
Abstract
The intensive culture of characid teleosts for ornamental trade is highly dependent on live feed organisms, particularly Artemia nauplii, to provide nutrition through the larval stage. Live feeds have inherent disadvantages relative to prepared microparticulate diets (MDs), specifically availability, labor and cost. In this research, the dependence of larval Paracheirodon innesi on live Artemia was confirmed via a nutritional trial. Next, digestive system ontogeny was characterized from the onset of exogenous feeding through metamorphosis. P. innesi exhibited an agastric larval stage, as well as low digestive enzyme activity at the onset of exogenous feeding followed by abrupt increases in trypsin, lipase and pepsin activity. Differentiation of the stomach, including gastric gland formation and production of neutral mucopolysaccharides, as well as the onset of pepsin activity, did not occur until 20 days post hatch (dph; 5.24 ± 0.20 mm). This shift from agastric to gastric digestive modes is indicative of a proliferation of digestive capacity and subsequent prey diversity in other fish species exhibiting similar altricial larval stages.
Based on this information, different schedules for weaning from Artemia to a MD were evaluated. For P. innesi fed until 32 dph, weaning beginning at 12 dph and 17 dph resulted in similar survival to live Artemia (mean: 22.0 ± 1.7%), and the MD resulted in the lowest survival (0.8 ± 0.3%). These results indicate that weaning is possible prior to gastric differentiation, potentially resulting in the reduction of Artemia use in the larval culture P. innesi.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The neon tetra Paracheirodon innesi is a small, neotropical characid that is one of the most highly traded fish in the ornamental industry (Weitzman et al. 1983; Chapman et al. 1997, 1998). While an estimated 80% of the market demand, or 12–15 million individuals annually, is satisfied via import from wild collections originating in their native range, foreign and domestic aquaculture of P. innesi represents an economically significant endeavor with the potential for expansion (Chapman et al. 1998; Chao 2001; Kucharczyk et al. 2010). P. innesi, like many other species in the ornamental aquaculture trade, are characterized by having very small, poorly developed larvae at first feeding, which has led to the ubiquitous use of live zooplankters as first feed items (Lim et al. 2003; Sanaye et al. 2014). While rotifers (Brachionus spp.), Paramecium spp., and other live feed organisms have been used during the critical larval stage of P. innesi culture, newly hatched Artemia franciscana nauplii are the most widely used for this application (Chapman et al. 1998; Lim et al. 2003; Sanaye et al. 2014).
Commercial producers of freshwater ornamental fish rely heavily on newly-hatched 1–2 instar stage A. franciscana for early larval nutrition in intensive and semi-intensive aquaculture operations (Lim et al. 2003). This wild caught feed item, while convenient and nutritious relative to some other live feeds, is characterized by increasing costs, limited availability, and variable quality, with periods of shortage being a primary concern to ornamental fish producers. Commercially available microparticulate diets (MDs) can provide more complete nutrition than live feeds and are more convenient and reliable (Hamre et al. 2013; Kolkovski 2013; Lipscomb et al. 2020a). While total replacement of live feeds with MDs is ideal, this is not feasible for all species, particularly those with small, altricial larval stages (Rønnestad et al. 2003). In many marine fishes, as well as some freshwater species, first feeding larvae are physiologically incapable of utilizing complex nutrients found in MDs, therefore reduced reliance on live feeds requires optimizing the timing of weaning following critical developmental milestones (Cahu and Zambonino Infante 2001; Kolkovski 2001; Faulk and Holt 2009).
The gastrointestinal (GI) tract in teleosts begins as a straight and undifferentiated tube that generally resembles the intestine of a more developed juvenile (Govoni et al. 1986). At the earliest stages, collective digestive enzyme activities are generally low, and proteolytic hydrolysis is primarily catalyzed in the intestinal lumen by trypsin and other trypsin-like enzymes of pancreatic origin (Holt 2011). In gastric fishes, developing larvae exhibit incremental increases in activities of individual enzymes, as well as the differentiation of the anterior GI into a functioning stomach. Gastric glands, a pyloric valve, and folding and elongation of the GI, in conjunction with the production of hydrochloric acid, pepsin, and protective neutral mucopolysaccharides all characterize this critical developmental milestone (Holt 2011). The development of the stomach affords the larvae a greatly increased aptitude for digesting complex nutrients, particularly those present in MDs (Rønnestad et al. 2003; Thompson et al. 2019). Previous authors have used the timing of the differentiation of the stomach, as well as peaks in digestive enzyme activities, to develop data-based weaning protocols for use in the production of fishes that exhibit this delayed mode of larval development (Faulk and Holt 2009; Thompson et al. 2019).
Many members of the order Characiformes have been identified as species that exhibit extended agastric larval phases, including pacu Piaractus mesopotomicus, bocón Brycon amazonicus, and dorado Salminus brasiliensis, as reviewed by Portella et al. (2014). Additionally, it has been shown that live-feed dependence of the black tetra Gymnocorymbus ternetzi larvae at first feeding was due to an agastric larval period and weaning to MD is possible shortly before stomach differentiation (Lipscomb et al. 2020b). By categorizing the GI development and digestive enzyme ontogeny of P. innesi, a reduction in weaning time and minimization of reliance on Artemia can be achieved.
In addition to utilizing biochemical assays to quantify digestive enzyme activities, histological and histochemical analysis of GI structures provides further evidence for timing of stomach differentiation, including the formation of gastric glands, which house oxynticopeptic cells, responsible in teleosts for producing both pepsinogen and hydrochloric acid. Additionally, the presence of neutral pH mucopolysaccharides, produced by the epithelial cells of the stomach to protect the tissues from hydrochloric acid and autolytic hydrolysis catalyzed by pepsin, indicate the presence of a functional stomach organ (Zambonino Infante and Cahu 2007; Ronnestad et al 2013; Yúfera et al. 2018; Nolasco-Soria et al. 2020). The appearance of these milestones can be used to identify the timing of stomach differentiation and subsequently optimize weaning schedules.
Here, we used an applied nutritional trial to evaluate the effectiveness of feeding protocols associated with three commercially available MDs relative to Artemia nauplii for first feeding in P. innesi. We also characterized the development of the gastrointestinal tract using histology (H & E stain) and histochemistry (PAS), as well as the ontogeny of three critical digestive enzymes (trypsin, bile-salt dependent lipase, and pepsin) through 32 days post hatch (dph), a time period inclusive of stomach differentiation. Subsequently, the appearance of gastric glands, PAS positive neutral mucopolysaccharides, and initiation of pepsin activity were used to guide empirical evaluations of weaning from Artemia to a MD. Lastly, the impacts of different weaning regimes on survival, growth, and digestive enzyme activity were assessed. The results of this study can be used to optimize the larval culture of P. innesi, reducing the overall reliance of commercial producers on live feeds during this critical production stage.
Materials and methods
General methods
Larval P. innesi for each trial were acquired from a commercial ornamental producer (VW Fish Hatcheries Inc, Lakeland, FL). At the hatchery, brood fish were placed in pairs in individual 10-L spawning tanks with spawning media and were allowed to volitionally spawn. Following spawning, brood fish were removed, and embryos hatched in-situ. At 1 dph, larvae were pooled and transferred to the University of Florida’s Tropical Aquaculture Laboratory, Ruskin, FL where they were then counted by hand into experimental tanks. Dissolved oxygen (DO), pH, temperature, total ammonia nitrogen (TAN), and nitrite-N were monitored weekly for each tank in each trial. DO (7.40 ± 0.34 mg l−1, average ± standard deviation) and temperature (24.96 ± 0.60 °C) were measured using an optical dissolved oxygen meter (ProComm II, YSI Industries, Yellow Springs, OH). TAN (0.2 ± 0 mg l−1), nitrite (0), and pH (8 ± 0) were measured using colorimetric assays according to manufacturer’s protocols (Hach Company, Loveland, CO).
Nutritional trial
Two dph larval P. innesi were stocked into twenty-four 3 L tanks at a density of 200 larvae per tank. Tanks were maintained on flow-through, degassed well water for the duration of the feeding trial. Beginning at 4 dph, concurrent with yolk-sac depletion, larvae were offered one of four feeding treatments (n = 6 per treatment) ad libitum to satiation, as indicated by the presence of feed in the tank following 15 min of feeding time. The treatments were as follows: newly hatched, 1st – 2nd instar stage A. franciscana nauplii twice per day, or one of three commercial MDs twice per day [MD1 (Larval AP-100, < 100 µm, Ziegler Bros Inc., Gardners, PA; 50% protein, 12% fat), MD2 (Golden Pearls 50 -100 µm, Brine Shrimp Direct, Ogden, UT, USA; 60% protein, 8% fat), and MD3 (Otohime A1, 75—150 µm, Marubeni Nisshin Feed Co. LTD, Tokyo, Japan; 53% protein, 8% fat)]. Artemia cysts were decapsulated in 10% sodium hypochlorite and stored at 4 °C prior to hatching in a 30 g/L saline solution.
For evaluation of larval feeding, two feeding techniques were employed, congruent with established methods for live feed delivery and MD delivery, respectively (Siccardi et al. 2009). For feeding Artemia, nauplii were harvested using a small brine net and transferred to a 100 ml sample cup filled with degassed well water. Nauplii were then introduced to the experimental tanks to reach a concentration of 100 nauplii/L per feeding. MDs were simply added directly to the tank at a rate of ~ 13.3 mg/L per day, spread across two feedings. Feeding frequency was based established feeding protocols employed by commercial producers of P. innesi (personal communication Eric Cassiano, Assistant Extension Scientist, University of Florida). Following 10 days of feeding (14 dph; 4.71 ± 0.10 mm), all larvae remaining in each tank were harvested and enumerated.
Digestive ontogeny characterization study
Two dph larvae were stocked into each of three 10 L tanks maintained on flow-through, degassed well water at a density of 600 per tank. Prior to the first feeding of the day at 3, 5, 7, 9, 11, 14, 17, 20, 23, 26, and 28 dph, 10 larvae from each tank were harvested and comingled in the same sampling cup (n = 30 larvae per sampling day). All 30 larvae were photographed on a Sedgewick rafter cell and measured for notochord length (NL) prior to hypural plate formation (20 dph, 5.23 ± 0.20 mm) or standard length (SL) thereafter (mm). After photographs were taken, 20 larvae were placed on a Kimwipe to absorb excess moisture, placed in 1.5 ml microcentrifuge tubes, and stored at -80°C for future processing for pancreatic and gastric digestive enzyme assays (10 larvae for each category of enzyme). The remaining 10 larvae were placed in a 1.5 ml microcentrifuge tube filled with 1 ml of Trump’s fixative (Electron Microscopy Sciences, Hatfield, PA) and stored at 4°C for future histological and histochemical processing.
All histological and histochemical processing was completed at the Fish and Wildlife Research Institute, operated by the Florida Fish and Wildlife Conservation Commission, St. Petersburg, FL. Samples were embedded in epoxy, longitudinally sectioned, and stained with either routine H & E (all sampling days), or periodic acid Schiff’s (PAS)/iron-hematoxylin with metanil yellow for contrast (14, 17, 20, and 23 dph; 4.71 ± 0.10, 5.56 ± 0.12, 5.24 ± 0.20, and 5.87 ± 0.13 mm, Quintero-Hunter et al. 1991) to identify neutral mucopolysaccharides produced in the functioning, glandular stomach (Çinar and Şenol 2006; Ghosh and Chakrabarti 2015).
Weaning study
Following characterization of GI development and digestive enzyme ontogeny, a study was conducted to evaluate the effect of different weaning schedules on survival, growth and digestive enzyme activity. Twenty 10 L tanks maintained on flow-through, degassed well water were each stocked with 500 P. innesi larvae at 1 dph. Fish were fed twice daily beginning at 4 dph using one of five different feeding regimes: Artemia nauplii for the entirety of the trial, MD3 for the entirety of the trial, or one of three weaning schedules (n = 4, Fig. 1). During periods of cofeeding, half rations of each MD3 and Artemia were given. MD3 was chosen for the weaning trial based on having the highest numerical survival following the nutritional trial. Weaning schedules consisted of a period of feeding exclusively Artemia from first feeding, followed by a 5-day period of cofeeding with Artemia and MD3, and finally a transition to exclusively MD3. Schedules were as follows: W1 (Artemia 4 – 12 dph; cofeed 13 – 17 dph; MD1 18 – 32 dph), W2 (Artemia 4 – 17 dph; cofeed 18 – 22 dph; MD3 23 – 32 dph), and W3 (Artemia 4 – 22 dph; cofeed 23 – 27 dph; MD3 28 – 32 dph). At 32 dph, each tank was harvested, fish were enumerated, and 20 fish were photographed, measured and frozen at -80°C for future processing for pancreatic and gastric digestive enzyme assays as previously described. Tanks fed exclusively MD3 exhibited low survival and were thus excluded from subsequent enzymatic assays.

Experimental feeding regimes for weaning optimization trial. Exclusive feeding with Artemia nauplii is indicated by black sections, while exclusive feeding with MD3 is indicated by white. Checkered sections indicate periods of cofeeding. Weaning schedules were as follows: W1 (Artemia 4 – 12 dph; cofeed 13 – 17 dph; MD1 18 – 32 dph), W2 (Artemia 4 – 17 dph; cofeed 18 – 22 dph; MD3 23 – 32 dph), and W3 (Artemia 4 – 22 dph; cofeed 23 – 27 dph; MD3 28 – 32 dph). Dph = days post hatch
Digestive enzyme activity analysis
Digestive enzyme activity assays were performed according to Zambonino Infante and Cahu (1994, pepsin), and Faulk et al. (2007, trypsin and bilesalt-dependent lipase). On the morning of each assay, samples were thawed in an aluminum cooling chamber nested in ice. For measuring pepsin activity, pooled samples of 10 larvae were homogenized with a motorized micro-pestle in 400 µl reagent grade deionized water. Samples were then centrifuged for 10 min at 1700 G at 4 \(^\circ{\rm C}\), and 100 µl of each supernatant was transferred to a new microcentrifuge tube. In addition, a negative control with 100 µl of reagent grade deionized water, as well as a positive control with 100 µl of 50 µg/ml pepsin solution were aliquoted into microcentrifuge tubes. With tubes in a water bath at 37 \(^\circ{\rm C}\), 500 µl of hemoglobin substrate (60 mM HCl, 2% w/v equine hemoglobin) was added to each sample. After 20 min, 1 ml of 5% trichloroacetic acid was added to the tubes to quench the reaction. For each sample, a blank was created by adding 1 ml of 5% trichloroacetic acid prior to the addition of the hemoglobin substrate. Samples and blanks were centrifuged for 10 min at 1700 G and 20 µl of each supernatant was pipetted into a 96-well plate in triplicate, in addition to positive and negative controls. The concentration of tyrosine liberated from hydrolysis of the substrate was measured as absorbance at 280 nm using a BioTek spectrophotometer (Synergy HTX Multi-Mode Microplate Reader, BioTek Instruments, Inc., Winooski, VT, USA). Pepsin activity was expressed as µmol of tyrosine liberated per minute.
Samples of 10 fish for both trypsin and bile-salt dependent lipase were homogenized in a buffer solution (20 mM Tris–HCl, 1 mM EDTA, 1 mM CaCl2, 7.5 pH) in 1.5 ml microcentrifuge tubes and centrifuged for 10 min at 1700 G at 4 \(^\circ{\rm C}\). To measure trypsin activity, 20 µl of each sample supernatant, as well as 15 µg/ml porcine trypsin solution (positive control) and homogenization buffer (negative control) were pipetted in triplicate into a 96-well plate resting on an aluminum cooling plate in ice. Trypsin substrate solution (100 µL, 1 mM Nα-benzoyl-DL-arginine p-nitroanilide dissolved in dimethyl sulfoxide) was quickly added to each well using a multichannel pipette. At 30 \(^\circ{\rm C} ,\) liberation of p-nitroaniline from hydrolysis of the substrate was measured as absorbance at 410 nm every 45 s for 30 min. Trypsin activity was expressed as µmol of p-nitroaniline liberated per minute (U).
For bile-salt dependent lipase, 10 µl of each sample supernatant, 7.5 µg/ml lipase enzyme standard solution (positive control) and homogenization buffer (negative control), were pipetted in triplicate into a 96-well plate resting on an aluminum cooling plate in ice. Quickly, 200 µl of lipase substrate solution (0.35 mM 4-nitrophenyl n-Caproate (4-NPC) dissolved in 70 µl ethanol, 0.5 M Tris–HCl, 6 mM sodium taurochlorate, 1 M NaCl, pH = 7.4) was added to each well. At 30°C, hydrolyzation of 4-NPC was measured at 400 nm absorbance and units were expressed as 1 µM 4-NPC hydrolyzed per minute (U).
Statistical analysis
Water quality parameters among treatments were compared using one-way ANOVA following verification of assumptions of normality using a Shapiro-Wilks test and homogeneity of variance using a Bartlett’s test. Growth rate recorded during the GI characterization study was determined using linear regression. Proportional survival data for each trial was evaluated using the glm function (family = binomial) in R to perform a logistic regression on the Bernoulli distributed survival data (Crawley 2005). Effect of treatment on survival was evaluated using a likelihood ratio test, followed by a post hoc pairwise Tukey HSD test on the log odds of survival if a significant effect of treatment was detected. All survival data are reported as mean percent survival ± standard error (SE) of the proportion. Standard lengths and digestive enzyme activities resulting from different treatment groups were compared using one-way ANOVA following confirmation of assumptions of normality and homogeneity of variance. If a significant effect of treatment on standard length or enzyme activity was detected, a post hoc pairwise Tukey HSD test was used to compare differences between individual treatments. Values for SL and digestive enzyme activity (U) are reported as mean ± standard error of the mean. If individual tanks had zero surviving larvae at the conclusion of the trial, these tanks were omitted from analysis of SL and an unbalanced design was employed. A priori significance levels were set at 〈 = 0.05 and all analyses were completed in Program R (RStudio, V. 0.99.903 2015. RStudio: Integrated Development for R. RStudio, Inc., Boston, MA).
Results
Nutritional trial
Survival varied significantly among treatments following 10 days of initial feeding with either Artemia, MD1, MD2, or MD3 (X24 = 1,187.300, P < 0.001, Fig. 2a). Feeding behavior in each treatment group was confirmed visually. Survival of P. innesi larvae fed Artemia nauplii from first feeding (4 dph) until 14 dph (39.8 ± 3.3%; 4.71 ± 0.09 mm) was significantly higher than the survival resulting from MDs (mean: 1.4 ± 0.6%, P < 0.001).

Proportional survival ± standard error following 10 days of feeding with either newly hatched Artemia nauplii or one of three commercial microparticulate diets: MD1 (Larval AP-100, < 100 µm, Ziegler Bros Inc., Gardners, PA, 50% protein, 12% fat), MD2 (Golden Pearls, 50–100 µm, Brine Shrimp Direct, Ogden, UT, USA, 60% protein, 8% fat), and MD3 (Otohime A1, 75–150 µm, Marubeni Nisshin Feed Co. LTD, Tokyo, Japan, 53% protein, 8% fat). Different letters above columns denote statistically significant differences (P ≤ 0.050)
Digestive ontogeny characterization study
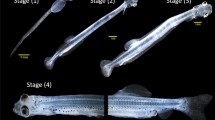
Larval P. innesi growth from 3 to 29 dph was linear (R2 = 0.887) and NL or SL increased at a rate of 0.154 mm/day (Fig. 3). Mean NL at 3 dph was 3.26 ± 0.03 mm and increased to a mean SL of 7.25 ± 0.26 mm at 29 dph. Yolk was present at 3 dph (3.26 ± 0.03 mm) and was nearly completely absorbed by 5 dph (3.61 ± 0.04 mm). The GI tract persisted as a straight, uniform tube divided into esophagus, intestine (including stomach anlage), and rectum from 3 dph (3.26 ± 0.03 mm) until 20 dph (5.24 ± 0.20 mm), when differentiation of the stomach was observed (Fig. 4). The buccopharyngeal cavity and esophagus were characterized by simple squamous epithelial cells and lacked extensive mucous cells prior to 9 dph (3.98 ± 0.07 mm). At 9 dph (3.98 ± 0.07 mm), PAS positive mucous cells began to proliferate and flame cells were apparent in the esophagus, a pattern that persisted throughout the observed developmental period. Intestinal rugae proliferated and deepened at 7 dph (3.50 ± 0.06 mm) in the stomach anlage region. Intestinal epithelial cells were columnar in shape, becoming cuboidal in the stomach region following differentiation. The muscularous layer of the esophageal region began to thicken and was more pronounced following 14 dph (4.71 ± 0.10 mm). While there was a distinct lack of teeth in the mouth region throughout the observed developmental stages, pharyngeal tooth buds appeared in the pharynx at 17 dph (5.56 ± 0.12 mm) and erupted pharyngeal teeth appeared at 20 dph (5.24 ± 0.20 mm). Intestinal epithelial cells exhibited extensive supranuclear vacuoles beginning at 20 dph (5.24 ± 0.20 mm), which proliferated distally throughout the GI. Mucous cells proliferated in the rectal region of the GI beginning at 14 dph. At 20 dph (5.24 ± 0.20 mm), gastric glands were present in the stomach and a pyloric valve separated the stomach and the anterior intestine (Figs. 4 and 5). The GI exhibited a fold initiating between the pyloric region of the stomach and the anterior intestine (Fig. 4c). By 29 dph (7.25 ± 0.26 mm), further folding along the entirety of the GI occurred, as well as proliferation of gastric glands in the lamina propria and the appearance of sections of 6 distinct pyloric caeca (Figs. 4d and 5d). The muscularous layer thickened in conjunction with stomach differentiation at 20 dph. Histochemical preparations with PAS stain revealed a distinct absence of neutral mucopolysaccharides in the stomach anlage prior to gastric gland formation (Fig. 6a). Following the appearance of gastric glands and concurrent stomach differentiation, the apical surface of the stomach epithelium as well as much of the stomach lumen were strongly PAS positive, indicating the presence of neutral mucopolysaccharides in these regions (Fig. 6c, d).

Growth curve for P. innesi fed exclusively newly hatched Artemia nauplii from 3 to 29 days post hatch (dph) from larvae in digestive ontogeny characterization trial. Values are either notochord length (mm, < 20 dph) or standard length (mm, ≥ 20 dph) ± standard error

Photomicrographs of histological preparations (H & E stain) of P. innesi gastrointestinal tract at a) 5 days post hatch (dph, 200x), b) 14 dph (100x), c) 20 dph (100x), and d) 29 dph (100x) from larvae in digestive ontogeny characterization trial. Esophagus (E), stomach anlage (SA), stomach (S), gastric glands (GG), pyloric valve (PV), pyloric caeca (PC), intestine (I), ileo-rectal valve (IR), rectum (R), yolk (Y)

Photomicrographs of histological preparations (H & E stain) of P. innesi gastrointestinal tract at a) 20 dph (400x), b) 29 dph (100x), c) 29 dph (400x), and d) 29 dph (400x) from larvae in digestive ontogeny characterization trial. Esophagus (E), stomach (S), gastric glands (GG), pyloric caeca (PC), Intestine (I)

Photomicrographs of histochemical preparations of P. innesi gastrointestinal tract stained with periodic acid Schiff (PAS) at a) 14 dph (400x), b) 23 dph (200x), and c) 23 dph (400x) from larvae in digestive ontogeny characterization trial. Esophagus (E), PAS + neutral mucopolysaccharides (M), stomach anlage (SA), stomach (S), gastric glands (GG)
While trypsin activity was detectable from 3 dph (3.26 ± 0.03 mm), activity on a per fish basis remained relatively low (δ 144.95 U/fish) until 29 dph (7.25 ± 0.26 mm), when a 6.42-fold increase to 931.06 U/fish occurred (Fig. 7a). Lipase exhibited a similar pattern, with enzyme activities of ≤ 12,655.71 U/fish occurring between 3 (3.26 ± 0.03 mm) and 20 dph (5.24 ± 0.20 mm), followed by sharp increases at 23 (5.87 ± 0.13 mm), 26 (6.88 ± 0.32 mm), and 29 dph (7.25 ± 0.26 mm) to 45,672.15 U/fish (Fig. 7b). Pepsin activity was undetectable until 11 dph (3.99 ± 0.09 mm), when very slight activity (0.342 U/fish) was detected. Pepsin activity was low or not detected (1.58 U/fish) until 29 dph (7.25 ± 0.26 mm), when activity increased 3.78 times relative to the previous peak (5.97 U/fish, Fig. 7c).

Develpoment of digestive enzyme activities [trypsin (a), lipase (b), pepsin (c)] of P. innesi fed exclusively newly hatched Artemia nauplii between 3 and 29 days post hatch (dph) from larvae in digestive ontogeny characterization trial. Errors bars are absent as each data point represents enzyme activity of a pooled sample of ten larvae standardized to enzyme activity per fish
Weaning study
Survival at 32 dph (7.38 ± 0.08 mm) varied significantly among weaning treatments (X25 = 712.64, P < 0.001), with exclusively feeding MD1 from first feeding resulting in the lowest survival (0.75 ± 0.33%). Fish fed exclusively Artemia and those receiving weaning treatments W1 and W2 resulted in similar survival (22.20 ± 1.62%, 21.50 ± 1.72%, and 22.20 ± 1.85%, respectively, P ≥ 0.988). Treatment W3 resulted in higher survival (16.20 ± 1.30%) than MD1, but lower survival than the other treatments (P < 0.001, Fig. 8a). Because survival was exceedingly low for fish fed exclusively MD1, SL and digestive enzyme activities for this treatment group were not included in those analyses. Growth among the remaining treatments was similar (F3 = 1.830, P = 0.142), with an overall final mean standard length of 7.38 ± 0.08 mm (Fig. 8b).

Proportional survival ± standard error (a) and standard length ± standard error (mm, b) at 32 dph following feeding with either exclusively newly hatched Artemia nauplii, exclusively MD3, or one of three weaning regimes: W1 = cofeeding beginning at 12 dph followed by exclusivley MD3at 17 dph; W2 = cofeeding beginning at 17 dph followed by exclusively MD3 at 22 dph; W3 = cofeeding beginning at 22 dph followed by exclusively MD3 at 27 dph. Different letters above columns denote statistically significant differences (P ≤ 0.050)
Trypsin activity varied significantly among treatments (F3 = 3.755, P = 0.049), however, post hoc pairwise comparison did not reveal significant relationships between individual treatments, due to the conservative nature of Tukey HSD for protection against type I error (P ≥ 0.066, Fig. 9a). Mean trypsin activity at 32 dph (7.38 ± 0.08 mm) for all treatments was 1,281.91 ± 124.91 U/fish. Activities for both lipase and pepsin did not vary significantly among treatments (F3 = 0.520, P = 0.678, and X23 = 1.752, P = 0.625, respectively). Mean lipase activity for all treatments at 32 dph (7.38 ± 0.08 mm) was 28,316.35 ± 2,698.40 U/fish and mean pepsin activity was 18.87 ± 1.23 U/fish (Fig. 9b, c).

Digestive enzyme activities [trypsin (a), lipase (b), pepsin (c)] of P. innesi at 32 dph following feeding with either exclusively newly hatched Artemia nauplii or one of three weaning regimes: W1 = cofeeding beginning at 13 dph followed by exclusivley MD3 at 18 dph; W2 = cofeeding beginning at 18 dph followed by exclusively MD3 at 23 dph; W3 = cofeeding beginning at 23 dph followed by exclusively MD3 at 28 dph
Discussion
Evaluation of the larval nutritional trial confirmed that, at least for the three MDs evaluated, P. innesi was incapable of thriving on an MD from first feeding, particularly when compared to larvae fed Artemia (Fig. 2). This pattern was confirmed in the weaning trial, where larvae fed exclusively MD3 from first feeding survived very poorly (0.75 ± 0.33%) relative to all other treatments (Fig. 8a). When taken with the evidence for an extended, agastric larval phase revealed through the physiological characterization of developmental ontogeny (Figs. 4, 5, 6 and 7), it appears that limited digestive capacity due to low digestive enzyme activity and the absence of acidic digestive function likely contributed to this result. However, inherent physical characteristics of each type of diet, including palatability, color, sinking rate, shape of the pellet, etc., may have also contributed to the limited observed success. Further investigations, including nutritional trials evaluating ration and feeding frequency for each diet, are required to optimize feeding protocols for this species.
During early development of larval teleosts there exist three modes of digestive system ontogeny: agastric, precocial, and altricial (Ronnestad et al. 2013). In agastric fishes (i.e. Cyprinidae, Gobiidae), there is a complete reliance on alkaline, pancreatic proteases for hydrolysis of proteins over the entirety of the fishes life, and digestive capacity increases as a function of gut length as the larvae develops, as well as the development of other structures like pharyngeal teeth (Dabrowski 1984; Dabrowski and Poczyczyński 1988; Hur et al. 2016). In precocial fishes (i.e. Salmonidae, Bagridae), larvae develop a fully functioning stomach with concomitant acid protease activity prior to the onset of exogenous feeding, vastly increasing their digestive capacity relative to agastric fishes and allowing for introduction of MDs from first feeding (Dabrowski 1984; Yang et al. 2010). Finally, altricial fishes (i.e. Sparidae, Paralichthyidae, Characidae) exhibit an agastric larval phase, where protease activity is due to alkaline proteases at first feeding until which time the stomach differentiates and gastric digestion begins (Zambonino Infante and Cahu 2007; Ronnestad et al 2013; Yúfera et al. 2018; Nolasco-Soria et al. 2020). In altricial fishes, the successful introduction of MDs in an aquaculture setting largely hinges on the differentiation of a functional stomach, as established by previous authors (Faulk and Holt 2009; Ronnestad et al. 2013; Thompson et al. 2019). While histological evaluation of the GI in P. innesi is absent from the literature, P. axelrodi gastrointestinal physiology has been described in adult samples, which indicate gastric digestive function (Gómez-Ramírez et al. 2011).
Stomach differentiation in P. innesi appeared to occur by 20 dph (5.24 ± 0.20 mm), evidenced by appearance of a pyloric valve, gastric glands, and PAS positive neutral mucopolysaccharide production (Figs. 4, 5 and 6). Pepsin activity increased sharply at 29 dph, indicating a delay between histologically evident organ differentiation and functional gastric digestion, which has been observed in meagre Argyrosomus regius (Solovyev et al. 2016). Also, this is similar timing to the ontogeny of the stomach of another freshwater altricial neo-tropical fish, Hemisorubim platyrhynchos, in which the first gastric glands were observed at 15 dph and the fully differentiated stomach develop at 20 dph (Faccioli et al. 2016). The digestive ontogeny of the Pacu is more accelerated with gastric gland formation occurring at 10–13 dph and functional stomach differentiation occurring from 10–16 dph (Portella et al. 2014). Slight peaks at 11 (3.99 ± 0.09 mm) and 17 dph (5.56 ± 0.12 mm) (Fig. 7c) may be the result of accelerated development of a few fish that were present in the pooled sample of ten fish for the pepsin assays, introducing variation in the sample result. Alternatively, previous authors who observed a similar phenomenon in leopard grouper Mycteroperca rosacea ascribed these early peaks in acid protease activity to other acid enzymes in other regions of the body of the larvae (Martínez-Lagos et al. 2014). Thompson et al. (2019) performed micro-dissections of pigfish Orthopristis chrysoptera larvae, effectively eliminating the potential for this type of false positive, which could have clarified the results in this study. However, given that feeding began at 4 dph and yolk absorption was nearly complete by 5 dph (3.61 ± 0.04 mm) (Fig. 4a), and that there was a complete lack of pepsin activity until 11 dph (3.99 ± 0.09 mm), a period of agastric digestion is apparent.
The presence of PAS positive neutral mucopolysaccharides beginning at 20 dph (5.24 ± 0.20 mm) provides further evidence for the initiation of acidic digestive function beginning at this age. Neutral mucins containing PAS positive mucopolysaccharides have been theorized to be produced by epithelial cells located in the mucosa of the stomach to protect this layer from the activity of the concurrently produced hydrochloric acid and pepsin by the oxynticopeptic cells of the gastric glands (Scocco et al. 1996; Pradhan et al. 2013; Ghosh and Chakrabarti 2015). The mucins in question effectively create a boundary between the acidic environment of the stomach lumen and the active pepsin, protecting the apical surface of the stomach epithelium from autolysis. The production of these compounds signals a functional shift, where acid and pepsin are actively being produced (Scocco et al. 1996).
Despite some species of small bodied, altricial, freshwater fish being capable of digesting MDs at fist feeding (Lipscomb et al 2020a), other studies have implicated a similar pattern of agastric larval digestive function as causative in low survival following first feeding with MDs. Hamlin et al. (2000) described the larval digestive system ontogeny in haddock Melanogrammus aeglefinus, which begin exogenous feeding at 2 dph, but do not exhibit stomach differentiation and gastric gland formation until 33 dph. The authors of this study indicated that weaning M. aeglefinus from live feeds to an MD should be conducted following this transition (Hamlin et al. 2000). Likewise, a study that described digestive enzyme ontogeny and evaluated weaning from live feeds to an MD in larval O. chrysoptera showed that pepsin activity remained low until 30 dph and weaning at 32 dph yielded significantly higher survival than earlier weaning times (Thompson et al. 2019). Additionally, we showed a similar pattern of development in G. ternetzi, a characid relative of P. innesi (Lipscomb et al. 2020b). In the case of G. ternetzi, pepsin activity, gastric gland formation, stomach differentiation and neutral mucopolysaccharide production all occurred at 22 dph, although weaning with cofeeding initiated at 13 dph resulted in similar survival to later weaning times (Lipscomb et al. 2020b).
While P. innesi exhibit histological and histochemical stomach differentiation at 20 dph (5.24 ± 0.20 mm), cofeeding beginning at 13 dph resulted in similar survival and growth to fish fed exclusively Artemia from first feeding through 32 dph, as well as cofeeding initiated at 17 dph. Given that cofeeding ceased and exclusive feeding with MD3 occurred at 17 dph for weaning treatment W1, it appears that weaning to an MD prior to completion of stomach formation is possible for P. innesi (Fig. 8). This may be explained by the increased activity of lipase and a slight increase in pepsin that were concurrent with this weaning period (Fig. 7).
Alternatively, authors have suggested that initiating larval nutrition with live feeds may infer digestive benefits due to an increase the production of endogenous enzymes in larval fish (Hjelmeland et al. 1988). Hjelmeland et al. (1988) observed that ingestion of either copepods or Artemia in larval herring Clupea harengus elicited higher levels of trypsin in the intestine than inert particles, although the inert particles did elicit some response. This interaction between live prey and larval digestive function could help explain the observed results of successful weaning prior to complete stomach differentiation in P. innesi. Curiously, larvae weaned beginning at 23 dph (treatment W3; 5.87 ± 0.13 mm) exhibited reduced survival relative to those weaned at 13 and 18 dph (treatments W1 and W2, respectively Fig. 8a). While further research is required to elucidate the cause of this result, one possible explanation is that the prolonged live feed phase in W3 resulted in difficulty in transitioning to an inert particle, potentially due to habituation to Artemia.
Timing of weaning in P. innesi did not significantly impact pepsin or lipase activities, and although a significant effect on trypsin activity was detected (F3 = 3.755, P = 0.049), pairwise comparisons did not reveal differences between individual treatments (Fig. 9). Given that samples were taken after a period of overnight fasting, positive feedback interactions between ingested feed items and enzyme activity were likely limited. This means that observed digestive enzyme activities can be interpreted as basal zymogen production, and not acutely modulated by ingesta. This observation agrees with the previous assertion by Zambonino Infante, and Cahu (2001) that diet may modulate peaks in enzyme activities following ingestion, but timing of increases and decreases in activity is largely genetically determined.
The results of this study show that live feed dependence of P. innesi at first feeding is likely due to an altricial pattern of GI development, where the stomach and associated functions are absent during the initiation of exogenous feeding, and do not appear until between 11 (3.99 ± 0.09 mm) and 20 dph (5.24 ± 0.20 mm). Weaning prior to the differentiation of the stomach organ and appearance of gastric glands, pyloric valve, and PAS positive neutral mucins is possible, with weaning beginning at 13 dph inferring similar survival to exclusively feeding Artemia. This result is consistent with previous work with another characid, G. ternetzi, which potentially reveals a pattern of digestive function common to small-bodied characids, although further research into more species is warranted (Lipscomb et al. 2020b). In conclusion, commercial production of P. innesi can be accomplished with minimal live feed inputs if the early weaning protocols established herein are implemented. This potentially increases the resilience of this aquaculture endeavor in periods of Artemia shortage, as well as provides an overall economic advantage for the critical larval culture stage in this ornamental fish.
Data availability
The datasets generated during and/or analyzed during the current study are not publicly available but are available from the corresponding author on reasonable request.
Code availability
The statistical analyses and figures herein were created in Program R (RStudio, V. 0.99.903 2015. RStudio: Integrated Development for R. RStudio, Inc., Boston, MA). The custom code is available upon reasonable request.
References
Cahu C, ZamboninoInfante J (2001) Substitution of live food by formulated diets in marine fish larvae. Aquaculture 200:161–180. https://doi.org/10.1016/S0044-8486(01)00699-8
Chao NL (2001) The fishery, diversity, and conservation of ornamental fishes in the Rio Negro Basin, Brazil - A review of Project Piaba (1989–99). In conservation and management of ornamental fish resources of the Rio Negro Basin, Amazonia, Brazil: Project Piaba. Ning L Chao, et al, eds. Manaus, Amazonas: Editora Universidade do Amazonas. pp 161–204
Chapman FA, Colle DE, Rottmann RW, Shireman JV (1998) Controlled Spawning of the Neon Tetra. Progress Fish-Culturist 60:32–37. https://doi.org/10.1577/1548-8640(1998)060%3c0032:CSOTNT%3e2.0.CO;2
Chapman FA, Fitz-Coy SA, Thunberg EM, Adams CM (1997) United States of America trade in ornamental fish. J World Aquac Soc 28:1–10. https://doi.org/10.1111/j.1749-7345.1997.tb00955.x
Çinar K, Şenol N (2006) Histological and histochemical characterization of the mucosa of the digestive tract in flower fish (Pseudophoxinus antalyae). J Vet Med Ser C Anat Histol Embryol 35:147–151. https://doi.org/10.1111/j.1439-0264.2005.00629.x
Crawley MJ (2005) Statistics: an introduction using R. John Wiley & Sons, West Sussex, England
Dabrowski K, Poczyczyński P (1988) Comparative experiments on starter diets for grass carp and common carp. Aquaculture 69:317–332. https://doi.org/10.1016/0044-8486(88)90339-0
Dabrowski KR (1984) The feeding of fish larvae: present (state of the art) and prespective. Reprod Nutr Dév 24:807–833. https://doi.org/10.1051/rnd:19840701
Faccioli CK, Chedid RA, Mori RH et al (2016) Organogenesis of the digestive system in Neotropical carnivorous freshwater catfish Hemisorubim platyrhynchos (Siluriformes: Pimelodidae). Aquaculture 451:205–212. https://doi.org/10.1016/j.aquaculture.2015.09.009
Faulk CK, Benninghoff AD, Holt GJ (2007) Ontogeny of the gastrointestinal tract and selected digestive enzymes in cobia Rachycentron canadum (L.). J Fish Biol 70:567–583. https://doi.org/10.1111/j.1095-8649.2007.01330.x
Faulk CK, Holt GJ (2009) Early weaning of southern flounder, Paralichthys lethostigma, larvae and ontogeny of selected digestive enzymes. Aquaculture 296:213–218. https://doi.org/10.1016/j.aquaculture.2009.08.013
Ghosh SK, Chakrabarti P (2015) Histological and histochemical characterization on stomach of Mystus cavasius (Hamilton), Oreochromis niloticus (Linnaeus) and Gudusia chapra (Hamilton): Comparative study. J Basic Appl Zool 70:16–24. https://doi.org/10.1016/j.jobaz.2015.04.002
Gómez-Ramírez E, Obando M, Tovar MO et al (2011) Histological Study of Digestive Tract of the Neon Cardenal Paracheirodon axelrodi (Characidae). Int J Morphol 29(3):782–786
Govoni JJ, Boehlert GW, Watanabe Y (1986) The physiology of digestion in fish larvae. Environ Biol Fishes 16:59–77. https://doi.org/10.1007/BF00005160
Hamlin HJ, Hunt Von Herbing I, Kling LJ (2000) Histological and morphological evaluations of the digestive tract and associated organs of haddock throughout post-hatching ontogeny. J Fish Biol 57:716–732. https://doi.org/10.1006/jfbi.2000.1347
Hamre K, Yúfera M, Rønnestad I et al (2013) Fish larval nutrition and feed formulation: Knowledge gaps and bottlenecks for advances in larval rearing. Rev Aquac 5:1–87. https://doi.org/10.1111/j.1753-5131.2012.01086.x
Hjelmeland K, Pedersen BH, Nilssen EM (1988) Trypsin content in intestines of herring larvae, Clupea harengus, ingesting inert polystyrene spheres or live crustacea prey. Mar Biol 98:331–335. https://doi.org/10.1007/BF00391108
Holt GJ (2011) Larval Fish Nutrition. John Wiley & Sons, Hoboken, NJ, USA
Hur S-W, Kim S-K, Kim D-J et al (2016) Digestive Physiological Characteristics of the Gobiidae: - Characteristics of CCK-producing Cells and Mucus-secreting Goblet Cells of Stomach Fish and Stomachless Fish. Dev Reprod 20:207–217. https://doi.org/10.12717/DR.2016.20.3.207
Kolkovski S (2013) Microdiets as alternatives to live feeds for fish larvae in aquaculture: Improving the efficiency of feed particle utilization. In: Allan G, Burnell G (eds) Advances in Aquaculture Hatchery Technology. Elsevier Science & Technology, Philadelphia, PA, pp 203–222
Kolkovski S (2001) Digestive enzymes in fish larvae and juveniles—implications and applications to formulated diets. Aquaculture 200:181–201. https://doi.org/10.1016/S0044-8486(01)00700-1
Kucharczyk D, Targońska K, Zarski D et al (2010) The reproduction of neon tetra, paracheirodon innesi (Myers, 1936), under controlled conditions. Polish J Nat Sci 25:81–92. https://doi.org/10.2478/v10020-010-0006-x
Lim LC, Dhert P, Sorgeloos P (2003) Recent developments in the application of live feeds in the freshwater ornamental fish culture. Aquaculture 227:319–331. https://doi.org/10.1016/S0044-8486(03)00512-X
Lipscomb TN, Patterson JT, Wood AL, Watson CA, DiMaggio MA (2020a) Larval growth, survival, and partial budget analysis related to replacing Artemia in larval culture of six freshwater ornamental fishes. J World Aquac Society 51(5):1132–1144
Lipscomb TN, Yanong RP, Ramee SW, DiMaggio MA (2020b) Histological, histochemical and biochemical characterization of larval digestive system ontogeny in black tetra Gymnocorymbus ternetzi to inform aquaculture weaning protocols. Aquaculture 520:734957. https://doi.org/10.1016/j.aquaculture.2020.734957
Martínez-Lagos R, Tovar-Ramírez D, Gracia-López V, Lazo JP (2014) Changes in digestive enzyme activities during larval development of leopard grouper (Mycteroperca rosacea). Fish Physiol Biochem 40:773–785. https://doi.org/10.1007/s10695-013-9884-5
Nolasco-Soria H, Nolasco-Alzaga HR, Gisbert E (2020) The importance of pepsin-like acid protease quantification in aquaculture studies: a revision of available procedures and presentation of a new protocol for its assessment. Rev Aquac 12(3):1928–1943
Portella MC, Jomori RK, Leitão NJ et al (2014) Larval development of indigenous South American freshwater fish species, with particular reference to pacu (Piaractus mesopotamicus): A review. Aquaculture 432:402–417. https://doi.org/10.1016/j.aquaculture.2014.04.032
Pradhan PK, Jena J, Mitra G et al (2013) Ontogeny of the digestive enzymes in butter catfish Ompok bimaculatus (Bloch) larvae. Aquaculture 372–375:62–69. https://doi.org/10.1016/j.aquaculture.2012.10.024
Quintero-Hunter I, Grier H, Muscato M (1991) Enhancement of histological detail using metanil yellow as counterstain in periodic acid Schiff’s hematoxylin staining of glycol methacrylate tissue sections. Biotech Histochem 66:169–172
Rønnestad I, Tonheim SK, Fyhn HJ et al (2003) The supply of amino acids during early feeding stages of marine fish larvae: a review of recent findings. Aquaculture 227:147–164. https://doi.org/10.1016/S0044-8486(03)00500-3
Ronnestad I, Yúfera M, Ueberschär B et al (2013) Feeding behaviour and digestive physiology in larval fish: current knowledge, and gaps and bottlenecks in research. Rev Aquac 5:S59–S98. https://doi.org/10.1111/raq.12010
Sanaye SV, Dhaker HS, Tibile RM, Mhatre VD (2014) Effect of Green Water and Mixed Zooplankton on Growth and Survival in Neon Tetra, Paracheirodon innesi (Myers, 1936) during Larval and Early Fry Rearing. Int J Biol Biomol Agric Food Biotechnol Eng 8:159–163
Scocco P, Ceccarelli P, Menghi G (1996) Glycohistochemistry of the Tilapia spp. stomach. J Fish Biol 49:584–593. https://doi.org/10.1006/jfbi.1996.0188
Siccardi AJ, Garris HW, Jones WT et al (2009) Growth and survival of Zebrafish (Danio rerio) fed different commercial and laboratory diets. Zebrafish 6:275–280. https://doi.org/10.1089/zeb.2008.0553
Solovyev MM, Campoverde C, Öztürk S et al (2016) Morphological and functional description of the development of the digestive system in meagre (Argyrosomus regius): An integrative approach. Aquaculture 464:381–391
Thompson KL, Faulk CK, Fuiman LA (2019) Applying the ontogeny of digestive enzyme activity to guide early weaning of pigfish, Orthopristis chrysoptera (L.). Aquac Res 50:1404–1410. https://doi.org/10.1111/are.14015
Weitzman SH, Weitzman SH, Fink WL (1983) Relationships of the neon tetras, a group of South American freshwater fishes (Teleostei, Characidae), with comments on the phylogeny of New World characiforms. Bull Museum Comp Zool Harvard Coll 150:339–395
Yang R, Xie C, Fan Q et al (2010) Ontogeny of the digestive tract in yellow catfish Pelteobagrus fulvidraco larvae. Aquaculture 302:112–123. https://doi.org/10.1016/j.aquaculture.2010.02.020
Yúfera M, Moyano FJ, Martínez-Rodríguez G (2018) The digestive function in developing fish larvae and fry from molecular gene expression to enzymatic activity. In: Yúfera M (ed) Emerging Issues in Fish Larvae Research. Springer, Cham
ZamboninoInfante JL, Cahu CL (1994) Influence of diet on pepsin and some pancreatic enzymes in sea bass (Dicentrarchus labrax) larvae. Comp Biochem Physiol 109A:209–212. https://doi.org/10.1016/0300-9629(94)90122-8
ZamboninoInfante JL, Cahu CL (2001) Ontogeny of the gastrointestinal tract of marine fish larvae. Comp Biochem Physiol Part C Toxicol Pharmacol 130:477–487. https://doi.org/10.1016/S1532-0456(01)00274-5
ZamboninoInfante JL, Cahu CL (2007) Dietary modulation of some digestive enzymes and metabolic processes in developing marine fish: applications to diet formulation. Aquaculture 268(1–4):98–105
Acknowledgements
The authors acknowledge Noretta Perry of the Fish and Wildlife Research Institute, Florida Fish and Wildlife Conservation Commission, for histological and histochemical processing. In addition, we acknowledge VW Fish Hatcheries Inc, Lakeland, FL, for providing fish for this research.
Funding
The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
TL conceptualized the research, designed and conducted the experiments, conducted associated assays, analyzed resulting data and wrote the manuscript. RY facilitated histological processing, interpreted histological slides, and reviewed/edited the manuscript. SR assisted in conducting the experiments, conducted associated assays, and reviewed/edited the manuscript. MD conceptualized the research, assisted in conducting the experiments, and reviewed/edited the manuscript.
Corresponding author
Ethics declarations
Ethics approval
All research herein was conducted under approval by and in compliance with standards developed by the Institutional Animal Care and Use Committee of the University of Florida (IACUC study #202011169).
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflicts of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Lipscomb, T.N., Yanong, R.P., Ramee, S.W. et al. Larval digestive system ontogeny and early weaning in neon tetra Paracheirodon innesi. Fish Physiol Biochem 49, 1241–1255 (2023). https://doi.org/10.1007/s10695-023-01254-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10695-023-01254-w