems were left un uprooted.
Above ground biomass
Total above ground biomass of trees (leaves, twigs and wood) was measured at fallow clearing by separating the biomass components into foliage (leaves and twigs), branches and stems. These components were then weighed as fresh biomass after which samples of each component were collected on plot basis and oven dried at 70 °C to equilibrium moisture content. Surface litter was measured from three quadrants of 2 m2 in the net plot at fallow clearing. This data was used to estimate dry weight on plot basis and extrapolated to a hectare basis. The tree mulch and herbaceous legumes were incorporated in the soil by hand hoeing. After the fallow, hybrid maize (Zea mays variety MM 604) was sown by hand at 25 cm within-row and 100 cm between-row spacing (44,444 plant ha−1). Fertilizer was applied only to the fertilized control plots at the recommended rates of 20, 18 and 17 kg K ha−1 of Compound-D (basal dressing) at sowing and 92 kg N ha−1 as urea (top dressing) at 4 weeks after sowing (WAS). All the plots were manually cultivated at the beginning of each season and weeded twice during the crop season using hand-hoes.
Nitrogen dynamics
Soil samples were taken from six positions in the net plot by systematically sampling along the diagonals at fallow clearing (pre-season, November 2000) using a metal sampler (4.2 cm diameter G. I. Pipe) from 0–20, 20–40, 40–60 and 60–100 cm soil depths at Msekera. Soil samples were air-dried on the same day that samples were extracted. This was to avoid loss of carbon due to ongoing chemical and biochemical processes when moist soil samples are stored in warm conditions. Once air-drying was complete any visible organic debris in the soil including leaves, stalks and roots were removed. Each air-dried soil sample was weighed to obtain the total air-dry mass of the sample. The soils were then sieved before analysis. The soils were determined for ammonium-N by colorimetric method (Anderson and Ingram 1993) and nitrate–N concentrations were determined by cadmium reduction (Dorich and Nelson 1984). The sum of inorganic ammonium-N and inorganic nitrate–N constituted the total inorganic-N.
Bulk density and pore size distribution
Bulk density (ρb) and pore size distribution was calculated from undisturbed triplicate core (100 cm3) soil samples from 0 to 10 cm soil layer at fallow clearing (November 2000).
Soil water dynamics
Infiltration and cumulative water intake
Infiltration and cumulative water intakes were monitored at fallow clearing (October 2000) and after first cropping (October 2001) at Msekera during the dry season in 8 treatments only due to limitation of the equipment. The Double Ring Infiltrometer method by Bouwer (1986) was used to measure infiltration rates and cumulative water intakes. Water measurements were recorded for 3 h at 0, 5, 10, 15, 20, 30, 45, 60, 90, 120, 150 and 180 min intervals. The average readings were used to calculate infiltration and cumulative water intake per plot using the Kostiakov (1932) and Philips (1957) models.
Volumetric water content
A calibrated neutron probe (Belcher et al. 1950) was used to measure volumetric soil–water content in 10 cm increments to a depth of 100 cm bi-weekly from all plots at fallow clearing and during the cropping phase. One aluminum access tube was installed in the center of the 10 × 10 m plot. Neutron probe calibration was based on simultaneous probe readings and gravimetric measurements. Soil water storage on 19 December 2000 and 11 April 2001 in the 1.0 m soil profile was calculated by summing the water storage in the 0–100 cm soil profile. The measurements in the top 20 cm of the soils were determined gravimetrically and converted into volumetric values using the measurements of bulk density. This is because the sphere of influence of the neutron probe would extend into the air above the soil surface.
Data analysis
All data are presented as means of three replicates with standard error of difference (s.e.d.). Differences between treatments were tested with General Analysis of variance (ANOVA) using GENSTAT version 5 (Genstat 5 committee 1988). Simple linear regressions were used to determine the relationship between maize grain yield and pre-season inorganic nitrate-N and total-N for different soil depths. Maize grain yield was determined in the net plot, with grain yield adjusted to 12.5 % moisture content and converted to per hectare basis. Where necessary, treatments mean separation test was done using the least significant difference (LSD) method.
Results
Surface litter and weed biomass
At 24 months of fallow, significant differences (P < 0.01) were recorded for weed biomass (Table 1). The highest surface litter of 3.5 t ha−1 was recorded in Gliricidia + Archer dolichos mixture as compared with 1.4 t ha−1 in sesbania + velvet beans (Table 1). On the other hand weed cover of 0.1 t ha−1 was recorded in sole Gliricidia as compared with 3.8 t ha−1 in Sesbania + velvet (Table 1). The highest weed biomass of 7.0 t ha−1 was recorded in natural fallow.
Tree biomass
At fallow clearing the lowest survival of 36 % was in tephrosia + Archer dolichos mixture as compared with 97 % in sole Gliricidia (Table 1). The highest total biomass production of 9.2 t ha−1 was in sole Gliricidia as compared with 4.5 t ha−1 in tephrosia + Archer mixture (Table 1). No significant difference was recorded in seasonal input coppice biomass for the two seasons (Table 2). Despite there being no significant difference in coppice biomass among the three Gliricidia treatments, sole Gliricidia had the highest coppice growth input compared with Gliricidia + Archer dolichos or Gliricidia + velvet beans.
Maize grain yield
Among the 7 mixtures and 3 sole species, Sesbania + Archer dolichos mixture produced the highest maize yield of 2.7 and 1.9 t ha−1 in 2001 and 2002 season, respectively (Table 3). Maize with fertilizer (control) treatment had the highest maize grain yield of 3.7 and 3.1 t ha−1 during 2001 and 2002 season, respectively (Table 3). All the treatments recorded a reduction of maize grain yield in the second year of cropping as compared with the first year grain yields, except for sole Gliricidia that increased yields to 122 % over the maize without fertilizer control (Table 3).
Nitrogen dynamics
Pre-season soil total inorganic-N in 0–20 cm in tree and herbaceous mixture was highest in tree and Archer dolichos mixture than the tree and velvet mixture treatment (Table 4). Sesbania, Gliricidia and Tephrosia mixture with Archer dolichos had higher pre-season total inorganic-N than the sole trees and velvet mixtures.
Relationship between inorganic nitrogen and maize grain yield
Significant (P < 0.01) relationships were found between first year maize grain yield and pre-season inorganic nitrate-N and total-N. The coefficient (r) for the relationship between first year maize grain yield and total inorganic-N was 0.83 for 0–20 cm soil depth. While the coefficient (r) for the relationship between inorganic nitrate–N and grain yield was 0.54 for the same soil depth. On the other hand no relationship was observed for the second year maize yield with pre-season inorganic-N for 2001–02 season.
Bulk density and pore size distribution
Bulk density and pore size distribution was significantly affected by land use system in 0–10 cm soil layer after fallow clearing (Table 5). Bulk density ranged from 1.35 to 1.48 g cm−3 for sole sesbania and Gliricidia + Archer dolichos, respectively. Macropores ranged from 15.4 to 27.3 % for sole Gliricidia and sesbania + Archer dolichos mixture, respectively. Where as mesopores ranged from 14.3 to 30.3 % for sole sesbania and sole Gliricidia, respectively. Micropores ranged from 50.6 to 68.2 % for sesbania + Archer dolichos mixture, respectively.
Soil water dynamics
Cumulative water intake at fallow clearing
Significant differences (P < 0.01) were observed at fallow clearing (October 2000) (Table 6) and after first year of cropping (October 2001) (Table 6) in cumulative water intake. The order of cumulative water intake in October 2000, after 3 h was: Sesbania + Archer dolichos = Sesbania = Sesbania + velvet beans > Gliricidia + Archer dolichos > Gliricidia = Gliricidia + velvet beans = natural fallow = maize + fertilizer. Cumulative water intake in October 2001, after 3 h was in the order of: Gliricidia + velvet beans = natural fallow > Sesbania = maize + fertilizer > Gliricidia + Archer dolichos > Sesbania + Archer dolichos = Gliricidia = Sesbania + velvet beans (Table 6).
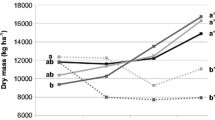
Sorptivity values at fallow clearing were in the order of Sesbania > Sesbania + Archer dolichos > Sesbania + velvet beans > Gliricidia + Archer dolichos > maize + fertilizer > natural fallow > Gliricidia > Gliricidia + velvet beans (Table 7). On the other hand after first year of cropping the order was natural fallow > Gliricidia + velvet beans > Sesbania > maize + fertilizer > Sesbania + Archer dolichos > Gliricidia + Archer dolichos > Sesbania + velvet beans > Gliricidia (Table 8). Water storage in December 2000 was not significantly affected by the different land use system (Fig. 1). In April 2001 soil water storage was in the order of Gliricidia + Archer dolichos > Sesbania + Archer dolichos > Gliricidia > Sesbania > maize + fertilizer and was significantly affected by the different land use system (Fig. 1). The change in water storage varied from −10.4 to −4.0 mm for Gliricidia + Archer dolichos to maize with fertilizer.

Change in water storage after a period of 113 days at Msekera as affected by land use system during 2000–2001 season. Note identical letters indicate that bars are not significantly different at P < 0.05
Discussion
The litter fall obtained in this study appeared low compared to results obtained elsewhere. Torquebiau and Kwesiga (1996) showed that litter fall from a 2 year fallow of sole sesbania can be as much as 4.0 t ha−1. The reason could be that litter fall collected by the quadrant method could be underestimated as what falls during the fallow period might be lost through decomposition or blown away from the plots. Also the poor survival, low biomass and choking of the trees by the herbaceous legumes could have contributed to low litter fall after 2 years of fallow. Similarly, our total above ground biomass yield were lower than those reported elsewhere in eastern Zambia (Kwesiga and Coe 1994; Kwesiga et al. 1994; Chirwa et al. 2000). The low Sesbania biomass could have been due to development of the Sesbania defoliating mesoplatys beetles, which reduce biomass production of Sesbania, as reported by Sileshi and Mafongoya (2002).
Subsequent maize yields were also lower than those reported else where after 2 years of fallow (Kwesiga and Coe 1994; Kwesiga et al. 1994) This reduction in yield was due to excessive rains experienced in 2000/2001 season (1342 mm instead of the usual annual average of 1029 mm), which could have leached most of the nutrients beyond the rooting depth of maize (Mekonnen et al. 1997; Conyers et al. 2012). Also in 2001/2002 season there was erratic rainfall pattern (832 mm instead of the usual annual average of 1029 mm). In general high rainfall, water logging and out break of maize grey leaf spot disease Cercospora zeae-maydes around grain filling stage during the growing season of maize could explain the low maize yield from the fallows and maize with fertilizer. Low biomass production at fallow clearing in the sole tree and tree herbaceous mixture could also have contributed to low maize yields during the first year and second year of cropping. Tree biomass production at fallow clearing has been known to have significant effect to subsequent maize production (Kwesiga and Coe 1994). When species such as Gliricidia are managed as coppicing fallows they tend to have considerable potential in sustaining maize grain yield as seen from the increase in maize yield in year two (Mafongoya et al. 2000). This sustainability aspect comes from the coppices that are continuously cut and incorporated in the soil to replace the lost nutrients in the surface from the subsoil in form of fresh leaf and litter (Bah and Rahman 2001; Murphy et al. 2013).
Maize yields in the first year followed the preseason inorganic total inorganic-N in the soil (0–20 cm soil layer). This is in agreement with the findings of Angus et al. (2006), Maroko et al. (1998) and Ikerra et al. (1999). However, maize yields in the second year did not show any relationship with preseason inorganic nitrate-N. This could be due to factors other than soil fertility such as low rainfall and outbreak of maize grey leaf spot disease, which also negatively affected the yields during the season.
The addition of above ground biomass during fallow phase and improved soil aggregation under sesbania could have lowered the soil bulk density. Bulky density can be used to determine if the soil is too compact to allow root penetration or adequate aeration (McCarty et al. 2016). The lower the bulk density, the easier the roots will penetrate the soil and exploit soil nutrients (Rowell 1994). Torquebiau and Kwesiga (1996) under similar conditions reported improvement in bulk density and penetrometer resistance after 2 years of sesbania fallow. Rowell (1994) classified macropores as those pores, which allow rapid drainage of water after heavy rainfall or irrigation, and once these pores are emptied drainage becomes very slow. On the other hand meso-pores, which conduct water by rapid capillary flow, have pore diameters of between 30 and 100 μm. Micropores, whose diameters are <30 μm, are responsible for moisture retention and slow capillary flow. Therefore, our results indicate that sesbania + Archer dolichos, sole Gliricidia and Gliricidia + Archer dolichos will promote both macro- and meso-pores that allow good root proliferation and rapid drainage of the soil profile. Natural fallow, maize with fertilizer and sole sesbania will promote development of micropores that will enhance more soil moisture storage as supported by the high sorptivity values under sole sesbania. The high cumulative water intake in sesbania + Archer dolichos observed in our study could be due to less runoff, high root mass, less compaction, low bulk density and high soil aggregation (Mafongoya et al. 2000). On the other hand the low water intake in the maize with fertilizer could be linked to continuous cultivation, which destroys soil structure (Lal 1989). Increased soil infiltration after 2 years of Sesbania fallow on a Ferric lixisols at Msekera has also been reported (Torquebiau and Kwesiga 1996). Similarly, fallowing with various legumes and grass cover crops is also known to improve soils infiltrability (Pereira et al. 1958; Lal et al. 1978, 1979). After 1 year of cropping cumulative water intake reduced in all the land use systems. The highest percentage of reduction in cumulative water intake was recorded in sesbania + velvet beans and the lowest being recorded in Gliricidia + velvet beans. Less runoff and water logging was observed under mixtures probably due to improved soil physical properties (Mafongoya et al. 2000). The increase in water storage under mixtures could be associated with high biomass produced during the fallow phase and decomposition of roots of herbaceous legumes and greater improvement in soil bulk density, infiltration and soil aggregation (Sheoran et al. 2010).
However, despite the improvement in soil properties under tree and herbaceous mixtures, the herbaceous legumes of M. axillare and Mucuna pruriens coiled and chocked the trees. Such herbaceous legumes are undesirable for mixing with trees (Dubeux et al. 2015).
Conclusion
The results show that mixed species have the potential to increase the amount of surface litter biomass and total N added to the soil and increase nutrient cycling. Weed suppression by herbaceous legumes in mixtures could be a good indicator for farmers to adopt improved planted fallow technology where labour is a constraint in weeding the fallows. Although Archer dolichos was able to suppress weeds it had negative attributes on associated tree biomass production of choking of the trees due to its coiling nature. Although mixtures with herbaceous legumes and trees improved soil physical properties, maize grain yields were not significantly improved as compared with single species due to low biomass production of the trees. This was attributed to the twining habit of both legumes used, which choked the trees and hence less tree biomass produced. In addition, these mixtures provided a favorable environment for development of the sesbania defoliating mesoplatys beetles, which reduced biomass production of Sesbania. There was also a problem of disentangling herbaceous legume vines that had extensively coiled around stems and branches of tree legumes. It took more time and labour to cut trees in mixtures as compared with tree sole species fallows. Therefore, these mixtures are not recommended for farmers. What would be ideal mixtures are trees with food grain legumes and non-twining herbaceous species such as siratro. The benefits from grain legumes could offset the maize yield loss.
References
Anderson JM, Ingram JSI (1993) Tropical soil biology and fertility: a handbook of methods, 2nd edn. CAB International, Wallingford
Angus JF, Bolger TP, Kirkegaard JA, Peoples MB (2006) Nitrogen mineralisation in relation to previous crops and pastures. Aust J Soil Res 44:355–365
Bah AR, Rahman ZA (2001) Gliricidia (Gliricidia sepium) Green manures as a potential source for maize production in the tropics. In: Optimizing nitrogen management in food and energy production and environmental protection: Proceedings of the 2nd International Nitrogen Conference on Science and Policy. The Scientific World 1
Belcher DJ, Cuykendall TR, Sack HS (1950) The measurement of soil moisture and density by neutron and gamma-ray scattering. Tech. Dev. Rep. 127. Civ. Aeronaut. Admin., Washington, DC
Bouwer H (1986) Intake rate: cylinder infiltrometer. In: Klute A (ed) Methods of soil analysis, part 1, Physical and mineralogical methods—agronomy monograph number, vol 9. American Society Agronomy, Madison, pp 825–844
Brussaard L, deRuiter PC, Broan GG (2007) Soil biodiversity for agricultural sustainability. Agric Ecosyst Environ 121:233–244
Buresh RJ, Sanchez PA, Calhoun FG (eds) (1997). Replenishing soil fertility in Africa SSSA Special Publication no 51: Soil Science Society of America, Madison, WI, USA. Proceedings of an International Symposium
Chirwa TS, Mafongoya PL, Mbewe DNM, Chishala BH (2000) Changes in soil properties and their effects on maize productivity following Sesbania and Pigeonpea Improved Fallow System in eastern Zambia. M.Sc. thesis, University of Zambia
Commissaris ALTM (1975) Detailed Soil Survey of Msekera Research Station. Soil Survey report no. 19
Conyers M, Newton P, Condon J, Poil G, Mele P, Ash G (2012) Three long-term trials end with a quasi-equilibrium between C, N and pH and implication for C sequestration. Soil Res 50:527–535
Dasaeger J, Rao MR (2000) Parasitic nematode populations in natural fallows and improved cover crops, and their effects on subsequent crops in Kenya. Field Crops Res 65:41–56
Dorich RA, Nelson DW (1984) Evaluation of manual cadmium reduction methods for determination of nitrate in potassium chloride extracts of soil. Soil Sci Soc Am J 48:72–75
Dubeux JC Jr, Muir JP, Nair PR, Sollenberger LE, Silva HM, de Mello AC (2015) The advantages and challenges of integrating tree legumes into pastoral systems. In: Proceedings of the 1st international conference on forages in warm climates, Lavras, Brazil, pp 141–164
FAO-UNESCO (1998) Soil Map of the World (Revised Legend), Rome
Gathumbi SM (2000) Nitrogen sourcing by fast-growing legumes in pure and mixed species fallows in western Kenya. Ph.D thesis, Wye College, University of London, Wye, Kent, UK
Genstat 5 Committee (1988) Genstat 5 reference manual. Oxford University Press, Oxford
Gladwin C (1992) Gender impacts of fertilizer subsidy removal program in Malawi and Cameroon. Agric Econ 7:141–153
Ikerra S, Maghembe J, Smithson P, Buresh R (1999) Soil nitrogen dynamics and relationships with maize yields in gliricidia–maize intercrop in Malawi. Plant Soil 211:155–164
Jama B, Buresh RJ, Place FM (1998) Sesbania tree fallows on phosphorus-deficient sites: maize yield and financial benefit. Agron J 90:717–726
Kostiakov AN (1932) On the dynamics of the coefficient of water-percolation in soils and on the necessity for studying it from a dynamic point of view for purposes of amelioration. In: Trans. Sixth International. Society of Soil Science, Russian Part A, pp 17–21
Kwesiga F, Coe R (1994) Effect of short rotation Sesbania sesban planted fallows on maize yield. For Ecol Manag 64:199–208
Kwesiga F, Phiri D, Simwanza CP, Mwanza S (1994) Zambia/ICRAF Agroforestry Research Project annual Report. Afrena Report no. 87
Kwesiga F, Franzel S, Place F, Phiri D, Simwanza CP (1999) Sesbania sesban improved fallows: their inception, development and farmer enthuasm. Agrofor Syst 47:49–66
Lal R (1985) Mechanized tillage systems effects on properties of a tropical Alfisol in watersheds cropped to maize. Soil Tillage Res 6:149–161
Lal R (1989) Agroforestry systems and soil surface management of a tropical alfisol: V. Water infiltrability, trasmissivity and soil water sorptivity. Agrofor Syst 8:217–238
Lal R, Wilson GF, Okigbo BN (1978) No-tillage farming after various grasses and leguminous cover crops in tropical Alfisols. I. Crop performance. Field Crops Res 1:71–84
Lal R, Wilson GF, Okigbo BN (1979) Changes in properties of an Alfisol produced by various crop covers. Soil Sci 127:377–382
Mafongoya PL, Katanga R, Mkonda A, Chirwa TS, Chintu R, Matibini J (2000) Zambia/ICRAF Agroforestry Research Project, 2000 Annual Report
Maroko JB, Buresh RJ, Smithson PC (1998) Soil nitrogen availability as affected by fallow-maize systems on two soils in Kenya. Biol Fertil Soils 26:229–234
McCarty LB, Hubbard LR, Quisenberry V (2016) Applied soil physical properties, drainage, and irrigation strategies. Springer International Publishing, Cham, pp 1–72. doi:10.1007/978-3-319-24226-2_1
Mekonnen K, Buresh RJ, Jama B (1997) Root and inorganic nitrogen distributions in sesbania fallow, natural fallow and maize fields. Plant Soil 188:319–327
Murphy BW, Crawford MH, Duncan DA, McKenzie DC, Koen TB (2013) The use of visual soil assessment schemes to evaluate surface structure in a soil monitoring program. Soil Use Manag 127:3–12
Pereira HC, Wood RA, Brozostowski HW, Hosegood PA (1958) Water conservation by fallowing in semi-arid tropical East Africa. Emp J Exp Agric 26:203–228
Philips JR (1957) The theory of infiltration: the infiltration equation and its solution. Soil Sci 83:345–357
Rowell DL (1994) Soil science, methods and application. Longman Group UK Limited, Harlow
Sheoran V, Sheoran AS, Poonia P (2010) Soil reclamation of abandoned mine land by revegetation : a review. Int J Soil Sediment Water 3(2):1–21
Sileshi G, Mafongoya PL (2002) Incidence of Mesoplatys ochroptera Stal (Coleoptera: Chrysomelidae) on Sesbania sesban in pure and mixed species fallows in eastern Zambia. Agrofor Syst 56:225–231
Sileshi G, Maghembe JA, Rao MR, Ogol CKPO, Sithanantham S (2000) Insects feeding on Sesbania species in natural stands and agroforestry systems in southern Malawi. Agrofor Syst 49:41–52
Torquebiau EF, Kwesiga F (1996) Root development in Sesbania sesban fallow-maize system in eastern Zambia. Agrofor Syst 34:193–211
Wilkinson GE (1975) Effect of grass fallow rotations on the infiltration of water into a savanna zone soil of northern Nigeria. Trop Agric 52:97–103
Acknowledgments
We thank the Swedish International Development Agency (SIDA) and the Government Republic of Zambia (GRZ) for their financial support to this research. We are grateful to Messers K. Linyunga, S. Chikale, and P. Phiri for field data collection, laboratory analysis and data analysis respectively. Dr. G. Sileshi is also acknowledged for his useful comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Mafongoya, P.L., Jiri, O. Soil nitrogen and physical properties and maize yields after mixed planted fallows of tree and herbaceous legumes. Nutr Cycl Agroecosyst 105, 75–84 (2016). https://doi.org/10.1007/s10705-016-9776-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10705-016-9776-z