
Abstract
This study was carried out to evaluate whether silymarin supplementation influences growth, lipid metabolism, and health status in grass carp fed elevated dietary lipid levels. The juvenile fish (27.43 ± 0.17 g/tail) were fed six isonitrogenous and isocaloric diets in a factorial design containing 0, 100, or 200 mg kg−1 silymarin (SM0, SM100, SM200) associated with either 4 or 8 % lipid level (low lipid, LL, and high lipid, HL, respectively) for 82 days. The results showed that both dietary silymarin supplementation and high lipid level significantly enhanced growth performance (WG, SGR), protein efficiency ratio, and feed utilization. Silymarin supplementation significantly reduced the VSI, hepatic lipid content, and the total bilirubin concentration in the serum. The gallbladdersomatic index displayed higher in the SM100 groups than SM200 groups. Serum total cholesterol content exhibited lower in the SM100 groups than SM0 groups. Meanwhile, significant interactions were shown for hepatic gene expression of HSL and CPT1 by two factors, and SM100 group had higher hepatic gene expression of HSL and CPT1 in fish fed with the HL diets. The SM100 groups up-regulated hepatic gene expressions of HMGCR and CYP7A1 compared with the SM0 groups. Silymarin supplementation notably reduced the elevated serum MDA content induced by HL treatments. Thus, silymarin supplementation markedly promoted growth and protein efficiency, suppressed lipid accumulation, and improved health status in grass carp fed with high-lipid diets, which might be associated with its enhancement of lipolysis and β-oxidation, antioxidant capacity.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Silymarin, a flavonolignan from the milk thistle (Silybum marianum L.), is mainly composed of four isomeric mixtures of active flavonolignans, including silibinin A and B, isosilibinin A and B, silychristin, and silydianin (Colturato et al. 2012). Silymarin has been employed as an antioxidant due to its anti-lipid peroxidative and anti-inflammatory effects and has been used in the treatment of various liver diseases (Chen et al. 2012; El-Lakkany et al. 2012; Surai 2015; Yao et al. 2011). However, some studies have reported that silymarin and silibinin may reduce fat deposition in the viscera of rats by suppressing lipogenic gene expression and/or enhancing lipolysis and hypolipidemic effects by regulating plasma lipoproteins, such as LDL, VLDL, and HDL (Chou et al. 2012; El-Sheikh and Abd El-Fattah 2011; Metwally et al. 2009; Škottová et al. 2003; Sobolová et al. 2006; Vecera et al. 2011; Yao et al. 2013). In fish, few reports on the application of silymarin are available. The available studies have indicated that silymarin supplementation in diets boosts growth performance in Carassius auratus gibelio (Yi et al. 2012), reduces plasma cholesterol in rainbow trout (Banaee et al. 2011), prevents liver damage induced by CCl4 in common carp (Jia et al. 2013), and strengthens the immune system in rainbow trout and the antioxidative stress capacity of SHK-1 cells in Atlantic salmon (Ahmadi et al. 2012; Sanchez et al. 2016). In addition, silibinin has anti-adipogenic effects on 3T3-L1 cells and in zebrafish (Ka et al. 2009; Suh et al. 2015). However, to our knowledge, little is known about the effects of silymarin as a promising feed additive on the improvement of growth, lipid metabolism, and health status in fish fed the diets with elevated lipid levels.
Lipids are an important source of energy for growth in fish (Tocher 2003). Some studies have shown that increasing dietary lipids induces rapid growth and efficient feed conversion in several fish species (Steffens 1996; Vergara et al. 1996; Wang et al. 2005), indicating that lipids exhibit protein-sparing effects (Ai et al. 2004; Dias et al. 1998; Morais et al. 2001). However, a dietary lipid intake surpassing an upper limit in some fish species not only reduces growth performance but also leads to unwanted fat accumulation in the liver or other tissues and can lead to hyperlipidemia, fatty liver, and lipid peroxidation (Du et al. 2005, 2008; Ji et al. 2011; Jin et al. 2013; Li et al. 2012; Lu et al. 2013a; Stéphan et al. 1995). Increases in lipid peroxidation also increase the occurrence of fatty liver (Rolo et al. 2012). Fatty liver not only represents wasted energy but also affects the health of fish (Lu et al. 2013b; Nanton et al. 2003).
The grass carp, a typical herbivorous finfish, is one of the major freshwater aquaculture species in China, with the production of 5.37 million tons in 2014, accounting for 20.65 % of freshwater fish production in China (Fisheries Bureau, Ministry of Agriculture of China 2015). Some studies have shown that grass carp present a relatively low lipid utilization capacity and that 6 % higher lipid levels in the diets induce negative effects on growth and body lipid deposition (Du et al. 2005, 2006; Gao et al. 2011). Because of the availability of inexpensive oil sources, the use of lipid-rich feeds is a current trend in intensive aquaculture. However, compared with wild grass carp, feeding higher lipid diets to artificially reared grass carp is often associated with fatty liver, which correlates closely with a high mortality rate and poor growth (Lu et al. 2013b). Based on previous findings regarding the effects of silymarin, we hypothesized that dietary silymarin might reduce the negative side effects of a high lipid intake in grass carp.
Therefore, the present study aimed to investigate whether supplementation of silymarin in diets could enhance the utilization of dietary lipids, based on examining growth performance, serum biochemical values and antioxidative indexes, the hepatic composition, and lipid metabolism-related gene expression levels in grass carp. We propose a practical approach to silymarin supplementation for improving the growth performance and promoting the lipid metabolism and health status of fish.
Materials and methods
Experimental diets
Six isonitrogen (Crude protein 32 %) and isoenergy (Gross energy 19.5 MJ kg−1) semi-purified experimental diets were formulated and randomized design with a 3 × 2 factorial arrangement, each containing three silymarin concentrations at 0, 100, or 200 mg kg−1 diets (SM0, SM100, and SM200) associated with either 4 or 8 % lipid level (low lipid, LL, and high lipid, HL, respectively). Levels of corn starch and cellulose were adjusted to maintain isocaloric diets. Silymarin was kindly provided by Beijing sunpu biochem. Teck.co., Ltd. (Beijing, China). Silymarin content was 77.8 % measured by UV spectrophotometry, which consisted of a large number of flavonolignans. According to high-performance liquid chromatography (HPLC) analysis, silymarin contained 20.49 % of silibinin A and B, 6.05 % of isosilibinin A and B, 3.38 % of silidianin, and 10.30 % of silicristin.
Ground ingredients were thoroughly blended in a food mixer. Powdered silymarin was mixed with a vitamin mixture to achieve uniformity and was then added to the blend, followed by homogenization. Next, soybean oil was added, followed by homogenization. Finally, distilled water was added to create a dough, and the mixture was further homogenized and extruded through a 2-mm-diameter die in a meat grinder. Pellets were air-dried at 20 °C and then stored at −20 °C in a freezer until feeding. The analyzed proximate composition of the experimental diets is presented in Table 1.
Experimental fish
Grass carp (mean initial weight, 27.43 ± 0.17 g) were obtained from Hua cheng aquaculture farm (Jiangsu, China) and reared in 300-L aquaria in a closed recirculation system with mechanical filtration in the aquaculture base of School of Basic Medicine and Biological Science, Soochow University. Before the experiment, all fish were acclimatized to indoor conditions for 2 weeks with the feeding of basal diet (LL-SM0). A total of 360 grass carp were randomly distributed into 24 fiberglass tanks, with 15 fish per tank for 82 days. Each treatment was randomly assigned to 4 replicate tanks. The feeding rate was 2.5–3 % wet body weight per day. Daily feed ration was daily adjusted depending on the feed consumed by fish to be kept close to apparent satiation. In addition, fish from two randomly selected tanks per treatment were group-weighed every other week, and daily feed rations were adjusted accordingly. The pre-weighed daily feed ration was divided into two equal portions and administered to fish at 08:00 and 16:00 h, respectively.
During the experimental period, air blowers provided aeration via air stones to maintain dissolved oxygen >6.0 mg L−1. Water temperature was ranged from 25.0 to 28.1 °C. Water quality parameters (total ammonia <0.1 mg L−1, nitrite <0.1 mg L−1, and pH 7.0–7.4) were measured weekly to ensure the safe levels, respectively. Photoperiod was maintained at 12:12-h light–dark schedule.
Sampling and chemical analysis
Sample collection
At the end of growth trial, fish were starved for 24 h and then anesthetized with tricaine methanesulfonate (MS222) at 100 mg L−1. All fish were counted and weighed, and then each fish was weighted individually and body length measured. Blood was randomly sampled from six to eight fish each tank. After centrifugation (825×g, 10 min, 4 °C), serum samples of two or three fish were pooled and stored frozen at −80 °C until analysis. Two pooled serum samples for each tank were tested. After blood withdrawal, viscera, hepatopancreas, and gallbladder were dissected out and weighed individually to calculate viscerosomatic index (VSI), hepatosomatic index (HSI), and gallbladdersomatic index (GI). Hepatopancreas tissues from six fish per tank were pooled and then stored at −20 °C for proximate analysis. Hepatopancreas from another three fish were rapidly excised, frozen in liquid nitrogen, and stored at −80 °C for genes expression. At the same time, part of fresh liver samples was analyzed for total cholesterol (T-cho) contents. The procedures were carried out in accordance with the EU Directive 2010/63/EU for animal experiments.
To evaluate the growth performance of fish and feed utilization, the following variables were calculated:
-
Weight gain (WG, %) = [final average body weight (g) − initial average body weight (g)] × 100/initial average body weight (g)
-
Specific growth rate (SGR, % d−1) = [Ln final average body weight (g) − Ln initial average body weight (g)] × 100/days
-
Feed conversion ratio (FCR) = amount of wet feed intake (g)/weight gain (g)
-
Day feed intake (DFI, % d−1) = total wet feed intake (g) × 100/((initial weight (g) + final weight (g))/2)/days
-
Protein efficiency ratio (PER) = (final weight (g) − initial weight (g))/[(feed intake (g) × wet feed protein content (g 100 g−1)]
-
Protein production value (PPV, %) = (wet fish protein gain (g)) × 100/(wet protein intake (g))
-
Lipid retention (LR, %) = (wet fish lipid gain (g)) × 100/(wet lipid intake (g))
-
Survival rate (SR, %) = final number of fish × 100/initial number of fish
-
Condition factor (CF, %) = wet body weight × 100/body length3
-
Viscerosomatic index (VSI, %) = viscerosomatic weight × 100/wet body weight
-
Gallbladdersomatic index (GI, %) = gallbladder weight × 100/wet body weight
-
Hepatosomatic index (HSI, %) = hepatopancreas weight × 100/wet body weight
Proximate composition analysis
Composition analyses of diets and hepatopancreas were made following standard methods (AOAC 1998). Samples were dried to a constant weight at 105 °C for 24 h to determine moisture. Crude protein was determined by measuring nitrogen (N × 6.25) using the Kjeldahl method after acid digestion (KDN-04 III, Shanghai XianJian Instruments Co., Ltd., Shanghai, China; LNK-872 versatile combustion, Yixing Science and Instrument Research Institute, Wuxi, China). Crude lipid content was determined by the petroleum ether extraction method for 12 h using a Soxhlet system (Yixing Science and Instrument Research Institute, Wuxi, China). Ash content was determined after by incineration at 550 °C for 12 h. Gross energy of feed was analyzed by Oxygen Bomb Calorimeter (XRY-1C, Shanghai Changji Geological Instrument Co., Ltd, China).
Serum biochemical parameters and hepatic total cholesterol contents
Serum biochemical parameters including triacylglycerol, glucose, total cholesterol, low-density and high-density lipoprotein cholesterol, and total bilirubin (TG, Glu, T-cho, LDL-c, LDL-c, and TBil, respectively) were assayed by enzymatic procedure using automatic biochemical analyzer and attached kit (Abbott Architec, C8000, Princeton, New Jersey, USA) from a clinic laboratory of Suzhou Jiulong hospital. Nonesterified fatty acid (NEFA), superoxide dismutase (SOD), and malondialdehyde (MDA) contents in the serum were measured with commercial assay kits (Nanjing Jiancheng Biochemical Corporation, Nanjing, China).
Hepatic T-cho contents were measured with commercial assay kits (Applygen Technologies Inc., Beijing, China). Soluble protein contents of liver homogenates were determined by the bicinchoninic acid assay using bovine serum albumin as a standard (BCA Kit, Nanjing Jiancheng Biochemical Corporation, Nanjing, China).
Determination of gene expression by quantitative real-time PCR
Total RNA extraction of hepatopancreas using RNAiso Plus (TaKaRa, Otsu, Shiga, Japan) was performed by the specification of the manufacturer. Agarose gel electrophoresis and spectrophotometer (NanoDrop 1000, Thermo Scientific, Wilmington, USA) were used to check the RNA integrity. Total RNA per sample was reverse-transcribed into cDNA using the PrimeScript™ RT reagent Kit with gDNA eraser (TaKaRa, Otsu, Shiga, Japan), following the manufacturer’s instructions. Total RNA was treated with 0.5 μL gDNA Eraser (Takara, Otsu, Shiga, Japan) for 2 min at 42 °C according to the manufacture’s instructions. The DNase-treated total RNA (500 ng) was used for cDNA synthesis in a 10 μL reaction volume containing 2 μL 5 × prime script buffer, 0.5 μL RT prime mix, 0.5 μL prime script RT enzyme mix I, 2 μL RNase free H2O. The cDNA was synthesized for 15 min at 37 °C followed by 5 s at 85 °C and 10 min at 4 °C. The qRNA amplifications were carried out in triplicate (CFX 96 Real-Time PCR Detection System, Bio-Rad, California, USA) in a final volume of 20 μL containing 3 μL of each primer (4 μM), 1 μL diluted first-strand cDNA product, 10 μL 2 × SYBR® Premix Ex TaqTM II (TaKaRa, Otsu, Shiga, Japan), and 6 μL sterilized double-distilled water. The qPCR profiles contained an initial activation step at 95 °C for 30 s followed by 40 cycles of 95 °C for 5 s and 60 °C for 30 s. The primer sequences for β-actin, FAS, ACC, ATGL, HSL, CPT1, HMGCR, and CYP7A1 are listed in Table 2. After the PCR, melting curve was analyzed to confirm that a single product was present in these reactions. β-actin mRNA was used as the internal control. A relative quantification method was used to calculate the gene expression values using the comparative CT method (2−ΔΔCt) described in the literature (Livak and Schmittgen 2001; Pfaffl 2001).
Statistical analysis
Value of parameters assayed was presented as the mean. All data were analyzed by the analysis of variance (ANOVA) using the general linear model procedure of the PASW Statistics (version 18.0, Chicago, IL, USA). Two-way ANOVA and Duncan’s new multiple range tests were performed to assess the effect of dietary lipid level, silymarin supplementation, and the interaction of two variables. Differences were considered significant at P < 0.05. The graphs were created using Prism 6 (GraphPad Software Inc., San Diego, USA).
Results
Growth performance
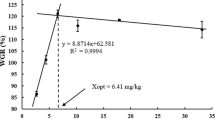
Growth performance and feed efficiency were strongly affected by dietary silymarin supplementation and lipid levels, as shown in Table 3. Both the SM100 and SM200 groups displayed significantly higher WG, SGR, and PER and lower FCR compared with the SM0 groups. The HL diets significantly increased FBW, WG, SGR, PER, and PPV and decreased FCR and LR compared with the LL diets. In addition, no significant interactions between dietary silymarin supplementation and lipid levels were observed for WG, SGR, PER, PPV, FCR, and LR. DFI and SR were not affected by dietary silymarin supplementation, lipid levels, or their interaction.
Biological indices
The biological indices of grass carp fed experimental diets are reported in Table 4. Both the SM100 and SM200 groups exhibited significantly lower VSI compared with the SM0 groups. GI was markedly affected by silymarin supplementation, and GI was significantly higher in fish from the SM100 groups than those from the SM200 groups. Despite no statistical differences, HSI presented a decreasing trend with silymarin supplementation in diets by using two-way ANOVA. Additionally, the SM200 group had a significantly lower HSI than the SM0 group under the HL treatments. Moreover, increasing lipid levels resulted in significant increases in VSI and GI and a decrease in HSI. No significant interactions between dietary silymarin supplementation and lipid levels were observed for VSI, HSI, and GI. CF was not affected by the dietary treatments.
Hepatic proximate composition and T-cho contents
The hepatic proximate composition and T-cho content data for grass carp fed the experimental diets are reported in Table 5. Both the SM100 and SM200 groups showed significantly reduced hepatic lipid contents compared with the SM0 groups. The hepatic lipid contents were much lower under the HL treatments than the LL treatments. The moisture, protein, and T-cho contents in the hepatopancreas were not affected by dietary silymarin supplementation, lipid levels, or their interactions, although there appeared to be an elevated trend of T-cho with increased silymarin levels (P = 0.067).
Serum biochemical parameters
The serum biochemical parameter data for grass carp fed different experimental diets are shown in Table 6. The serum T-cho and TBil contents were affected by dietary silymarin supplementation. Compared with the SM0 groups, the serum T-cho content was markedly lower in fish fed diets with SM100 groups, and the serum TBil content was significantly decreased in both the SM100 and SM200 groups. The serum LDLc content in fish given feed with dietary silymarin supplementation showed a declining trend (P = 0.0561). However, increasing dietary lipid levels significantly elevated the concentrations of TG, NEFA, and Glu in the serum, but markedly decreased serum T-cho and LDLc contents. There was a decreasing tendency of serum HDLc contents with increasing dietary lipid levels (P = 0.0789). In addition, the serum TG, NEFA, HDLc, and Glu concentrations were not affected by silymarin supplementation, and the serum TBil content was not influenced by dietary lipid levels. No significant interactions between any two factors were found for any of the biochemical indices in the serum.
Antioxidant capacity and lipid peroxidation status
The serum SOD and MDA values of grass carp fed the experimental diets are summarized in Table 7 and Fig. 1. Serum SOD activities were not affected by dietary silymarin supplementation, lipid levels, or their interaction. The serum MDA concentration was higher under the HL treatments than the LL treatments, but notably reduced by dietary silymarin supplementation.

Serum SOD activities (A) and MDA content (B) in grass carp fed the six experimental diets for 82 days subjected to two-way ANOVA showing the effects of the two factors and their interaction. For each lipid level, values indicated with different letters are significantly different; lowercase or uppercase letters correspond to the LL or HL treatments, respectively. Within each silymarin level, significant differences between the LL and HL treatment values are marked with an asterisk. HL high lipid, 8 % lipid level, LL low lipid, 4 % lipid level, SM0 no silymarin supplementation, SM100 silymarin supplementation at 100 mg kg−1, SM200 silymarin supplementation at 200 mg kg−1. n = 4 per group
Expression of lipid metabolism-related genes
The effects of dietary silymarin supplementation and different lipid levels on the hepatic mRNA levels of lipid metabolism-related genes of grass carp are shown in Fig. 2. The hepatic mRNA expression of FAS and ACC was significantly down-regulated in the HL treatments, but not affected by silymarin supplementation (Fig. 2A, B). Significant interactions between silymarin supplementation and dietary lipid levels were observed for the expression of HSL mRNA (Fig. 2D). The expression of CPT1 mRNA was markedly influenced by both silymarin supplementation and their interaction (Fig. 2E). The SM100 groups showed markedly higher mRNA expression of HSL compared with the SM0 and SM200 groups under the HL treatments, while the HL treatments resulted in notably higher mRNA expression of HSL compared with the LL treatments in the SM100 groups (Fig. 2D). Both SM100 and SM200 groups displayed dramatically higher mRNA expression of CPT1 compared with the SM0 group under the HL treatments. The gene expression of CPT1 mRNA was significantly down-regulated under the HL treatments compared with the LL treatments in the SM0 groups, but notably up-regulated under the HL treatments compared with the LL treatments for the SM200 groups (Fig. 2E). Hepatic mRNA expression of HMGCR and CYP7A1 was only affected by dietary silymarin supplementation (Table 7, Fig. 2F, G). The SM100 and SM200 groups notably up-regulated gene expressions of HMGCR compared with the SM0 groups. The SM100 groups resulted in higher hepatic mRNA expression of CYP7A1 than the SM0 groups. Hepatic mRNA expression of ATGL was not influenced by the dietary treatments (Fig. 2C).

Results of quantitative real-time PCR analysis carried out for genes associated with FAS, ACC, ATGL, HSL, CPT1, HMGCR, and CYP7A1 (A–G) in the hepatopancreas of grass carp, subjected to two-way ANOVA, showing the effects of the two factors and their interaction. For each lipid level, values indicated with different letters are significantly different; lowercase or uppercase letters correspond to the LL or HL, respectively. Within each silymarin level, significant differences between LL and HL values are marked with an asterisk. HL, high lipid, 8 % lipid level; LL, low lipid, 4 % lipid level; SM0, no silymarin supplementation; SM100, silymarin supplementation at 100 mg kg−1; SM200, silymarin supplementation at 200 mg kg−1. n = 12 per group
Discussion
As feed additives, plant extracts have been shown to promote growth performance, act as antioxidants, and improve the immune system in fish. These effects are due to the active components of these extracts, such as flavonoids, phenolics, and pigments (Ahmadi et al. 2012; Banaee et al. 2011; Cao et al. 2015; Citarasu 2010). A significant growth-promoting effect of dietary silymarin supplementation was observed in this study. Specifically, dietary silymarin supplementation resulted in higher WG, SGR, and PER, and lower FCR, indicating that silymarin supplementation improved feed efficiency and protein utilization (Table 3). Similar effects were previously observed in Carassius auratus gibelio (Yi et al. 2012) and common carp (Jia et al. 2013), suggesting a beneficial effect of silymarin supplementation.
Increasing dietary lipid levels also improved several growth parameters, including FBW, WG, SGR, PER, and PPV, and reduced FCR, which suggests obvious growth performance and protein-sparing effects of increased lipid levels, which has previously been observed in grass carp (Köprücü 2012) and other fish species (Ai et al. 2004; Karalazos et al. 2007, 2011; Morais et al. 2001). Moreover, the lower LR observed under the HL treatments indicated an increased proportion of lipids used to supply energy, which is consistent with observations made in rainbow trout (Cho and Watanabe 1985) and European sea bass (Peres and Oliva-Teles 1999), further demonstrating that energy from lipids had protein-sparing effects. Nevertheless, some studies have found that grass carp have a relatively low capacity to endure a high energy intake and that no protein-sparing effects of dietary lipids can be achieved (Du et al. 2005; Gao et al. 2011). These discrepancies in results may be related to different dietary protein levels, fish sizes, or aquaculture conditions. Additionally, the lower FCR and higher PER in the silymarin supplementation compared to no silymarin supplementation under the HL diets further suggested that silymarin not only promotes feed efficiency, but also influences protein retention in fish fed with the high-lipid diets, which might be caused by increased catabolism of fatty acids for energy production through an enhanced β-oxidation capacity due to up-regulated hepatic expression of CPT1 mRNA (Fig. 2E), indicating the necessity of silymarin supplementation in high-lipid diets.
In general, increases in dietary lipid levels are associated with increases in lipid deposits in the viscera (Martino et al. 2002; Halver and Hardy 2002; Du et al. 2006). The elevated VSI observed under increased dietary lipid levels in this study has also been observed in grass carp (Gao et al. 2010), hybrid striped bass (Gaylord and Gatlin 2000), and cobia (Wang et al. 2005). However, diets supplemented with silymarin resulted in a decreased VSI. Previous studies showed that silibinin, an active component of silymarin, effectively reduced visceral fat and ameliorated visceral obesity in rats (Yao et al. 2013), indicating that silymarin may inhibit lipid deposition. Furthermore, GI indirectly indicates the size of the endogenous bile salt pool (Yamamoto et al. 2010). Bile acids play a critical role in the absorption of dietary fats in the intestine. In this study, the significant increases in GI observed under increasing dietary lipid levels might be connected with the amount of lipids in diets (Reddy et al. 1977), indicating that grass carp required more bile salts to address increased dietary lipid levels and to promote their delivery to the liver (Russell 2003). Cholesterol 7α-hydroxylase (CYP7A1) encodes the key rate-limiting enzyme in bile salt synthesis and plays an important role in the regulation of bile acid biosynthesis and cholesterol homeostasis (Chiang et al. 2001). Hepatic CYP7A1 mRNA was not influenced by dietary lipid levels (Table 7), in contrast to the increase in GI, which may be caused by feedback regulation of the elevated bile acid pool size. However, silymarin supplementation markedly affected GI and CYP7A1 mRNA in this study (P = 0.0450 and P = 0.0180, respectively). The SM100 groups showed significantly up-regulated hepatic CYP7A1, but displayed a slightly higher GI (despite the absence of a significant difference) compared with the SM0 groups, suggesting that silymarin has a positive effect on promoting bile acid synthesis, which is consistent with previous studies (Crocenzi et al. 2000).
Interestingly, increasing dietary lipid levels significantly decreased HSI in the present study. A similar phenomenon has been reported in silver perch (Yang et al. 2012), hybrid striped bass (Gaylord and Gatlin 2000), and European sea bass (Peres and Oliva-Teles 1999). In our study, the hepatic lipid content decreased as dietary lipid levels increased, which induced the same change in HSI. This phenomenon might be associated with the dietary carbohydrate levels (Brauge et al. 1995; Peres and Oliva-Teles 1999). Furthermore, to better understand the regulatory mechanism of dietary lipid levels associated with lipid homeostasis in fish, the expression patterns of genes involved in lipid biosynthesis (FAS and ACC) and catabolism (ATGL, HSL, and CPT1) were investigated in this study. FAS is the enzyme that catalyzes de novo synthesis of fatty acids (Jensen-Urstad and Semenkovich 2012), and ACC is required for the carboxylation of acetyl-CoA to produce malonyl-CoA (Davis et al. 2000). ATGL is the rate-limiting enzyme that catalyzes the first step of TG hydrolysis to diacylglycerol and fatty acids (Kratky et al. 2014). HSL is also the rate-limiting enzyme in the hydrolysis of stored triacylglycerol (Lampidonis et al. 2011). CPT1 serves as the main regulatory enzyme in mitochondrial fatty acid β-oxidation, catalyzing the conversion of acyl-CoAs into acylcarnitines for entry into the mitochondrial matrix (Kerner and Hoppel 2000). In the present study, high dietary lipid levels reduced the expression of lipogenesis-related gene, including FAS and ACC, but did not affect the expression of genes encoding proteins related to lipid catabolism (ATGL, HSL, and CPT1) (Table 7), indicating that the reduced capacity for de novo synthesis of fatty acids might result in lower hepatic lipid deposition in fish fed with dietary high lipid levels. Similarly, inhibition of lipogenesis and a decreased hepatic lipid content in response to increased dietary lipid levels have been observed in grass carp (Leng et al. 2012), gift tilapia (Wang et al. 2010), black sea bream (Ma et al. 2009), European sea bass (Dias et al. 1998), and rainbow trout (Brauge et al. 1995). However, a decreasing tendency of HSI was observed in association with the silymarin supplementation in the diets, and the SM200 group exhibited a dramatically lower HSI than the SM0 group under the HL treatments, which is in line with the results from previous studies in common carp (Jia et al. 2013). These findings indicate that silymarin treatment may normalize liver size. Similarly, silymarin supplementation significantly decreased the liver lipid content in the current study, which further confirms that silymarin has lipid-lowering effects (Sobolová et al. 2006; Metwally et al. 2009).
The mechanism of silymarin supplementation in the reduction in hepatic lipid deposition was also explored. HSL contributed to lipolysis (Reid et al. 2008), and CPT1 is considered to be the key regulatory enzyme in mitochondrial β-oxidation (Bruce et al. 2009). The presence of high-lipid diets without silymarin suppressed the gene expression of CPT1 mRNA (compared with LL-SM0 group), which was consistent with the results in blunt snout bream (Lu et al. 2014), probably because of the dietary fatty acid compositions that contained higher soybean oil content (rich in linoleic acid) (Coccia et al. 2014; Peng et al. 2014), or the changes in the mitochondrial membrane composition (Kolodziej and Zammit 1990; Morash et al. 2009) induced by a high-fat diet (Lu et al. 2014), those were found to be related to the CPT1 gene expression. However, up-regulation of hepatic HSL gene in HL-SM100 group (compared with LL-SM100 group) and hepatic CPT1 gene in HL-SM200 group (compared with LL-SM200 group) reflected silymarin supplemented with high-lipid diets had positive benefits for promoting lipid catabolism. Interestingly, we found that the up-regulation of the HSL and CPT1 mRNA levels only occured in high-fat diet with silymarin supplementation (HL-SM100 > HL-SM0 for HSL; both HL-SM100 and HL-SM200 > HL-SM0 for CPT1). To our knowledge, no information about the effects of silymarin on lipid metabolism was observed in fish, expect that silibinin, a major bioactive component of silymarin, has been reported to inhibit lipid accumulation in zebrafish by downregulating the mRNA expression of adipogenic factors (FAS) (Suh et al. 2015). However, in rat, previous studies have been shown that silymarin enhanced lipid catabolism and energy expenditure via β-oxidation (Yao et al. 2013; Chou et al. 2012) as well as reduced lipid accumulation against high-fat-diet-induced nonalcoholic fatty liver disease development (Zhang et al. 2013), and modulated liver lipid homeostasis (Salamone et al. 2012). It is well known that silymarin is a natural extract with hepatoprotective properties composed mainly of flavonolignans and has antioxidant effects (Vargas-Mendoza et al. 2014; GaΖák et al. 2007). Silymarin not only influenced hepatocellular plasma membrane stability, but quantities and qualities of membrane lipids such as cholesterol and phospholipids (Basiglio et al. 2009; Roberti et al. 1973), which may explain the responsibility to silymarin that was found only in the case of high-fat feeding. Moreover, in contrast to previous study (Suh et al. 2015), our results found that the expression of lipogenic gene (FAS and ACC) was not affected by silymarin supplementation, demonstrating silymarin supplementation might have no effect on the capacity of lipogenesis. Thus, these data suggest that silymarin has positive effects on improving lipolysis and fatty acid oxidation of the liver in fish fed with a high-fat diet.
Blood cholesterol derives from dietary cholesterol (exogenous cholesterol) and de novo cholesterol synthesis in the liver (endogenous cholesterol), the concentration of which depends on the nutritional status of fish (Regost et al. 2001). HDL carries cholesterol from peripheral tissues to the liver, whereas LDL carries cholesterol from the liver to peripheral tissues. In the present study, the serum T-cho and LDLc contents were significantly reduced as dietary lipid levels increased, while HDLc content showed a decreasing trend (but without a significant influence) associated with dietary lipid levels. The lower T-cho contents observed under the high-lipid diets are believed to largely be due to decreasing LDLc contents (Jain et al. 1993). Additionally, the lower serum T-cho and LDLc contents in the SM100 groups compared to the SM0 groups (P = 0.0120, 0.0561, respectively) suggest that silymarin might inhibit the absorption of dietary cholesterol owing to positively modifying plasma lipoprotein profile (decrease LDLc) (Škottová et al. 2003; Sobolová et al. 2006), which has been observed in rainbow trout (Banaee et al. 2011). In addition, silymarin supplementation up-regulated the gene expression of 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR) mRNA in the hepatopancreas, which is the rate-limiting enzyme of cholesterol synthesis (Friesen and Rodwell 2004), suggesting the capacity of endogenous cholesterol synthesis was improved, which was also confirmed by an elevated trend of hepatic T-cho contents (P = 0.067). Meanwhile, the SM100 groups exhibited higher hepatic CYP7A1 mRNA levels than the SM0 groups (P = 0.0188), supported by the GI, further proved that silymarin might enhance bile acid synthesis. The liver is the center of cholesterol homoeostasis (Sobolová et al. 2006). Cholesterol is a precursor of bile acid synthesis. Therefore, we deduced that, to compensate the cholesterol requirement, the endogenous cholesterol synthesis was up-regulated. However, studies on cholesterol and bile acids are scarce in fish, and further investigation will be required to understand the mechanisms involved.
TG, a water-insoluble energy-rich lipid, acts as a short-term indicator of nutritional status. In the present experiment, serum TG values increased in fish fed high-lipid diets, as previously shown in grass carp (Du et al. 2008) and grouper (Cheng et al. 2006). Plasma NEFA is mobilized from extrahepatic or hepatic lipid stores for oxidation and for certain anabolic processes in peripheral tissues in teleosts (Henderson and Tocher 1987). The major contribution to the plasma NEFA pool comes from adipose tissue, followed by dietary lipids and de novo lipogenesis (Donnelly et al. 2005). The current results regarding the NEFA contents in serum suggest that high lipid levels in diets might contribute to increased serum NEFA contents. The serum Glu values were dramatically increased by high dietary lipid levels in our study, which has also been found in grouper (Cheng et al. 2006). Nevertheless, the TG, NEFA, and Glu contents in serum were not affected by silymarin supplementation in this study, in contrast to some previous reports (Banaee et al. 2011; Tuorkey et al. 2015; Prakash et al. 2014), which might be due to differences in animal models, experimental conditions, research methods, or the composition of the diet (such as different doses of silymarin).
Bilirubin, the breakdown product of the heme moiety of hemoglobin and other hemoproteins, generally acts as an important indicator of liver dysfunction (Wang et al. 2006). In our experiment, the serum TBil content was reduced by silymarin supplementation, suggesting beneficial effects of silymarin on hepatic function (Ludovico et al. 2011; Crocenzi and Roma 2006).
Antioxidant defenses in fish are usually affected by nutritional factors. Fish have developed antioxidant defenses and are susceptible to attack by ROS (Lewis-McCrea and Lall 2007). SOD serves as an antioxidative enzyme, scavenging free radicals and thereby protecting tissue against peroxidation, while MDA is a final product of lipid peroxidation and may be generated during hydrolysis through the oxidation of polyunsaturated fatty acids and the degradation of preexisting oxidation products (Draper and Hadley 1990). Lipid peroxidation is thought to cause destruction and damage to cell membranes, with potential pathological effects on cells and tissues (Halver and Hardy 2002). Our results showed that SOD activities were not influenced by the dietary lipid levels or silymarin supplementation, while MDA contents were affected by both dietary lipid levels and silymarin supplementation. The increase in dietary lipid levels was highly correlated with the increase in MDA (Du et al. 2006), in accord with results previously obtained in grass carp (Du et al. 2008; Ji et al. 2011; Jin et al. 2013), blunt snout bream (Li et al. 2012), and turbot (Stéphan et al. 1995). However, silymarin supplementation significantly decreased the elevation of MDA levels induced by high-lipid diets in the current study, and similar results have been reported in common carp (Jia et al. 2013). In accordance with previous studies (Škottová et al. 2004; Shaker et al. 2010), silymarin supplementation was shown to generally exert protective effects against lipid peroxidation.
In conclusion, the results of this study suggest that both dietary silymarin supplementation and high lipid levels promote growth performance and feed efficiency, and enhance protein utilization. Moreover, dietary silymarin may inhibit lipid deposition due to the up-regulation of lipolysis and β-oxidation-related gene expression and positive effects on bile acid synthesis. In addition, silymarin improves health status via hepatoprotective effects and reduces the increase in lipid peroxidation induced by high lipid intake. These results provide new insights into the application of plant extracts as feed additives in high-lipid diets for freshwater fish.
Abbreviations
- ACC:
-
Acetyl coenzyme A carboxylase
- ATGL:
-
Adipose triglyceride lipase
- CF:
-
Condition factor
- CL:
-
Crude lipid
- CP:
-
Crude protein
- CPT-1:
-
Carnitine palmitoyl transferase 1
- CYP7A1:
-
Cholesterol 7α-hydroxylase
- DFI:
-
Day feed intake
- FAS:
-
Fatty acid synthase
- FBW:
-
Final average body weight
- FCR:
-
Feed conversion ratio
- GI:
-
Gallbladdersomatic index
- Glu:
-
Glucose
- HDL-c:
-
High-density lipoprotein cholesterol
- HL:
-
High lipid
- HMGCR:
-
3-Hydroxy-3-methylglutaryl-CoA reductase
- HPLC:
-
High-performance liquid chromatography
- HSI:
-
Hepatosomatic index
- HSL:
-
Hormone-sensitive lipase
- IBW:
-
Initial average body weight
- LDL-c:
-
Low-density lipoprotein cholesterol
- LL:
-
Low lipid
- LR:
-
Lipid retention
- MDA:
-
Malondialdehyde
- NEFA:
-
Nonesterified fatty acid
- PER:
-
Protein efficiency ratio
- PPV:
-
Protein productive value
- SGR:
-
Specific growth rate
- SM:
-
Silymarin
- SOD:
-
Superoxide dismutase
- SR:
-
Survival rate
- T-cho:
-
Total cholesterol
- TBil:
-
Total bilirubin
- TG:
-
Triacylglycerol
- VSI:
-
Viscerosomatic index
- WG:
-
Weight gain
References
Ahmadi K, Banaee M, Vosoghei AR, Mirvaghefei AR, Ataeimehr B (2012) Evaluation of the immunomodulatory effects of silymarin extract (Silybum marianum) on some immune parameters of rainbow trout, Oncorhynchus mykiss (Actinopterygii: Salmoniformes: Salmonidae). Acta Ichthyol Piscat 42:113–120
Ai Q, Mai K, Li H, Zhang C, Zhang L, Duan Q, Liufu Z (2004) Effects of dietary protein to energy ratios on growth and body composition of juvenile Japanese seabass, Lateolabrax japonicus. Aquaculture 230:507–516
AOAC (1998) Official methods of analysis. Association of Official analytical chemists, Washington, DC
Banaee M, Sureda A, Mirvaghefi AR, Rafei GR (2011) Effects of long-term silymarin oral supplementation on the blood biochemical profile of rainbow trout (Oncorhynchus mykiss). Fish Physiol Biochem 37:885–896
Basiglio CL, Pozzi EJS, Mottino AD, Roma MG (2009) Differential effects of silymarin and its active component silibinin on plasma membrane stability and hepatocellular lysis. Chem Biol Interact 179:297–303
Brauge C, Corraze G, Médale F (1995) Effects of dietary levels of carbohydrate and lipid on glucose oxidation and lipogenesis from glucose in rainbow trout, Oncorhynchus mykiss, reared in freshwater or in seawater. Comp Biochem Phys A 111:117–124
Bruce CR, Hoy AJ, Turner N, Watt MJ, Allen TL, Carpenter K et al (2009) Overexpression of carnitine palmitoyltransferase-1 in skeletal muscle is sufficient to enhance fatty acid oxidation and improve high-fat diet–induced insulin resistance. Diabetes 58:550–558
Cao L, Ding W, Du J, Jia R, Liu Y, Zhao C, Shen Y, Yin G (2015) Effects of curcumin on antioxidative activities and cytokine production in Jian carp (Cyprinuscarpio var. Jian) with CCl4 -induced liver damage. Fish Shellfish Immun 43:150–157
Chen I, Chen Y, Chou C, Chuang R, Sheen L, Chiu C (2012) Hepatoprotection of silymarin against thioacetamide-induced chronic liver fibrosis. J Sci Food Agr 92:1441–1447
Cheng A, Chen C, Liou C, Chang C (2006) Effects of dietary protein and lipids on blood parameters and superoxide anion production in the Grouper, Epinephelus coioides (Serranidae: Epinephelinae). Zool Stud 45:492–502
Chiang J, Kimmel R, Stroup D (2001) Regulation of cholesterol 7α-hydroxylase gene (CYP7A1) transcription by the liver orphan receptor (LXRα). Gene 262:257–265
Cho C, Watanabe T (1985) Dietary energy and lipid requirements of rainbow trout (Salmo gairdneri) at different water temperatures, energy metabolism of farm animals, proceedings of the 10th symposium EAAP, Airlie, Virginia, pp 206–209
Chou C, Chen Y, Hsu M, Tsai W, Chang C, Chiu C (2012) Effect of silymarin on lipid and alcohol metabolism in mice following long-term alcohol consumption. J Food Biochem 36:369–377
Citarasu T (2010) Herbal biomedicines: a new opportunity for aquaculture industry. Aquacult Int 18:403–414
Coccia E, Varricchio E, Vito P, Turchini GM, Francis D, Paolucci M (2014) Fatty acid-specific alterations in leptin, PPARα, and CPT-1 gene expression in the rainbow trout. Lipids 49:1033–1046
Colturato CP, Constantin RP, Maeda AS, Constantin RP, Yamamoto NS, Bracht A, Ishii-Iwamoto EL, Constantin J (2012) Metabolic effects of silibinin in the rat liver. Chem Biol Interact 195:119–132
Crocenzi FA, Roma MG (2006) Silymarin as a new hepatoprotective agent in experimental cholestasis: new possibilities for an ancient medication. Curr Med Chem 13:1055–1074
Crocenzi FA, Pellegrino JM, Pozzi EJS, Mottino AD, Garay EAR, Roma MG (2000) Effect of silymarin on biliary bile salt secretion in the rat. Biochem Pharmacol 59:1015–1022
Davis MS, Solbiati J, Cronan JE (2000) Overproduction of acetyl-CoA carboxylase activity increases the rate of fatty acid biosynthesis in Escherichia coli. J Biol Chem 275:28593–28598
Dias J, Alvarez M, Diez A, Arzel J, Corraze G, Bautista J, Kaushik S (1998) Regulation of hepatic lipogenesis by dietary protein/energy in juvenile European seabass (Dicentrarchus labrax). Aquaculture 161:169–186
Donnelly KL, Smith CI, Schwarzenberg SJ, Jessurun J, Boldt MD, Parks EJ (2005) Sources of fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty liver disease. J Clin Invest 115:1343–1351
Draper HH, Hadley M (1990) Malondialdehyde determination as index of lipid Peroxidation. Method Enzymol 186:421–431
Du Z, Liu Y, Tian L, Wang J, Wang Y, Liang G (2005) Effect of dietary lipid level on growth, feed utilization and body composition by juvenile grass carp (Ctenopharyngodon idella). Aquacult Nutr 11:139–146
Du Z, Clouet P, Zheng W, Degrace P, Tian L, Liu Y (2006) Biochemical hepatic alterations and body lipid composition in the herbivorous grass carp (Ctenopharyngodon idella) fed high-fat diets. Brit J Nutr 95:905–915
Du Z, Clouet P, Degrace P, Zheng W, Frøyland L, Tian L, Liu Y (2008) Hypolipidaemic effects of fenofibrate and fasting in the herbivorous grass carp (Ctenopharyngodon idella) fed a high-fat diet. Brit J Nutr 100:1200–1212
El-Lakkany NM, Hammam OA, El-Maadawy WH, Badawy AA, Ain-Shoka AA, Ebeid FA (2012) Anti-inflammatory/anti-fibrotic effects of the hepatoprotective silymarin and the schistosomicide praziquantel against Schistosoma mansoni-induced liver fibrosis. Parasite Vector 5:21–32
El-Sheikh NM, Abd El-Fattah HM (2011) Counteracting methionine choline-deficient diet-induced fatty liver by administration of turmeric and silymarin. J Appl Sci Res 7:1812–1820
Friesen JA, Rodwell VW (2004) The 3-hydroxy-3-methylglutaryl coenzyme-A (HMG-CoA) reductases. Genome Biol 5:60
Gao W, Liu Y, Tian L, Mai K, Liang G, Yang H, Huai M, Luo W (2010) Effect of dietary carbohydrate-to-lipid ratios on growth performance, body composition, nutrient utilization and hepatic enzymes activities of herbivorous grass carp (Ctenopharyngodon idella). Aquacult Nutr 16:327–333
Gao W, Liu Y, Tian L, Mai K, Liang G, Yang H, Huai M, Luo W (2011) Protein-sparing capability of dietary lipid in herbivorous and omnivorous freshwater finfish: a comparative case study on grass carp (Ctenopharyngodon idella) and tilapia (Oreochromis niloticus × O. aureus). Aquacult Nutr 17:2–12
Gaylord T, Gatlin D (2000) Dietary lipid level but not l-carnitine affects growth performance of hybrid striped bass (Morone chrysops♀ × M. saxatilis♂). Aquaculture 190:237–246
GaΖák R, Walterová D, Kren V (2007) Silybin and silymarin-new and emerging applications in medicine. Curr Med Chem 14:315–338
Halver JE, Hardy RW (2002) Fish nutrition. Academic press, San Diego
Henderson RJ, Tocher DR (1987) The lipid composition and biochemistry of freshwater fish. Prog Lipid Res 26:281–347
Jain AK, Vargas R, Gotzkowsky S, McMahon FG (1993) Can garlic reduce levels of serum lipids? A controlled clinical study. Am J Med 94:632–635
Jensen-Urstad AP, Semenkovich CF (2012) Fatty acid synthase and liver triglyceride metabolism: housekeeper or messenger? BBA Mol Cell Biol L 1821:747–753
Ji H, Li J, Liu P (2011) Regulation of growth performance and lipid metabolism by dietary n-3 highly unsaturated fatty acids in juvenile grass carp, Ctenopharyngodon idellus. Comp Biochem Phys B 159:49–56
Jia R, Cao L, Du J, Xu P, Jeney G, Yin G (2013) The protective effect of silymarin on the carbon tetrachloride (CCl4)-induced liver injury in common carp (Cyprinus carpio). In Vitro Cell Dev An 49:155–161
Jin Y, Tian L, Zeng S, Xie S, Yang H, Liang G, Liu Y (2013) Dietary lipid requirement on non-specific immune responses in juvenile grass carp (Ctenopharyngodon idella). Fish Shellfish Immun 34:1202–1208
Ka SO, Kim KA, Kwon KB, Park JW, Park BH (2009) Silibinin attenuates adipogenesis in 3T3-L1 preadipocytes through a potential upregulation of the insig pathway. Int J Mol Med 23:633–637
Karalazos V, Bendiksen EÅ, Dick JR, Bell JG (2007) Effects of dietary protein and fatlevel and rapeseed oil on growth and tissue fatty acid composition and metabolism in Atlantic salmon (Salmo salar L.) reared at low water temperatures. Aquacult Nutr 13:256–265
Karalazos V, Bendiksen EÅ, Bell JG (2011) Interactive effects of dietary protein/lipid level and oil source on growth, feed utilisation and nutrient and fatty acid digestibility of Atlantic salmon. Aquaculture 311:193–200
Kerner J, Hoppel C (2000) Fatty acid import into mitochondria. BBA Mol Cell Biol L 1486:1–17
Kolodziej MP, Zammit VA (1990) Sensitivity of inhibition of rat liver mitochondrial outer-membrane carnitine palmitoyltransferase by malonyl-CoA to chemical-and temperature-induced changes in membrane fluidity. Biochem J 272:421–425
Köprücü K (2012) Effects of dietary protein and lipid levels on growth, feed utilization and body composition of juvenile grass carp (Ctenopharyngodon idella). J Fish Sci 6:243–251
Kratky D, Obrowsky S, Kolb D, Radovic B (2014) Pleiotropic regulation of mitochondrial function by adipose triglyceride lipase-mediated lipolysis. Biochimie 96:106–112
Lampidonis AD, Rogdakis E, Voutsinas GE, Stravopodis DJ (2011) The resurgence of hormone-sensitive lipase (HSL) in mammalian lipolysis. Gene 477:1–11
Leng X, Wu X, Tian J, Li X, Guan L, Weng D (2012) Molecular cloning of fatty acid synthase from grass carp (Ctenopharyngodon idella) and the regulation of its expression by dietary fat level. Aquacult Nutr 18:551–558
Lewis-McCrea LM, Lall SP (2007) Effects of moderately oxidized dietary lipid and the role of vitamin E on the development of skeletal abnormalities in juvenile Atlantic halibut (Hippoglossus hippoglossus). Aquaculture 262:142–155
Li X, Liu W, Lu K, Xu W, Wang Y (2012) Dietary carbohydrate/lipid ratios affect stress, oxidative status and non-specific immune responses of fingerling blunt snout bream, Megalobrama amblycephala. Fish Shellfish Immun 33:316–323
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using realtimequantitative PCR and the 2−ΔΔCT method. Methods 25:402–408
Lu K, Xu W, Li J, Li X, Huang G, Liu W (2013a) Alterations of liver histology and blood biochemistry in blunt snout bream Megalobrama amblycephala fed high-fat diets. Fish Sci 79:661–671
Lu K, Xu W, Li X, Liu W, Wang L, Zhang C (2013b) Hepatic triacylglycerol secretion, lipid transport and tissue lipid uptake in blunt snout bream (Megalobrama amblycephala) fed high-fat diet. Aquaculture 408:160–168
Lu K, Xu W, Wang L, Zhang D, Zhang C, Liu W (2014) Hepatic β-oxidation and regulation of carnitine palmitoyltransferase (CPT) I in blunt snout bream Megalobrama amblycephala fed a high fat diet. PLoS ONE 9:e93135
Ludovico A, Gabriella A, Raffaele C, Natasa M, Francesco C (2011) Milk thistle for treatment of nonalcoholic fatty liver disease. Hepat Mon 11:173–177
Ma J, Shao Q, Xu Z, Zhou F, Zhong G, Song W, Owari NB (2009) Effects of dietary n-3 HUFA on growth performance and lipid metabolism in juvenile black sea bream, Sparus macrocephalus. J Fish China 33:639–648
Martino RC, Cyrino JEP, Portz L, Trugo LC (2002) Effect of dietary lipid level on nutritional performance of the surubim, Pseudoplatystoma coruscans. Aquaculture 209:209–218
Metwally M, El-Gellal A, El-Sawaisi S (2009) Effects of silymarin on lipid metabolism in rats. World Appl Sci J 6:1634–1637
Morais S, Bell JG, Robertson DA, Roy WJ, Morris PC (2001) Protein/lipid ratios in extruded diets for Atlantic cod (Gadus morhua L.): effects on growth, feed utilisation, muscle composition and liver histology. Aquaculture 203:101–119
Morash AJ, Bureau DP, McClelland GB (2009) Effects of dietary fatty acid composition on the regulation of carnitine palmitoyltransferase (CPT) I in rainbow trout (Oncorhynchus mykiss). Comp Biochem Phys B 152:85–93
Nanton DA, Lall SP, Ross NW, McNiven MA (2003) Effect of dietary lipid level on fatty acid β-oxidation and lipid composition in various tissues of haddock, Melanogrammus aeglefinus L. Comp Biochem Phys B 135:95–108
Peng M, Xu W, Mai K, Zhou H, Zhang Y, Liufu Z, Zhang K, Ai Q (2014) Growth performance, lipid deposition and hepatic lipid metabolism related gene expression in juvenile turbot (Scophthalmus maximus, L.) fed diets with various fish oil substitution levels by soybean oil. Aquaculture 433:442–449
Peres H, Oliva-Teles A (1999) Effect of dietary lipid level on growth performance and feed utilization by European sea bass juveniles (Dicentrarchus labrax). Aquaculture 179:325–334
Pfaffl MW (2001) A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29:e45
Prakash P, Singh V, Jain M, Rana M, Khanna V, Barthwal MK, Dikshit M (2014) Silymarin ameliorates fructose induced insulin resistance syndrome by reducing de novo hepatic lipogenesis in the rat. Eur J Pharmacol 727:15–28
Reddy BS, Mangat S, Sheinfil A, Weisburger JH, Wynder EL (1977) Effect of type and amount of dietary fat and 1, 2-dimethylhydrazine on biliary bile acids, fecal bile acids, and neutral sterols in rats. Cancer Res 37:2132–2137
Regost C, Arzel J, Cardinal M, Robin J, Laroche M, Kaushik S (2001) Dietary lipid level, hepatic lipogenesis and flesh quality in turbot (Psetta maxima). Aquaculture 193:291–309
Reid BN, Ables GP, Otlivanchik OA, Schoiswohl G, Zechner R, Blaner WS, Goldberg IJ, Schwabe RF, Chua SC Jr, Huang LS (2008) Hepatic overexpression of hormone-sensitive lipase and adipose triglyceride lipase promotes fatty acid oxidation, stimulates direct release of free fatty acids, and ameliorates steatosis. J Biol Chem 283:13087–13099
Roberti R, Mozzi R, De Medio GE, Francescangeli E, Porcellati G (1973) The activity of silymarin on membrane-bound phospholipid metabolism in brain and liver tissues of the rat. Pharmacol Res Commun 5:249–257
Rolo AP, Teodoro JS, Palmeira CM (2012) Role of oxidative stress in the pathogenesis of nonalcoholic steatohepatitis. Free Radical Bio Med 52:59–69
Russell DW (2003) The enzymes, regulation, and genetics of bile acid synthesis. Ann Rev Biochem 72:137–174
Salamone F, Galvano F, Cappello F, Mangiameli A, Barbagallo I, Volti GL (2012) Silibinin modulates lipid homeostasis and inhibits nuclear factor kappa B activation in experimental nonalcoholic steatohepatitis. Transl Res 159:477–486
Sanchez R, Olivares P, Carmona E, Astuya A, Herrera H, Parodi J (2016) Fish Nutrition Additives in SHK-1 Cells: protective effects of silymarin. Adv Biosci Biotechnol 7:55–62
Shaker E, Mahmoud H, Mna S (2010) Silymarin, the antioxidant component and Silybum marianum extracts prevent liver damage. Food Chem Toxicol 48:803–806
Škottová N, Večeřa R, Urbánek K, Váňa P, Walterová D, Cvak L (2003) Effects of polyphenolic fraction of silymarin on lipoprotein profile in rats fed cholesterol-rich diets. Pharmacol Res 47:17–26
Škottová N, Kazdová L, Oliyarnyk O, Večeřa R, Sobolová L, Ulrichová J (2004) Phenolics-rich extracts from Silybum marianum and Prunella vulgaris reduce a high-sucrose diet induced oxidative stress in hereditary hypertriglyceridemic rats. Pharmacol Res 50:123–130
Sobolová L, Škottová N, Večeřa R, Urbánek K (2006) Effect of silymarin and its polyphenolic fraction on cholesterol absorption in rats. Pharmacol Res 53:104–112
Steffens W (1996) Protein sparing effect and nutritive significance of lipid supplementation in carp diets. Arch Anim Nutr 49:93–98
Stéphan G, Guillaume J, Lamour F (1995) Lipid peroxidation in turbot (Scophthalmus maximus) tissue: effect of dietary vitamin E and dietary n − 6 or n − 3 polyunsaturated fatty acids. Aquaculture 130:251–268
Suh HJ, Cho SY, Kim EY, Choi HS (2015) Blockade of lipid accumulation by silibinin in adipocytes and zebrafish. Chem Biol Interact 227:53–62
Surai PF (2015) Silymarin as a natural antioxidant: an overview of the current evidence and perspectives. Antioxidants 4:204–247
Tocher DR (2003) Metabolism and functions of lipids and fatty acids in teleost fish. Rev Fish Sci 11:107–184
Tuorkey MJ, El-Desouki NI, Kamel RA (2015) Cytoprotective effect of silymarin against diabetes-induced cardiomyocyte apoptosis in diabetic rats. Biomed Environ Sci 28:36–43
Vargas-Mendoza N, Madrigal-Santillán E, Morales-González A, Esquivel-Soto J, Esquivel-Chirino C, García-Luna Y, Gayosso-de-Lucio JA, Morales-González JA (2014) Hepatoprotective effect of silymarin. World J Hepatol 6:144–149
Vecera R, Zacharova A, Orolin J, Skottova N, Anzenbacher P (2011) The effect of silymarin on expression of selected ABC transporters in the rat. Vet Med Czech 56:59–62
Vergara JM, Robainà L, Izquierdo M, Higuera MDL (1996) Protein sparing effect of lipids in diets for fingerlings of gilthead sea bream. Fish Sci 62:624–628
Wang J, Liu Y, Tian L, Mai K, Du Z, Wang Y, Yang H (2005) Effect of dietary lipid level on growth performance, lipid deposition, hepatic lipogenesis in juvenile cobia (Rachycentron canadum). Aquaculture 249:439–447
Wang X, Chowdhury JR, Chowdhury NR (2006) Bilirubin metabolism: applied physiology. Curr Paediatr 16:70–74
Wang A, Han G, Wei X, Liu B, Lv F, Feng G, Qi Z, Wang T, Xu P, Yang Z (2010) Molecular cloning of fatty acid synthase from gift tilapia (Oreochromis niloticus): response of its expression to refeeding and different lipid levels in diet. J Fish China 34:1113–1120
Yamamoto T, Iwashita Y, Matsunari H, Sugita T, Furuita H, Akimoto A, Okamatsu K, Suzuki N (2010) Influence of fermentation conditions for soybean meal in a non-fish meal diet on the growth performance and physiological condition of rainbow trout Oncorhynchus mykiss. Aquaculture 309:173–180
Yang S, Liu F, Liou C (2012) Effects of dietary L-carnitine, plant proteins and lipid levels on growth performance, body composition, blood traits and muscular carnitine status in juvenile silver perch (Bidyanus bidyanus). Aquaculture 342:48–55
Yao J, Zhi M, Minhu C (2011) Effect of silybin on high-fat-induced fatty liver in rats. Braz J Med Biol Res 44:652–659
Yao J, Zhi M, Gao X, Hu P, Li C, Yang X (2013) Effect and the probable mechanisms of silibinin in regulating insulin resistance in the liver of rats with non-alcoholic fatty liver. Braz J Med Biol Res 46:270–277
Yi D, Gu L, Ding B, Li M, Hou Y, Wang L, Gong J (2012) Effects of dietary silymarin supplementation on growth performance and oxidative status in Carassius auratus gibelio. J Anim Vet Adv 11:3399–3404
Zhang Y, Hai J, Cao M, Zhang Y, Pei S, Wang J, Zhang Q (2013) Silibinin ameliorates steatosis and insulin resistance during non-alcoholic fatty liver disease development partly through targeting IRS-1/PI3K/Akt pathway. Int Immunopharmacol 17:714–720
Acknowledgments
Financial support was provided by the National Basic Research Program of China Project No. 2014CB138603, Plan for Outstanding Young of Beijing Academy of Science and Technology Project No. 201420 and the fund of Beijing sunpu biochem. tech. co., Ltd, China. We thank Hua cheng aquaculture farm (Jiangsu, China) for their juvenile fish and School of Basic Medicine and Biological Science, Soochow University, for rearing condition support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Xiao, P., Ji, H., Ye, Y. et al. Dietary silymarin supplementation promotes growth performance and improves lipid metabolism and health status in grass carp (Ctenopharyngodon idellus) fed diets with elevated lipid levels. Fish Physiol Biochem 43, 245–263 (2017). https://doi.org/10.1007/s10695-016-0283-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10695-016-0283-6