
Abstract
Nitrogen (N) is the most important yield-limiting factor in agricultural systems, however, N application can lead to emissions and environmental problems such as global warming (N2O) and groundwater contamination (NO3 −). This study analyses the N balance, nitrogen-use efficiency, and N loss potential of conventional farming systems (arable farming, improved arable farming, and agroforestry) and organic farming systems (mixed farming, arable farming, and agroforestry) based on long-term field experiments in southern Germany. The effects of the conversion of farm structure and N management are identified. The conventional farming systems in this study were high N-input and high N-output systems. The conventional arable farming system had the lowest nitrogen-use efficiency and the highest N surplus. An optimised N management and the use of high-yielding crop varieties improved its nitrogen-use efficiency. The establishment of conventional agroforestry resulted in the reduction of N input, N output and N surplus, while maintaining high yields. The organic mixed farming system is characterised by a relatively high N input and N output, the accumulation of soil organic nitrogen, the highest nitrogen-use efficiency, and the lowest N surplus of all analysed systems. These good results can be attributed to the intensive farm N cycle between soil–plant–animal. The shift from organic mixed farming to organic arable farming system extensified the N cycle, reduced N input, crop yield and N output. The change from organic arable farming to organic agroforestry reduced the N input, increased the biomass yield, and remained the N surplus within an optimal range.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Highlights
-
1.
A comprehensive N balance method was adapted to agroforestry systems.
-
2.
We quantify the effects of changing farm structures on the nitrogen-use efficiency.
-
3.
Organic mixed farming has the highest nitrogen-use efficiency of all farming systems.
-
4.
Agroforestry can improve nitrogen-use efficiency and reduce N surpluses.
Introduction
Nitrogen (N) is the most important yield-limiting factor in agricultural systems (Tilman 1999). It is mainly supplied by mineral fertilisers, farm manure, symbiotic N2 fixation, and atmospheric wet and dry deposition. Stewart et al. (2005) reported that at least 30–50 % of the crop yield is attributed to the application of mineral fertiliser. However, when the applied N is not completely taken up by plants nor accumulated as soil organic N (SON), it results in emissions and associated environmental problems such as global warming (N2O), groundwater contamination (NO3 −), air pollution and eutrophication (NH3), and loss of biodiversity (Crutzen et al. 2008; European Environmental Bureau 2013; Ju et al. 2006; Millar et al. 2010; Sutton et al. 2011b). A recent study pointed out that the threats posed by N emissions cost the European Union more than double the value which N fertiliser adds to European farm income (Sutton et al. 2011a, b). The high energy consumption and greenhouse gas emissions generated by the production and use of mineral fertiliser (Dawson and Hilton 2011; Hülsbergen et al. 2001; Snyder et al. 2009) are also threats to the environment and society.
N-management and N-balance in conventional and organic farming
Both high N-input systems (e.g. conventional farming) and low N-input systems (e.g. organic farming) are found in Western Europe. Spiertz (2010) suggested that the focus in high N-input systems should be to improve the efficiency of applied N (more yield with less fertiliser N), while in low-input systems additional N input is required to increase yield level and yield stability. In Germany, the high N input and N surplus of conventional farms are mainly from the use of high doses of mineral N, often in combination with organic N (e.g. slurry, biogas slurry). The mean N surplus of Germany at farm-gate scale is about 100 kg N ha−1 yr−1 (Taube et al. 2013; Umweltbundesamt 2014). However, whether a significant reduction of N surplus without negative yield effects is possible has not been studied comprehensively.
The most important N sources in organic farming are green and animal manure and symbiotic N2 fixation. These N sources can be combined in mixed farming systems perfectly: N-fixing crops (e.g. clover and alfalfa) could be harvested as fodder for animals, and the animal manure will be returned to the soil as an N source, therefore the N cycle is nearly closed and N losses are minimised (Küstermann et al. 2010). However, current policies and economic benefits encourage European farmers to enlarge and specialise their farms (Ryschawy et al. 2012) and consequently reduce the amount of both organic and conventional mixed farms. According to a survey done in the agrarian region north of Munich, southern Germany, 40 % of the organic farms are managed as specialised arable farming systems (AF) (Vockinger 2013) and this number is still increasing. However, this change may affect the farm’s nitrogen cycle, soil fertility, and yield potential (Van Keulen and Schiere 2004). The long-term effects of the shift from organic mixed farming to specialised organic arable farming systems are so far not being looked into, therefore leaves a research gap in this field.
N-management and N-balance of agroforestry systems
Agroforestry systems (AGFS) are multifunctional land use systems with trees incorporated into agricultural crop and/or animal production. The interaction between woody and non-woody components benefits these systems economically, ecologically, environmentally and socially (Jose 2009; Reynolds et al. 2007).
The combination of short-rotation trees for energy wood production and agricultural crops is one of the modern forms of AGFS gaining most of the interest in Germany (Johann-Heinrich von Thünen-Institut 2012; Nerlich et al. 2013). Even though this farming system is still not widespread in Germany, the establishment of AGFS could help to reduce the N surplus and N emissions of farming systems because (1) the N demand of short-rotation trees is generally less than that of agricultural crops (Dawson 2007; Musshoff 2012; Sevel et al. 2014), thus the external N inputs as well as the N surplus of the whole AGFS system can be reduced, and (2) it has the potential to optimise the nitrogen-use efficiency (NUE) of the system due to its ability to enhance soil quality and nutrient uptake (Bambrick et al. 2010; Ilany et al. 2010; Isaac et al. 2007; Patra 2013). However, the effects on yields, nitrogen-use efficiency, and soil fertility of N-limited organic farms through the establishment of AGFS have not yet been analysed to the authors’ best knowledge.
Study scope
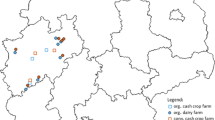
Scheyern Research Farm in southern Germany has been established with different conventional and organic farming systems. Thus, the analysis and comparison of the NUE and nitrogen surplus from different N management strategies under the same soil and climate conditions is possible, using a unique long-term data set (Fig. 1). This article is based on (1) data collected from the organic mixed farming system and conventional arable farming system between 1999 and 2002 (partly been reported by Küstermann et al. 2010, see Tables 1 and 4), and (2) data collected from the organic and conventional arable farming and agroforestry systems from 2009 to 2012 (the results from the first stage of a 20-year long-term agroforestry experiment). Therefore, the effects of the change from organic mixed farming to organic arable farming and organic agroforestry, as well as from conventional arable farming to conventional agroforestry, could be analysed.

The development of farming systems in Scheyern Research Farm
This study determines the NUE of the above-mentioned organic and conventional farming systems by analysing N input, N uptake, N output, accumulation or depletion of soil organic nitrogen (Δ SON), and N surplus. The aims of this study are (1) improving a comprehensive N balance method (Küstermann et al. 2010) in order to make it applicable to mixed farming, arable farming, and agroforestry systems; (2) analysing the effects of changing farm structures and nitrogen management, and identifying the systems with the highest N output, highest NUE, and the lowest N loss potential; and (3) examining whether it is possible to increase the DM yield, optimise the NUE, sustain soil fertility, and reduce N surplus at the same time.
Methodology
Experimental site
The experimental data analysed in this study was collected at the Scheyern Research Farm, 40 km north of Munich in southern Germany. The research farm is located at 445–498 m above sea level in a hilly landscape with soils characterised as loamy to sandy cambisols derived from tertiary sediments partly covered by loess (Schröder et al. 2002). The mean annual precipitation and temperature is 887 mm and 8.3 °C, respectively (Wetterdienst 2012).
The experimental farm was established with an organic mixed farming system (31.3 ha arable fields with crop rotation and 18.2 ha permanent grassland) and a conventional arable farming system (30.4 ha arable fields with crop rotation) in 1992 (Küstermann et al. 2010).
In the organic mixed farming system, a suckler cow herd was kept. The stock density had increased gradually from 0 livestock units ha−1 (in 1992) to 1.4 livestock units ha−1 (averaged from 1999 to 2002). 1999–2002 was the period with the highest production intensity (highest livestock density, highest N input) (Küstermann et al. 2010). The stock numbers of the organic mixed farming system were reduced in 2005, in order to simulate changes brought by the shift to a specialised arable farming system.
The organic rotation comprises: (1) grass-clover-alfalfa (GCA, Lolium perenne L. + Trifolium pretense L. + Medicago sativa L.), (2) potato (Solanum tuberosum L.) + catch crop mustard (Sinapis alba L.) or catch crop Egyptian clover (Trifolium alexandrinum L.) and bean (Vicia faba L.), (3) winter wheat (Triticum aestivum L.), (4) sunflower (Helianthus annuus L.) + undersown GCA, (5) GCA, (6) winter wheat, and (7) winter rye (Secale cereale L.) + undersown GCA, with a complete duration of 7 years. GCA was harvested as fodder during 1999–2002, but used as green manure during 2009–2012.
The conventional crop rotation includes: (1) forage maize (Zea mays L.) + catch crop mustard, (2) winter wheat, (3) potato + catch crop mustard, and (4) winter wheat. The cultivation of catch crops (mustard or clover) varied slightly from year to year, depending on the soil and management conditions. Compared to the period 1999–2002, the management in 2009–2012 was improved by better adaption of N fertiliser input to the N demand of the crops, and by using new crop varieties with higher yield potential.
The agroforestry systems were established in 2009 as long-term experiments on four fields (two with organic, two with conventional farming). They are a combination of strips of short rotation trees for bioenergy production (TAGFS, 22 % of AGFS area) and of crop area for arable farming for food production (CAGFS, 78 % of AGFS area). The investigated tree species were poplar (Populus maximowiczii × P. nigra), willow (Salix triandra x S. viminalis), black locust (Robinia pseudoacacia), and black alder (Alnus glutinosa). The agroforestry experiments were designed with a 4-year growing period for trees, and a total experimental period of 20 years. Further information on the design of the field experiment can be found in Lin et al.
Nitrogen balance and NUE
The N balance and the NUE were computed for the crop subsystem (soil surface balance), the animal subsystem (barn balance), and the whole farm (farm-gate balance) for the period 1999–2002. These balances can be combined into a system balance, and thus allows the quantification of all relevant N fluxes. From 2009 to 2012, no animal subsystem was involved in the farming systems and thus, the N balance was computed only for the plant (crop and tree) subsystem and the whole farm.
The modelling approach used in this study was described in Küstermann et al. (2010). In this study, we derivate N balance parameters for tree subsystems in agroforestry based on experimental data and parameter from literature (e.g. specific N contents of tree biomass to estimate the N output; N contents of leafs to calculate the N uptake). We integrate a method to calculate symbiotic N2 fixation of trees (black locust, black alder) as well as an approach to estimate SON-accumulation of trees. So the approach of Küstermann et al. (2010) was expanded and can be used in agroforestry systems with different tree species.
Parameters in this study were N input, N uptake, N output, Δ SON, NUE, and N surplus. The N input included the N fluxes from symbiotic N2 fixation, mineral N, slurry, farmyard manure, and N deposition. N in seed/seedlings was not included because it is of minor importance regarding environmental issues. The N uptake was defined as the nitrogen contained in the whole aboveground crop/tree biomass, whereas N output considered only the nitrogen contained in the harvested products. NUE was defined as N output in relation to N input. It was analysed with and without the consideration of Δ SON:
The parameter Δ SON was included in the N balance to quantify NUE and N surplus (N loss potential) more precisely (Küstermann et al. 2010).
N surplus, which represents the potential N loss by the system, was defined as the difference between N input and N output. It was also analysed with and without the consideration of Δ SON:
The N content of farming products and harvested wood was taken from Hülsbergen (2003), Seidl et al. (2014), and experimental data.
Symbiotic N2 fixation is an important N source for the tree strips and organic crop rotation in this study. The methods for calculating the symbiotic N2 fixation of legume crops and trees are described below.
Symbiotic N2 fixation of legume crops
The symbiotic N2 fixation of legume crops was calculated based on the assumption that (1) N2 fixation increases with increasing yield (Carlsson and Huss-Danell 2003; Høgh-Jensen et al. 2004) and (2) the N fixed by crops contributes to a specific share of N uptake. The equations were presented by Küstermann et al. (2010) as:
where NYsym = symbiotically fixed nitrogen in the yield (kg N ha−1 yr−1), Y = fresh matter yield (kg ha−1 yr−1), DM = dry matter content (%), N = N content in dry matter (kg N kg−1 DM), L = share of legumes among the plants (%) and %Ndfa = proportion of nitrogen derived from atmosphere (differentiated according to cropping conditions and the content of plant-available N in the soil) (%).
where NRsym = symbiotically fixed nitrogen in crop residues (kg N ha−1 yr−1), rDM = DM residues/DM yield and rN = N content residues/N content yield
Symbiotic N2 fixation by trees
We used mean %Ndfa values of locust and alder to determine the amount of symbiotic N2 fixation. The equation was expressed as:
where NTsym = symbiotically fixed nitrogen in the whole tree (wood, roots, and leaves) (kg N ha−1 yr−1), Nh = N in harvested woody biomass (= N output of trees) (kg ha−1 yr−1), Nl = N in leaf litter (kg ha−1 yr−1) and Nr = N in roots (kg ha−1 yr−1).
The parameters Nh and Nl were measured, and Nr was derived from the ratio of Nr to Nh (Uri et al. 2011). The mean value of %Ndfa of black alder and black locust in Germany is 70 %, however, it ranges from 60 to 80 % (Dittert 1992; Veste et al. 2013). Due to the good soil quality and nitrogen availability of the experimental sites, we used 60 % as the %Ndfa of black alder and black locust in this study.
Nitrogen in tree leaves was not part of the N output in the N balance because it was recycled in litterfall to the soil. Thus, it is presented in the N balance as N uptake and N in green manure.
Soil organic nitrogen
Soil organic N depletion or accumulation (Δ SON) in crop areas was calculated based on the algorithm of the model REPRO (Hülsbergen 2003; Küstermann et al. 2010). This method considered the specific effects of crops (depending on site, yield, and mineral N doses) and organic fertilisers (depending on quality and quantity) on the organic nitrogen pool of the soil, with parameters derived in long-term field experiments with various crop rotations and fertilization patterns in regions of different soil and climate conditions for more than 20 years (Küstermann et al. 2008). The Δ SON of TAGFS area was assumed to be 0 kg N ha−1 yr−1 for trees not fixing N (Petzold et al. 2010) and 30 % of the fixed N for N-fixing trees (Dulormne et al. 2003).
For the sake of clarity, only the main N fluxes are shown in our tables and figures.
Results
Nitrogen soil surface balance of the conventional farming systems
From 1999 to 2002, the N supply in the conventional arable farming system (AF1999–2002) was mainly from mineral fertiliser (75 % of the N input) (Table 1). The mean N output in the harvested biomass (150 kg ha−1) was 61 % of the N uptake. Forage maize had the highest DM yield (13.8 Mg ha−1), and thus the highest N output (178 kg ha−1) as the whole plant was harvested (uptake = output). Potato had the highest aboveground crop biomass and N uptake, but 37 % of the N was returned to the soil in the form of potato residues. However, its N output was still higher than the N supply, resulting in a negative N surplus without considering Δ SON. In winter wheat the N input was high, but, although N uptake was of a comparable order of magnitude to maize, N output was relatively low. Therefore, the N surplus in winter wheat was the highest among the crops. The highest SON depletion occurred during the cultivation of potato and forage maize, however, the catch crop mustard had a positive effect on SON (green manure). For the crop rotation, a decline of the SON (−24 kg N ha−1 yr−1) was computed. Taking Δ SON into account increases the N surplus from 44 to 68 kg ha−1, which decreases the NUE from 0.79 to 0.69.
From 2009 to 2012, the N input in the conventional arable farming system (AF2009–2012) was similar to that of AF1999–2002. The mineral N input (+23 %) was adapted to the higher yield potential and N uptake of new crop varieties (forage maize and wheat), and to compensate for the reduction in other N inputs (no slurry application). The DM yield increased by 18 % in conventional AF2009–2012, mainly due to the improved yield of forage maize (+36 %). Maize was also the crop which removed the most N from the system. Because of the higher DM yield, the N output of conventional AF2009–2012 was higher than that of conventional AF1999-2002 (+15 %). As a result, the N surplus of conventional AF2009–2012 was lower and the NUE improved compared to the reference period.
The lower yield and N uptake of catch crop mustard (after forage maize) in conventional AF2009–2012 was the result of the later harvest time of forage maize due to the type of cultivar and the weather conditions in this period. Compared to AF1999–2002, the modelled SON depletion was higher, which can be partly explained by the reduced input of green manure and no slurry application.
When comparing conventional AF2009–2012 and the conventional CAGFS area2009–2012 in the agroforestry system, the management and DM yields differed only slightly according to the soil characteristics, tree-crop interactions, and different management conditions (e.g. smaller field sizes in AGFS). The N surplus and the nitrogen-use efficiency of the CAGFS area2009–2012 are similar to the N surplus and NUE of AF2009–2012 (see Tables 2, 3).
The TAGFS area in conventional AGFS was a low-N-input (60 kg N ha−1 yr−1) and low-N-output (46 kg N ha−1 yr−1) system, although we measured relatively high N contents of the wood biomass (0.62–0.70 % N in DM) due to young plant material with a high amount of bark. The N surplus of TAGFS area was 14 kg N ha−1 yr−1. However, when taking Δ SON into consideration, the N surplus became 1 kg N ha−1 yr−1, which was much lower than that of the conventional CAGFS area. When the N stored in roots (19 kg N ha−1 yr−1, not shown in Table 3) was also considered, the N surplus decreased to −18 kg N ha−1 yr−1.
The tree species poplar and willow had a high NUE (2.87 and 1.12, respectively) and a negative N surplus (−30 and −2 kg N ha−1 yr−1, respectively) (both with and without Δ SON, Table 3) due to the low N input. The N2-fixing tree species black alder and black locust had higher N inputs compared to poplar and willow. The calculated N2 fixation amounted to more than 80 kg N ha−1 yr−1. Therefore, the NUE of black alder and black locust was considerably lower and N surplus was discernibly higher. The establishment of tree strips resulted in a 15 % lower N input in conventional AGFS, reduced the depletion of SON, and reduced the N surplus of the whole system compared to conventional AF2009–2012. However, there was no substantial effect on the NUE.
Nitrogen soil surface balance of the organic farming systems
During the period 1999–2002, the main N sources in the organic crop rotation were the symbiotic N2 fixation from grass-clover-alfalfa (47 %), and farmyard manure and slurry from animal husbandry (44 %) (Table 4). The relatively high N output of the crop rotation (140 kg N ha−1 yr−1) resulted mainly from the high-yielding grass-clover-alfalfa. GCA was also the crop with the highest N uptake, contributed the highest amount of SON accumulation, but had also a negative N balance. The whole crop rotation brought a SON accumulation of 35 kg ha−1 yr−1, a N surplus of 38 kg ha−1 yr−1 without and 3 kg ha−1 yr−1 with Δ SON, and a NUE of 0.77 without and 0.98 with Δ SON.
There was no application of FYM and slurry in the organic arable farming system between 2009 and 2012 after the shift from mixed farming to cash crop production. There was also no harvest of GCA (used as green manure) under this new management system. The N input of the organic AF was only 41 % of the N input in the crop rotation of the organic mixed farming system (Table 5). The lower N input and harvest index (DM yield/biomass production; organic AF: 0.30, organic mixed farming: 0.85,) resulted in a very low DM yield, as well as in an extremely low N output. The Δ SON of organic AF was 11 kg N ha−1 yr−1. The N surplus (8 kg N ha−1 yr−1, with Δ SON) stayed at the same level as during the organic mixed farming period. The NUE of organic AF was lower than that of the organic mixed farming system.
GCA was the crop with the highest N uptake and the highest SON accumulation in organic AF. The N2 fixation, N uptake, and SON accumulation of mulched GCA differed from the harvested GCA in the organic mixed farming system as a result of different DM yields, proportion of legumes in the mixture, and %Ndfa rate caused by specific conditions and management. The nitrogen fixed by GCA was partly accumulated in soil organic nitrogen and transferred to the other crops in the crop rotation after mineralization. The N surplus of GCA in organic AF was positive as the biomass was not removed from the system. Wheat after GCA had the highest N output, and sunflower the highest N surplus in organic AF.
The management, DM yields, and N balance of organic AF and the organic CAGFS area (2009–2012) (Table 6) differed slightly, but both were low-N-input and low-N-output systems.
In the TAGFS area of organic AGFS, poplar and willow had higher NUE (4.22 and 2.68, respectively) and lower N surplus (−52 and −27 kg N ha−1 yr−1, respectively) compared to N2-fixing black alder and black locust. Due to the higher tree yields of poplar and willow in the organic TAGFS area, the NUE of the whole organic TAGFS area was higher than the NUE of the conventional system.
Nitrogen cycle of organic mixed farming, organic arable farming, organic agroforestry, and conventional agroforestry systems
Because of the different structure and subsystems, the N fluxes and N pools of the analysed farming systems were very different.
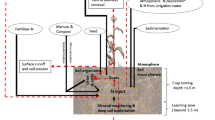
Figure 2 shows the N cycle of the organic mixed farming system (1999–2002). There were two kinds of N input: for crops (e.g. N2 fixation) and for animals (e.g. forage and straw). Nitrogen left the system via cash products (crops: 28 kg N ha−1, animals: 22 kg N ha−1) and various N losses. The greatest part (78 %) of the N in the harvested products from the crop area entered the animal subsystem as forage, was returned partly to the soil as FYM and slurry, and was taken up by the crops again. The input of forage and straw into the animal subsystem and the repeated use of N intensified the farm internal N cycle of the organic mixed farming system. In total, 69 % of the N uptake by plants was returned to the soil as manure. When the system boundary was expanded to the farm gate, the NUE of the organic mixed farming system was 0.47, and the N surplus 60 kg N ha−1 yr−1.

On-farm nitrogen cycle of the organic mixed farming system (31.1 ha arable land, 18.2 ha permanent grassland), 1999–2002. Adapted from Küstermann et al. (2010). Unit: kg N ha−1 yr−1. aLosses of ammonia in animal housing systems, bN losses during storage of slurry and farmyard manure
The N cycle of the organic arable farming system was simpler than that of the organic mixed farming system (Fig. 3). Furthermore, the N form, N availability, and N yield effects were different. There was an important N transfer from GCA to the other crops in the organic AF, so that its DM yield was sustained at a certain level without extra N input. At the farm-gate level, the NUE of the organic AF amounted to 0.74, and the N surplus amounted to 8/19 kg N ha−1 (with/without ΔSON).

On-farm nitrogen cycle of the organic arable farm, 31.1 ha arable land, 2009–2012. Unit: kg N ha−1 yr−1. a N losses from soil = NH3 losses, denitrification and leaching losses (not specified)
The N cycle of the organic CAGFS area was similar to the organic AF (Fig. 4). The N input of the organic TAGFS area (mean value of the analysed tree species) was lower than the N input of the organic CAGFS area due to the smaller amount of N2 fixation. The N uptake and N in plant residues (leaf litter) were less than those of the CAGFS area. The N output via the harvest of aboveground woody biomass (55 kg N ha−1 yr−1) was similar to the N output of the CAGFS area. The ratio of N taken up to N returned to soil in the CAGFS area, TAGFS area and the whole AGFS system was 68, 49 and 65 %, respectively. Because the N cycle of the TAGFS area was less intensive compared to the N cycle of the CAGFS area, the establishment of AGFS led to an extensification of the low-N-input organic system.

On-farm nitrogen cycle of the organic agroforestry system, 2009–2012. Unit: kg N ha−1 yr−1. a N losses from soil = NH3 losses, denitrification and leaching losses (not specified). b Interaction between TAGFS area and CAGFS area (not analysed in this study)
The intensities of N cycling, the effects on Δ SON, and the N surplus of the CAGFS and TAGFS areas in conventional AGFS were very different (Fig. 5). In the conventional CAGFS area, the N input was mainly from mineral N, and the output was via the harvest of cash crops; both were at a very high level. The input (60 kg N ha−1 yr−1) and output (46 kg N ha−1 yr−1) of the TAGFS area were much lower (no mineral fertiliser input, low N content in harvested wood). Because 78 % of the N in the conventional CAGFS area was removed by harvesting the crops, only 22 % of the N taken up by plants was returned to the soil. In contrast, 52 % of the total N uptake was returned in the conventional TAGFS area.

On-farm nitrogen cycle of the conventional agroforestry system, 2009–2012. Unit: kg N ha−1 yr−1. a N losses from soil = NH3 losses, denitrification and leaching losses (not specified). b Interaction between TAGFS area and CAGFS area (not analysed in this study)
Interactions and N fluxes between the subsystems (TAGFS area and CAGFS area) of organic and conventional AGFS were not analysed in this study and are therefore not shown in Figs. 4 and 5.
Discussion
Characteristics and restrictions of the modelling approach
System boundaries and N fluxes
Our N-balance model described agricultural farms as systems which respond to interferences like structural changes and alterations in intensity and technology. All subsystems of a farm (soil–plant–animal–environment) are linked via N fluxes, which enable the interactions between crop production and animal husbandry to be simulated. In order to analyse the effects on the N balance, NUE, and N loss potential brought by the change of farming systems, examining only the relevant N fluxes was sufficient. Our N-balance model is based on the algorithms of the model REPRO, which the N losses can be further specified as N losses via NH3 emission, denitrification, and leaching (see method description in Küstermann et al. 2010). However, these N losses were not analysed in this study.
In agroforestry systems, nitrogen may be transferred between the tree strips and the crop area, in the form of litter or through N taken up by tree roots reaching deeper soil horizons. Hence, the N input, N surplus, and Δ SON may be affected in the interactive zone. Such interactions between the subsystems were not analysed in this study. However, because the results reported in this study were from the first stage of long-term experiments, we can expect that the interactions between the tree and crop subsystems were not yet that significant (Lin et al.).
Symbiotic N2 fixation
Because symbiotic N2 fixation was the most important N input for the organic farming and tree subsystems, the accuracy of its determination is decisive for the accuracy of the N balance sheets. However, modelling the symbiotic N2 fixation of trees based on site-specific conditions is challenging because of various influencing factors (e.g. pH, moisture, temperature and nutrition level of soils) (Danso et al. 1992; Noh et al. 2009). Because the aim of this study is to understand the effects of different farm management on the whole farm system, a certain level of uncertainty in the fluxes is acceptable. The %Ndfa method (description in section “Nitrogen balance and NUE”) seems to be appropriate in this context. Nygren et al. (2012) reviewed the %Ndfa of N2 fixing trees in AGFS from 38 case studies and 19 tree species and concluded a general average of 59 ± 16.6 %, which is influenced by several factors including tree species, sampling season, and pruning frequency.
The calculated symbiotic N2 fixation of black alder and black locust in this study (88–104 kg N ha−1 yr−1 and 86–101 kg N ha−1 yr−1) was within the range found in literature (alder: 10–266 kg N ha−1 yr−1 (Bormann et al. 1993; Hurd et al. 2001; Lee and Son 2005; Sanborn et al. 2002; Son et al. 2007; Uliassi and Ruess 2002; Uri et al. 2011); black locust: 23–112 kg N ha−1 yr−1 (Boring and Swank 1984; Bormann et al. 1993; Danso et al. 1995; Noh et al. 2009)).
Soil organic nitrogen
Due to the lack of available data, it is commonly assumed that soil N is at steady state (Δ SON = 0 kg N ha−1 yr−1) when conducting N balances (Schröder et al. 2003). However, ignoring Δ SON can lead to non-negligible effects (positive or negative) on the N loss potential. In our N balance, Δ SON was calculated based on parameters and algorithms from long-term field experiments with consideration of crop yield, management, soil and climate conditions (Brock et al. 2012a, b; Leithold et al. 2015), but the derivation of Δ SON of trees was based on a comparatively small amount of existing literature. The accumulation or depletion of soil nitrogen by trees is largely influenced by site conditions (e.g. treatment before establishment of trees and soil N content), the soil layer measured, and the year after tree establishment (Jug et al. 1999; Uri et al. 2014). The actual Δ SON of N-fixing trees could be higher or lower than the amount calculated by our approach, and would accordingly change the amount of N surplus.
Nitrogen-use efficiency
Nutrient-use efficiencies can be defined in different ways: crop yield per unit of nutrient applied (partial factor productivity); crop yield increase per unit of nutrient applied (agronomic efficiency); nutrient in harvested crop per unit of nutrient applied (partial nutrient budget); or increase in aboveground crop uptake per unit of nutrient applied (recovery efficiency). In addition, some NUE calculations only consider nutrient inputs derived from fertilisers, others include nutrients from the mineralization of soil organic matter, crop residues, or manures over several crop cycles (Keating et al. 2010). These different definitions of nutrient-use efficiency have different areas of application in agricultural science and management. We defined NUE as N output in relation to N input because it considers all the relevant inputs and outputs, and reflects not only the relationship between agricultural production and resource consumption (Godinot et al. 2014) but also the management at farm level.
Godinot et al. (2014) proposed a new indicator, system nitrogen efficiency, to improve NUE. They argued NUE has the following disadvantages: (1) Δ SON is not considered, (2) indirect N losses outside the farm are not included, (3) external inputs but not producing them on farm is favoured, and (4) the quality of outputs are not distinguished (e.g. manure is not distinguished from other crop products). Gerber et al. (2014) also suggested the life cycle thinking should be incorporated in the assessment of NUE. The NUE in our study was analysed with the consideration of Δ SON. Because both soil surface balance and farm-gate balance are integrated in our model, the bias that “relying on external input is more efficient than being self-sufficient” is partly solved. The system boundary of this study is within the farm; it is set according to the aim of this study. If today we want to analyse the whole production process of food (or bioenergy), we would expand the system boundary and combine our method with life cycle assessment; N losses outside the farm would also be integrated.
Nitrogen balance of Scheyern Research Farm
N surplus
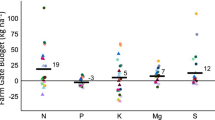
The N surplus of the conventional farming systems (68, 54, and 47 kg ha−1 yr−1, with Δ SON; 44, 21, 21 kg ha−1 yr−1, without Δ SON; see Tables 1, 2 , 3) was much lower than the average N surplus in Germany (100 kg N ha−1 yr−1, Umweltbundesamt 2014). Our results show that under the conditions of high-N-input farming, a considerable reduction in the N surplus is possible without negative yield effects. The N surplus of the organic farming systems (3, 8 and 5 kg ha−1, with Δ SON; 38, 19, 16 kg ha−1 yr−1, without Δ SON; see Tables 4, 5, 6) was within the optimum range (0–50 kg N ha−1 yr−1; Christen et al. 2009; Hülsbergen 2003) recommended from an environmental perspective.
Even though the N balance of the whole system was positive and resulted in a positive N surplus, some crops had a negative N balance. The negative N balances of crops can be partly explained by the N uptake of mineralised soil N and/or N from crop residues (N transfer within the crop rotation), but they can also indicate an over- or underestimation of N balance parameters (Δ SON, N deposition, N2 fixation, etc.).
Figure 6 shows the relation between N input and N surplus of the Scheyern Research Farm, as well as of organic and conventional farms in southern Germany with comparable soil and climatic conditions (Hülsbergen et al. 2012). Even though a lower N input does not guarantee a lower N surplus, a positive correlation is indicated. Most of the organic arable farms were low N-input systems (110–210 kg N ha−1 yr−1) with a low N surplus (<0–30 kg N ha−1 yr−1). The organic mixed farms had a medium level of N input (160–200 kg N ha−1 yr−1) and low N surplus (0–35 kg N ha−1 yr−1); the conventional arable farms were high-N-input systems (210–300 kg N ha−1 yr−1) with moderate to high N surpluses (45 to >100 kg N ha−1 yr−1).

Correlation between nitrogen input and nitrogen surplus. Data analysed for Scheyern Research Farm and 56 organic and conventional farms in Germany (revised from Hülsbergen et al. 2012)
At the Scheyern Research Farm, the conversion of the farm structure and management, i.e. from mixed farming (1999–2002) to arable farming (2009–2012), led to a considerable reduction of N input in the organic farming systems. However, this change did not further decrease the N surplus. In contrast, for the conventional farming systems, the positive effect of the improved management (2009–2012) was clear, and a further reduction of both N input and N surplus was found after the conversion to agroforestry. The transition and optimization resulted in lower N inputs for the farming systems in Scheyern (2009–2012) compared to other commercial farms. However, the regression function shown in Fig. 6 indicates that the N surplus of farming systems in Scheyern, could, potentially, be reduced further.
The results from the organic AGFS were consistent with the conclusion made by Rosenstock et al. (2014) that the theoretical N surplus of legume-based AGFS with low N input should be around 0 kg N ha−1 yr−1. However, this also depends on the N mineralization potential (soil fertility) and the N deposition. In both conventional and organic agroforestry systems, poplar and willow had negative N surpluses while black alder and black locust had medium (positive) N surpluses. The negative N surplus of poplar and willow was due to the uptake of N, although no N was applied to the tree strips. This N may be N remaining in the system from previous land use (mineral N and N in crop residues from previous arable farming) or was transferred from the CAGFS area. It is also possible that this N was from SON and therefore indicates the depletion of SON. Further research will be conducted in the future to analyse these N fluxes, interactions, and to measure the Δ SON of our experiments.
Nitrogen-use efficiency
Tilman et al. (2002) warned of a globally decreasing trend of cereal yield obtained per unit of N fertiliser applied since 1960, which implies a reduction of the nitrogen-use efficiency. They reported that only 30–50 % of the applied N fertiliser is taken up by crops. The highest N fertiliser efficiency is achieved with the first increments of N fertiliser, and declines with further additional N.
In this study, the NUE of crop areas of organic farming systems (0.87–0.98, with Δ SON) was higher than that of conventional farming systems (0.69–0.76). Torstensson et al. (2006) reported the NUE of organic arable fields in southern Sweden, which ranged from 0.34 (arable farming system) to 0.91 (mixed farming system). The NUE of conventional systems ranged from 0.71 to 0.74, with a higher NUE found in the system with cover crops. Spiertz (2010) estimated the NUE of high-N-input systems (150–300 kg N ha−1) and low-N-input systems (100–150 kg N ha−1) and found values between 0.30–0.60 and 0.40–0.70, respectively. Li et al. (2007) showed that the NUE of conventional farming systems in the North China Plain increased from 0.18 to 0.75 with a decrease in N fertiliser. These examples illustrate the high variability of the NUE of organic and conventional farming systems depending on site and management conditions. The high NUE presented in this study may be the result of ideal soil–climatic conditions, and thus high yield potential, of optimised N-management and technologies, and of the high-yielding varieties used at the Scheyern Research Farm. However, a high NUE does not necessarily indicate that the N surplus does not exceed critical environmental thresholds. The amount of mineral N remaining after the harvest plays an even more important role in the environmental N pollution issue (Spiertz 2010).
Soil organic nitrogen
We found an accumulation of SON in organic farming systems but a depletion of SON in conventional farming systems. The modelled results were confirmed by the SON contents measured at defined measuring points (Küstermann et al. 2008). The latest soil inventory conducted at Scheyern Research Farm, based on measured SON contents, revealed an increase in SON of 44 kg ha−1 yr−1 in organic mixed farming and a decrease of 38 kg ha−1 yr−1 in conventional arable farming since 1991 (Küstermann et al. 2010). The SON accumulation of the organic arable farming system was the result of the crop rotation (with legumes), straw and green manure.
Jug et al. (1999) found that the soil N in the top 30 cm in short rotation forestry in Germany could be both positive and negative (range from −50 to 20 kg N ha−1 yr−1) 8–10 years after establishment. Uri et al. (2014) reported the N in the top 10 cm soil of grey alder in Estonia increased by 26.4 kg N ha−1 yr−1 after 14 years of cultivation. This study was conducted based on data collected in the first 4 years after the establishment of AGFS and the change in SON may not yet be detectable. The literature suggests that a SON accumulation is detectable, at the earliest, 5–10 years after the change in farm management (Hülsbergen 2003; Körschens 1992). Due to the difficulty in assuming a correct value of SON for poplar and willow and due to the short duration of the experiment, we used the assumption of Petzold et al. 2010 and calculated the N balance with a SON of 0 kg N ha−1 yr−1. Less literature exists for locust and alder and the assumption of an SON accumulation of 30 % of the fixed N seemed to be appropriate, also with regard to the comparison of N-fixing and non-N-fixing tree species.
Opportunities to improve the use of nitrogen in farming systems
One of the central questions of this study is: can we increase the DM yield, optimise the NUE, sustain soil fertility, and reduce N surplus at the same time? Our results showed that (1) with better crop varieties and optimised farm management, the DM yield and NUE can be increased with a simultaneous reduction in N surplus and the related negative environmental effects, and (2) establishing conventional AGFS has positive effects on the increase of NUE, the accumulation of SON, and the reduction of N surplus.
The results of this study also showed that the crop area of the organic mixed farming system had the highest DM yield, NUE, Δ SON, and the lowest N surplus among the three organic farming systems. The good results from the organic mixed farming system were attributed to the intensive internal N cycle between soil–plant–animal, compared to the soil–plant cycle of organic arable farming. In organic mixed farming systems, animal manure and slurry can also be applied flexibly in both space and time, according to expected plant needs.
Bryzinski and Hülsbergen (2015) detected significantly higher DM yields in fields applying FYM and slurry (organic mixed farming systems) compared to fields applying straw and green manure (organic arable farming systems) in a long-term field experiment in southern Germany. Schmid et al. (2013) also found that crop areas in organic mixed farming systems performed better than organic arable farming systems regarding DM yield, energy recovery, nitrogen use efficiency, SON accumulation, and N surplus, in 28 farms in Germany. The effects from establishing organic mixed farming systems (compared to those from arable farming systems) shown in these studies confirm the results of this study.
However, even though there were clear differences in DM yield and N output between the crop rotation of the organic mixed farming system (DM yield: 6.9 kg DM ha−1 yr−1; N output: 140 kg N ha−1 yr−1) and the organic arable farming system (DM yield: 2.5 kg DM ha−1 yr−1; N output: 54 kg N ha−1 yr−1) in this study, the N output of their cash crops were comparable (organic mixed farming system: 50 kg N ha−1 yr−1; organic arable farming system: 54 kg N ha−1 yr−1, see Figs. 2, 3). This reduction in cash crop-N in the mixed farming system was expected because of the conversion loss of N from plant protein to animal protein (Oenema and Tamminga 2005). Other N loss sources in an animal subsystem are, for example, the emission of ammonia in animal housing systems and during storage of slurry (Leip et al. 2011). The NUE of mixed farming systems is therefore lower than NUE of arable farming systems, when the N balance is calculated at farm scale (Leip et al. 2011).
The negative N balance of non-N2 fixing trees (poplar and willow) indicated a low N loss potential. However, it also indicated that there is no potential for SON accumulation. The N uptake of poplar was also rather high. Therefore, in the long run, an additional N input for poplar and willow could be necessary. Their negative N balance offers the chance to reduce the N surplus and N loss potential of the whole AGFS system. In contrast, the N2 fixing trees (black alder and black locust) lead to positive N balances. The SON accumulated by them offers the chance for N transfer and to improve soil fertility in the interaction zone between the TAGFS and the CAGFS areas in an agroforestry system.
Conclusion
Case studies on farms, like the Scheyern Research Farm, represent a valuable and necessary supplement to field experiments. A decisive advantage is the complete and realistic description of farm internal mass fluxes in the soil–plant–animal–environment system.
N balancing tools have become widely used by scientists, policymakers, consultants, and farmers as useful instruments for planning and control of on-farm nitrogen management. This study improved an existed N balancing method by integrating the N parameters for crop rotation and tree strips, so the improved method can be used in mixed farming, arable farming, and agroforestry systems. Simple N balance approaches neglect internal pools and flows of nitrogen on farms. However, if special emphasis is to be given to system analysis and optimization, farm internal structures and processes have to become the focus of attention. An analysis of nitrogen flux relationships facilitates the comprehensive understanding of a system. Our approach reveals the causes of NUE differences and is the precondition for scenario calculations aimed at reducing N losses.
The results in conventional and organic agroforestry showed that poplar and willow are especially suitable for high-N-input systems (conventional farming) because the negative N surplus balances the high surplus of those systems; black alder and black locust are, in contrast, ideal for low-N-input-systems (organic farming) because of their ability to accumulate SON and offer additional N sources. The improved conventional arable farming and conventional agroforestry system in this study showed that a significant reduction of N surplus without negative yield effects is possible. Mixed farming systems are one of the best ways to run organic farming systems sustainably regarding the use of N, and organic AGFS has the potential to increase the DM yield of the whole system, to improve nitrogen-use efficiency, and to reduce negative environmental effects. Our findings can be used in setting up the strategies and policies for agricultural N surplus reduction (e.g. encouraging the establishment of different agroforestry systems for organic and conventional farms), therefore mitigate the environmental problems brought by agricultural N emissions.
References
Bambrick AD, Whalen JK, Bradley RL, Cogliastro A, Gordon AM, Olivier A, Thevathasan NV (2010) Spatial heterogeneity of soil organic carbon in tree-based intercropping systems in Quebec and Ontario, Canada. Agroforest Syst 79:343–353
Boring LR, Swank WT (1984) Symbiotic nitrogen fixation in regenerating black locust (Robinia pseudoacacia L.) stands. Forest Sci 30:528–537
Bormann BT, Bormann FH, Bowden WB, Pierce RS, Hamburg SP, Wang D, Snyder MC, Li CY, Ingersoll RC (1993) Rapid N2 fixation in pines, alder, and locust: evidence from the sandbox ecosystems study. Ecology 74:583–598
Brock C, Hoyer U, Leithold G, Hülsbergen K-J (2012a) The humus balance model (HU-MOD): a simple tool for the assessment of management change impact on soil organic matter levels in arable soils. Nutr Cycl Agroecosyst 92:239–254
Brock C, Oberholzer H-R, Schwarz J, Fließbach A, Hülsbergen K-J, Koch W, Pallutt B, Reinicke F, Leithold G (2012b) Soil organic matter balances in organic versus conventional farming—modelling in field experiments and regional upscaling for cropland in Germany. Org Agric 2:185–195
Bryzinski T, Hülsbergen K-J (2015) Energiebilanzen und Erträge ökologischer und konventioneller Anbausysteme: erste Analyseergebnisse eines Dauerfeldversuchs in Süddeutschland. In: Beiträge zur 13. Wissenschaftstagung Ökologischer Landbau. Verlag Dr. Köster, Berlin, pp 248–251
Carlsson G, Huss-Danell K (2003) Nitrogen fixation in perennial forage legumes in the field. Plant Soil 253:353–372
Christen O, Hövelmann L, Hülsbergen K-J, Packeiser M, Rimpau J, Wagner B (2009) Nachhaltige landwirtschaftliche Produktion in der Wertschöpfungskette Lebensmittel. Erich Schmidt Verlag, Berlin
Crutzen PJ, Mosier AR, Smith KA, Winiwarter W (2008) N2O release from agro-biofuel production negates global warming reduction by replacing fossil fuels. Atmos Chem Phys 8:389–395
Danso S, Bowen G, Sanginga N (1992) Biological nitrogen fixation in trees in agro-ecosystems. Plant Soil 141:177–196
Danso S, Zapata F, Awonaike K (1995) Measurement of biological N2 fixation in field-grown Robinia pseudoacacia L. Soil Biol Biochem 27:415–419
Dawson M (2007) Short rotation coppice willow best practice guidelines. Renew Project. http://www.spiritsolar.co.uk/wp-content/uploads/2013/05/short-rotation-coppice-willow.pdf. Accessed 23 Sept 2015
Dawson CJ, Hilton J (2011) Fertiliser availability in a resource-limited world: production and recycling of nitrogen and phosphorus. Food Policy 36:S14–S22
Dittert K (1992) Die stickstoffixierende Schwarzerle-Frankia-Symbiose in einem Erlenbruchwald der Bornhöveder Seenkette. EcoSys Suppl 5:1–98
Dulormne M, Sierra J, Nygren P, Cruz P (2003) Nitrogen-fixation dynamics in a cut-and-carry silvopastoral system in the subhumid conditions of Guadeloupe, French Antilles. Agroforest Syst 59:121–129
European Environmental Bureau, ClientEarth, AirClim (2013) Ammonia control options for better air quality. http://www.eeb.org/?LinkServID=E1B5ACC1-5056-B741-DB92489757EB9AC0&showMeta=0&aa. Accessed 23 Sept 2015
Gerber PJ, Uwizeye A, Schulte RPO, Opio CI, de Boer IJM (2014) Nutrient use efficiency: a valuable approach to benchmark the sustainability of nutrient use in global livestock production. Curr Opin Environ Sustain 9–10:122–130
Godinot O, Carof M, Vertès F, Leterme P (2014) SyNE: an improved indicator to assess nitrogen efficiency of farming systems. Agric Syst 127:41–52
Høgh-Jensen H, Loges R, Jørgensen FV, Vinther FP, Jensen ES (2004) An empirical model for quantification of symbiotic nitrogen fixation in grass-clover mixtures. Agric Syst 82:181–194
Hülsbergen K-J (2003) Entwicklung und Anwendung eines Bilanzierungsmodells zur Bewertung der Nachhaltigkeit landwirtschaftlicher Systeme. Shaker, Aachen
Hülsbergen K-J, Feil B, Biermann S, Rathke G-W, Kalk W-D, Diepenbrock W (2001) A method of energy balancing in crop production and its application in a long-term fertilizer trial. Agric Ecosyst Environ 86:303–321
Hülsbergen K-J, Küstermann B, Engelmann K (2012) Indikatoren zur Beurteilung der N-Effizienz in ökologischen und konventionellen Betriebssystemen. In: N-Effizienz im Spannungsfeld “Ertrags- und Qualitätssicherung – Nachhaltigkeit – Umweltauflagen”. Wissenschaftliche Tagung am 1./2. März 2012. LEUCOREA, Lutherstadt Wittenberg. Martin-Luther-Universität Halle-Wittenberg, Halle, pp 25-30
Hurd T, Raynal D, Schwintzer C (2001) Symbiotic N2 fixation of Alnus incana ssp. rugosa in shrub wetlands of the Adirondack Mountains, New York, USA. Oecologia 126:94–103
Ilany T, Ashton MS, Montagnini F, Martinez C (2010) Using agroforestry to improve soil fertility: effects of intercropping on Ilex paraguariensis (yerba mate) plantations with Araucaria angustifolia. Agroforest Syst 80:399–409
Isaac ME, Ulzen-Appiah F, Timmer VR, Quashie-Sam SJ (2007) Early growth and nutritional response to resource competition in cocoa-shade intercropped systems. Plant Soil 298:243–254
Johann-Heinrich von Thünen-Institut–Institut für Forstgenetik (2012) Poplars and Willows in Germany: report of the National Poplar Commission. Federal Ministry of Food, Agriculture and Consumer Protection (BMELV), Bonn
Jose S (2009) Agroforestry for ecosystem services and environmental benefits: an overview. Agroforest Syst 76:1–10
Ju XT, Kou CL, Zhang FS, Christie P (2006) Nitrogen balance and groundwater nitrate contamination: comparison among three intensive cropping systems on the North China Plain. Environ Pollut 143:117–125
Jug A, Makeschin F, Rehfuess K, Hofmann-Schielle C (1999) Short-rotation plantations of balsam poplars, aspen and willows on former arable land in the Federal Republic of Germany. III. Soil ecological effects. Forest Ecol Manag 121:85–99
Keating BA, Carberry PS, Bindraban PS, Asseng S, Meinke H, Dixon J (2010) Eco-efficient agriculture: concepts, challenges, and opportunities. Crop Sci 50:109–119
Körschens M (1992) Simulationsmodelle für den Umsatz und die Reproduktion der organischen Substanz im Boden. Berichte über Landwirtschaft 206:140–154
Küstermann B, Kainz M, Hülsbergen K-J (2008) Modeling carbon cycles and estimation of greenhouse gas emissions from organic and conventional farming systems. Renew Agric Food Syst 23:38–52
Küstermann B, Christen O, Hülsbergen K-J (2010) Modelling nitrogen cycles of farming systems as basis of site- and farm-specific nitrogen management. Agric Ecosyst Environ 135:70–80
Lee YY, Son Y (2005) Diurnal and seasonal patterns of nitrogen fixation in an Alnus hirsuta plantation of central Korea. J Plant Biol 48:332–337
Leip A, Britz W, Weiss F, de Vries W (2011) Farm, land, and soil nitrogen budgets for agriculture in Europe calculated with CAPRI. Environ Pollut 159:3243–3253
Leithold G, Hülsbergen K-J, Brock C (2015) Organic matter returns to soils must be higher under organic compared to conventional farming. J Plant Nutr Soil Sc 178:4–12
Li X, Hu C, Delgado JA, Zhang Y, Ouyang Z (2007) Increased nitrogen use efficiencies as a key mitigation alternative to reduce nitrate leaching in north china plain. Agric Water Manage 89:137–147
Lin HC, Huber JA, Gerl G, Hülsbergen K-J Effects of changing farm management and farm structure on the energy balance and energy-use efficiency: a case study in organic and conventional farming systems in Southern Germany. Eur J Agron (submitted)
Millar N, Robertson GP, Grace PR, Gehl RJ, Hoben JP (2010) Nitrogen fertilizer management for nitrous oxide (N2O) mitigation in intensive corn (Maize) production: an emissions reduction protocol for US Midwest agriculture. Mitig Adapt Strateg Glob Change 15:185–204
Musshoff O (2012) Growing short rotation coppice on agricultural land in Germany: a real options approach. Biomass Bioenerg 41:73–85
Nerlich K, Graeff-Hönninger S, Claupein W (2013) Agroforestry in Europe: a review of the disappearance of traditional systems and development of modern agroforestry practices, with emphasis on experiences in Germany. Agroforest Syst 87:475–492
Noh NJ, Son Y, Koo JW, Seo KW, Kim RH, Lee YY, Yoo KS (2009) Comparison of nitrogen fixation for north- and south-facing Robinia pseudoacacia stands in central Korea. J Plant Biol 53:61–69
Nygren P, Fernández MP, Harmand J-M, Leblanc HA (2012) Symbiotic dinitrogen fixation by trees: an underestimated resource in agroforestry systems? Nutr Cycl Agroecosys 94:123–160
Oenema O, Tamminga S (2005) Nitrogen in global animal production and management options for improving nitrogen use efficiency. Sci China Ser C 48:871–887
Patra AK (2013) Tree–crop interaction in agroforestry. Sci Horiz 4:12–17
Petzold R, Schubert B, Feger K-H (2010) Biomasseproduktion, Nährstoffallokation und bodenökologische Veränderungen einer Pappel-Kurzumtriebsplantage in Sachsen (Deutschland). Die Bodenkultur 61:23–35
Reynolds PE, Simpson JA, Thevathasan NV, Gordon AM (2007) Effects of tree competition on corn and soybean photosynthesis, growth, and yield in a temperate tree-based agroforestry intercropping system in southern Ontario, Canada. Ecol Eng 29:362–371
Rosenstock T, Tully K, Arias-Navarro C, Neufeldt H, Butterbach-Bahl K, Verchot L (2014) Agroforestry with N2-fixing trees: sustainable development’s friend or foe? Curr Opin Environ Sustain 6:15–21
Ryschawy J, Choisis N, Choisis J, Joannon A, Gibon A (2012) Mixed crop-livestock systems: an economic and environmental-friendly way of farming? Animal 6:1722–1730
Sanborn P, Preston C, Brockley R (2002) N2-fixation by Sitka alder in a young lodgepole pine stand in central interior British Columbia, Canada. Forest Ecol Manag 167:223–231
Schmid H, Braun M, Hülsbergen K-J (2013) Treibhausgasbilanzen und ökologische Nachhaltigkeit der Pflanzenproduktion – Ergebnisse aus dem Netzwerk der Pilotbetriebe. In: Hülsbergen K-J, Rahmann G (eds) Klimawirkungen und Nachhaltigkeit ökologischer und konventioneller Betriebssysteme: Untersuchungen in einem Netzwerk von Pilotbetrieben, Thünen Report, No. 8. Johann Heinrich von Thünen-Institut, Braunschweig, p 259–293
Schröder P, Huber B, Olazábal U, Kämmerer A, Munch J (2002) Land use and sustainability: FAM research network on agroecosystems. Geoderma 105:155–166
Schröder J, Aarts H, Ten Berge H, Van Keulen H, Neeteson J (2003) An evaluation of whole-farm nitrogen balances and related indices for efficient nitrogen use. Eur J Agron 20:33–44
Seidl F, Mastel K, Aust C, Kannenwischer N, Dieterich M (2014) Kurzumtriebsplantagen (KUP) und Miscanthus in Baden-Württemberg–Ergebnisse aus dem Forschungsprojekt„ Biomasse aus Kurzumtrieb” 28.01.2008–31.12.2013. Landwirtschaftliches Technologiezentrum Augustenberg, Karlsruhe
Sevel L, Nord-Larsen T, Ingerslev M, Jørgensen U, Raulund-Rasmussen K (2014) Fertilization of SRC willow, I: biomass production response. Bioenergy Res 7:319–328
Snyder CS, Bruulsema TW, Jensen TL, Fixen PE (2009) Review of greenhouse gas emissions from crop production systems and fertilizer management effects. Agric Ecosyst Environ 133:247–266
Son Y, Lee YY, Lee CY, Yi MJ (2007) Nitrogen fixation, soil nitrogen availability, and biomass in pure and mixed plantations of alder and pine in central Korea. J Plant Nutr 30:1841–1853
Spiertz J (2010) Nitrogen, sustainable agriculture and food security. A review. Agron Sustain Dev 30:43–55
Stewart W, Dibb D, Johnston A, Smyth T (2005) The contribution of commercial fertilizer nutrients to food production. Agron J 97:1–6
Sutton MA, Howard CM, Erisman JW, Billen G, Bleeke A, Grennfelt P, van Grinsven H, Grizzetti B (2011a) The European nitrogen assessment: sources, effects and policy perspectives. Cambridge University Press, Cambridge
Sutton MA, Oenema O, Erisman JW, Leip A, van Grinsven H, Winiwarter W (2011b) Too much of a good thing. Nature 472:159–161
Taube F, Balmann A, Bauhus J, Birner R, Bokelmann W, Christen O, Gauly M, Grethe H, HolmMüller K, Horst W, Knierim U, Latacz-Lohmann U, Nieberg H, Qaim M, Spiller A, Täuber S, Weingarten P, Wiesler F (2013) Novellierung der Düngeverordnung: Nährstoffüberschüsse wirksam begrenzen. Berichte über Landwirtschaft-Zeitschrift für Agrarpolitik und Landwirtschaft 219:1–12
Tilman D (1999) Global environmental impacts of agricultural expansion: the need for sustainable and efficient practices. Proc Natl Acad Sci USA 96:5995–6000
Tilman D, Cassman KG, Matson PA, Naylor R, Polasky S (2002) Agricultural sustainability and intensive production practices. Nature 418:671–677
Torstensson G, Aronsson H, Bergström L (2006) Nutrient use efficiencies and leaching of organic and conventional cropping systems in Sweden. Agron J 98:603–615
Uliassi DD, Ruess RW (2002) Limitations to symbiotic nitrogen fixation in primary succession on the Tanana River floodplain. Ecology 83:88–103
Umweltbundesamt (2014) Nährstoffeinträge aus der Landwirtschaft und Stickstoffüberschuss. http://www.umweltbundesamt.de/daten/land-forstwirtschaft/landwirtschaft/naehrstoffeintraege-aus-der-landwirtschaft. Accessed 07 July 2015
Uri V, Lõhmus K, Mander Ü, Ostonen I, Aosaar J, Maddison M, Helmisaari H-S, Augustin J (2011) Long-term effects on the nitrogen budget of a short-rotation grey alder (Alnus incana (L.) Moench) forest on abandoned agricultural land. Ecol Eng 37:920–930
Uri V, Aosaar J, Varik M, Becker H, Ligi K, Padari A, Kanal A, Lõhmus K (2014) The dynamics of biomass production, carbon and nitrogen accumulation in grey alder (Alnus incana (L.) Moench) chronosequence stands in Estonia. Forest Ecol Manag 327:106–117
van Keulen H, Schiere H (2004) Crop-livestock systems: old wine in new bottles. In: New directions for a diverse planet. Proceedings of the 4th international crop science congress, Brisbane, Australia, 26 September–1 October
Veste M, Böhm C, Quinkenstein A, Freese D (2013) Biologische Stickstoff-Fixierung der Robinie. AFZ-Der Wald 2:40–42
Vockinger F (2013) Analyse der Flächenentwicklung, Anbaustrukturen und Fruchtfolgen ökologisch bewirtschafteter Ackerflächen Bayerns auf der Basis agrarstruktureller Daten. Bachelor Thesis, Technische Universität München
Wetterdienst D (2012) Klimadaten für Messstationen in Deutschland - Mittelwerte 30-jähriger Perioden (1981–2010). http://www.dwd.de/bvbw/appmanager/bvbw/dwdwwwDesktop. Accessed 12 Oct 2015
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Lin, HC., Huber, J.A., Gerl, G. et al. Nitrogen balances and nitrogen-use efficiency of different organic and conventional farming systems. Nutr Cycl Agroecosyst 105, 1–23 (2016). https://doi.org/10.1007/s10705-016-9770-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10705-016-9770-5