
Abstract
The development of hybrid cultivars is one of the most important contributions of a breeding program. A high allelic fixation of the parent lines is essential to guarantee the uniformity of developed hybrids. In this sense, papaya segregation has been observed in the parent lines, impacting the pattern of the hybrids obtained and, consequently, their commercialization. Thus, this study aimed to evaluate the JS-12, Sekati, and SS-72/12 genotypes, which are parents of important papaya hybrids, to identify possible segregations and proceed with the generation progress through self-fertilization of selected genotypes for agronomic traits of interest. A mass selection was carried out, selecting 400 plants based on the phenotype, being: 100 plants from the Sekati parent, 100 from the SS-72/12 parent, and 200 plants from the JS-12 parent, of which 100 plants with smaller fruits (JS12-P) and 100 plants with larger fruits (JS12-G). These plants were agronomically evaluated for 15 traits in three evaluation periods. Descriptive statistics of the observations, diversity analysis, as well as the combined selection index and direct selection were performed. Through the descriptive and phenotypic variability analysis, it was possible to verify the existence of variability in the parents, enabling the selection of 80 genotypes, 72 of which were selected via combined selection and eight by direct selection. Given the existence of parents' segregation, the extreme importance of selecting lines with different patterns to be explored in the papaya breeding program is emphasized to purify the parents and adjust the hybrids to meet the consumer market's demand.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Papaya is one of the tropical fruit trees of great importance in Brazil, mainly grown in the northeast and southeast regions. In these regions, the states of Bahia, Espírito Santo, Ceará, and Rio Grande do Norte are the largest national producers, emphasizing the Bahia and Espírito Santo, representing approximately 66.8% of the national papaya production (IBGE 2019). However, the papaya crop demands increasingly yielding genotypes, nutritional and sensory quality, and disease resistance (Pereira et al. 2019a).
Papaya breeding programs have been carrying out various works to develop new cultivars to meet the demands mentioned above. It is worth mentioning that the knowledge of the inheritance of the traits is an important element to define the type of cultivar to be developed by a breeding program. Studies carried out by Cattaneo (2001) on the analysis of averages and variances in different generations allowed us to verify that the additive effects had magnitudes greater than the deviations of dominance for the number and fruit weight. On the other hand, the dominance deviations showed greater magnitude for fruit yield per plant. Thus, the study revealed that papaya has the viability of developing both pure line and hybrid cultivars. Works like these allow the breeder to direct a more efficient breeding program. Additionally, Vettorazzi (2020) observed that the trait average weight of the fruits might suffer from the action of overdominance effects, indicating that heterosis can be explored.
Because of the heterosis exploring feasibility in papaya and considering two heterotic groups (Solo and Formosa), studies were carried out to identify promising hybrid combinations for cultivars development. The research explored the heterotic intra- and intergroup variability, culminating in lines and hybrids developing with Solo and Formosa patterns (Luz et al. 2015; Luz et al. 2018; Pereira et al. 2019a, b, c; Vettorazzi 2020).
It is expected that lines present allelic fixation equal to 1.00, however, molecular characterization studies (Pirovani et al. 2021; Rodrigues 2021) point to a level of allelic fixation below that expected for the JS-12 lines, showing the existence of variability. This intragenotypic variation is not desirable in hybridization programs, indicating the need to direct efforts towards advancing self-fertilization generations, guarantee the fixation of loci in heterozygosis, and, consequently, achieve the uniformity of hybrids.
This intragenotypic variation in parent lines has been observed in papaya crops, culminating in variations in the agronomic traits of hybrids, especially regarding yield and quality aspects. One reason for the lack of purity in the parental lines may be the mixture of seeds (physical contamination) and pollen contamination in the seed production fields (genetic contamination) (MAPA 2011). According to the Ministry of Agriculture, Livestock, and Supply (2011), genetic and physical contamination sources must be studied, and their degree of occurrence estimated. In genetic contamination, plants from other cultivars of the same crop and plants of similar species pollinate the crop. This crossing alters the genetic constitution of seeds, which is no longer representative of the production cultivar (MAPA 2011).
In cultivars of 'Formosa' papaya plants tend to have a marked allogamy rate, promoting genetic recombinations and, therefore, preserving a certain variability genetics (Silva et al. 2007). According to the authors due to the high allogamy rate in the 'Formosa' genotypes, the parents available have variable degrees of loci in heterozygous and the evolution of the fixation degree of the parents must therefore be monitored in order to produce vigorous and uniform hybrids.
The purification of parental lines was reported in rice (Ingale and Waghmode 2005) however, the literature still does not report work on adjustments in papaya hybrids, with parent purification and the possibility of generating new hybrid combinations a challenge for the breeding program. Thus, this research aimed to study the segregation of the JS-12, Sekati, and SS-72/12 parents, who are parents of important hybrids, to reduce the size of the 'UC10' and increase the 'Calimosa' fruit according to market demand, as well as increasing uniformity during the production process. To this end, a diversity study in these genotypes was carried out, as well as the combined and direct selection based on the analysis of fifteen morpho-agronomic traits, to identify and select parents with the traits mentioned above and proceed with the advance of self-fertilization generation for the increase the fixation index in the lines.
Material and methods
Genetic material and experimental conditions
The 'Sunrise Solo' 72/12 (SS-72/12) from the Solo group and the JS-12 from the Formosa group are elite genotypes used to obtain hybrids (Barros et al. 2017; Santa-Catarina et al. 2019; Pereira et al. 2019c). The parent Sekati is used as a source of favorable alleles for reducing the severity of black spots and powdery mildew (Vivas et al. 2013a), besides its good general combining ability regarding resistance to phoma-spot in papaya (Vivas et al. 2013b).
The JS-12 and Sekati parents belong to the same heterotic group (Formosa); however, they contrast agronomic and sensory traits (Cardoso et al. 2014; Cortes et al. 2019). These genotypes also differ in plant and fruit morphology, with Sekati being represented by shorter plants with more dense leaves and fruits. In comparison, JS12 has taller plants and elongated fruits with pointed ends and a smaller diameter.
The experiment was carried out at the Caliman Agrícola S/A company, located at 19º15' S and 40º10' W, in Linhares, Espírito Santo state, from 2018 to 2020. The climate of the region is classified as AWI-type (humid tropical), with rainy summer and dry winter (Alvares et al. 2013). The seeds of the JS-12, Sekati, and SS-72/12 genotypes used by the company were sown in a greenhouse at the Santa Terezinha Farm in December 2018, using plastic trays, with a capacity for 96 tubes of 55 cm3. The plant substrates HT Tropstrato and the Basacote mini 3 M® fertilizer, NPK + (Mg) formulation 13–06-16 (1.4) with micronutrients: 0.0; 2.5; 5.0; 7.5; 10.0, and 12.5 kg m−3 of Controlled Release Fertilizer.
Thirty days after sowing, the Sekati, JS-12, and SS-72/12 seedlings genotypes were planted in the seed production area of the company, using three seedlings per hole at a distance of 3.6 m between the rows and 1.5 m between plants in the row. Ninety days after planting, the thinning of the plants was carried out, just hermaphrodite plants were used for the work since they produce the piriform fruits of interest for commercialization. The cultural treatments were the same used in the commercial plantations of the company.
Mass selection
Ninety days after thinning, a mass selection was carried out based on the phenotype, selecting plants with smaller and larger fruits: 100 plants from the parent Sekati and 100 plants from the parent SS-72/12, and 200 from the parent JS- 12, totaling 400 plants. The latter parent was divided into two categories, with 100 plants selected for each. The first category is for plants with smaller fruits (JS12-P), and the second for plants with larger fruits (JS12-G). Papaya cultivation is very susceptible to disease attack, causing plant loss. Thus, agronomic evaluations were completed in 264 plants used in the analyses carried out in this study.
Evaluated traits
Digital phenotyping in the field
The plant height (PH), first fruit insertion height (FFIH), stem diameter (SD), number of commercial fruits (NCF), number of deformed fruits (NDF), fruitless nodes (FN) were assessed via digital phenotyping according to a methodology developed and validated by Cortes et al. (2017). A Sony DSCHX 300 digital camera was used, using two complete photos per plant, one photo from side A and the other side B. The evaluations were carried out in three periods: 180, 270, and 360 days after planting. Next to each plant stem, a label was placed to facilitate identification during analysis and image processing. In each evaluation, the plants were also marked with red wool to identify the last fruit evaluated in the bunch. A known measurement scale was placed on the soil, close to the stem of the evaluated plants, for later calibration of the processed and analyzed images using the ImageJ v1.50c software. The FW traits was obtained through the average of five fruits measured with digital scale, Toledo brand—model 9094, and expressed in grams and the fruit yield (YIELD) was obtained by multiplying the FW by the NCF.
Determinations of physical and chemical traits
The evaluations were carried out at 270, 360, and 450 days after planting. In each evaluation, the fruits that showed the same maturation stage (RST1) were harvested, where the fruit reaches its maximum physical development and could be harvested (Barragán-Iglesias et al. 2018).
After weighing, each fruit was cut in half along its longitudinal axis. Subsequently, half of the fruit was scanned using a Pro A320 optical model bench scanner coupled to a personal notebook to obtain the images and perform the evaluation of the following morphological traits: fruit length (FL); fruit diameter (FD); ovarian cavity length (OCL); ovarian cavity diameter (OCD), and pulp thickness (PT) using the digital image analysis and processing methodology described and validated by Santa-Catarina et al. (2018), using the ImageJ v1.50c software. The fruit volume (FV), pulp volume (PV), and the percentage of pulp volume (PV%) were calculated using the following equations:
where: VF, OCV, and VP are the fruit, ovarian cavity, and pulp estimated volumes, respectively. FL and OCL are the fruit and the ovarian cavity lengths, FD and OCD are the fruit and the ovarian cavity diameters. The other half of the fruit was used to measure the fruit and the pulp firmness (FF and PF, respectively). Firmness was assessed by penetration resistance using a digital Bench Penetrometer (Fruit Pressure Tester, Italy, Model 53,205). For FF, perforations were performed at three points equidistant in the equatorial region of three fruits, and the PF was measured from the perforation of three points equidistant in the pulp. The soluble solids content (SST) was obtained through a portable digital refractometer, as stated by the Association of Official Analytical Chemists (AOAC 1990).
Descriptive statistics
The FN, NDF, NCF traits were obtained by adding the three evaluation periods. The FFIH trait was evaluated only in the first evaluation (where we have the first fruit). In contrast, the average values of PH, SD, FW, FF, PF, SST, FL, FD, PT, and PV% traits were estimated in the three periods evaluated. The YIELD was obtained by multiplying the sum of the NCF with the FW. The mass selection was carried out in a seed production field, that is why the experimental design was not used. Thus, it followed with descriptive statistics (minimum, maximum, first quartile, median, and third quartile) and presentation of results in a boxplot graphic obtained through the R program (R Core Team 2018).
Phenotypic variability
Multivariate analysis was used to estimate the phenotypic variability between the analyzed genotypes. Initially, the Euclidean distance was estimated to obtain the phenotypic dissimilarity matrix, considering the 15 morphological traits evaluated (PH, FFIH, SD, NCF, NDF, FN, FW, YIELD, FF, PF, SST, FL, FD, PT, and PV%). Subsequently, cluster analysis was performed using the Unweighted Pair-Group Method using an Arithmetic Average (UPGMA). Such analyses were performed with the R software aid (R Core Team 2018; Gu 2014). The relative contribution of each trait to phenotypic diversity was also estimated using Singh's methodology (1981). The analysis of genetic relationships among genotypes was also assessed with graphic dispersion via principal coordinate analysis (PCA).
Combined selection and direct selection
Fifteen traits were considered simultaneously through the combined selection index (CSI) to select the genotypes. This index was proposed by Silva et al. (2008a) and rectified by Ramos et al. (2014) and can be estimated using the following equation:
where: VFt is the standardized phenotypic means of trait i, (obtained using the following equation: (Xg − Ẋg) / SXg, where: Xg is the individual measured value for the x trait; Ẋg is the general mean of the trait, and SXg is the standard deviation), and p is the agronomic weight established for each I trait and can be positive or negative according to the direction of the selection.
The agronomic weights established per attempt for the fifteen traits were: PH (1), FFIH (-10), SD (5), NCF (100), NDF (-20), FN (-20), FW (200 (JS12 -G and SS-72/12) and -200 (Sekati and JS12-P)), YIELD (100), FF (100), PF (100), SST (100), FL (1), FD (1), PT (70), and PV% (50). These weights were established experimentally established, based on the breeders' knowledge of the UENF/CALIMAN papaya breeding program, the agronomic importance of the evaluated traits, and the work's objective. As in the study, each plant is a different individual, negative (-200) and positive (200) weights were attributed to the FW trait to identify possible variations of the respective parents with different fruit sizes. Microsoft Office Excel (2019) was used for CSI estimations. Two genotypes per parent were also selected via direct selection (DS) based on the FW trait to maintain in the lines to be purified alleles genotypes sources for this attribute.
Results
Descriptive statistics
Figure 1 shows the observations centrality measure, using the average and the dispersion of the observations around this parameter. The quartiles represent the dispersion measure for 15 morpho-agronomic traits (PH, FFIH, SD, NCF, NDF, FN, FW, YIELD, FF, PF, SST, FL, FD, PT, and PV%) evaluated in the parents JS12-P, JS12-G, Sekati, and SS-72/12. For the NCF, FF, PF, FW, YIELD, and SST traits considered the most relevant in the papaya breeding program, we can see great variation among the evaluated parents, especially FW, allowing genotype selection for larger or smaller fruits. The FF and PF traits in the JS12-P, JS12-G, Sekati genotypes showed less phenotypic variation than the SS-72/12 genotype. Additionally, the SS-72/12 genotype also showed low phenotypic variation for FN, FW, YIELD, FL, FD, and PT.

Boxplot based on 15 traits evaluated in 264 papaya genotypes of the S12-P, JS12-G, Sekati, and SS-72/12 parents. PH plant height; FFIH height of insertion of the first fruit; SD stem diameter; NCF number of commercial fruits; NDF number of deformed fruits; FN fruitless nodes; FW average fruit weight; YIELD fruit yield; FF fruit firmness; PF pulp firmness; SST soluble solids content; FL fruit length; FD fruit diameter; PT pulp thickness; PV% pulp volume percentage
Phenotypic variability
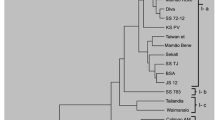
The diversity analysis, obtained by the Euclidean distance, considering the morpho-agronomic traits (PH, FFIH, SD, NCF, NDF, FN, AMF, YIELD, FF, PF, SST, FL, FD, PT, and PV%) indicated the formation of six clusters (Fig. 2).

Dendrogram obtained by the Euclidean distance using the UPGMA clustering method based on the analysis of 264 plants for 15 evaluated traits (co-phenetic correlation coefficient = 0.80)
Cluster I (yellow color) has 33 genotypes (20 genotypes from Sekati and 13 from JS12-G), followed by cluster II (green color) with 82 genotypes (4 Sekati, 48 JS12-G, and 30 JS12- P), cluster III (purple color) with five genotypes (4 Sekati and 1 JS12-G), cluster IV (blue color) with 49 genotypes (11 Sekati and 38 JS12-G), cluster V (black color) with 47 genotypes (46 JS12-P and 1 Sekati), cluster VI (red color) with 48 genotypes (all SS-72/12) (Fig. 2).
Figure 3 shows the relative contribution of traits to phenotypic variability among the individuals evaluated. It appears that the trait that provided the greatest relative contribution was the average weight of the fruits with 98.59%, followed by the plant height with 0.79%, the number of commercial fruits with 0.18%, fruit yield with 0.16%, first fruit insertion height with 0.14%, and fruit firmness with 0.02%.

Relative genetic contribution of the traits evaluated in 264 papaya genotypes of the S12-P, JS12-G, Sekati, and SS-72/12 parents. FW average fruit weight; PH plant height; NCF number of commercial fruits; YIELD fruit yield; FFIH height of insertion of the first fruit and FF fruit firmness, estimated by the methodology proposed by Singh (1981)
Figure 4 shows the average values of the morpho-agronomic traits FW, PH, NCF, YIELD, FFIH, and FF for each cluster. The FW trait ranged from 380.90 g for cluster VI to 2146 g for cluster III. The number of fruits with commercial standard varied from 27.48 fruits for cluster I (formed by genotypes of the Formosa cluster) to 69.75 fruits for cluster VI (formed by the Solo cluster genotype). The YIELD ranged from 26.41 kg plant−1 (cluster VI) to 62.08 kg plant−1 (cluster III), and the FF ranged from 124.14 N for cluster VI to 131.44 N for cluster II (Fig. 4). From these numbers, it is possible to verify the existence of variability in the genetic materials under study, both for the traits of fruits related to yield and quality.

Graph of average values of clusters for the six morpho-agronomic traits evaluated in 264 plants. FW average fruit weight; PH plant height; NCF number of commercial fruits; YIELD fruit yield; FFIH height of the first fruit insertion and FF fruit firmness
The analysis of genetic relationships among genotypes was also assessed with graphic dispersion via principal coordinate analysis (Fig. 5a). The first 2 coordinates together explained 54.9% of the total data variation, whereas 40.1% of this variation was explained by coordinate 1 and 14.8% by coordinate 2 for 15 traits (Fig. 5b).

Principal coordinates analysis based on 15 traits for Carica papaya. a Principal coordinates analysis for papaya genotypes of the JS12-P, JS12-G, Sekati, and SS-72/12 parents. b Variables analyzed for papaya genotypes of the S12-P, JS12-G, Sekati, and SS-72/12 parents. PH plant height; FFIH height of insertion of the first fruit; SD stem diameter; NCF number of commercial fruits; NDF number of deformed fruits; FN fruitless nodes; FW average fruit weight; YIELD fruit yield; FF fruit firmness; PF pulp firmness; SST soluble solids content; FL fruit length; FD fruit diameter; PT pulp thickness; PV% pulp volume percentage
Combined selection and direct selection
Based on phenotypic averages, 80 genotypes were selected by combined and direct selection, considering the 15 traits evaluated in the 264 plants of the three parents. Tables 1, 2, 3, and 4 show the genotypes selected for the parents JS12-P, JS12-G, Sekati, and SS-72/12, respectively. For each parent, 18 genotypes were selected through the CSI according to their standardized phenotypic averages. Two genotypes were selected via direct selection for the FW trait to maintain in the lines to be purified, alleles genotypes sources for the standard of desired fruit, totaling 20 genotypes per parent.
Based on the selection differential (Table 1), a reduction in NDF, FN, and fruit size, as well as increased values of important traits such as NCF, YIELD, FF, PF, SST, PV%, were observed. For the FW attribute, the average of the selected genotypes for the parent JS12-P (Table 1) was 918.10 g, lower than the general average (951.7 g), and the average of JS12-G (Table 2) was 1567.00 g, higher than the general average (1355.87 g).
Regarding the genotypes selected for the Sekati parent (Table 3), a reduction in the average values for the FW, FFIH, FN, FL, FD, and PT, as well as increased values of important traits such as NCF, YIELD, FF, PF, SST, PV%, were observed.
When analyzing the traits related to the production of plants and fruit quality selected for the parent SS-72/12 (Table 4), the phenotypic average for the genotypes selected by the CSI was higher than the general average except for FFIH, NDF, FN.
Discussion
The 'Calimosa' hybrid, the first Brazilian papaya hybrid, has shown superiority for fruit production and quality compared to the 'Tainung 01' hybrid of the Formosa group, constituting an alternative for the papaya cultivation, not only in the country but also for farmers in other countries by exporting its seeds (Pereira et al. 2015). The 'UC10' hybrid, according to Pereira et al. (2019c), has excellent agronomic performance with standard Formosa fruits and excellent flavor. However, a loss of these hybrids pattern has been observed, with a considerable increase in fruit size in 'UC10' and reduced fruit size in 'Calimosa'. This fact points to an important and not addressed detail: the hybrids are segregating, so it is of fundamental importance to monitor the parent lines.
It is evident that the parental lines can be purified and improve the commercial hybrid in Fig. 1, we can see that the parents JS12-P, JS12-G, Sekati, and SS-72/12 showed genetic variation, mainly for the FW, which is extremely important for determining the fruit pattern of cultivars UC10 and Calimosa. This observation indicates the possibility of selecting parents with larger fruits, increasing the average fruit weight of the 'Calimosa' hybrid, and parents with smaller fruits to reduce the average fruit weight of the 'UC10' hybrid.
Concerning phenotypic variability, cluster II brought together the largest genotype number presenting in its composition Sekati, JS12-G, and JS12-P, all from the Formosa group. Clusters I, III, and IV were also made up of Sekati and JS12-G genotypes, while cluster V brought together Sekati and JS12-P genotypes. On the other hand, the genotypes of the Solo group (SS-72/12) formed a single cluster (VI) (Fig. 2).
Thus, it is evident that the genotypes are grouped by the heterotic groups Solo and Formosa, which differ basically by the size of the fruit. Some Sekati genotypes were grouped with JS12-P and JS12-G, indicating variability in these genotypes and the possibility of selecting genotypes with larger fruits for the parents of 'Calimosa' and smaller fruits for the parents of 'UC10'.
It can be seen in Fig. 3 that the FW is a trait that has the greatest contribution to explain the total variation available in the population evaluated in this study, while the FF has a low contribution and can be discarded in evaluations of genetic divergence. This result can be attributed to evaluating individuals from different heterotic groups, where the variation in fruit sizes is extremely noticeable.
The average values of clusters for the 15 morpho-agronomic traits (PH, FFIH, SD, NCF, NDF, FN, FW, YIELD, FF, PF, SST, FL, FD, PT, and PV%) evaluated in 264 plants (Fig. 4), demonstrate the existence of genetic variability in the different genotypes evaluated, in which the selection can be applied. It is worth highlighting the FW trait for having the greatest variation. This fact clarifies the existing variability between and within the evaluated genotypes, enabling the selection of genotypes that may be used in hybridization programs as parent lines with different FW in the future, making it possible to obtain hybrids with different fruit sizes.
Based on the selection differential of the genotypes selected for the JS12-P parent (Table 1), a reduction in NDF, FN, and fruit size was observed without affecting extremely important traits such as NCF, YIELD, FF, PF, SST, and PV%. Thus, we can select genotypes with the potential to reduce fruit size without compromising quality and yield.
It is known that the selection differential is not the gain in the selection and that the measures of heritability/genotypic determination coefficients are inherent to the populations studied and the experimental conditions. However, the estimates of genotypic determination coefficients (H2) obtained in Linhares-ES for morpho-agronomic traits evaluated in papaya have shown high values, indicating a great chance of genetic gains in the selection procedures. Silva et al. (2008b) evaluated papaya segregating populations and obtained H2 varying between 53% and 99.75% in several morpho-agronomic traits (plant height, stem diameter, insertion height of the first fruit, number of flowers (total, deformed and sterile flowers), number of commercial and deformed fruits, mean fruit weight, plant yield, fruit firmness and content of soluble solids of fruit pulp). The authors reported values superior than 70% for plant height (79.19), stem diameter (85.36), plant yield (89.89), external fruit firmness (92.72), internal fruit firmness (88.41) and content of soluble solids of fruit pulp (73.68).
Based on the high magnitude of the genotypic determination coefficients reported on literature and the selection differential obtained, there are expectations of obtaining gains with the selected genotypes. Thus, we can select the genotypes with traits of interest and, after the progeny test, evaluate in future works the parents' genetic potential to generate hybrids with the same traits.
The genotypes selected for the JS12-G parent (Table 2) showed a positive selection differential for all traits except FN (-3.6), which points to its reduction and possible gains in other traits, especially FW (211.1 g) and NCF (12.0). Regarding the genotypes selected for the Sekati parent (Table 3), a reduction in terms of FW, FFIH, FN, FL, FD, and PT, as well as increased important traits such as NCF, YIELD, FF, PF, SST, and PV%, were observed. Santa-Catarina et al. (2020b) points out that the FF, PF, and SST traits variability allow the selected lines, whether Formosa or Solo, to present quality and fruit flavor.
Studies on the genetic divergence, that is, differences in allele frequencies of populations, are of fundamental importance in choosing varieties to be used as parents since the genetic distance of parents indicates heterotic expression in the progenies (Falconer 1981). Thus, the results obtained indicate the possibility of selecting genotypes with higher and lower FW, confirming the need for purification of the parent lines targeting the expected standard for the hybrids to be developed from these purified lines, highlighting the demand for FW reduction of 'UC10' and increased values for 'Calimosa'.
Among the selected genotypes for the SS-72/12 (Table 4), the FW trait is worth noting, with 411.6 g higher than the general average (380.9 g). The values of the traits related to quality were also higher than the general average. Only FFIH, NDF, FN showed values below the general average. This result is interesting because the set of selected genotypes presents plants with a lower insertion of the first fruits and has few nodes without fruits and deformed fruits.
According to Santa-Catarina et al. (2020a), associated with the combined selection, direct selection can be used to select genotypes that are potential sources of favorable alleles for specific traits. In this sense, the direct selection of this study, based on the FW, allowed us to maintain eight genotypes in the lines (two genotypes per parent).
Considering the simultaneous selection objective, we found that the CSI (using standardized phenotypic means) was consistent in identifying possible genotypes to form the progeny test. This finding shows the advantage of standardizing observations in the evaluation and selection of superior genotypes. Thus, it is evident that this strategy can allow greater precision in the selection process, thus increasing the chances of success of the breeding programs (Ramos et al. 2014). By associating CSI and SD, 80 genotypes were selected (20 Sekati, 20 JS-12-P smaller fruits, 20 JS-12-G larger fruits, and 20 SS-72/12) to compose the next work stage, which is the evaluation of the 80 progenies in a competition trial.
Given the results, we can show that the maintenance of agronomic traits in hybrid cultivars is extremely important in using pure lines as parents in hybridization tests since allelic variations in loci genes of agronomic importance in parents can change the standard of developed hybrids over time. In this sense, some actions are essential to guarantee the high level of allelic fixation of the parent lines and, consequently, the genetic purity of the hybrid seeds: care in the crossing procedures to avoid contamination; monitoring the inbreeding of parent lines and, if necessary, purifying them from time to time, to avoid such segregation. Ingale and Waghmode (2005) worked with the purification of Sahdyari rice hybrid parental lines and also reported that lack of purity in parental lines and improper isolation conditions in seed production result in poor hybrid seed quality.
It is expected that with the selected genotypes and the continuity of the work, we will be able to achieve purified lines, contributing to the maintenance of the commercial standard of the developed hybrids. This process aims to obtain the ‘UC10' hybrid with a maximum of 2.0 kg fruits and the hybrid 'Calimosa' with fruits from 1.2 to 1.4 kg and more yielding.
Conclusions
There is a possibility to improve a developed hybrid through the purification of parental lines since there is segregation in the JS-12-P, JS12-G, Sekati and SS-72/12 parents that can be explored in the papaya breeding program to obtain 'UC10' and 'Calimosa' hybrids with different fruit weight so that they meet the needs of the markets.
References
Alvares CA, Stape JL, Sentelhas PC, Goncalves JLM, Sparovek G (2013) Koppen’s climate classification map for Brazil. Meteorol Z 22:711–728. https://doi.org/10.1127/0941-2948/2013/0507
AOAC (1990) Official Methods of Analysis, 13th ed. Association of Official Analytical Chemists, Washington, DC, USA
Barragán-Iglesias J, Méndez-Lagunas LL, Rodríguez-Ramírez J (2018) Ripeness indexes and hysicochemical changes of papaya (Carica papaya L.cv. Maradol) during ripening ontree. Sci Hortic 236:272–278. https://doi.org/10.1016/j.scienta.2017.12.012
Barros GB, Aredes FAS, Ramos HCC, Santa-Catarina R, Pereira MG (2017) Combining ability of recombinant lines of papaya from backcrossing for sexual conversion. Rev Ciênc Agron 48:166–174. https://doi.org/10.5935/1806-6690.20170019
Cardoso DL, Da Luz LN, De Macêdo CMP, Gonsalves LSA, Pereira MG (2014) Heterosis in papaya: Inter and intragroup analysis. Rev Bras Frutic 36:610–619. https://doi.org/10.1590/0100-2945-279/13
Cattaneo LF (2001) Avaliação da divergência genética e análise de gerações em mamoeiro (Carica papaya L.). Dissertation, Universidade Estadual do Norte Fluminense Darcy Ribeiro
Cortes DFM, Santa-Catarina R, Barros GBA, Aredes FAS, Silveira SF, Ferreguetti GA, Ramos HCC, Viana AP, Pereira MG (2017) Model-assisted phenotyping by digital images in papaya breeding program. Sci Agric 74:294–302. https://doi.org/10.1590/1678-992x-2016-0134
Cortes DFM, Santa-Catarina R, Vettorazzi JCF, Ramos HCC, Viana AP, Pereira MG (2019) Development of superior lines of papaya from the Formosa group using the pedigree method and REML/Blup procedure. Bragantia 78:1–11. https://doi.org/10.1590/1678-4499.20180253
Falconer DS (1981) Introdução à genética quantitativa. Impr Univ, Viçosa
Gu Z (2014) Circlize implements and enhances circular visualization in R. Bioinformatics 30:2811–2812. https://doi.org/10.1093/bioinformatics/btu393
IBGE (2019) Instituto Brasileiro de Geografia e Estatística. http://www.ibge.gov.br/home/. Accessed on Mar. 31, 2020
Ingale BV, Waghmode BD (2005) Purification of Sahdyari rice hybrid parental lines through paired crosses for nucleus seed production. Int Rice Res Notes 30:18–19
Luz LNDA, Pereira MG, Barros FR, Barros GBA, Ferreguetti GA (2015) Novos híbridos de mamoeiro avaliados nas condições de cultivo tradicional e no semiárido brasileiro. Rev Bras Frutic 37:159–171. https://doi.org/10.1590/0100-2945-069/14
Luz LNDA, Santa-Catarina R, Barros GBA, Barros FR, Vettorazzi JCF, Pereira MG (2018) Adaptability and stability of papaya hybrids affected by production seasonality. Crop Breed Appl Biotechnol 18:357–364. https://doi.org/10.1590/1984-70332018v18n4a54
Ministério da Agricultura Pecuária e Abastecimento (MAPA) (2011) Guia de inspeção de campos para produção de sementes / Ministério da Agricultura Pecuária e Abastecimento. Secretaria de Defesa Agropecuária. – 3. ed. revisada e atualizada – Brasília: Mapa/ACS, 41 p
Pereira MG, Ferreguetti GA, Nair T, et al (2015) Melhoramento Genético do Mamoeiro: Programa UENF/CALIMAN. In: David dos Santos Martins. (Org.). VI Simpósio do Papaya Brasileiro. Vitória, ES: DCM/Incaper 6:01–41
Pereira MG, Luz LN, Santa-Catarina R, Ramos HCC, Pereira TNS, Barros GBA, Ferreguetti GA, Vivas M, Cortes DFM, Vettorazzi JCF, Azevedo AON, Silveira SF, Oliveira JG, Viana AP (2019a) UC10: a new early Formosa papaya cultivar. Crop Breed Appl Biotechnol 19:131–134. https://doi.org/10.1590/1984-70332019v19n1c18
Pereira MG, Luz LN, Santa-Catarina R, Ramos HCC, Pereira TNS, Barros GBA, Ferreguetti GA, Vivas M, Cortes DFM, Vettorazzi JCF, Azevedo AON, Silveira SF, Oliveira JG, Viana AP (2019b) ‘UC14’: a new papaya cultivar with intermediate fruit size. Crop Breed Appl Biotechnol 19:226–229. https://doi.org/10.1590/1984-70332019v19n2c31
Pereira MG, Poltronieri TPS, Pereira TNS, Ramos HCC, Santa-Catarina R, Vettorazzi JCF, Aredes FAZ, Boechat MSB, Venâncio TM, Azevedo AON, Cortes DFM, Moreira NF, Bohry D, Silveira SF, Vivas M, Ferreguetti GA (2019c) Twenty-two-year papaya breeding program: from breeding strategy establishment to cultivar development. Funct Plant Breed J 1:9–27. https://doi.org/10.35418/2526-4117/v1n2a2
Pirovani AAV, Ramos HCC, Duarte RP, Miranda DP, Bohry D, Santana JGS, Santos PHAD, Pereira MG (2021) Molecular characterization of elite lines of papaya (Carica papaya L.) via SSR markers. Funct Plant Breed J 3:49–58. https://doi.org/10.35418/2526-4117/v3n1a5
R Core Team (2018) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Ramos HCC, Pereira MG, Viana AP, Luz LN, Cardoso DL, Ferreguetti GA (2014) Combined selection in backcross population of papaya (Carica papaya L.) by the mixed model methodology. Am J Plant Sci 05:2973–2983. https://doi.org/10.4236/ajps.2014.520314
Rodrigues AS (2021) Caracterização molecular em mamoeiro visando a seleção e purificação de linhagens genitoras de híbridos comerciais. Tese (Mestrado em Genética e Melhoramento de Plantas) - Universidade Estadual do Norte Fluminense Darcy Ribeiro, Centro de Ciências e Tecnologias Agropecuárias, Campos dos Goytacazes, RJ, 70 p
Santa-Catarina R, Cortes DFM, Vettorazzi JCF, Ramos HCC, Ferreguetti GA, Pereira MG (2018) Image-based phenotyping of morpho-agronomic traits in papaya fruits (Carica papaya L. THB var.). Aust J Crop Sci 12(11):1750–1756. https://doi.org/10.21475/ajcs.18.12.11.p1437
Santa-Catarina R, Cortes DFM, Vettorazzi JCF, Poltronieri TPS, Barros GBA, Arêdes FAS, Azevedo AON, Ramos HCC, Pereira MG (2019) Combining ability for fruit yield and quality in papaya recombinant inbred lines from the sexual conversion backcrossing. Euphytica 215:154–164. https://doi.org/10.1007/s10681-019-2485-3
Santa-Catarina R, Pereira MG, Vettorazzi JCF, Cortes DFM, Poltronieri TPS, Azevedo AON, Moreira NF, Miranda DP, Moraes R, Pirovani AAV, Ramos HCC, Vivas M, Viana AP (2020a) Papaya (Carica papaya L.) S1 family recurrent selection: opportunities and selection alternatives from the base population. Sci Hortic 260:108848. https://doi.org/10.1016/j.scienta.2019.108848
Santa-Catarina R, Vettorazzi JCF, Cortes DFM et al (2020b) Phenotypic characterization of recurrent selection S1 Papaya (Carica papaya L.) families by multivariate approach. Euphytica 216:117. https://doi.org/10.1007/s10681-020-02655-1
Silva FF, Pereira MG, Damasceno Júnior PC, Daher RF, Pereira TNS, Filho GAS, Viana AP, Ferreguetti GV (2007) Monitoring of the genetic variability in papaya parent “Formosa” of “UENF/CALIMAN 01” hybrid via RAPD. Crop Breed Appl Biotechnol 7(1):36–42. https://doi.org/10.12702/1984-7033.v07n01a06
Silva FF, Pereira MG, Ramos HCC, Damasceno Júnior PC, Pereira TNS, Gabriel APC, Viana AP, Ferreguetti GA (2008a) Selection and estimation of the genetic gain in segregating generations of papaya (Carica papaya L.). Crop Breed Appl Biotechnol 8:1–8. https://doi.org/10.12702/1984-7033.v08n01a01
Silva FF, Pereira MG, Ramos HCC, Damasceno Júnior PC, Pereira TNS, Viana AP, Daher RF, Ferreguetti GA (2008b) Estimation of genetic parameters related to morpho agronomic and fruit quality traits of papaya. Crop Breed Appl Biotechnol 8:65–73. https://doi.org/10.12702/1984-7033.v08n01a09
Singh D (1981) The relative importance of characters affecting genetic divergence. Indian J Genet e Plant Breed 41:237–245
Vettorazzi, JCF (2020) Linhagens endogâmicas recombinantes: variabilidade genética, avaliação per se, capacidade combinatória e heterose em mamoeiro. Dissertation Universidade Estadual do Norte Fluminense Darcy Ribeiro
Vivas M, Silveira SF, Amaral Júnior AT, Cardoso DL, Pereira MG (2013) Inheritance of papaya resistance to fungal disease based on Hayman’s diallel analysis. Bragantia 72:332–337. https://doi.org/10.1590/brag.2013.047
Vivas M, Silveira SFDA, Pereira MG, Cardoso DL, Ferreguetti GA (2013) Diallel analysis of papaya for resistance to phoma spot. Cienc Rural 43:945–950. https://doi.org/10.1590/S0103-84782013005000054
Acknowledgements
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES) – Finance Code 001. We also thank the Carlos Chagas Filho Foundation for Research Support of Rio de Janeiro (FAPERJ), the Council National Scientific and Technological Development (CNPq) for the scholarship granting and project financing, and Caliman Agrícola S/A for logistical support.
Funding
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) – Finance Code 001. We also thank the Carlos Chagas Filho Foundation for Research Support of Rio de Janeiro (FAPERJ), the Council National Scientific and Technological Development (CNPq) for the scholarship granting and project financing, and Caliman Agrícola S/A for logistical support.
Author information
Authors and Affiliations
Contributions
AAVP: Evaluation the experiments, analysis of the results and wrote the manuscript; HCCR: supervised the project and contributed to the writing of the manuscript; RS-C: contributed to the design and implementation of the research, helped in planting and evaluating the experiment and to the analysis of the results and to the writing of the manuscript; DPM: helped in planting and evaluating the experiment; ASR: helped in planting and evaluating the experiment; JCFV: helped in planting, evaluating the experiment and contributed to the final version of the manuscript; IRSJ: helped in planting and evaluating the experiment; MGP: supervised the project and contributed to the final version of the manuscript; APV: contributed to the final version of the manuscript.
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Pirovani, A.A.V., Ramos, H.C.C., Santa-Catarina, R. et al. Is there a possibility to improve a developed hybrid? A current demand on papaya (Carica papaya L.). Euphytica 218, 64 (2022). https://doi.org/10.1007/s10681-022-03014-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10681-022-03014-y