
Abstract
Fish are faced with a wide range of hydrostatic pressure (HP) in their natural habitats. Additionally, freshwater fish are occasionally exposed to rapid changes in HP due to heavy rainfall, flood and/or dam release. Accordingly, variations in HP are one of the most important environmental cues for fish. However, little information is available on how HP information is perceived and transmitted in the central nervous system of fish. The present study examined the effect of HP (water depth of 1.3 m) on the quantities of monoamines and their metabolites in the telencephalon, optic tectum, diencephalon, cerebellum (including partial mesencephalon) and vagal lobe (including medulla oblongata) of the goldfish, Carassius auratus, using high-performance liquid chromatography. HP affected monoamine and metabolite contents in restricted brain regions, including the telencephalon, cerebellum and vagal lobe. In particular, HP significantly increased the levels of dopamine (DA) in the telencephalon at 15 min and that of norepinephrine (NE) in the cerebellum at 30 min. In addition, HP also significantly increased locomotor activity at 15 and 30 min after HP treatment. It is possible that HP indirectly induces locomotion in goldfish via telencephalic DA and cerebellar NE neuronal activity.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Aquatic organisms are constantly exposed to hydrostatic pressure (HP). Notably, many fish are faced with a wide range of HPs in their natural habitats, such as during vertical migration (Godø and Michalsen 2000; Janssen and Brandt 1980; McCleave and Arnold 1999; Raffaelli et al. 1990). Additionally, when water levels increase rapidly in freshwater due to heavy rainfall, flood and/or dam release, freshwater fish migrate to safer places in response to increasing HP and flow velocity (Koizumi et al. 2013; Nunn et al. 2010). Marine fish are always exposed to semidiurnal-cyclic HP changes caused by tidal amplitude (Northcott et al. 1991b), and they synchronize their migration, feeding and spawning with tidal cycles (Gibson 1984, 1992; Takemura et al. 2010b). Therefore, it seems likely that HP is an important environmental cue for fish (Northcott et al. 1991a, b).
The sensor for HP has been found only in the crab, Carcinus maenas, and the dogfish, Scyliorhinus canicula (cartilaginous fish) (Fraser et al. 2003; Fraser and Macdonald 1994; Fraser and Shelmerdine 2002). In the crab, sensory hair receptors in the balancing organ (statocyst) respond to HP with a piston mechanism: differential compression of the outer cuticle and the internal fluid of thread hairs in the balancing system lead to nanometer-level displacements that are sensed by mechanoreceptors (Fraser and Macdonald 1994). In the dogfish, vestibular hair cells located in the labyrinth sense HP due to changes in resting and angular acceleration-derived activity after a change in HP (Fraser and Shelmerdine 2002).
After perceiving HP, this information is transmitted to the central nervous system (CNS). In fact, when isolated labyrinths are exposed to cyclic HP, a positive relationship is observed between nerve spike frequency and HP in the dogfish (Fraser and Shelmerdine 2002). Some studies have reported on the effects of HP on monoamines (known as neurotransmitters and neuromodulators) and their metabolites in the CNS of marine fish (Damasceno-Oliveira et al. 2006, 2007; Sebert et al. 1985, 1986; Takemura et al. 2010a, 2012). Exposing the European eel, Anguilla anguilla, to constant HP (101 ATA, correspondence to a water depth of 1000 m) for 1 h results in a rise in brain epinephrine (E) and dopamine (DA) contents, whereas HP does not affect brain serotonin (5-HT) or its metabolite 5-hydroxyindoleacetic acid (5-HIAA) contents (Sebert et al. 1985, 1986). In the threespot wrasse, Halicoeres trimaculatus, constant HP (water depth of 3 m) for 3–6 h decreases the DA and 5-HT turnover ratio (Takemura et al. 2010a, 2012). Furthermore, in the European flounder, Platichthys flesus, tidal-related-cyclic changes in HP (800 kpa, corresponding to a water depth of ~70 m) decrease norepinephrine (NE), DA, 3-methoxytyramine (DA metabolite), 5-HT and 5-HIAA contents in the brain, whereas constant HP does not affect those compounds (Damasceno-Oliveira et al. 2006, 2007). Therefore, it seems likely that brain monoamines act as neurotransmitters and/or neuromodulators in the CNS in response to HP in fish.
Monoamines are involved in different behaviors and physiological processes in some vertebrates. For example, dopaminergic perikarya are located mainly in the hypothalamus and mesencephalon in mammals. Hypothalamic DA regulates prolactin release from the pituitary, whereas mesencephalic DA modulates motor control and emotional behavior (Gnegy 2012). In fish, a wealth of dopaminergic perikarya is concentrated in the hypothalamus (Corio et al. 1991; Goping et al. 1995; Kah and Chambolle 1983; Yoshida et al. 1983). One of the functions of hypothalamic DA is to inhibit a gonadotropin release from the pituitary (Vidal et al. 2004). In addition, dopaminergic perikarya are present in the telencephalon (Kapsimali et al. 2000; Vetillard et al. 2002), and telencephalic DA modulates locomotion (Goping et al. 1995; Pollard et al. 1992, 1996; Thompson et al. 2008). Therefore, it is possible that HP indirectly affects behaviors and/or physiological processes by affecting monoaminergic neuron activities in fish.
The goldfish, Carassius auratus, has been used as a neuro-experimental model for decades, and the distribution of brain monoaminergic neurons is well known when compared with that in other fish species (Bonn 1987; Geffard et al. 1984; Gotow et al. 1990; Kah and Chambolle 1983; Yoshida et al. 1983). In addition, DA and 5-HT functions in some specific brain regions have been already clarified in the goldfish (De Pedro et al. 1998; Dietrich et al. 2002; Goping et al. 1995; Kah and Chambolle 1983; Olcese et al. 1979; Pollard et al. 1992, 1996; Somoza and Peter 1991; Wollmuth et al. 1989). Accordingly, the present study clarified the effect of HP on brain monoamines and HP-affected monoamine functions in the goldfish. First, the effect of HP (water depth of 1.3 m) on the quantities of monoamines and their metabolites was examined in the telencephalon, optic tectum, diencephalon, cerebellum (including partial mesencephalon) and vagal lobe (including medulla oblongata) using high-performance liquid chromatography (HPLC). Second, the effect of HP on locomotor activity was examined because HP increased telencephalic contents of DA and DA metabolites and cerebellar content of NE, which were expected to be involved in locomotion.
Materials and methods
Experimental fish
Goldfish (body weight, 4.4 ± 0.1 g) were purchased from a local dealer and maintained in an aquarium (350-mm height × 300-mm width × 600-mm depth) at 25 °C under a 12-h light:12-h dark photoperiod for 2 weeks. Fish were fed commercial goldfish pellets equivalent to 1 % of body weight per day.
Effect of HP on monoamine levels in goldfish brain
Sixty-three fish were transferred to nine cages (200 × 350 × 350 mm) set at the water surface of a 3-metric-ton-capacity concrete tank with a depth of 1.3 m. Seven individuals were housed in each cage. After a 24-h acclimation under these conditions, the cages were either kept at the surface (control group, n = 7; depth of 0 m, 101 kpa) or moved to the tank bottom (HP group, n = 7; depth of 1.3 m, 115 kpa). At 0, 15, 30, 60 and 180 min, fish were removed from cages and anesthetized in 0.03 % tricaine methanesulfonate (Sigma-Aldrich, St. Louis, MO, USA) buffered with an equal amount of sodium bicarbonate. Then fish were killed by decapitation, and the brain was removed. The brains were trimmed into the telencephalon, optic tectum, diencephalon, cerebellum (including partial mesencephalon) and vagal lobe (including medulla oblongata), as shown in Fig. 1, and immediately frozen in liquid nitrogen and stored at −80 °C until monoamine measurements.

Schematic illustration of goldfish brain trimming. 1 Telencephalon; 2 optic tectum; 3 diencephalon; 4 cerebellum (including partial mesencephalon); 5 vagal lobe (including medulla oblongata)
Monoamine quantification in brain tissues
Monoamines and their metabolites were measured in the telencephalon, optic tectum, diencephalon, cerebellum and vagal lobe. Each brain tissue was homogenized in 0.2 M ice-cold perchloric acid, and the homogenates were cooled on ice for 30 min for deproteinization. The homogenates were centrifuged at 20,000×g for 15 min at 0 °C. Then, pH of the supernatant was adjusted to 3.0 by adding 1 M sodium acetate, and the precipitate was used to quantify protein. The samples were filtered through a 0.45-µm filter (Millipore, Billerica, MA, USA). NE, DA, 3,4-dihydroxyphenylacetic acid (DOPAC), 5-HT and 5-HIAA concentrations were analyzed in filtrates by HPLC (Eicom, Kyoto, Japan) with a 150 × 3.0-mm ODS column (EICOMPAK SC-5ODS, Eicom) and an electrochemical detector (ECD-300, Eicom) at an applied potential of +750 mV versus an Ag/AgCl reference analytical electrode. The change in electric current (nA) was recorded using a computer interface (Power Chrom ver 2.3.2.J; AD Instruments, Tokyo, Japan). The mobile phase was pH 3.5 and consisted of 0.1 M sodium acetate, 0.1 M citric acid, sodium 1-octan sulphanate and disodium ethylenediaminetetraacetic acid (5 mg/ml). Flow rate was 0.5 ml/min. The retention time and height of the peaks in the tissue homogenates were measured and compared to samples of external calibrating standard solution (0.2–50 pg/µl each of NE, DA, DOPAC, 5-HT and 5-HIAA). Protein was quantified using the BCA method according to the manufacturer’s protocol (Sigma-Aldrich). The concentrations of these monoamines in the sample were calculated and expressed as pg/mg protein.
Effect of HP on locomotor activity of goldfish
Individual fish was transferred to the cage sets (100 × 90 × 177 mm) at the water surface of a 10-metric-ton-capacity concrete tank, and the cages were set either at the surface (control group, n = 9; depth of 0 m, 101.3 kpa) or at the tank bottom (HP group, n = 9; depth of 1.3 m, 114 kpa). Fifteen and 30 min later, fish were individually transferred to an observation tank (150 × 130 × 170 mm) filled with 1.6-L tap water (water depth of 80 mm). A video camera (HDR-PJ590, Sony, Tokyo, Japan) directly above the tank was used to record locomotion. Locomotor activity was measured by counting the frequency that the goldfish’s head crossed grid lines (one grid size, 30 × 30 mm) during a 10-min video observation. Locomotion consisted of swimming movement.
Statistical analysis
Values are expressed as means ± standard errors. Comparisons of the monoamine and metabolite contents among the five brain regions were performed by one-way analysis of variance followed by Tukey’s post hoc test for multiple-group comparisons. Comparisons between monoamines, their metabolites and locomotion between the control and the HP groups were performed by Student’s t test. Statistical significance was set at P < 0.05. Statistical analyses were performed using SPSS for Windows (SPSS Inc., Chicago, IL, USA).
Results
Levels of monoamines and their metabolites in the goldfish brain
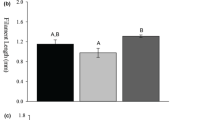
Levels of monoamines and their metabolites in five brain regions of the initial control fish (at 0 min) were compared (Fig. 2). The highest NE level was found in the telencephalon (Fig. 2a), whereas the highest DA, 5-HT and 5-HIAA levels were found in the diencephalon (Fig. 2b, d, e). No significant differences were observed in DOPAC levels among the five brain regions (Fig. 2c).

Distribution of monoamines and their metabolites in goldfish brain. Distribution of a norepinephrine (NE), b dopamine (DA), c 3,4-dihydroxyphenylacetic acid (DOPAC), d serotonin (5-HT) and e 5-hydroxyindoleacetic acid (5-HIAA) in the goldfish brain. T telencephalon, O optic tectum, D diencephalon, C cerebellum, V vagal lobe. Probability values (P) are indicated for significant differences among brain regions (one-way analysis of variance). Values with different characters are significantly different among brain regions (P < 0.05)
Effect of HP on levels of monoamines and their metabolites in the telencephalon, cerebellum and vagal lobe
Of the brain areas (telencephalon, optic tectum, diencephalon, cerebellum, and vagal lobe) tested, the effect of HP on the levels of monoamines and their metabolites was found only among the telencephalon (Fig. 3), cerebellum (Fig. 4) and vagal lobe (Fig. 5). HP increased telencephalic DA and DOPAC levels at 15 and 30 min, respectively (Fig. 3a, b). In addition, HP also increased cerebellar NE and 5-HIAA levels at 30 min (Fig. 4a, c) while HP did not affect cerebellar 5-HT level (Fig. 4b). In contrast, HP inhibited 5-HT and 5-HIAA levels in the vagal lobe at 30 and 60 min, respectively (Fig. 5a, b).

Effect of hydrostatic pressure (HP) on a dopamine (DA) and b 3,4-dihydroxyphenylacetic acid (DOPAC) contents in the telencephalon. Asterisk and double asterisk indicate significant differences at P < 0.05 and P < 0.001 in DA and DOPAC contents between the control and HP groups, respectively

Effect of hydrostatic pressure (HP) on a norepinephrine (NE), b serotonin (5-HT) and c 5-hydroxyindoleacetic acid (5-HIAA) contents in the cerebellum. Asterisk indicates significant differences at P < 0.05 in NE and 5-HIAA contents between the control and HP groups

Effect of hydrostatic pressure (HP) on a serotonin (5-HT) and b 5-hydroxyindoleacetic acid (5-HIAA) contents in the vagal lobe. Asterisk indicates significant differences at P < 0.05 in 5-HT and 5-HIAA contents between the control and HP groups
Effect of HP on locomotor activity
The effect of HP on locomotor activity was examined at 15 and 30 min after exposure to HP using a video camera for 10 min. HP induced locomotion both at 15 and 30 min (Fig. 6): Locomotor activity was three- and fourfold higher in the HP group than in the control group at 15 and 30 min, respectively.

Effect of hydrostatic pressure (HP) on locomotor activity. Asterisk indicates significant differences at P < 0.05 in locomotor activity between the control and HP groups
Discussion
We clarified brain levels of monoamines and their metabolites in goldfish in the present study (Fig. 2a–e). Excellent agreement was observed in the rank order of DA, 5-HT and 5-HIAA levels in the brain regions between the goldfish and crucian carp, Carassius carassius (Nilsson 1989). The highest DA, 5-HT and 5-HIAA levels were found in the diencephalon (Fig. 2b, d, e). In addition, immunohistochemical studies have revealed a wealth of DA-immunoreactive (DA-IR) perikarya and 5-HT-IR perikarya in the hypothalamus of the goldfish (Goping et al. 1995; Kah and Chambolle 1983; Yoshida et al. 1983), zebrafish, Danio rerio (Lillesaar, 2011), African sharptooth catfish, Clarias gariepinus (Corio et al. 1991) and rainbow trout, Oncorhynchus mykiss (Vetillard et al. 2002), demonstrating that dopaminergic and serotonergic perikarya are highly concentrated in the hypothalamus. These results suggest that the distribution of DA and 5-HT neurons is conserved in fish species.
The rank order of NE in the brain regions of goldfish was in agreement with that reported in rainbow trout (Linard et al. 1996) and European eel (Sébert et al. 2008). In those studies, the highest NE level was found in the telencephalon (Fig. 2a). In fact, in the goldfish telencephalon, dense innervation of tyrosine hydroxylase-IR (TH-IR) and dopamine β-hydroxylase-IR fibers has been noted within the area dorsalis, pars lateralis and pars dorsalis of the telencephalon (Hornby and Piekut 1990). Similarly, a formaldehyde-induced fluorescence study in the telencephalon demonstrated that monoaminergic fibers in the European eel contain predominantly NE fluorophores (Fremberg et al. 1977). In the crucian carp, the highest NE level was found in the hypothalamus (Nilsson 1989). Therefore, the distribution of NE neurons may vary depending on species.
HP increased DA and DOPAC levels in the telencephalon at 15 and 30 min, respectively (Fig. 3a, b), whereas HP did not affect NE level in the telencephalon, suggesting that HP-induced DA is not merely a precursor of NE. An immunohistochemical study in goldfish demonstrated that DA-IR perikarya are present in the medial part of the central telencephalon [nucleus pars medialis, (NPM)] (Goping et al. 1995). Additionally, TH-IR cells have been found in a similar region of the telencephalon in rainbow trout (Vetillard et al. 2002). In goldfish, DA neurons in the NPM are likely to be equivalent to those in the substantia nigra of the mesencephalon in mammals, which play a key role in locomotion (Goping et al. 1995; Pollard et al. 1992). When goldfish are treated with the neurotoxin 1-methyl-4-phenyl1-1,2,3,6-tetrahydropyriine (MPTP), MPTP destroys major DA neurons in the NPM and causes reduced movement, as seen in Parkinsonian syndrome (Goping et al. 1995; Pollard et al. 1992, 1996; Weinreb and Youdim 2007). A similar report in European river lamprey, Lampetra fluviatilis, indicated that MPTP injection causes greater DA depletion in the forebrain (i.e., nucleus tuberculi of the telencephalon) compared with other brain regions and impairs motor behavior (Thompson et al. 2008). These observations reinforce involvement of forebrain dopaminergic neurons in fish locomotion. Additionally, locomotion is regulated by DA receptors (D1-like receptors) expressed in the telencephalon of the cichlid genus Oreochromis (Mok and Munro 1998). When apomorphine (non-selective DA agonist that activates both D1-like and D2-like receptors) is added to the water, it causes a significant increase in locomotion. This effect is abolished by the D1 antagonist SCH-23390. In contrast, the D2 antagonist metoclopramide has no effect. Additionally, removing both hemispheres of the telencephalon abolishes the response to apomorphine, whereas removal of only one hemisphere has no effect on locomotion. These observations suggest that locomotion is regulated by DA, which is synthesized and released in the telencephalon of fish. In the present study, HP increased DA level in the telencephalon at 15 min (Fig. 3a). In addition, HP also increased locomotion at 15 min (Fig. 6). Therefore, it seems likely that HP leads to an increase in locomotion by inducing synthesis and release of DA in the telencephalon of goldfish at 15 min. However, the present study could not clarify direct relationship between HP-induced DA and HP-induced locomotion. HP also increased NE and 5-HIAA levels in the cerebellum at 30 min (Fig. 4a, c). However, HP did not affect 5-HT levels (Fig. 4b). Meanwhile, the effect of HP on levels of NE metabolites including 3-methoxy-4-hydroxyphenylglycol and 3-methoxy-4-hydroxymandelic acid was unclear because we could not measure those levels. An immunocytochemical study in goldfish demonstrated a wealth of NE and 5-HT fibers in the corpus cerebelli (CC) of the cerebellum, but no NE and 5-HT perikarya in the cerebellum (Hornby and Piekut 1990; Kah and Chambolle 1983), demonstrating that NE and 5-HT in the CC are transported through axons from other brain regions in goldfish. Similarly, NE and 5-HT are transported to the cerebellum through axons from the locus coeruleus of the dorsal pons and the dorsal raphe nucleus in mammals, respectively (Gnegy 2012; Hensler 2012). These results suggest that HP induces transport of NE through axons to the cerebellum and that HP induces release of 5-HT in the cerebellum. The CC plays a key role in controlling tail-beat frequency (swimming speed) in rainbow trout (Roberts et al. 1992) and goldfish (Matsumoto et al. 2007). In mouse, NE in the cerebral cortex, stratum and cerebellum is involved in modulating locomotion (Nishi et al. 1991). When NE terminals were destructed by N-ethyl-2-bromobenzylamine (DSP4) in the cerebral cortex, striatum and cerebellum, locomotion significantly was reduced (Nishi et al. 1991). In the present study, HP significantly increased cerebellar NE contents at 30 min. In addition, HP also increased locomotion at 30 min. These results suggest that HP-induced NE in the cerebellum might be involved in locomotion. Meanwhile, it is also possible that cerebellar 5-HT is involved in locomotion. However, cerebellar 5-HT seems to be involved in other function such as neuromodulation because it is considered that function of 5-HT in locomotion reduces activity in goldfish (Fenwick 1970).
HP decreased 5-HT and 5-HIAA contents in the vagal lobe at 30 and 60 min, respectively (Fig. 5a, b). Serotonergic perikarya are not present in the vagal lobe, but numerous fibers are present in goldfish (Kah and Chambolle 1983). These results suggest that HP inhibits axonal transport of 5-HT to the vagal lobe and release of 5-HT in the vagal lobe. The vagal lobe is involved in swallowing in goldfish (Finger 2008), crucian carp (Morita et al. 1980) and bullhead catfish, Ictalurus natalis (Atema 1971): When selective ablations of the vagal lobe were performed in the bullhead catfish, the fish could not swallow food (Atema 1971). However, the effect of HP on swallowing in the present study was unclear due to overhead video recording of fish behavior.
5-HT is a well-known modulator of γ-aminobutyric acid (GABA), glutamate and aspartate neurons in addition to neurotransmitter function. In fact, 5-HT is involved in modulating GABA-elicited inhibition of cerebellar Purkinje cells (Strahlendorf et al. 1989), glutamate-induced excitation of Purkinje cells (Lee et al. 1985) and aspartate-induced excitation of deep cerebellar nuclei neurons in rats (Gardette et al. 1987). In the CC of goldfish, the Purkinje cells and eurydendroid cells utilize GABA and aspartate, respectively, as neurotransmitters (Ikenaga et al. 2005, 2006). Additionally, GABA is likely to play an important role in the sensory layer of the goldfish vagal lobe (Contestabile et al. 1986), whereas glutamate mainly acts as a neurotransmitter for both the primary gustatory afferent fibers and the reflex interneurons (Finger 2008). Therefore, HP-affected 5-HT might be involved in modulation of GABA, aspartate and/or glutamate neuronal activities in the cerebellum and vagal lobe.
Locomotor activity was higher in the HP group (water depth of 1.3 m) than that in the control group (water depth of 0 m) (Fig. 6). In addition, Macdonald et al. (1987) have reported that HP induced swimming activity within habitat depth in European plaice Pleuronectes platessa, European eel and Atlantic cod Gadus morhua. At HP 1–50 atm (water depth 1–50 m), fish were stationary (European plaice), resting (Atlantic cod), slow exploratory (European eel). In contrast, at HP 50–100 atm, fish were upward swimming (European plaice and European eel) and attempted to upward swimming (Atlantic cod) (Macdonald et al. 1987). Thus, we hypothesized that fish locomotion (swimming speed) varies depending on water depth (HP) within their normal habitat depth: Fish choose deeper waters when they need to swim faster. Some supporting results have been reported on the relationship between swimming speed and water depth in some fish (Block et al. 1992; Tanaka et al. 2005). For example, an acoustic telemetry study in the blue marlin, Makaira nigricans, demonstrated that when fish are at a depth >50 m, they swim faster (80–120 cm/s). In contrast, the fish swim slowly (15–25 cm/s) in near-surface (within 10 m of surface) waters (Block et al. 1992). Additionally, a data logger study in chum salmon, O. keta, showed that hourly average swimming speeds were positively correlated with water depth during oceanic migration (Tanaka et al. 2005). Accordingly, fish might choose water depth according to the swimming speed demand. However, it is difficult to evaluate the relationship between water depth and swimming speed in fish in the field because several factors influence the choice of water depth, such as pursuit of prey and/or avoidance of predators. It will be necessary to evaluate the effect of water depth on swimming speed by excluding other factors without HP under laboratory conditions. Therefore, goldfish would be a good model for any such study. However, the protocol for testing the effect of HP on locomotion is defective in the present study because we recorded locomotion in HP group after transfer from high pressure to control pressure. Accordingly, we could not exclude another possibility that the increase in goldfish locomotion observed is the result of the transfer from high pressure to lower pressure.
Swimming activities of most marine fish are synchronized with tidal cycles. In fact, there are many reports on tidal-synchronized locomotion rhythm with peaks during high tides in intertidal fish, including shanny, Lipophrys pholis (Northcott et al. 1991a, b); Montagu’s blenny, Coryphoblennius galerita (Gibson 1970) and rocky goby, Gobius pagnellus (Northcott 1991). Additionally, when those fish are exposed to tidal-cyclic HP in an aquarium, locomotion is synchronized with changes in HP with a peak during the highest HP (Gibson 1970; Northcott 1991; Northcott et al. 1991a, b). Accordingly, Northcott (1991) reported that HP is probably the most reliable tidal cue for intertidal fish because it is less affected by weather conditions and seasonal changes. Therefore, the tidal-related locomotion in intertidal fish could also be explained by HP-induced locomotion in goldfish.
In conclusion, HP (water depth of 1.3 m) affected the contents of monoamines and their metabolites in the telencephalon, cerebellum and vagal lobe of goldfish. In particular, HP increased DA content in the telencephalon at 15 min and NE content in the cerebellum at 30 min. In addition, HP also induced locomotion at 15 and 30 min. These results suggest that HP induced fish locomotion via the telencephalic DA and cerebellar NE neuronal activity. However, we could not exclude the possibility that locomotion observed is the result of the transfer from high pressure to lower pressure. Further studies will be needed to clarify direct relationship between HP-induced locomotion and telencephalic DA and cerebellar NE system by inhibitory experiment such as DA and NE antagonists.
References
Atema J (1971) Structures and functions of the sense of taste in the catfish (Ictalurus natalis). Brain Behav Evol 4:273–294
Block BA, Booth D, Carey FG (1992) Direct measurement of swimming speeds and depth of blue marlin. J Exp Biol 166:267–284
Bonn U (1987) Distribution of monoamine-containing neurons in the brain of a teleost, Carassius auratus (Cyprinidae). J Hirnforsch 28:529–544
Contestabile A, Villani L, Bissoli R, Poli A, Migani P (1986) Cholinergic, GABAergic and excitatory amino acidic neurotransmission in the goldfish vagal lobe. Exp Brain Res 63:301–309
Corio M, Peute J, Steinbusch HWM (1991) Distribution of serotonin- and dopamine-immunoreactivity in the brain of the teleost Clarias gariepinus. J Chem Neuroanat 4:79–95
Damasceno-Oliveira A, Fernández-Durán B, Gonçalves J, Serrão P, Soares-Da-Silva P, Reis-Henriques MA, Coimbra J (2006) Effects of cyclic and constant hydrostatic pressure on norepinephrine and epinephrine levels in the brain of flounder. J Fish Biol 68:1300–1307
Damasceno-Oliveira A, Fernandez-Duran B, Goncalves J, Serrao P, Soares-da-Silva P, Reis-Henriques MA, Coimbra J (2007) Effects of cyclic hydrostatic pressure on the brain biogenic amines concentrations in the flounder, Platichthys flesus. Gen Comp Endocrinol 153:385–389
De Pedro N, Pinillos ML, Valenciano AI, Alonso-Bedate M, Delgado MJ (1998) Inhibitory effect of serotonin on feeding behavior in goldfish: involvement of CRF. Peptides 19:505–511
Dietrich M, Hofmann MH, Bleckmann H (2002) Effects of dopaminergic drugs and telencephalic ablation on eye movements in the goldfish, Carassius auratus. Brain Res Bull 57:393–395
Fenwick JC (1970) Brain serotonin and swimming activity in the goldfish, Carassius auratus. Comp Biochem Physiol 32:803–806
Finger TE (2008) Sorting food from stones: the vagal taste system in goldfish, Carassius auratus. J Comp Physiol A 194:135–143
Fraser PJ, Macdonald AG (1994) Crab hydrostatic pressure sensors. Nature 371:383–384
Fraser PJ, Shelmerdine RL (2002) Dogfish hair cells sense hydrostatic pressure. Nature 415:495–496
Fraser PJ, Cruickshank SF, Shelmerdine RL (2003) Hydrostatic pressure effects on vestibular hair cell afferents in fish and crustacea. J Vestib Res 13:235–242
Fremberg M, van Veen T, Hartwig HG (1977) Formaldehyde-induced fluorescence in the telencephalon and diencephalon of the eel (Anguilla anguilla l.). A fluorescence-microscopic and microspectrofluorometric investigation with special reference to the innervation of the pituitary. Cell Tissue Res 176:1–22
Gardette R, Krupa M, Crepel F (1987) Differential effects of serotonin on the spontaneous discharge and on the excitatory amino acid-induced responses of deep cerebellar nuclei neurons in rat cerebellar slices. Neuroscience 23:491–500
Geffard M, Kah O, Onteniente B, Seguela P, Le Moal M, Delaage M (1984) Antibodies to dopamine: radioimmunological study of specificity in relation to immunocytochemistry. J Neurochem 42:1593–1599
Gibson RN (1970) The tidal rhythm of activity of Coryphoblennius galerita (L.) (Teleostei, Blenniidae). Anim Behav 18:539–543
Gibson RN (1984) Hydrostatic pressure and the rhythmic behaviour of intertidal marine fishes. Trans Am Fish Soc 113:479–483
Gibson RN (1992) Tidally-synchronised behaviour in marine fishes. In: Ali MA (ed) Rhythms in fishes. Springer, New York, pp 63–81
Gnegy ME (2012) Catecholamine. In: Brady ST, Siegel GJ, Albers RW, Price DL (eds) Basic neurochemistry: Principles of molecular, cellular, and medical neurobiology. Elsevier Academic Press, Amsterdam, pp 283–293
Godø OR, Michalsen K (2000) Migratory behaviour of north-east Arctic cod, studied by use of data storage tags. Fish Res 48:127–140
Goping G, Pollard HB, Adeyemo OM, Kuijpers GA (1995) Effect of MPTP on dopaminergic neurons in the goldfish brain: a light and electron microscope study. Brain Res 687:35–52
Gotow T, Triller A, Korn H (1990) Differential distribution of serotoninergic inputs on the goldfish Mauthner cell. J Comp Neurol 292:255–268
Hensler JG (2012) Serotonin. In: Brady ST, Siegel GJ, Albers RW, Price DL (eds) Basic neurochemistry: Principles of molecular, cellular, and medical neurobiology. Elsevier Academic Press, Amsterdam, pp 300–322
Hornby PJ, Piekut DT (1990) Distribution of catecholamine-synthesizing enzymes in goldfish brains: presumptive dopamine and norepinephrine neuronal organization. Brain Behav Evol 35:49–64
Ikenaga T, Yoshida M, Uematsu K (2005) Morphology and immunohistochemistry of efferent neurons of the goldfish corpus cerebelli. J Comp Neurol 487:300–311
Ikenaga T, Yoshida M, Uematsu K (2006) Cerebellar efferent neurons in teleost fish. Cerebellum 5:268–274
Janssen J, Brandt SB (1980) Feeding ecology and vertical migration of adult alewives (Alosa pseudoharengus) in lake Michigan. Can J Fish Aquat Sci 37:177–184
Kah O, Chambolle P (1983) Serotonin in the brain of the goldfish, Carassius auratus. An immunocytochemical study. Cell Tissue Res 234:319–333
Kapsimali M, Vidal B, Gonzalez A, Dufour S, Vernier P (2000) Distribution of the mRNA encoding the four dopamine D1 receptor subtypes in the brain of the european eel (Anguilla anguilla): comparative approach to the function of D1 receptors in vertebrates. J Comp Neurol 419:320–343
Koizumi I, Kanazawa Y, Tanaka Y (2013) The fishermen were right: experimental evidence for tributary refuge hypothesis during floods. Zool Sci 30:375–379
Lee M, Strahlendorf JC, Strahlendorf HK (1985) Modulatory action of serotonin on glutamate-induced excitation of cerebellar purkinje cells. Brain Res 361:107–113
Lillesaar C (2011) The serotonergic system in fish. J Chem Neuroanat 41:294–308
Linard B, Bennani S, Jego P, Saligaut C (1996) Tyrosine hydroxylase activity and dopamine turnover of rainbow trout (Oncorhynchus mykiss) brain: the special status of the hypothalamus. Fish Physiol Biochem 15:41–48
Macdonald AG, Gilchrist I, Wardle CS (1987) Effect of hydrostatic pressure on the motor activity of fish from shallow water and 900 m depths: some results of challenger cruise 6B/85. Comp Biochem Physiol 88:543–547
Matsumoto N, Yoshida M, Uematsu K (2007) Effects of partial ablation of the cerebellum on sustained swimming in goldfish. Brain Behav Evol 70:105–114
McCleave JD, Arnold GP (1999) Movements of yellow- and silver-phase European eels (Anguilla anguilla L.) tracked in the western North Sea. ICES J Mar Sci 56:510–536
Mok EY, Munro AD (1998) Effects of dopaminergic drugs on locomotor activity in teleost fish of the genus Oreochromis (Cichlidae): involvement of the telencephalon. Physiol Behav 64:227–234
Morita Y, Ito H, Masai H (1980) Central gustatory paths in the crucian carp, Carassius carassius. J Comp Neurol 191:119–132
Nilsson GE (1989) Regional distribution of monoamines and monoamine metabolites in the brain of the crucian carp (Carassius carassius L.). Comp Biochem Physiol C 94:223–228
Nishi K, Kondo T, Narabayashi H (1991) Destruction of norepinephrine terminals in 1-metyl-4phenyl-1,2,3,6-tetrahydropyridine (MPTP)-treated mice reduces locomotor activity induced by L-DOPA. Neurosci Lett 123:244–247
Northcott SJ (1991) A comparison of circatidal rhythmicity and entrainment by hydrostatic pressure cycles in the rock goby, Gobius paganellus L. and the shanny, Lipophrys pholis (L.). J Fish Biol 39:25–33
Northcott SJ, Gibson RN, Morgan E (1991a) Phase responsiveness of the activity rhythm of Lipophrys pholis (L.) (Teleostei) to a hydrostatic pressure pulse. J Exp Mar Biol Ecol 148:47–57
Northcott SJ, Gibson RN, Morgan E (1991b) The effect of tidal cycles of hydrostatic pressure on the activity of Lipophrys pholis (L.) (Teleostei). J Exp Mar Biol Ecol 148:35–45
Nunn AD, Copp GH, Vilizzi L, Carter MG (2010) Seasonal and diel patterns in the migrations of fishes between a river and a floodplain tributary. Ecol Freshw Fish 19:153–162
Olcese JM, Hall TR, Figueroa HR, de Vlaming VL (1979) Hypothalamic monoamine oxidase, a component in the serotonergic control of pituitary prolactin content in Carassius auratus L. Gen Comp Endocrinol 38:309–313
Pollard HB, Dhariwal K, Adeyemo OM, Markey CJ, Caohuy H, Levine M, Markey S, Youdim MB (1992) A parkinsonian syndrome induced in the goldfish by the neurotoxin MPTP. FASEB J 6:3108–3116
Pollard HB, Kuijpers GA, Adeyemo OM, Youdim MB, Goping G (1996) The MPTP-induced parkinsonian syndrome in the goldfish is associated with major cell destruction in the forebrain and subtle changes in the optic tectum. Exp Neurol 142:170–178
Raffaelli D, Richner H, Summers R, Northcott S (1990) Tidal migrations in the flounder (Platichthys flesus). Mar Behav Physiol 16:249–260
Roberts BL, van Rossem A, de Jager S (1992) The influence of cerebellar lesions on the swimming performance of the trout. J Exp Biol 167:171–178
Sebert P, Barthelemy L, Caroff J (1985) Serotonin levels in fish brain: effects of hydrostatic pressure and water temperature. Experientia 41:1429–1430
Sebert P, Barthelemy L, Caroff J (1986) Catecholamine content (as measured by the HPLC method) in brain and blood plasma of the EEL: effects of 101 ATA hydrostatic pressure. Comp Biochem Physiol C 84:155–157
Sébert ME, Weltzien FA, Moisan C, Pasqualini C, Dufour S (2008) Dopaminergic systems in the European eel: characterization, brain distribution, and potential role in migration and reproduction. In: Dufour S, Prévost E, Rochard E, Williot P (eds) Fish and diadromy in Europe (ecology, management, conservation). Springer, Netherlands, pp 27–46
Somoza GM, Peter RE (1991) Effects of serotonin on gonadotropin and growth hormone release from in vitro perifused goldfish pituitary fragments. Gen Comp Endocrinol 82:103–110
Strahlendorf JC, Lee M, Strahlendorf HK (1989) Modulatory role of serotonin on GABA-elicited inhibition of cerebellar Purkinje cells. Neuroscience 30:117–125
Takemura A, Uchimura M, Shibata Y (2010a) Dopaminergic activity in the brain of a tropical wrasse in response to changes in light and hydrostatic pressure. Gen Comp Endocrinol 166:513–519
Takemura A, Rahman MS, Park YJ (2010b) External and internal controls of lunar-related reproductive rhythms in fishes. J Fish Biol 76:7–26
Takemura A, Shibata Y, Takeuchi Y, Hur SP, Sugama N, Badruzzaman M (2012) Effects of hydrostatic pressure on monoaminergic activity in the brain of a tropical wrasse, Halicoeres trimaculatus: possible implication for controlling tidal-related reproductive activity. Gen Comp Endocrinol 175:173–179
Tanaka H, Naito Y, Davis ND, Urawa S, Ueda H, Fukuwaka M (2005) First record of the at-sea swimming speed of a Pacific salmon during its oceanic migration. Mar Ecol Prog Ser 291:307–312
Thompson RH, Menard A, Pombal M, Grillner S (2008) Forebrain dopamine depletion impairs motor behavior in lamprey. Euro J Neurosci 27:1452–1460
Vetillard A, Benanni S, Saligaut C, Jego P, Bailhache T (2002) Localization of tyrosine hydroxylase and its messenger RNA in the brain of rainbow trout by immunocytochemistry and in situ hybridization. J Comp Neurol 449:374–389
Vidal B, Pasqualini C, Le Belle N, Holland MC, Sbaihi M, Vernier P, Zohar Y, Dufour S (2004) Dopamine inhibits luteinizing hormone synthesis and release in the juvenile European EEL: a neuroendocrine lock for the onset of puberty. Biol Reprod 71:1491–1500
Weinreb O, Youdim MB (2007) A model of MPTP-induced Parkinson’s disease in the goldfish. Nat Proto 2:3016–3021
Wollmuth LP, Crawshaw L, Panayiotides-Djaferis H (1989) The effects of dopamine on temperature regulation in goldfish. J Comp Physiol B 159:83–89
Yoshida M, Nagatsu I, Kawakami-Kondo Y, Karasawa N, Spatz M, Nagatsu T (1983) Monoaminergic neurons in the brain of goldfish as observed by immunohistochemical techniques. Experientia 39:1171–1174
Acknowledgments
We would like to thank Ms. Chihiro Yamauchi, University of the Ryukyus, and Dr. Yoriko Akimoto, Mr. Takahiro Nakashita, Mr. Seiya Mochinaga and Mr. Takeshi Yoshihara, Kyushu University, for their support and help in sampling. Dr. Sethu Selvaraj, Kyushu University, Dr. Takeshi Onuma, Osaka University, and Mr. Makoto Yoshida, The University of Tokyo, provided stimulating discussions and helpful advice. This study was supported by a Grant-in-Aid for JSPS fellows (Grant Numbers 10J02886 and 12J02083) to TI from the Japan Society for the Promotion of Science.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ikegami, T., Takemura, A., Choi, E. et al. Increase in telencephalic dopamine and cerebellar norepinephrine contents by hydrostatic pressure in goldfish: the possible involvement in hydrostatic pressure-related locomotion. Fish Physiol Biochem 41, 1105–1115 (2015). https://doi.org/10.1007/s10695-015-0072-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10695-015-0072-7