
Abstract
Wheat breeding has significantly contributed to enhancing yield potential and stability. However, one of the most promising technologies for further sustainable yield increases, hybrid wheat, is still limited due to lack of well-established heterotic groups and insufficient heterosis to compensate cost of hybrid seed production. Thus, optimizing the appropriate parents and combinations of floral and flowering traits is important to enhance outcrossing ability. Phenotyping methods for floral and flowering traits were applied in this study. Wide phenotypic variation was observed for all the traits assessed. High heritability coupled with high genetic advance for anther extrusion, pollen mass, duration of floret opening, and openness of florets were observed. Ideally, visual anther extrusion and pollen shedding appear as promising traits allowing indirect selection of other associated floral traits, such as pollen mass and duration and degree of floret opening. Adopting these two traits in further selection will allow scoring of large numbers of genotypes in a very short time.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Wheat (Triticum aestivum L.) is one of the main staple crops in the world and demand for wheat is almost synonymous with demand for food. Worldwide, wheat is the primary source of calories for millions of people, accounting for around 28% of cereals used as food (FAOSTAT 2014). Globally, wheat is grown on more than 220 million hectares with annual production likely to reach 750 million tons (FAOSTAT 2018). Despite remarkable progress in wheat productivity, greater efforts need to be made to cope with an ever-increasing world population growth that will reach 9.8 billion by 2050. Global wheat production needs to increase by 2% annually to meet escalating demands.
Over the years, wheat breeding has significantly contributed to enhancing yield potential and stability. Comparing the 1961–1965 and 2013–2017 periods, average yield rose from 1.18 to 3.15 t/ha, and wheat production increased from 247 to 741 million tons (FAOSTAT 2019). However, opportunities for further sustained increases are limited due to the limited genetic variability in wheat germplasm, insufficient exploitation of heterosis and the gap between the technology and farm achievement (Venske et al. 2019). To bridge this gap and to make wheat cultivation more attractive, there is a need to explore new technologies with potential to break yield barriers and substantially enhance the level of productivity (Jordaan 1996). In this context, the exploitation of heterosis through hybrid wheat is one possibility that could capture significant yield benefits.
Hybrid wheat has shown superior grain potential for enhanced yield performance and stability compared to pure-line varieties (Mühleisen et al. 2014). Longin et al. (2013) reported average best parent heterosis levels ranging from − 15.7 to 25.7% for grain yield of elite hybrids compared to their respective higher yielding parent (average of 12.45 t ha−1).
Several attempts have been undertaken to establish suitable seed production systems for hybrid wheat. However, these were only partially successful. The constraints included inadequate levels of heterosis, inefficient seed production systems, and lack of defined heterotic groups of germplasm to serve as parents (Zhou et al. 2005). A major hindrance was also the stringent autogamous nature of wheat. Thus, optimization of floral and flowering traits could contribute to increased production of hybrid seed. Several traits such as flowering time, plant height, anther extrusion, pollen mass and degree and duration of floret opening were seen as essential for maximum pollination (De Vries 1971; Whitford et al. 2013). Moreover, the respective male and female parental lines need to possess specific trait combinations. The male ideotype was taller than the female, with long extruded anthers producing large amounts of pollen over an extended time period enabled by differential flowering timing between tillers. The female ideotype was envisaged as shorter with wide opening florets and long hairy stigmas that were receptive for extended periods (De Vries 1972; Longin et al. 2012). Obviously, flowering time has to be synchronized.
Identification of potential parental lines with these features is challenging and phenotyping of most floral and flowering traits is difficult and time consuming. Hence, it is important to use suitable approaches to expedite selection for enhanced outcrossing capacity. The present study was undertaken to evaluate diverse wheat germplasm for appropriate floral and flowering traits and to investigate potential of genomic approaches for use in hybrid wheat programs. In particular, our objectives were to (1) evaluate genetic variability in bread wheat for male and female traits with potential relevance for hybrid wheat breeding programs, (2) assess correlation, heritability and genetic advance for hybrid related traits, and (3) identify time- and cost-effective proxies for good outcrossing ability.
Materials and methods
Plant materials
A panel of 400 genotypes, including mainly elite lines from the ICARDA bread wheat breeding program, 15 commercial checks and 63 synthetics and synthetic-derived lines was assembled to study the genotypic variability in hybrid wheat-related traits (Supplemental Table S1).
Phenotyping
The panel was phenotyped under plastic house conditions at ICARDA Rabat Guich, Morocco (33°97′N, 6°86 W, 57 masl) in the 2015/16 cropping season and a subset of 200 genotypes were selected based on initial phenotypic assessment of potential hybrid traits. This subset was grown under field conditions at Marchouch Station, Morocco (33°36′N, 6°43′W, 394 masl) during the 2016–2017 and 2017–2018 cropping seasons. The genotypes were grown in 3m2 plots in an augmented design. All experiments were managed according to standard cultural practices.
The traits assessed are listed in Table 1:

Phenotypic evaluation of traits: a Pollen mass assessment; b Pollen viability. Viable pollen grains are indicated with arrowheads; c Openness of florets (gapping)
Statistical analyses
Statistical analyses were performed on a yearly basis as well as across years. A linear mixed model using restricted maximum likelihood (REML) method (Patterson and Thompson 1971) was used for data analysis. For yearly analyses, genotypes and checks were considered as fixed whereas blocks were considered as random effects. Best linear unbiased estimates (BLUEs) were calculated for genotypes for all traits specifically to get the estimated means in order to compute the between trait correlations. The same analysis was done using genotypes as random to get the genotypic variance components and hence to compute heritability and the genotypic coefficient of variation. For the across year analysis, years, checks and their interaction were fitted as fixed effects and genotypes and genotype by year interaction were random to get the genotypic and genotype by year interaction variance components. Best linear unbiased predictors (BLUPs) were calculated for genotype by year interactions and used for ranking genotypes and selection of the best genotypes for further experiments.
Pearson correlation coefficients and their corresponding P values were calculated between traits in each year. Repeatability among biological replicates was calculated from the expression σ2g / (σ2g + σ2e/nr). Broad sense heritability of the evaluated traits was calculated from the expression σ2g/(σ2g + σ2g · e/ne + σ2e/ne · nr), where σ2g represented the genetic variance, σ2e the error variance, ne was the number of environments and nr the number of replicates. All statistical analyses were done and models were fitted in ASReml v3.0-1 (Butler et al. 2007) in R v3.3.1 (R Core Team 2016).
Phenotypic and genotypic coefficients of variation were estimated according to the method suggested by Burton and DeVane (1953) as follows:
where\({\upsigma }^{2}p\) was the phenotypic variance; \({\upsigma }^{2}g \mathrm{was the}\) genotypic variance and \({\bar{\hbox{x}}}\) was the sample mean. Genetic advance (GA) and percentage of the mean (GAM) assuming selection intensity at 5% was estimated as per the formulae given by Allard (1960)
where K = standardized selection differential (at 5% selection intensity K = 2.063); and H is broad sense heritability
Genotypic analyses
The panel was genotyped by whole genome scanning using the 15 K Single Nucleotide Polymorphism (SNP) marker array (TraitGenetics Gmbh). SNP raw data were filtered according to marker criteria; minor allele frequency (MAF) > 5% and missing data ≤ 10%. This resulted in 10,477 SNPs for this study. Cluster analysis was performed by the unweighted pair group method with arithmetic mean (UPGMA) method to find discrete groups based on linkage criteria between genotypes.
Results
The results of the mixed model analysis showed large phenotypic and genotypic variations as well as important differences in broad sense heritability for all flowering and floral traits evaluated for the 200-genotype subset (Table 2). Heritabilities ranged from low to high and genotypic variances were significantly different from zero (P < 0.001) for all traits across the three data sets.
Floral traits showed a wide range of variability across years. Genotypes SOMAMA-9/ICARDA-SRRL-2 and QADANFER-11/REBWAH-11 among others showed very good performance for floral traits favoring outcrossing. FLAG-1 was the check having highest values for anther extrusion and anther length but low values for the other floral traits. Among the synthetic and synthetic-derived lines, STY-US/CELTA//PALS/3/SRN_5/4/AE.SQUARROSA (431) was the only genotype with good performance for duration of floret opening, pollen mass and anther extrusion with values of 100 min, 38.42 mg and 79.17%, respectively. Most of the synthetic and synthetic-derived genotypes had long anthers but produced lower pollen mass compared with the elite lines used in this study. For example, synthetics DOY1/AE. SQUARROSA (1026) and SORA/AE. SQUARROSA (208) had large anthers (> 5 mm) but shed small amounts of pollen (< 14 mg).
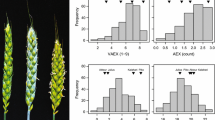
The trait distributions of the BLUES followed approximate normal distributions except for heading date, flowering start and flowering peak (Fig. 2). Absolute values of phenotypic correlations among the 20 traits ranged between − 0.40 and 0.92 (Table 3). There were high correlations among pollen mass, openness of the floret, duration of floret opening, pollen shedding, anther extrusion and visual anther extrusion ranging from 0.65 to 0.90 (P < 0.001). The highest correlation for pollen mass was with anther extrusion (r = 0.90). The correlation between anther extrusion and visually scored anther extrusion was highly significant (r = 0.75, P < 0.001). Spikelet number per spike was near-independent of pollen shedding and pollen mass (r = − 0.18* and r = − 0.15*, respectively) but moderately positively correlated with spike length (r = 0.47, P < 0.001). Anther length was weakly correlated with plant height (r = 0.16).

Histograms of variation in phenotypic values for some evaluated traits in 200 genotypes
Among flowering traits, days to heading was strongly correlated with days to maturity, flowering start, flowering peak and flowering end with the highest value for flowering start and flowering peak (r = 0.92).
The across year analysis showed high heritability of important floral traits in Table 4 except for anther length. The highest heritability (0.93) was observed for anther extrusion. However, anther length and spike length showed low to medium heritability across years. The percentage of estimated genotypic variances and variances due to genotype-by-year interaction were significant for most of the floral and flowering traits except for spike compactness (Fig. 3). Genetic variation dominated the total variation for anther extrusion (59.94%), pollen mass (59.09%), duration of floret opening (57.01%) and floret openness (61.64%). However, genotype-by-year variation had the greatest effect on anther (62.56%) and stigma length (46.81%). The genotypic variance for agronomic traits accounted for a maximum 5% of the total variation.

Partition of total variance for some investigated traits. See Table 1 for trait abbreviations. Ve: error variance, Vg: genotypic variance, Vg/y: genotype by year variance
The phenotypic and genotypic coefficients of variability ranged from 4.01 and 44.24 and from 1.46 and 41.92, respectively. The highest coefficient of variability for both phenotypic and genotypic levels was recorded for anther extrusion. Highest values were also found for pollen mass, duration and degree of floret opening (Table 4). Similarly, the highest genetic advance was for anther extrusion (GA = 38.09). Genetic advance estimated at 5% selection intensity was high for anther extrusion, pollen mass, duration and degree of floret opening.
Discussion
Although hybrids are well established for allogamous crops such as maize, hybrid wheat has not been successful due to relatively low levels of hybrid vigor and lack of an efficient hybrid seed system. Floral biology will play an important role in increasing out-crossing. Several studies have described the floral and flowering biology of wheat. Although male and female ideotypes for efficient hybrid seed production are well defined, slow progress has been made in key trait improvement. In this study, we applied several approaches to evaluate floral and flowering traits and discuss their contribution in improving pollination capacity.
Variation in floral and flowering traits
A high outcrossing ability is most likely influenced by a combination of floral traits such as anther extrusion, pollen shedding, anther size, openness of the floret, and stigma length (Singh et al. 2010). The approaches suggested by Langer et al. (2014) to assess pollen mass and anther extrusion are generally used as proxies to determine the amount of pollen released and percentage of extruded anthers. Anther extrusion can be visually and repetitively assessed under optimum weather conditions, but assessments could be disrupted by wind or rain. In the present study, anther extrusion assessment was carried out using counted and visual scoring methods and the results showed that both were successful in identifying highly extruding genotypes. Given the high correlation between anther extrusion and visual anther extrusion, visual anther extrusion is a promising method for initial selection of male parents for hybrid seed production.
It is not surprising that anther extrusion depends on opening florets and the duration of opening. A line showing these traits should serve as a potential pollen donor as well as a more receptive female parent. As in the present study, previous research showed that a high percentage of anther extrusion was directly related to high numbers of open florets (Rajki 1960, 1962; Singh and Joshi 2003). In addition to anther extrusion, visual anther extrusion also showed a close relationship with floret openness and duration of floret opening.
Another important determinant for outcrossing is the quantity of shed pollen. The wide variation in pollen shedding in this study was in concordance with previous studies. D’Souza (1970) reported differences of 5–80% between genotypes for the amount of pollen shed. However, such differences will dependent on traits like anther extrusion, openness of the floret and duration of floret opening. Generally, there was profuse pollen shedding outside the florets when there was adequate floret widening and high anther extrusion. Our results were in agreement with previous studies (De Vries 1974a, b; Singh et al. 2007; Langer et al. 2014).
Pollen viability is also an important factor that enhances cross-pollination. The longevity of pollen grains is less than 3 h (Fritz and Lukaszewski 1989). Regardless of low viability values, most lines showed high variability. The low values might be due to moisture loss during transportation from the field to the laboratory.
Anther length, used as proxy for anther size, is also a key spike trait that contribute to enhance outcrossing in wheat. We found ranges of 2.52–4.95 mm over the three years. Song et al. (2018) showed anther length to be 2.7–6 mm in a set of 305 lines. Anther size was associated with pollen grain number in wheat/rye addition lines (Athwal and Kimber 1970; De Vries 1974a; Nguyen et al. 2015). Other studies showed that anther size per se had no impact on the amount of pollen shed outside the floret (Beri and Anand 1971). We also found no correlation between the anther length and pollen shedding or any other trait except for stigma length and plant height. The lack of correlation between the two characters suggests that genes controlling anther length differ from those controlling the other floral traits.
Open florets in hybrid seed production is crucial for both male and female parents. Lines with wide floret opening for an extended duration should be prone to greater outcrossing. Few studies have addressed female traits due to their intensive work and time requirements. In this study, we observed a wide range of variability in the degree of floret opening (9.05–45.6° as separation angles) and duration (8.61–109 min).
Selection for stigma size was shown to have positive effects for cross-pollination and seed set (Virmani and Edwards 1983; Blouet et al. 1999). Long fully extruded and receptive stigmas for extended periods should be advantageous. Wide ranges in stigma length have been published (2.13–5.2 mm) (Komaki and Tsunewaki 1981; Fábián et al. 2019) as well as 1.78–5.05 mm being recorded in this study.
An optimized difference in plant height and synchronized flowering between the male and female parents are prerequisite for an efficient hybrid wheat seed production system. Non-uniformity of flowering causes low seed set. The pollen shed by taller male parents can disperse longer distances and gradually fall to reach the shorter female parents. The flowering period is expected to be determined by genotype and weather conditions. For hybrid production, a prolonged flowering time should be advantageous.
Phenotyping specific traits for hybrid production is hard and time-consuming as floral and flowering traits are strongly dependent to environmental conditions. High temperatures accelerate the flowering process and shorten the flowering period, narrowing the time window for pollination.
Genetic advance
Improvement of a given trait is through the available genetic variation and its heritability. Estimated broad-sense heritability alone is not very helpful because it includes the effect of both additive and non-additive genetic variation. Prediction of genetic advance is therefore useful to indicate the expected genetic progress from selection for a particular trait and helps to determine gene action affecting that trait (Sravan et al. 2012). Hazel and Lush (1942) defined the genetic advance as the improvement in average phenotypic or genetic value due to selection within a population in each breeding cycles. Genetic advance as percent of the mean was classified as low for values from 0 to 10%, 10–20% for moderate and 20% and above for high values (Johnson et al. 1955). Since high heritability does not necessarly indicate high genetic gain, heritability coupled with genetic advance is a more reliable guide for selection of superior genotypes at early generation (Akinwale et al. 2011).
The genetic advances as percentages of means ranged from 4.58 to 88.20% for floral traits. Predicted genetic advances in anther extrusion, duration of floret opening, pollen mass, and floret openess were high at 88.20%, 65.06%, 64.83% and 45.20%, respectively. Moderate GAM was estimated for stigma length (13.51%) while that for anther length was low (4.58%). Singh (2006) predicted high genetic advances as percentage of mean values for the same floral traits. The agronomic traits underwent moderate genetic advances when compared to their mean values.
Traits like anther extrusion, pollen mass, openness of the floret and duration of floret opening showed high GAM coupled with high heritability estimates. This indicates the preponderance of additive gene action in determination of these traits and that a large proportion of the phenotypic variance was genotypic and effective selection could be made for such traits based on phenotypic expression. However, high heritability along with low to moderate genetic advance as percent of mean, i.e. spike compactness, was indicative of non-additive gene action and that selection might be less successful. Low heritability and predicted genetic advance for anther length was observed due to non-additive gene action and high environmental influence as reported by Akinwale et al. (2011).
The estimates of phenotypic variance were only slightly higher than genotypic variance for some floral traits indicating a low influence of environmental factors. Thus, selection based on phenotype could be successful for these traits. Anther extrusion, duration of floret opening, pollen mass, and floret openness showing high genotypic and phenotypic coefficients of variation along with high heritability and predicted genetic advance may be considered important traits in breeding for high hybrid seed production.
Associations among traits
The results of selection for hybrid parent attributes depend on the heritability and genetic variation of floral and flowering traits. Positive associations between traits could simplify parental selection. Most floral and flowering traits show positive correlations. Anther extrusion, a key trait for improving pollen dispersal, was highly correlated with released pollen mass and pollen shedding. Similar associations were reported by Langer et al. (2014). Given the high correlation between pollen shedding and both pollen mas and anther extrusion, pollen shedding is an easily scored trait that could be taken as proxy for selection based on the respective correlated traits. Several studies showed that genotypes with high anther extrusion are more likely to have resistance to Fusarium head blight (FHB) (Buerstmayr and Buerstmayr 2015; Xu et al. 2019). Therefore, it will also by interesting to check correlations between FHB and other evaluated floral traits.
Whether longer anthers influence pollen mass remains unclear. We assessed anther length, but no significant correlation was found. However, Langer et al. (2014) reported that anther size was positively correlated with pollen mass and anther extrusion. Boeven et al. (2018) reported that both counted and visual anther extrusion as well as filament length were correlated with hybrid seed set. In other hand, AL was found to be weakly associated with SL (r = 0.29) in the present study.
An extended period of open flowers should favor outcrossing. The degree and the duration of floret opening were highly associated (r = 0.85) over two years. OPF and DFO were both correlated with other traits including pollen mass, pollen shedding, and visual and counted anther extrusion. Singh et al. (2007) reported weak but significant associations between OPF and DFO (r = 0.30) and a high correlation between DFO and AEX (r = 0.89).
Langer et al. (2014) reported a moderate correlation between anther extrusion and plant height and therefore suggested that the correlation between these traits was worthy of further investigation. Boeven et al. (2016) later reported that reduced height genes had a negative effect on male floral traits. However, our results did not confirm this effect as we found only a weak association between anther length and plant height.
Floral traits in relation to flowering traits showed only weak negative correlations between flowering duration and pollen shedding and visual anther extrusion. Langer et al. (2014) reported a high negative correlation between AEX and DH. It would be interesting to determine whether the early flowering is associated with greater anther extrusion and pollen mass in further studies.
AEX previously showed a moderately negative correlation (r = − 0.48) with numbers of spikelets per spike (Langer et al. 2014). Other weak negative correlations were between spikelets per spike and both pollen mass and pollen shedding. But no associations between tillers per plant, spike length, spike compactness and the floral traits were measured in this study.
The floret structure within spikes was reported to influence flowering type characteristics with tighter spikelets tending to have narrower separation angles (Obermayer 1916). However, no significant relationship was observed between the spike compactness and floret gap in this study.
Clustering of germplasm
We made a cluster analysis of panel members based on phenotypic values (Fig. 4). We identified genotypes that combined high pollen mass and anther extrusion as promising traits for male parents and openness of the florets and duration of floret opening for female parents. This allowed us to confirm that most genotypes exhibiting high male potential carried features of good females as well. The selected genotypes were already used in hybrid wheat trials to study the combining ability and level of heterosis of hybrids using a chemical hybridizing agent to sterilize the female lines.

Prediction of a best female candidates with high rates of duration of floral opening and openness of the florets; and b best male candidates with high rates of anther extrusion and pollen mass
We also tried to determine the molecular relatedness of the characterized genotypes in relation to observed phenotypic values. From the cluster plot (Fig. 5) eight main clusters were identified at branch level 5. Except for QAFZAH-21/ICARDA-SRRL-9, lines derived from crosses involving QAFZAH and QIMMA in the first cluster showed a weak performance in floral traits. Six different lines derived from the cross QADANFER-11/REBWAH-11 grouped in the second cluster had high potential for both male and female parent traits. QAMAR-6, in the third cluster, was the best check having high values of anther extrusion (83.6%), pollen mass (38.6 mg) duration of floral opening (89.3 min), and floret openness (39°). The fifth and sixth clusters included the most diverse genotypes in terms of potential parents. SOMAMA-9/ICARDA-SRRL-2 and FLAG-1 showed high potential as male/female for hybrid seed production. Some genotypes were closely related but differed in hybrid parent potential. Hence pedigree was not an acceptable guide for prediction of potential hybrid parents. However, we could identify different groups of genotypes with favorable trait combinations for both male and female parents with high rates of extruded anthers, likely to release more pollen and with florets that remained open for extended time periods. The identification of these groups of genotypes appears interesting and can be classified as one heterotic pattern. It will therefore be interesting to investigate the heterotic potential among the selected genotypes to demonstrate high levels of heterosis. The identification of heterotic loci and exploring their expression levels will help to map stable QTLs and simplify the hybrid wheat breeding process. When the heterotic loci and genes are clearly known, it will be possible to characterize the genotypes of the key loci across the collection of parental lines and predict the potential hybrid combinations with strong heterosis performances. Genome-wide association mapping (GWAS) can be also an alternative to detect genomic regions harboring genes/markers for hybrid traits and thereby substantially reduce the time for selection.

Cluster analysis of genetic relationships among 200 genotypes and heat plots for evaluated traits. *, Genotypes with superior floral trait combinations. Trait abbreviations are provided in Table 1
Height indicates
Conclusions
The self-pollinating nature of wheat makes hybrid wheat development a major challenge. Considerable progress has been made in defining phenotyping approaches in order to identify parents for hybrid breeding. Visual anther extrusion and profuse pollen shedding are promising traits to predict associated target floral traits. While the genetic architecture of male floral traits has been intensively studied, female traits are far less dissected. We therefore recommend shorter lines with prolonged opening of florets and stigma receptivity as potential female parents. The superior potential male and female parents identified in this study must be converted into cytoplasmic male sterile and/or restorer parents for hybrid seed production. The phenotypic variability within the present screened panel is adequate and allowed us the identification of best performing male and female genotypes tested for cross-pollination in hybrid wheat crossing blocks.
Data Availability
Can be shared.
References
Akinwale MG, Gregorio G, Nwilene F et al (2011) Heritability and correlation coefficient analysis for yield and its components in rice (Oryza sativa L.). Afr J Plant Sci 5:207–212
Allard RW (1960) Principles of plant breeding. Wiley, New York, USA
Athwal RS, Kimber G (1970) Anther size and pollen longevity in wheat/rye addition lines. Wheat Inf Serv 30:30–32
Beri SM, Anand SC (1971) Factors affecting pollen shedding capacity in wheat. Euphytica 20:327–332. https://doi.org/10.1007/BF00056096
Blouet A, Streiff K, Guckert A (1999) Possibilities for hybrid seed production in wheat. In: Basra AS (ed) Heterosis and hybrid seed production in agronomic crops, Softcover edn. CBS Publishers & Distributors, New Delhi, pp 81–117
Boeven PHG, Longin CFH, Leiser WL et al (2016) Genetic architecture of male floral traits required for hybrid wheat breeding. Theor Appl Genet 129:2343–2357. https://doi.org/10.1007/s00122-016-2771-6
Boeven PHG, Würschum T, Rudloff J et al (2018) Hybrid seed set in wheat is a complex trait but can be improved indirectly by selection for male floral traits. Euphytica 214:110. https://doi.org/10.1007/s10681-018-2188-1
Buerstmayr M, Buerstmayr H (2015) Comparative mapping of quantitative trait loci for Fusarium head blight resistance and anther retention in the winter wheat population Capo × Arina. Theor Appl Genet 128:1519–1530. https://doi.org/10.1007/s00122-015-2527-8
Burton GW, DeVane EH (1953) Estimating Heritability in tall fescue (Festuca Arundinacea) from replicated clonal material 1. Agron J 45:478. https://doi.org/10.2134/agronj1953.00021962004500100005x
Butler DG, Cullis BR, Gilmour AR, Gogel BJ (2007) Analysis of mixed models for S–language Environments: ASReml-R Reference Manual. https://vsn.klever.co.uk/downloads/asreml/release3/asreml-R.pdf. Accessed 18 Dec 2019
D’Souza L (1970) Studies on the suitability of wheat as pollen donor for cross pollination, compared with rye, triticale and secalotricum. Zeitschrift fur Pflanzenzuchtung 63:246–269
De Vries AP (1971) Flowering biology of wheat, particularly in view of hybrid seed production—A review. Euphytica 20:152–170. https://doi.org/10.1007/BF00056076
De Vries AP (1972) Some aspects of cross-pollination in wheat (Triticum aestivum L.) 1. Pollen concentration in the field as influenced by variety, diurnal pattern, weather conditions and level as compared to the height of the pollen donor. Euphytica 21:185–203. https://doi.org/10.1007/BF00036759
De Vries AP (1974a) Some aspects of cross-pollination in wheat (Triticum aestivum L.). 3. Anther length and number of pollen grains per anther. Euphytica 23:11–19. https://doi.org/10.1007/BF00032735
De Vries AP (1974b) Some aspects of cross-pollination in wheat (Triticum aestivum L.). 4. Seed set on male sterile plants as influenced by distance from the pollen source, pollinator: Male sterile ratio and width of the male sterile strip. Euphytica 23:601–622. https://doi.org/10.1007/BF00022483
Fábián A, Sáfrán E, Szabó-Eitel G et al (2019) Stigma functionality and fertility are reduced by heat and drought co-stress in wheat. Front Plant Sci 10:244. https://doi.org/10.3389/fpls.2019.00244
FAO (2014) FAOSTAT database. Food and Agriculture Organization of the United Nations, Rome, Italy
FAO (2018) FAOSTAT database. Food and Agriculture Organization of the United Nations, Rome, Italy
FAO (2019) FAOSTAT database. Food and Agriculture Organization of the United Nations, Rome, Italy
Fritz SE, Lukaszewski AJ (1989) Pollen longevity in wheat, rye and triticale. Plant Breed 102:31–34. https://doi.org/10.1111/j.1439-0523.1989.tb00311.x
Hazel LN, Lush JL (1942) The efficiency of three methods of selection. J Hered 33:393–399. https://doi.org/10.1093/oxfordjournals.jhered.a105102
Johnson HW, Robinson HF, Comstock RE (1955) Estimates of genetic and environmental variability in soybeans1. Agron J 47:314. https://doi.org/10.2134/agronj1955.00021962004700070009x
Joppa LR, McNeal FH, Welsh JR (1966) Pollen and anther development in cytoplasmic male sterile wheat (Triticum aestivum L.). Crop Sci 6:296–297. https://doi.org/10.2135/cropsci1966.0011183x000600030026x
Komaki MK, Tsunewaki K (1981) Genetical studies on the difference of anther length among common wheat cultivars. Euphytica 30:45–53. https://doi.org/10.1007/BF00033658
Langer SM, Longin CFH, Würschum T (2014) Phenotypic evaluation of floral and flowering traits with relevance for hybrid breeding in wheat (Triticum aestivum L.). Plant Breed 133:433–441. https://doi.org/10.1111/pbr.12192
Longin CFH, Gowda M, Mühleisen J et al (2013) Hybrid wheat: Quantitative genetic parameters and consequences for the design of breeding programs. Theor Appl Genet 126:2791–2801. https://doi.org/10.1007/s00122-013-2172-z
Longin CFH, Mühleisen J, Maurer HP et al (2012) Hybrid breeding in autogamous cereals. Theor Appl Genet 125:1087–1096
Mühleisen J, Piepho HP, Maurer HP et al (2014) Yield stability of hybrids versus lines in wheat, barley, and triticale. Theor Appl Genet 127:309–316. https://doi.org/10.1007/s00122-013-2219-1
Nguyen V, Fleury D, Timmins A et al (2015) Addition of rye chromosome 4R to wheat increases anther length and pollen grain number. Theor Appl Genet 128:953–964. https://doi.org/10.1007/s00122-015-2482-4
Obermayer E (1916) Untersuchungen über das Blühen und die Befruchtung von Winterroggen und Winterweizen. Z PflZücht 4:347–403
Patterson HD, Thompson R (1971) Recovery of inter-block information when block sizes are unequal. Biometrika 58:545. https://doi.org/10.2307/2334389
R Core Team (2016) R: a language and envirenment for statistical computing. https://www.R-project.org/. Accessed 12 Dece 2019
Rajki E (1960) Open and closed flowering in some wheat varieties used in breeding. Növénytermelés 9:309–320. https://doi.org/10.1556/novenyterm.60.2011.suppl.0
Rajki E (1962) Some problems of the biology of flowering in wheat. Proc Symp Genet and Wheat Breed, Martonvásár, Hungary, pp 41–62
Jordaan JP (1996) Hybrid wheat: advances and challenges. In: Reynolds MP, Rajaram S, McNab A (eds) Increasing yield potential in wheat: breaking the barriers. CIMMYT, Mexico DF, Mexico, pp 66–74
Singh S, Joshi A (2003) Variability and character association for various floral characters in wheat [Triticum aestivum (L.) em Thell.]. Indian J Genet Plant Breed 63:153–154
Singh SK (2006) Evaluation of spring wheat [Triticum aestivum (L.) em Thell] germplasm for various floral characteristics. SAARC J of Agric 4:167–177
Singh SK, Arun B, Joshi AK (2007) Comparative evaluation of exotic and adapted germplasm of spring wheat for floral characteristics in the Indo-Gangetic Plains of northern India. Plant Breed 126:559–564. https://doi.org/10.1111/j.1439-0523.2007.01396.x
Singh SK, Chatrath R, Mishra B (2010) Perspective of hybrid wheat research: A review. Indian J Agric Sci 80:1013–1027
Song X, Feng J, Cui Z et al (2018) Genome-wide association study for anther length in some elite bread wheat germplasm. Czech J Genet Plant Breed 54:109–114. https://doi.org/10.17221/70/2017-CJGPB
Sravan T, Rangare NR, Suresh BG, Ramesh S (2012) Genetic variability and character association in rainfed upland rice (Oryza sativa L.). J Rice Res 5:24–29
Venske E, Dos Santos RS, Busanello C et al (2019) Bread wheat: a role model for plant domestication and breeding. Hereditas 156:16. https://doi.org/10.1186/s41065-019-0093-9
Virmani SS, Edwards IB (1983) Current status and future prospects for breeding hybrid rice and wheat. Adv Agron 36:145–214
Whitford R, Fleury D, Reif JC et al (2013) Hybrid breeding in wheat: Technologies to improve hybrid wheat seed production. J Exp Bot 64:5411–5428. https://doi.org/10.1093/jxb/ert333
Xu K, He X, Dreisigacker S et al (2019) Anther extrusion and its association with Fusarium head blight in CIMMYT wheat germplasm. Agronomy 10:47. https://doi.org/10.3390/agronomy10010047
Zadoks JC, Chang TT, Konzak CF (1974) A decimal code for the growth stages of cereals. Weed Res 14:415–421. https://doi.org/10.1111/j.1365-3180.1974.tb01084.x
Zhou W, Kolb FL, Domier LL, Wang S (2005) SSR markers associated with fertility restoration genes against Triticum timopheevii cytoplasm in Triticum aestivum. Euphytica 141:33–40. https://doi.org/10.1007/s10681-005-5067-5
Acknowledgements
This research was financially supported by the International Center for Agricultural Research in Dry Areas. We thank the bread wheat program technicians for their outstanding work in the field at ICARDA-Marchouch Research Station, Morocco.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare that they have no conflict of interest.
Ethical Approval
The authors declare that the experiments comply with the current laws of Morocco.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
El Hanafi, S., Bendaou, N., Kehel, Z. et al. Phenotypic evaluation of elite spring bread wheat genotypes for hybrid potential traits. Euphytica 216, 168 (2020). https://doi.org/10.1007/s10681-020-02695-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10681-020-02695-7