
Abstract
With the exception of Cajanus cajan (L.) Millspaugh (pigeonpea), the remaining species of genus Cajanus have not been domesticated. For millennia these taxa have persisted in natural habitats through self-sown seeds. These wild species are an asset for sustaining future pigeonpea breeding programmes since they contain certain traits (genes) that are necessary for encountering various breeding challenges related to crop improvement and adaptation. In this review we identify the key traits from wild Cajanus species, and discuss various physical and genetic constraints encountered in their utilization in introgression breeding. Some noteworthy achievements recorded from inter-specific breeding programmes in pigeonpea are also discussed. These include the development of (1) high protein (> 28%) genotypes (2) cytoplasmic nuclear male sterility systems (3) highly (> 95%) self-pollinating genotypes, and (4) resistance sources to sterility mosaic disease, nematodes, salinity, photo-insensitivity, pod borers, podfly, bruchids, and Phytophthora blight. To help pigeonpea breeders engaged in inter-specific breeding programmes, we suggest the division of the secondary gene pool germplasm into two sub-group/tiers on the basis of ease in hybridization.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
All the cultivated crop species have evolved from their respective wild ancestors through selection. The natural evolutionary processes such as mutation, recombination, segregation, cross-pollination, natural selection etc. have contributed to patterns of speciation and ecological diversification in the wild relatives of crops over timeframes of millennia. As expected the wild species of genus Cajanus are highly diverse and harbour some useful traits that are not found in the cultivated species Cajanus cajan (L.) Millspaugh popularly called as pigeonpea. It is a highly valued protein-rich food legume, grown on about 7 million hectares in the tropics and sub-tropics. Pigeonpea scientists believe that the productivity of the crop has reached its plateau at around 700 kg/ha (Saxena 2015) and any breakthrough in productivity would be possible only by enriching the genetic diversity through the introgression of genetic materials from wild species. In this review we have identified useful wild Cajanus species and various physical and genetic constraints encountered in inter-specific gene transfers. Besides this, breeding strategies using modern genomics approaches for the genetic enhancement of pigeonpea productivity and stability using wild Cajanus germplasm are also discussed.
The wild versus domesticated Cajanus
The wild Cajanus germplasm
The value of genetic resources in crop improvement has been well recognized and documented by scores of researchers. It is a reservoir of both exploited and unexploited genes which provides ample opportunities to breeders to use them in crop improvement programmes. Initially, the taxonomists established 13 genera within sub tribe Cajaninae, and among these the only cultivated genus was Cajanus and genus Atylosia was placed closest to Cajanus. Subsequently, van der Maesen (1986) sunk Atylosia within genus Cajanus and thus at present, only 12 genera are recognized in sub-tribe Cajaninae. According to van der Maesen (1986, 1990) the genus Cajanus now contains a total of 32 species. Of these, 13 are endemic to Australia, eight to Indian sub-continent, one to Africa, and the remaining across more than one country. At present, the ICRISAT Global Gene Bank maintains 555 accessions of wild relatives of pigeonpea which represents six genera and 57 species (Upadhayaya et al. 2007).
Considering the vastness and potential of germplasm in variety development and crop science, Harlan and de Wet (1971) suggested a systematic means of classifying the entire germplasm into three broad groups and designated them as “gene pools”. Primarily this classification was based on crossability of a given species with cultivated types. Accordingly, the ‘primary gene pool’ involved the cultivated germplasm that is easily crossed genotypes to produce fertile progeny. The ‘secondary gene pool’ contained wild germplasm which, on hybridization, produce hybrids with variable degree of male/female sterility due to various chromosomal aberrations or other incompatibilities. All of the non-crossable wild relatives of a species were placed in a ‘tertiary gene pool’.
Smartt (1990), while reviewing the Cajanus germplasm, proposed a slight modification in the gene pool concept of Harlan and de Wet (1971), suggesting a two-tiered grouping within primary gene pool based on ease of hybridization. He proposed that C. cajanifolius, the progenitor of cultivated pigeonpea with high crossability, should also be included in the primary gene pool, but in a separate (second order) tier. van der Maesen (1986), while revising the genera reported that “strophiole”, a raised growth on hilum (Fig. 1), is a characteristic trait of all the Cajanus wild species including C. cajanifolius, and this structure is absent in the entire cultivated C. cajan germplasm. He further elaborated that the raised strophiole separates the wild species as a group, from the cultivated types. Based on the above observations, the authors hereby propose that instead of primary gene pool the additional tier/order should be created within the secondary gene pool. This suggestion is based on the fact that there are vast differences among secondary gene pool species for their abilities to sexually hybridize with the cultivated type.

Charachteristic prominent strophiole on the seeds of a wild species (left) and its absence in cultivated type (right)
Reddy (1990), Dundas (1990), and Pundir and Singh (1985) observed that in some wild species crosses with the cultivated types are made with ease (20–35% success) while in others considerable difficulties were experienced in the hybridization (< 5% success). Hence on the basis of these arguments, it is proposed that within the secondary gene pool, species such as C. albicans, C. cajanifolius, C. sericeus, C. lineatus, and C. scarabaeoides be placed in the upper tier for their greater crossability; on the other hand, C. acutifolius, C. lanceolatus, C. reticulates, C. laticepalus, and C. trinervius should form the lower tier. Strong evidence in the favour of this argument comes from the fact that the wild species housed in the upper tier of the secondary gene pool also get cross-pollinated (Fig. 2) by insects under natural conditions (Saxena et al. 2016). However, at this stage, we believe that the more credible species allocation in the two tiers should be done only after studying the multiple data sets on the hybridization success rates across diverse environments and whole genome characterization in a well-planned study.

Photo source: ICRISAT
Three wild species (left) and their natural hybrids (right)
Center of origin
For a long time two distinct views persisted related to the centre of origin of pigeonpea; one favoured Africa and the other India. The theory of African origin of pigeonpea was based on the exclusive distribution of Atylosia (now Cajanus) kerstingii Haines, a non-crossable wild relative of pigeonpea. The alternate theory of Indian origin of this crop was constructed on the basis of vast genetic variability found among pigeonpea landraces and the presence of a range of its crossable and non-crossable wild relatives in the country (for details see van der Maesen 1990).
Archaeological evidences
The discovery of pigeonpea seeds in Egyptian tombs of the XII Dynasty (2200–2400 BC) at Dra Abu Negga (Zeven and Zhukossky 1975) supported the African origin of pigeonpea. Similarly, the findings of pigeonpea seeds in the excavations of second century BC to the third century in peninsular India (Kajale 1974) supported its Indian origin (van der Maesen 1995).
Molecular evidences
Advancements made in genomics during last decade have resolved the puzzle of geographical origin of pigeonpea. Initially, single nucleotide polymorphisms (SNPs) were assayed on pre-breeding material, landraces, as well as on wild species accessions of pigeonpea (Kassa et al. 2012; Saxena et al. 2014). Based on detailed phylogenetic, admixture, heterozygosity and molecular variance analyses in pair-wise manner at different scales representing continent, country and province, both the SNP based studies concluded India as the center of origin of pigeonpea (Kassa et al. 2012; Saxena et al. 2014). Further, a detailed study based on pairwise FST values calculated from whole genome re-sequencing (WGRS) data on 166 landraces and 7 accessions from three wild species, Cajanus cajanifolius, Cajanus scarabaeoides and Cajanus platycarpus have underpinned the center of origin of pigeonpea at province level (Madhya Pradesh in India) (Varshney et al. 2017).
Dispersal routes
The theory of Indian origin of pigeonpea is now well established and accepted worldwide. Varshney et al. (2017) also postulated that the pigeonpea crop would have travelled from India to Africa and Madagascar at least two millennia BC; to the new world along with the slave trade in post-Columbian time; and to Indo-China and Australia via the Malay Archipelago. The recent evidence generated from WGRS of pigeonpea landraces and their wild relatives from different geographic points (Varshney et al. 2017) also confirmed the hypotheses that pigeonpea migrated from India to sub-Saharan Africa and finally to South America and Meso-America.
Progenitor of Cajanus cajan/domestication
Genetic and morphological evidences
De (1974) opined that C. cajanifolius (Haines) van der Maesen, a wild species of secondary gene pool, is the critical link between the wild and domesticated species of genus Cajanus. This wild relative of pigeonpea is endemic to hilly forests of the eastern coast and central plateau of India. Dr. L. J. G. van der Maesen, ICRISAT’s former Principal Germplasm Botanist, collected a few live samples of this species which are still being maintained at the Institute’s gene bank. Based on various morphological, biochemical, and cytological evidence Pundir and Singh (1985) concluded that C. cajanifolius is the most probable progenitor of the cultivated pigeonpea. The karyotypic studies conducted by Ohri and Singh (2002) also supported this view. In general, the phenology including plant type, branching pattern, pod bearing habit, and most pod and seed size characters of C. cajanifolius match closely with that of traditional landraces of pigeonpea (van der Maesen 1980). The physical resemblance between these two species is so great that the local tribes consume it like pigeonpea and they call it as “Ban Arhar” meaning “wild pigeonpea”. De (1974) attributed the evolution of cultivated type from its C. cajanifolius to a single gene mutation. However, in a recent study, the two species were found to be separated by at least five genes (Mallikarjuna et al. 2012).
Molecular evidences
The theory of evolution of cultivated pigeonpea from C. cajanifolius was further strengthened when strong molecular evidence of genetic similarity between C. cajanifolius and C. cajan started emerging. Parsimony analysis, using restriction fragment length polymorphism (RFLP) and simple sequence repeat (SSR) markers revealed close genetic association between the two species (Nadimpalli et al. 1993; Ratnaparkhe et al. 1995) and they placed C. cajanifolius as the closest wild species to the cultivated type. This view was further supported by recent genomic studies which used SNP markers (Kassa et al. 2012; Saxena et al. 2014) and WGRS data (Varshney et al. 2017). The WGRS data generated from 292 Cajanus lines from the reference set, including breeding lines, landraces, and wild species accessions have identified the regions of pigeonpea genome that have undergone selective sweeps during domestication and modern breeding. While comparing WGRS data of wild species accessions with landraces and landraces with breeding lines, a total of 2945 and 1323 genomic regions, respectively, were identified with reduced diversity. Moreover, these genomic regions also had 69 structural variations (68 copy number variations and 1 presence and absence variation) detected as targets of domestication. Additionally, genomic regions with reduced diversities were also provided information on a number of genes went under the domestication process (Varshney et al. 2017). One such gene (C. cajan_22378), a homolog of EARLY FLOWERING3 (ELF3) gene, showed two different haplotypes in cultivated and wild species accession at five nucleotide positions. ELF3 gene homologs have been shown to be strongly associated with regulating photoperiod-dependent flowering and normal circadian regulation in plants (Weller and Ortega 2015). Another identified gene is SHATTERING1, which controls pod shattering. These genes/mutations are considered as ‘kick-off’ points for trait diversification in cultivated pigeonpea.
The major conclusions drawn from various studies related to the origin and domestication of pigeonpea are (1) the cultivated pigeonpea evolved through mutation in a wild Cajanus species (C. cajanifolius) about 3500–4000 years ago, (2) India is the centre of origin of pigeonpea, (3) Africa is a secondary centre of diversification, (4) Australia is an important centre of diversity of the genus Cajanus, and (5) the pigeonpea travelled from India to the east and west through trade and migration.
The economic drivers in wild Cajanus
Speciation and natural selection have crafted the diverse traits found across wild Cajanus, several of which are potentially important for cultivated pigeonpea. The wild types continued to grow in natural habitats while the cultivation of productive landraces and the targeted cultivar breeding programmes focussing on enhancing yield, adaptation, and quality traits resulted in reduced genetic diversity. With the changes in various climatic parameters and dynamics of crop damaging insects and diseases, the breeding of high yielding cultivars with stable productivity is becoming difficult. Further, despite intensive searching of the primary gene pool over the last few decades, limited resistance against key stresses has been uncovered. Therefore, plant breeders are now looking beyond primary gene pool to mine useful genes from wild relatives; screening of this genetic wealth has already yielded a few good resources materials as briefly described here.
Resistance to insects
Pigeonpea regularly suffers heavy losses due to insects such as pod borers (Helicoverpa armigera Hubner and Maruca testulalis Geyers), pod flies (Melanagromyza obtusa Molloch), and bruchids (Callosobruchus chinensis). Among the pod damaging insects, H. armigera is prevalent throughout the tropics and subtropics; and the annual estimated losses to pigeonpea are > US$ 310 million (Ranga Rao et al. 2013). To control this pest, the use of chemical pesticides has increased by 170 fold; from 2.2 g/ha of active ingredient in 1950 to 381 g/ha in 2007. Such dependence on pesticides may lead to insecticide resistance in insects, outbreak of secondary pests, insecticide residues in food chain, and/or loss to biodiversity.
Open-field screening of the wild relatives of pigeonpea such as C. scarabaeoides, C. sericeus, C. lineatus, C. acutifolius, and C. platycarpus revealed that some of their accessions carry high levels of resistance to Helicoverpa pod borer. For pod fly, the resistance can be transferred from C. sericeus; while the resistance to bruchids is available in C. scarabaeoides, C. platycarpus, and C. acutifolius. These resources offer ample opportunities to breeders for developing insect resistant pigeonpea cultivars.
Resistance to diseases
A reasonable number of resistance sources for fusarium wilt and sterility mosaic diseases are available within the primary gene pool. However, resistance to the P3 race of stem blight caused by Phytophthora drechsleri f.sp. cajani, the third most damaging pigeonpea disease, is available only in a wild species (C. platycarpus) from tertiary gene pool. Since this disease is now taking a form of an epidemic in the low-lying and high rainfall areas (Pandey et al. 2011), its genetic solution through inter-specific crop breeding involving C. platycarpus as a donor is essential.
Other economic traits
Soil salinity and drought are important abiotic stresses and four wild species have been found to have good tolerance to these traits. These include C. albicans, C. platycarpus, C. lineatus, and C. cajanifolius for salinity tolerance; and C. acutifolius, C. albicans, C. lineatus, and C. lanceolatus for drought tolerance. Besides these, C. platycarpus has also been reported to carry genes for photo-insensitivity and annual growth habit. Besides resistance to different stresses, the Cajanus wild species have been effectively used to develop unique traits (Table 1) such as cytoplasmic nuclear male sterility and cleistogamous lines (see “Success stories of utilizing wild species in pigeonpea breeding” section). Recently a mapping population segregating for cleistogamous trait have been developed and being used for identification of associated genomics segments (Saxena unpublished data).
The information summarised above indicate that the wild Cajanus species harbour genes for some very important traits. In fact the wild species such as C. scarabaeoides, C. sericeus, C. lineatus, and C. platycarpus can be used to transfer multiple traits to pigeonpea to breed high yielding cultivars with greater stability.
Hurdles in utilizing wild Cajanus in breeding
Plant breeding programmes based on inter-specific hybridizations may encounter several difficulties at any stage between hybridization and selection of desirable recombinants. The key limitations recognised by pigeonpea breeders are summarized in the following text.
Undesirable agronomic traits of wild species
The plants of the wild relatives of pigeonpea are agronomically inferior because of various inherent traits such as undesirable phenology, photo-sensitivity, perenniality, poor dry matter partitioning, pod shattering, high flower drop, and poor expression of yield contributing traits (fewer pods per plant, short fruiting branches, fewer small black seeds etc.). Therefore, these species cannot be expected to contribute directly towards significant gains in productivity; and therefore in most crop improvement programmes the wild relatives have been used only as donor parents for specific traits. However, in some cases wild material may help to remove mildly deleterious alleles fixed in cultivated materials during domestication bottlenecks, or contribute to heterosis. In this direction continuously evolving large scale genomics data in form of SNPs or WGRS can help in bracketing desirable genes/alleles from wild species.
Photo-sensitivity
The induction of flowering in Cajanus requires long nights with sunlight period of 10 h or less; and unfortunately, all the wild species are highly sensitive to photo-period. The only exception is C. platycarpus of the tertiary gene pool and it cannot be crossed to pigeonpea without using embryo rescue technology. Since the sensitivity to photo-period is controlled by dominant genes (Saxena 2008) it does not permit rapid generation advancements. The recent finding of a homolog of EARLY FLOWERING3 (ELF3) gene in pigeonpea (Varshney et al. 2017) can be first target of genomics approaches to develop photo-period insensitive lines.
Natural out-crossing
Unwanted cross-pollination is one of the key factors responsible for generating heterogeneity in a given population. In genus Cajanus, natural cross-pollination is a common event (Saxena et al. 2016) and it is mediated by various insect species such as Apis mellifera, A. dorsata, Megachile bicolor, M. conjuneta, M. lanata, and Xylocopa species (Pathak 1970; Williams 1977; Brar et al. 1992; Verma and Sandhu 1995).
Saxena and Kumar (2010) reported that both cultivated as well as some wild species of secondary gene pool are prone to insect-aided cross-pollination. They observed that the pollinating insects are attracted towards flowers of both cultivated and wild species to collect nectar; and during this process a load of pollen grains get stuck on different body parts. When the pigeonpea pollen-laden insects trip the flowers of certain wild species, the transfer of pollen takes place and it leads to cross-pollination (Fig. 2). The genetic contamination due to natural out-crossing could be as high as 25–30%; this is sufficient to obstruct the selection process and consequently may limit genetic gains in breeding programmes. In this direction introgression of cleistogamy or self-pollination behaviour in pure line varieties can be a genetic tool to maintain purity of elite lines. In pigeonpea an inbred line ICPL 99010 has been bred with cleistogamous flowers. This line was crossed with an open-flower line (ICP 5529) to generate recombinant inbred lines (RILs). These RILs have been used for phenotyping of cleistogamy and open flower behaviour and subsequently genotyped with Axiom®CajanusSNP Array with 56 K SNPs uniformly distributed in genome (Saxena unpublished data). These genomics efforts will provide opportunities to precisely mark the genomics segments with molecular markers for use in Genomics Assisted Breeding (GAB) to develop complete self-pollinating cultivars.
Hard seed coat
Most wild Cajanus species possess hard seed coat which protect seeds from dangerous storage insects, water-logging, and various soil-borne pathogens. Besides these, the hard seed coats also delay the process of germination by 2–3 weeks or even more. Seed hardness also prolongs cooking times and negatively affects grain quality. Since the hard seed coat in the wild species is governed by a single dominant gene (Reddy 1990), its frequency in the segregating generations is relatively high. Therefore, in early segregating populations, some recombinants with useful alleles may be lost due to the direct effect of late germination on plant growth and development leading to reduced plant vigour and inadequate expression of vital yield traits.
Hybrid sterility and cross-incompatibility
Pundir and Singh (1985) and Reddy (1990) reported that the success of inter-specific hybridizations varied across the species and accessions. A perusal of unpublished ICRISAT reports revealed that the hybridization success in C. acutifolius, C. lanceolatus, C. lineatus, C. reticulates, C. laticepalus, and C. trinervius with pigeonpea varied between 4 and 10%; while in C. sericeus, C. scarabaeoides, and C. albicans it was between 15 and 25%. On the other hand, C. platycarpus, C. volubilis and other species from the tertiary gene pool were cross incompatible with cultivated types. Also in the inter-specific hybrid plants, the prevalence of male and/or female sterility of various degrees is quite common and it is associated with molecular diversity of the parents leading to various pre- or post-meiotic abnormalities. In certain cases marginal improvement in the hybridization success rates were achieved by applying specific growth hormone on to the pollinated buds for retaining them on the mother plants for a few more days (Mallikarjuna and Moss 1995).
Linkage drag
Tight genetic linkages between certain economic traits and undesirable plant or seed characters interfere with the transferring of candidate genes from wild species into cultivated types. Such associations are not easy to break and such linked traits are inherited together. These situations are often referred to as “linkage drag” and make the gene transfer from wild species to the cultigen very difficult. Also, it not only fails to release expected recombinants but also discourage breeders to undertake inter-specific breeding programmes. At ICRISAT, while transferring the high protein genes from C. scarabaeoides and C. albicans to the cultivated types, it took 12–14 generations to select recombinant genotypes with high protein, high yield, and good pod and seed size (Saxena and Sawargaonkar 2016).
Pre-breeding: an ideal approach for transferring genes from wild to cultivated species
The entire process of gene transfers from wild to cultivated species is a long term endeavour involving about 10–15 generations. Therefore, the breeding programmes should be having clear objectives and their planning and execution should be done with great care and sufficient resources. Considering the complexities and limitations of such programmes, breeders advise that the entire process be divided into two major parts. In the first part elite advanced generation (F5/F6 lines) breeding products with confirmed presence of the target gene(s) are generated. This activity is popularly known as “pre-breeding”. In the second part, the pre-breeding genetic materials are used as parents in the variety development programmes.
Rick (1984) was the first to use the term “pre-breeding” to transfer one or more useful gene(s) from un-adapted sources into breeding materials. It refers to various plant breeding activities that precede the cultivar development and release. Pre-breeding aims to exploit un-adapted germplasm such as exotic landraces and wild species to enrich the genetic variability within the primary gene pool and develop elite introgression lines with certain combinations of desirable traits from both the wild as well as cultivated species (Sharma and Upadhyaya 2016; Sharma 2017). Since these inbred will be used in future breeding programmes, it would be necessary to characterize them at whole genome level using molecular markers and for key traits such as resistance to various stresses, productivity, combining ability, etc. The concept of pre-breeding is now attracting breeders and slowly it is emerging as a cost effective crop breeding tool. The main activities involved in a typical pre-breeding programme are highlighted in the following text.
Selection of traits and potential donors
The trait selection for inter-specific transfers should receive very high priority. The next logical step in breeding should be to identify the donor species that can be crossed easily with the cultivated type. This should be followed by selection of the best accession within the selected species. This recommendation is based on the fact that a considerable genetic variation has been reported among accessions of a given species (Saxena et al. 1990; Mallikarjuna et al. 2011). In pigeonpea, the most sought after trait is resistance to pod borer, H. armigera. Recently, the P3 race of Phytophthora drechsleri Tucker has also become a serious threat. Since there is no resistance to these constraints in the primary gene pool, a need to mine the desired genes from secondary and tertiary gene pools arises.
For effective transfer of the candidate gene(s), a good understanding about their inheritance is necessary because it will influence the planning and breeding efficiency. The number of genes and their mode of action will help breeders in making decisions related to mating design, population size, and selection strategy. To further clarify this point, an example of resistance to fusarium wilt disease is illustrated here. The resistance to this disease has been reported to be controlled by diverse genetic systems. These include a single recessive gene (Jain and Reddy 1995; Karimi et al. 2010); a single dominant gene (Kotresh et al. 2006; Singh et al. 2016), two dominant complimentary genes (Okiror 2002; Ajay et al. 2013), inhibitory genes (Saxena et al. 2012; Ajay et al. 2013), and duplicate dominant genes (Changaya et al. 2012; Patil et al. 2013). Such a complicated situation with respect to wilt inheritance may arise due to (1) more than one gene controlling the same trait, (2) different genetic backgrounds of the parental lines, (3) various epistatic effects, or (4) the presence of more than one biotype of the fungus. To develop an efficient breeding/selection scheme the selection of parents with known inheritance pattern would be a step in the right direction.
Inter-specific hybridizations
Deodikar and Thakar (1956) were the first to create successful inter-specific (then inter-generic) hybrids by crossing C. cajanus with C. lineatus and C. sericeus to establish genetic affinity between the two species through cytological evidences. To develop inter-specific populations it is important to address the problem of hard seed coat in the wild species and it is done by scarifying the seeds with a sharp blade. Further, for hybridization, it is better to select only a few representative plants within the parental lines. The methodology used in selecting buds, their emasculation and pollination in inter-specific hybridization is more or less similar to that of pigenpea and its details are elegantly described by Sharma and Green (1980). In general, it has been observed that in the inter-specific hybridizations the success is high when the wild species are used as a female parent (Pundir and Singh 1985). The unpublished observations at ICRISAT showed that such reciprocal differences in the hybridization were primarily related to the differences in pollen germination and pollen tube growth.
Hybridization of cultivated type with wild Cajanus species belonging to the tertiary gene pool under normal conditions is not possible due to various pre- and post-fertilization barriers. However, C. platycarpus was successfully crossed with cultivated types following the embryo rescue technique developed by Mallikarjuna et al. (2005).
Generation advancement
In the first (hybrid) generation it is important to assess the hybridity using polymorphic markers in the parents and just not by assessing its morphology with the female parent as done in past so that no self-pollinated plant is picked for generation advancement. The occurrence of abnormal seedlings such as twisted stem, twin seedlings, abnormal leaf shape and size, variable number of leaflets etc. can be observed in the first 2–3 generations. Such seedlings arise due to some cytological abnormalities (Reddy 1981) and should be removed. It has also been observed that the inter-specific populations continue to segregate much beyond F6 generation and keep throwing important segregants in the later generations. In early segregating generations the inter-specific populations will have a fewer plants of interest due to linkage drag and presence of undesirable dominant traits of wild species. Therefore, one must try to grow a large (about 2000 plants) population in F2 and F3 generations and if available use markers associated with the candidate genes for early generation selection. In later generations, the plants with target trait(s) should be selected and advanced further following pedigree method. The selections can further be screened for the target traits and associated markers.
In order to recover the genetic background of cultivated type with minimum linkage drag, at ICRISAT the focus of pre-breeding programmes is on developing large-sized advanced backcross populations. Depending upon the donor parent, usually two backcrosses are sufficient to generate pre-breeding populations or inbred lines having high frequency of useful alleles and acceptable agronomic background (Sharma 2017). The genetic materials developed through pre-breeding conserves a large variability with different trait combinations and it is meant for long term use in the breeding programmes. Therefore, it is necessary to characterize, document, and preserve (short term) the end products with proper evaluation, preferably under multiple environments. This will allow breeders to withdraw the lines of interest for breeding new cultivars with traits of interest. The introgressed lines (ILs) derived through advanced back-crossing approach involving C. acutifolius and C. cajanifolius have been used for multi-location phenotyping and high density genotyping using SNPs for identification of genomic segments associated with yield related traits (Saxena unpublished data).
Integration of genomics in wild Cajanus-based gene transfers
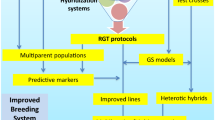
Tanksley and McCouch (1997) articulated the potential role of genome mapping in efficiently utilizing the genetic diversity of wild relatives and suggested that the continued sampling of wild germplasm would result in new gene discoveries and use. In pigeonpea, the wild relatives are quite inferior to their cultivated counterparts, with respect to productivity and other agronomic traits; but these are useful resources for some key traits (see “The economic drivers in wild Cajanus” section, Table 1). Transferring these traits to the cultivated types through traditional breeding approaches require concerted efforts to get rid of unwanted genome segments of wild species which are often inherited due to tight linkages. For this reason, this approach not only requires more time and resources but also the probability of success is relatively low. In this scenario the recently evolved genomics technologies can prove a boon for breeders. The molecular markers that are tightly linked to the target trait can guide the breeders in selecting the trait within large segregating populations with relatively fewer resources. The molecular makers and genetic maps can help in the identification of rare recombinant events leading to breakage of linkages and thereby reducing the proportions of deleterious alleles in the new genetic background. In this endeavour the first major task is to identify a marker that is associated with the target trait (Fig. 3).

A schimatic diagram of utilizing exotic germplasm through genomics approaches
To kick-off the research in this area, Bohra et al. (2011) constructed a genetic map using SSR markers in an inter-specific F2 population involving C. scarabaeoides. From a similar cross a relatively dense genetic map was also developed by Saxena et al. (2012) using SNP markers. Unfortunately, these two genetic maps showed limited genetic resolution and therefore could not be used to find a reliable marker associated with traits. Therefore, to incorporate the useful genes from wild species into pigeonpea, a programme to develop advanced backcrossed introgression lines (ILs) is in progress at ICRISAT. Under this programme, ILs have now been generated by crossing C. acutifolius, C. scarabaeoides, and C. cajanifolius as donor and popular cultivar “Asha” as recurrent parent using backcross breeding approach. In order to develop genetic maps, the introgression lines have been genotyped with SNPs for and it is anticipated that high resolution mapping along with trait phenotyping data will identify markers associated with agronomic important traits (Saxena unpublished data).
Success stories of utilizing wild species in pigeonpea breeding
Wild relatives of pigeonpea have been used sparsly in the crop improvement programmes because of various limitations arising primarily due linkage drag. Nevertheless, inter-specific breeding programmes have been used to transfer high protein and insect resistance from wild species to the cultivated types. Besides this, some recombinants with new useful traits, not available in either of the parental species, have also been recovered from the segregating generations. All the wild species of genus Cajanus have been eligantly described by van der Maesen (1986). Authors have identified the following wild species which have been used successfully in the pigeonpea breeding programmes and their brief information follows:
Cajanus albicans is a perennial climbing wild species with a hardened woody base. It is found in the dry deciduous forests of penninsular India and Sri Lanka. It has large yellow flowers and is highly sensitive to photo-period. Seeds are black and small. Another wild species C. lineatus is also endemic to southern India and Sri Lanka. The plants of this species are perennial, indeterminate, and erect with silky appearance. Its 2-cm long pods are covered with dense hairs. Cajanus sericeus is endemic to deciduous monsoon forests of western and Eastern Ghats of India. Its primary branches are erect and there are no secondary and tertiary branches. The yellow flowers of this species are sessile, axillary, and emerge in bunches of 1–3. C. scarabaeoides is often found climbing on grasses or other annuals. It is endemic to deciduous monsoon forests of Western Ghats and Satpura mountains of India and Sri Lanka. The plants grow vigorously and cover the ground rapidly. Its branches are characterized by winding tops. Pods are 12–20 mm in length with glandular and non-glandular trichomes present on its surfaces. C. cajanifolius, the progenator of pigeonpea, is found growing in the decidous forests of India’s eastern coast and central plains. Its plants are erect and 100–200 cm in height. The branching and flowering pattern, pods, and general appearance of this species are more or less similar to the cultivated type. All the species mentioned above can be crossed easily with the cultivated type.
The following wild species have also been used in breeding programme even though they cannot be crossed easily with pigeonpea. Among these, C. acutifolius, a native of Australia, is generally found growing in the wild habitats of dry, sandy or rocky soils of western and northern areas of the country (van der Maesen 1986). The green leaves of C. acutifolius are covered with numerous dense silvery small hairs. Its plants are 50–200 cm in height with erect or spreading growth habit. Pods are about 15–20 mm long with velvety hairs. Cajanus reticulatus is another wild species that is endemic to northern Australia and is found at the altitudes of around 1000 m. It is a shrub that usually grows to about 150 cm. This species is characterized by long hairs on the pods, stem, and branches. This is the only species where the pods have a prominent constriction between the two consecutive ovules.
Cajanus platycarpus is the only wild species from tertiary gene pool candidate that has been used successfully in pigeonpea breeding. This species is distributed in northwest and central India, Nepal, Pakistan, and Indonesia. It is a unique climbing genotype with annual growth habit. The plants of C. platycarpus are about one meter in height and produce only a small quantity of biomass.
Development of cytoplasmic nuclear male sterility systems
The development of cytoplasmic nuclear male sterility (CMS) systems has been the most valueable contribution of wild species towards pigeonpea breeding. CMS has been extensively used in commercial hybrid breeding in pigeonpea because of its role in large-scale hybrid seed production. In a plant system, CMS arises from an interaction between the cytoplasm of a wild species and nuclear genome of the cultivated type (Kaul 1988). Often these systems result from mutations in mitochondrial genomes (Rieseberg and Blackman 2010; Tuteja et al. 2013). In pigeonpea a breakthrough in this direction was achieved by Saxena et al. (2005) by crossing C. cajanifolius as female with C. cajan as male parent. This CMS system, designated as A4, has been used to develop three commercial pigeonpea hybrids, the first in any food legume (Saxena and Tikle 2015).
Using the approach of inter-specific hybridization a total of nine CMS systems have been bred in pigeonpea; and these can help in diversifyng the hybrid parental base. These new CMS systems (Table 2) involved different wild relatives of pigeonpea from the secondary and tertiary gene pools. Besides C. cajanifolius, the other species which produced CMS systems when crossed with the cultivated types are C.sericeus, C. scarabaeoides, C. lineatus, C. reticulatus, C. acutifolius, C. lanceolatus, and C. platycarpus (Saxena et al. 2010c; Saxena and Tikle 2015; Mallikarjuna et al. 2006; Srikanth et al. 2015).
Development of temperature-sensitive male sterility
The reversion of male sterility to fertility and the vice versa have been reported in a number of crop species (Kaul 1988). Various environmental factors such as photo-period, temperature, and specific stresses have been identified to alter the expression of genes controlling male sterility/fertility. Yuan (1986) and Sun et al. (1989) proposed that this type of male sterility can be used in producing hybrid rice seed economically. The recent success in breeding a temperature sensitive male sterility system in pigeonpea (Saxena 2014) has also opened up a similar option as of rice. Such genotypes when grown under < 24 °C, turn male fertile to produce self-pollinated seeds (Table 3); hence such male sterile lines will not require any maintainer line. The same line, when grown under high (> 25 °C) temperature regime, will remain male sterile; and hence can be used for large-scale F1 hybrid seed production when cross-pollinated by insects.
Delopment of high-protein lines
With a seed protein content of 20–22%, pigeonpea is a prime nutritional source for a great many people of the tropics and sub-tropics. By developing high-protein cultivars without losing grain productivity, the gross protein harvest can be increased significantly. With this objective, three wild relatives of pigeonpea C. albicans, C. sericeus, and C. scarabaeoides with 28–30% seed protein were identified as donor parents. The use of pedigree selection from inter-specific crosses yielded a number of high protein inbred lines (Saxena et al. 1987). Field evaluation of these lines revealed that besides high protein (Table 4), these lines had good yield levels and acceptable seed quality (Saxena and Sawargaonkar 2016).
Development of new plant types
Cleistogamous flowers
Natural out-crossing is a common event in genus Cajanus leading to a rapid genetic contamination of pure lines and germplasm. Within the segregating population of cross C. cajan × C. lineatus (Saxena et al. 1993) a unique recombinant with modified flowers was identified. The flowers had modified petal morphology and anther configuration (Fig. 4). In the genotypes with such flowers the natural out-crossing was found to be 0–2% (Saxena et al. 1994) and therefore, the maintenance of genetic purity became easy. This trait is easy to identify and it is controlled by single recessive gene and hence can be incorporated easily into popular cultivars.

Cleistogamous anthers and flower bud (left) compared with that of a normal cultivar (right)
Genetic dwarfs
In the absence of resistant cultivars, the insect management in pigeonpea is predominantly based on chemical pesticides; but often their efficiency is low due to practical difficulties in spraying the 2–3 m tall crop. Therefore, to realize good yields through effective insect management, breeding of genetically dwarf pigeonpea genotypes would be required. Saxena and Sharma (1995) reported 12 sources of genetic dwarfs in pigeonpea. Of these, D0 dwarf was identified from inter-specific breeding populations involving C. cajan and C. scarabaeoides. The height of the dwarf-inbred lines ranged around 50 cm and produced reasonable yields and it is a reliable genetic stock for future breeding programmes. Mallikarjuna et al. (2011) also reported deriving dwarfs from the population of the cross involving the cultivate type with C. platycarpus. Dwarf phenotypes may also prove to be quite useful in developing varieties well suited to mechanical harvesting and to shorter crop phenology in more water limited settings.
Photo-insensitivity
All the traditional pigeonpea germplasm and landraces are photo-sensitive and they flower during the period when the day length is 10 h or less. This has limited the adaptation of pigeonpea up to 30º latitudes; but the early flowering is linked to reduced degree of photo-sensitivity (Wallis et al. 1981). Hence the early maturing cultivars are adapted to higher latitudes also (Saxena 2008). However, so far no true photo-insensitivity gene has been identified for introgression into late the maturing types. Among the wild species, C. platycarpus has shown promise in this direction. According to Dundas (1990), Pundir and Singh (1987), and Mallikarjuna and Moss (1995) this species from tertiary gene pool is photo-insensitive which can be utilized in breeding. But unfortunately, no serious effort has been made so far to study and characterize this trait with respect to its genetics and photoperiods responses.
Development of phytophthora blight resistance inbreds
Some of the inbred lines derived from crosses involving C. cajan and two wild species C. acutifolious and C. platycarpus were found to have moderate to high levels of resistance to P3 race of phytophthora blight (Sharma 2017; Mallikarjuna et al. 2006). It was also reported that the resistance to phytophthora blight in C. platycarpus (ICPW 61) was under the control of a single recessive gene pair (Mallikarjuna et al. 2006). These inbred lines are being used as parents in breeding genotypes with high yield and resistance to phytophthora blight disease.
Development of insect resistant inbreds
Helicoverpa armigera is the most serious pest of pigeonpea throughout the tropics and subtropics, and breeding for resistance to this pest is a complex issue due to its polyphagous feeding habit and temporal variability in its population (Lateef 1992). Researchers found high levels of resistance to H. armigera in C. sericeus, C. acutifolius, C. albicans, C. scarabaeoides, and C. platycarpus (Pundir and Singh 1987; Lateef 1992; Shanower et al. 1997; Sujana 2005; Mallikarjuna et al. 2011). Unfortunately, so far no genotype with high level of resistance to this pest has been developed; but the efforts are still being made to achieve the goal. Also, efforts are being made to understand the mechanisms of resistance. In this context, Sharma et al. (2001) reported that the pod borer resistance in C. scarabaeoides was due to antibiosis and/or, non-preferential ovi-position. Shanower et al. (1997) observed that even the water extract of C. scarabaeoides pods exhibited significant antifeedant effects. The pod trichomes of the wild species have been reported to provide a natural defence system against insects in a variety of crops (see review by Aruna et al. 2005). Romies et al. (1999) reported that the pods of C. scarabaeoides have a layer of erect and non-glandular trichomes which helps in potecting the pods from borer damage. In contrast, the pods of cultivated types have glandular trichomes and these attract the pod boring insects. Aruna et al. (2005) reported that the trichomes were inherited as single dominant gene. Dodia et al. (1996) reported that the body mass, growth duration, and length of Helicoverpa larvae were adversely affected when they were fed on the flowers of C. cajanifolius, C. sericeus, and C. reticulatus.
Attempts are being made to develop pod borer resistant inbreds using C. acutifolious and C. scarabaeoides as donor parents (Malikarjuna and Saxena 2002; Sharma 2017). Mallikarjuna et al. (2011) evaluated BC4 derivatives from cross involving pigeonpea and C. platycarpus for field resistance to pod borer, podfly, and bruchids and reported high levels of resistance to these pests. The top five progenies identified in each group (Table 5) showed promising results with respect to insect resistance. In comparison to the control (68% pod damage), the pod damage in the selections was from 2–5 to 8.5%. Similar promising results were also recorded for pod fly and bruchids. These progenies are being bred further to enhance their productivity.
Prospects for more efficient uses of wild genetic materials provided by genomic resources
Rapid advances in the power and cost-effectiveness of DNA sequencing technologies have made available the genomic resources for various understudied and under-utilized crops like pigeonpea. In recent years the publication of the pigeonpea genome (Varshney et al. 2012), the development of a wide range of genomic resources (Krieg et al. 2017), and re-sequencing of several hundred accessions to facilitate Genome-Wide Association studies (Varshney et al. 2017) have opened the door to improved use of pigeonpea germplasm and the deployment of advanced approaches such as genomic selection. Importantly, these advances have included re-sequencing wild accessions in gene banks as well as the development of pan-genomes. As described above, these efforts have contributed to clarify the relationship among wild Cajanus, and to provide insight into patterns of genome evolution within the genus. We propose several future steps which can improve the utility of existing wild germplasm in breeding programs.
As the recovery of viable offspring from crosses in the secondary and tertiary gene pools is often quite low, and the labour and time involved in making such crosses and subsequent backcrosses can be quite high, the methodologies that help predict potentially successful crosses would be quite useful. Several pieces of information could help clarify which wild genotype may be most useful. One is to develop high-quality genome drafts for species in the secondary and tertiary gene pool. Importantly, using intra-specific crosses of wild species would allow for the development of wild draft sequence scaffold by recombination maps, as is common practice in developing complete genomes (e.g., Varshney et al. 2012). These de novo high quality wild genomes would help map major chromosomal reorganization across the genus Cajanus, providing insight into the large-scale rearrangements that are likely to greatly impact viability in the widest interspecific crosses. This information would complement pan-genome efforts, and provide insight into genome structural evolution across the genus. Secondly, surveys of CMS systems across plants have suggested that many arise from mitochondrial mutations (Rieseberg and Blackman 2010; Tuteja et al. 2013). Potential mutations can be predicted from whole-genome re-sequencing of wild relatives. These can be annotated. Furthermore, restorer loci can likely be identified from the same surveys.
Another potentially useful information is that a greater understanding the autecology and speciation patterns of the genus. Although agronomic examination of wild Cajanus has indicated many potentially useful traits in the uncultivated members of the genus, relatively little is known about the evolutionary patterns that have driven divergence among Cajanus species or the specific habitats to which different species are adapted. Geological patterns must have played some role in allopatric speciation of some Cajanus species, with the ancient split of the Indian subcontinent, Australia, and Africa likely separating species in these three ancient parts of Gonwanda. But the factors driving divergence within each of these regions have not yet been clarified. We do not know to what extent different wild Cajanus occupy distinct habitats, and to what extent they can be found to overlap with one another. It is not known to what extent wild Cajanus species are reproductively isolated when they in sympatry versus in allopatry, and to what extent natural hybridizations occur among wild Cajanus. Importantly, we do know the extent to which hybridization, if it occurs naturally, has fitness consequences in these natural populations. As some species described above are known to occur in sympatry, with overlapping natural ranges in India and Australia, natural gene flow among these taxa is at least possible between some sympatric populations. If this gene flow is disadvantageous, the reinforcement has exacerbated some differences among these species to reduce gene flow. Understanding these patterns can provide insight into mechanisms that interfere with interspecific mattings. In many other crop wild relatives, such as chickpea’s near relative Cicer echinospermum, wild populations vary in their compatibility with cultivated chickpea (Kahraman et al. 2017). Further the collection of wild relatives, with an emphasis on collecting from new populations that may differ in reproductive isolation, has the potential to enrich available germplasm by providing new compatible lines, new potential sources of disease and pest resistance, and more CMS systems. Furthermore, collecting wild relatives from more extreme habitats, such as areas with high incidence of drought, heat stress, or particular pests, may also capture adaptive alleles currently lacking in available germplasm.
Finally, we believe there is a potential benefit to mining the genomes of wild species directly for useful alleles. One of the most widespread uses of wild germplasm has been and will remain being a source of disease and pest resistance. Increasingly these alleles can be identified directly from sequences rather than from costly phenotypic screens. In particular, methods to identify resistance genes such as RENSEQ (Jupe et al. 2013) can profile wild genomes for potentially useful alleles of resistance genes. We believe these methods will eventually become more cost effective and reliable than field or laboratory phenotypic screens. These methods can prove extremely useful, and could allow for more rapid screening of larger wild germplasm collections.
Conclusions
By 2050, over nine billion human beings are expected to inhabit planet Earth and this will require about 70% more food than is currently required (Alexandratos and Bruinsma 2012). Achieving this target, particularly in the backdrop of looming climate changes will be a Herculean task. This would require new high yielding cultivars with greater resilience and new production technologies. The reality, however, is that the agriculture is becoming more and more expensive and difficult due to significant shifts in agricultural environmental and soil parameters. These result in frequent short term droughts, extreme temperatures, and emergence of new biotypes of pests. To encounter such threats, a search adaptive genes conferring climate resilience in crop species is essential.
In pigeonpea a lot of genetic variability exists for almost all the traits and a part of it has been been used in the release of 86 varieties for diverse agro-ecologies in India (Singh et al. 2016). This programme has helped in enhancing pigeonpea cultivated area and production in india, which accounts for over 85% of global acreage. In spite of these releases there has been no significant gain in the productivity of the crop (Saxena and Tikle 2015; Ahlawat et al. 2016). Therefore, to overcome the deficit of this commodity, it is important to a design and implement a long term crop improvement strategy involving different research disciplines. This may involve (1) search and incorporation of stable resistances to various diseases, insects and their biotypes, (2) development of efficient crop husbandry techniques, (3) enhanced use of modern crop improvement technologies such as genomics, transformation, rapid generation turnover, and hybrid breeding. In this context, the wild relatives of pigeonpea can play a significant role. Some of accomplishments are discussed in this review; but still a lot needs to be done, especially with respect to stable sources of resistances to key insects and diseases, broadening of nuclear and cytoplasmic divesity, and breeding heterotic hybrid parents.
The wild relatives of cultivated species are well-established resources of new genes. It was in the first half of twenteeth century when the wild relatives were increasingly described by collectors, breeders and geneticists by Vavilov and Freier (1951), and deployed in the genetic improvement of crops like sugarcane. Later, the utility of crop wild relatives was recognized in breeding of a number of other crops (Plucknett et al. 1987; Hajjar and Hodgkin 2007; Hoyt 1988). The genetic diversity in the primary gene pool of pigeonpea is limited (Yang et al. 2006) and there is a need to diversify the germplasm through the introgression of genetic materials from its secondary gene pool (Bohra et al. 2010).
The widening of genetic diversity in pigeonpea can be achieved by exploiting secondary gene pool through hybridization and precise marker based selection. The transfer of genes from their wild relatives is a resource intensive operation and often suffers from unwanted linkage drag. Therefore, selection of the candidate trait and is breeding approach should be given very careful consideration. In overcoming the problems associated with linkage drag, the emerging technologies of genomics can be a boon to breeders. In the last half century conventional research has failed to deliver the products most required by small holder farmers. Major issues of plateauing yield and pod borer damage still persist. We believe that the answers to these issues can come through the intelligent use of the genetic materials from the secondary gene pool and advanced genomics science in pigeonpea breeding programmes.
References
Ahlawat IPS, Sharma P, Singh U (2016) Production, demand, and import of pulses in India. Indian J Agron 61:S33–S41
Ajay BC, Prasad PS, Gowda MB, Ganapathy KN, Gnanesh BN, Fiyaz RA, Veerakumari GN, Babu HP, Venkatesha S, Ramya KT (2013) Inheritance of resistance to Bangalore race of Fusarium wilt disease in pigeon pea (Cajanus cajan L.). Aust J Crop Sci 7:1520–1524
Alexandratos N, Bruinsma J (2012) World agriculture towards 2030/2050: the 2012 revision ESA. Working paper no 12-03, FAO, Rome
Ariyanayagam RP, Rao AN, Zaveri PP (1995) Cytoplasmic male-sterility in interspecific matings of Cajanus. Crop Sci 35:981–985
Aruna R, Rao DM, Reddy LJ, Upadhyaya HD, Sharma HC (2005) Inheritance of trichomes and resistance to pod borer (Helicoverpa armigera) and their association in inter-specific crosses between cultivated pigeonpea (Cajanus cajan) and its wild relatives. Euphytica 145:247–257
Bohra A, Mallikarjuna N, Saxena KB, Upadhyaya HD, Vales MI, Varshney R (2010) Harnessing the potential of crop wild relatives through genomics tool for pigeonpea improvement. J Plant Biol 37:83–98
Bohra A, Dubey A, Saxena RK, Penmetsa RV, Poornima KN, Kumar N, Farmer AD, Srivani G, Upadhyaya HD, Gothalwal R, Ramesh S, Singh D, Saxena KB, Kishor PBK, Singh NK, Town CD, May GD, Cook DR, Varshney RK (2011) Analysis of BAC-end sequences (BESs) and development of BES-SSR markers for genetic mapping and hybrid purity assessment in pigeonpea (Cajanus spp.). BMC Plant Biol 11:56
Brar HS, Jhaljj HS, Gatoria GC (1992) Abundance and activity of the bee visits to pigeonpea (Cajanus cajan (L.) Millsp.), and role of Apis mellifera (L.) in its pollination. Indian Bee J 54:76–80
Changaya AG, Melis R, Derera J, Laing M, Saka V (2012) Inheritance of resistance to Fusarium wilt and yield traits in pigeonpea. Euphytica 186:883–896
De DN (1974) Pigeonpea: evolutionary studies in world crops; diversity, and change in the Indian sub-continent. Cambridge University Press, London, pp 79–87
Deodikar GB, Thakar CV (1956) Cytot-axonomic evidence for the affinity between C. indicus, Spreng and certain erect species of Atylosia W and A. Proc Indian Acad Sci B 43(B):37–45
Dodia DA, Patel AJ, Patel IS, Dhulia FK, Tikka SBS (1996) Antibiotic effect of pigeonpea wild relatives on Helicoverpa armigera. Int Chickpea Pigeonpea Newsl 3:100–101
Dundas IS (1990) Pigeonpea: cytology and cytogenetics—perspectives and prospects. In: The pigeonpea, CAB international, Wallingford, UK, pp 117–136
Hajjar R, Hodgkin T (2007) The use of wild relatives in crop improvement: a survey of developments over the last 20 years. Euphytica 156(1–2):1–13
Harlan JR, de Wet JMJ (1971) Towards a rational classification of cultivated plants. Taxon 20:507–517
Hoyt E (1988) Conserving the wild relatives of crops. IPGRI/IUCN/WWF, Rome
Jain KC, Reddy MV (1995) Inheritance of resistance to Fusarium wilt in pigeonpea (Cajanus cajan (L.) Millsp.). Ind J Genet Plant Breed 55:434–437
Jupe F, Witek K, Verweij W, Śliwka J, Pritchard L, Etherington GJ, Maclean D, Cock PJ, Leggett RM, Bryan GJ, Cardle L, Hein I, Jones JDG (2013) Resistance gene enrichment sequencing (RenSeq) enables reannotation of the NB-LRR gene family from sequenced plant genomes and rapid mapping of resistance loci in segregating populations. Plant J 76(3):530–544
Kahraman A, Pandey A, Khan MK, Lindsay D, Moenga S, Vance L et al (2017) Distinct subgroups of Cicer echinospermum are associated with hybrid sterility and hybrid breakdown in interspecific crosses with cultivated chickpea. Crop Sci 57:3101–3111
Kajale MD (1974) Plant economy at Bhokardan. In: Deo SB, Gupte RS (Eds) Excavations at Bhokardan (Bhogacardhana). Nagpur University, Aurangabad, pp 217–224
Karimi R, Owuoche JO, Silim SN (2010) Inheritance of fusarium wilt resistance in pigeonpea (Cajanus cajan (L.) Millspugh). Indian J Genet 70:271–276
Kassa MT, Varma RP, Garciel N, Sarma BK, Datta S, Upadhyaya HD, Varshney RK, Wettberg EJB, Cook DR (2012) Genetic patterns of domestication in pigeonpea (Cajanus cajan (L.) Millsp.) and wild Cajanus relatives. PLoS ONE 7:e39563
Kaul MLH (1988) Male sterility in higher plants. Springer, Berlin
Kotresh H, Fakrudin B, Punnuri SM, Rajkumar BK, Thudi M, Paramesh H, Lohithswa H, Kuruvinashetti MS (2006) Identification of two RAPD markers genetically linked to a recessive allele of a fusarium wilt resistance gene in pigeonpea (Cajanus cajan (L.) Millsp.). Euphytica 149:113–120
Krieg CP, Kassa MT, von Wettberg EJ (2017) Germplasm resources and trait characterization. In: Varshney RK, Saxena RK, Jackson SA (eds) The pigeonpea genome: a compendium. Springer, Berlin, pp 1–4
Lateef SS (1992) Scope and limitations of host plant resistance in pulses for the control of Helicoverpa armigera. In: Sachan JN (ed) Helicoverpa management—current status and future strategies in pigeonpea IIPR. Kanpur, India, pp 31–37
Malikarjuna N, Saxena KB (2002) Production of hybrid between C. acutifolius and Cajanus cajan. Euphytica 124:107–110
Malikarjuna N, Saxena KB (2005) A new cytoplasmic nuclear male-sterility system derived from cultivated pigeonpea cytoplasm. Euphytica 142:143–148
Mallikarjuna N, Moss JP (1995) Production of hybrid between Cajanus platycarpus and Cajanus cajan. Euphytica 83:43–46
Mallikarjuna N, Jadhav D, Reddy MV, Dutta-Tawar U (2005) Introgression of phytophthora blight disease resistance from Cajanus platycarpus into short duration pigeonpeas. Indian J Genet 65:261–264
Mallikarjuna N, Jadhav DR, Reddy P (2006) Introgression of Cajanus platycarpus genome into cultivated pigeonpea Cajanus cajan. Euphytica 149:161–167
Mallikarjuna N, Senthivel S, Deepak RJ, Saxena KB, Sharma HC, Upadhyaya HD, Rathore A, Varshney R (2011) Progress in the utilization of Cajanus platycarpus (Benth.) Maesen in pigeonpea improvement. Plant Breed 130(5):507–514
Mallikarjuna N, Saxena KB, Lakshami J, Varshney RK, Srikanth S, Jadbav DR (2012) Differences between Cajanus cajan (L.) Millsp. and Cajanus cajanifolius (Haines) van der Maesen, the progenitor species of pigeonpea. Genet Res Crop Evol 59:411–417
Nadimpalli RG, Jarret JL, Pathak SC, Kochert G (1993) Phylogenetic relationships of pigeonpea (Cajanus cajan (L.) Millsp.) based on nuclear restriction fragment length polymorphism. Genome 36:216–223
Ohri P, Singh SP (2002) Karyotypic and genome size variation in Cajanus cajan (L) Millsp. (pigeonpea) and some wild relatives. Genet Resour Crop Evol 49:1–10
Okiror MA (2002) Genetics of resistance to Fusarium udum in pigeonpea [Cajanus cajan (L.) Millsp.]. Ind J Genet Plant Breed 62:218–220
Pandey S, Sharma M, Mangala U, Ghosh R, Sundaresan G (2011) Phytophthora blight of pigeonpea (Cajanus cajan (L.) Millsp.): an updating review of biology, pathogenicity and disease management. Crop Prot 30:951–957
Pathak GN (1970) Red gram: pulse crops of India. Indian Council of Agricultural Research, New Delhi, pp 14–53
Patil P, Singh LP, Dhar V, Chaudhary RG, Datta S, Chaturvedi SK, Nadarajan N (2013) Variant specific studies on inheritance for resistance to Fusarium wilt in pigeonpea [Cajanus cajan (L.) Millsp.]. Legume Res 36:576–581
Plucknett D, Smith N, Williams J, Murthi AN (1987) Gene banks and the Worlds Food Princeton. University Press, Princeton
Pundir RPS, Singh RB (1985) Cross ability relationships among Cajanus, Atylosia and Rhynchosia species and detection of crossing barriers. Euphytica 34:303–308
Pundir RPS, Singh RB (1987) Possibility of genetic improvement in pigeonpea utilizing the wild genetic resources. Euphytica 36:33–37
Ranga Rao GV, Rao R, Ghaffar MA (2013) Hand book on chickpea and pigeonpea: insect pests identification and management. Information bulletin 57, International Crops Research Institute for the Semi-Arid Tropics (ICRISAT), Patancheru, 90 pp
Ratnaparkhe MB, Gupta VS, Murthy MRV, Ranjekar PK (1995) Genetic fingerprinting of pigeonpea (Cajanus cajan (L.) Millsp.) and its wild relatives using RAPD marker. Theor Appl Genet 91:893–898
Reddy LJ (1981) Pachytene analyses in Cajanus cajan, Atylosia lineatus and their hybrids. Cytologia 46:397–412
Reddy LJ (1990) Pigeonpea: morphology. In: The pigeonpea, CAB international, Wallingford, Oxon, UK, pp 47–88
Rick CW (1984) Plant germplasm resources. In: Evans DA, Sharp WR, Ammirato PV, Yamada Y (eds) Hand book of cell culture. MacMilan, New York, pp 9–37
Rieseberg LH, Blackman BK (2010) Speciation genes in plants. Ann Bot 106:439–455
Romies J, Shanower TG, Peter AJ (1999) Trichomes on pigeonpea (Cajanus cajan (L.) Millsp.) and two wild Cajanus spp. Crop Sci 39:564–569
Saxena KB (2008) Genetic improvement of pigeonpea—a review. Trop Plant Biol 1:159–178
Saxena KB (2013) A novel source of CMS in pigeonpea derived from Cajanus reticulatus. Indian J Genet 73:259–263
Saxena KB (2014) Temperature-sensitive male sterility system in pigeonpea. Curr Sci 107:277–281
Saxena KB (2015) From concept to field: evolution of hybrid pigeonpea technology in India. Indian J Genet 75:279–293
Saxena KB, Kumar RV (2003) Development of a cytoplasmic nuclear male-sterility system in pigeonpea using C. scarabaeoides (L.) Thouars. Ind J Genet 63:225–229
Saxena KB, Kumar RV (2010) Insect-aided natural out-crossing in four wild relatives of pigeonpea. Euphytica 173:329–335
Saxena KB, Sawargaonkar SL (2016) Genetic enhancement of seed proteins in pigeonpea—methodologies, accomplishments, and opportunities. Int J Sci Res 4:3–7
Saxena KB, Sharma D (1995) Sources of dwarfism in pigeonpea. Indian J Pulses Res 8:1–6
Saxena KB, Tikle AN (2015) Believe it or not, hybrid technology is the way to enhance pigeonpea yields. Intl J Sci Res 5:1–7
Saxena KB, Faris DG, Singh U, Kumar RV (1987) Relationship between seed size and protein content in newly developed high protein lines of pigeonpea. Food Hum Nutr 36:335–340
Saxena KB, Singh L, Reddy MV, Singh U, Lateef SS, Sharma SB, Remanandan P (1990) Intra species variation in Atylosia scarabaeoides (L.) Benth., a wild relative of pigeonpea (Cajanus cajan (L.) Millsp.). Euphytica 49:185–191
Saxena KB, Singh L, Ariyanayagam RP (1993) Role of cleistogamy in maintaining genetic purity of pigeonpea. Euphytica 66:225–229
Saxena KB, Jayasekara SJBA, Ariyaratne HP, Fonseka HHD (1994) Frequency of natural out-crossing in partially cleistogamous pigeonpea lines in diverse environments. Crop Sci 34:660–662
Saxena KB, Kumar RV, Srivastava N, Shiying B (2005) A cytoplasmic-nuclear male-sterility system derived from a cross between Cajanus cajanifolius and Cajanus cajan. Euphytica 145:289–294
Saxena KB, Kumar RV, Dalvi VA, Pandey LB, Gaddikeri G (2010a) Development of cytoplasmic nuclear male sterility, its inheritance, and potential use in pigeonpea hybrid breeding. J Heredity 101:497–503
Saxena KB, Kumar RV, Sultana R (2010b) Quality nutrition through pigeonpea - a review. Health 2:1335–1344
Saxena KB, Sultana R, Mallikarjuna N, Saxena RK, Kumar RV, Sawargaonkar SL, Varshney RK (2010c) Male sterility systems in pigeonpea and their role in enhancing yield. Plant Breed 129:125–134
Saxena RK, Varma Penmetsa R, Upadhyaya HD, Kumar A, Carrasquilla-Garcia N (2012) Large-scale development of cost-effective single-nucleotide polymorphism marker assays for genetic mapping in pigeonpea and comparative mapping in legumes. DNA Res 19:449–461
Saxena RK, von Wettberg E, Upadhyaya HD, Sanchez V, Songok S, Saxena KB, Kimurto P, Varshney RK (2014) Genetic diversity and demographic history of Cajanus spp. illustrated from genome-wide SNPs. PLoS ONE 9:e88568
Saxena KB, Tikle AN, Kumar RV, Choudhary AK, Bahadur B (2016) Nectarivore-aided cross-pollination and its exploitation in genetic enhancement of productivity in pigeonpea. Int J Sci Res 6:321–328
Saxena KB, Kumar RV, Ramarao Saxena RK (2018) Maternal inheritance of male sterility in the progeny of a natural hybrid between Cajanus lineatus and C. cajan. Plant Breed. https://doi.org/10.1111/pbr.12571
Shanower TG, Yoshida M, Peter AJ (1997) Survival, growth, fecundity, and behaviour of Helicoverpa armigera on pigeonpea and two wild Cajanus spp. J Eco Entmol 90:837–841
Sharma Shivali (2017) Pre-breeding using wild species for genetic enhancement of grain legumes at ICRISAT. Crop Sci 57:1132–1144
Sharma D, Green JM (1980) Pigeonpea. In: Hybridization of Crop Plants. Amer Soc Agron Crop Sci. Society of America, Wisconsin, pp 471–481
Sharma S, Upadhyaya HD (2016) Pre-breeding to expand primary genepool through introgression of genes from wild Cajanus species for pigeonpea improvement. Legume Perspect 11:17–20
Sharma HC, Green PW, Stevenson PC, Simmonds MJ (2001) What makes it so tasty for the pest. In: Identification of Helicoverpa armigera (Hubner) feeding stimulates and location of their production on the pod surface of pigeonpea [Cajanus cajan (L.) Millsp.]. Technical report, Competitive Research Facility Project R7029, ICRISAT, Patancheru
Singh N, Tyagi RK, Pandey C (2013) Genetic resources of pigeonpea: conservation for use. National Bureau of Plant Genetic Resources (NBPGR), New Delhi, pp 1–49
Singh IP, Bohra A, Singh F (2016) An overview of varietal development program of pigeonpea in India. Legume Perspect 11:39–42
Smartt J (ed) (1990) Evolution of genetic resources of Pigeonpea. In: Grain legumes, Cambridge University Press, Cambridge, pp 278–293
Srikanth S, Saxena RK, Rao MV, Varshney RK, Mallikarjuna N (2015) Development of a new CMS system in pigeonpea utilizing crosses with Cajanus lanceolatus (WV Fitgz) van der Maesen. Euphytica 204:289–302
Sujana G (2005) Mechanisms of resistance to pod borer Helicoverpa armigera (Hubner) in wild relatives of pigeonpea. PhD theses, Osmania University, Hyderabad, India
Sun ZX, Min SK, Xiong ZM (1989) A temperature-sensitive male sterile line found in rice. Rice Genet Newsl 6:116–117
Tanksley SD, McCouch S (1997) Seed banks and molecular maps: unlocking genetic potential from the wild. Science 277:1063–1066
Tikka SBS, Parmar LD, Chauhan RM (1997) First record of cytoplasmic-genic male-sterility system in pigeonpea [Cajanus cajan (L.) Millsp.] through wide hybridization. Gujarat Agric Univ Res J 22:160–162
Tuteja R, Saxena RK, Davila J, Shah T, Chen W, Xiao YL, Fan G, Saxena KB, Alverson AJ, Spillane C, Town C, Varshney RK (2013) Cytoplasmic male sterility-associated chimeric open reading frames identified by mitochondrial genome sequencing of four Cajanus genotypes. DNA Res 20:485–495
Upadhayaya HD, Reddy K, Gowda CLLG, Singh S (2007) Phenotypic diversity in the pigeonpea core collection. Genet Resour Crop Evol 54:1167–1184
van der Maesen LJG (1980) India is the native home of pigeonpea. In: de Wit HCD (ed) Liber Grantulatorius innonerem. Agricultural University, Wageningen, pp 257–262
van der Maesen LJG (1986) Cajanus DC and Alylosia W & A (Leguminosae). Agricultural University Wageningen papers 85–4. Agricultural University, Wageningen, 222 pp
van der Maesen LJG (1990) Pigeonpea: origin, history, evolution, and taxonomy. In: The pigeonpea, CAB international, Wallingford, UK, pp 15–46
van der Maesen LJG (1995) Pigeonpea. In: Smartt J, Sommond N (eds) Evolution of crop plants. Wiley, Hoboken, pp 251–255. ISBN 9780582086432
Varshney RK, Chen W, Li Y, Bharti AK, Saxena RK, Schlueter JA, Donoghue MT, Azam S, Fan G, Whaley AM, Farmer AD, Sheridan J, Iwata A, Tuteja R, Penmetsa RV, Wu W, Upadhyaya HD, Yang SP, Shah T, Saxena KB, Michael T, McCombie WR, Yang B, Zhang G, Yang H, Wang J, Spillane C, Cook DR, May GD, Xu X, Jackson SA (2012) Draft genome sequence of pigeonpea (Cajanus cajan), an orphan legume crop of resource-poorfarmers. Nat Biotechnol 30:83–89
Varshney RK, Saxena RK, Upadhyaya HD, Khan AW, Yu Y, Kim C, Rathore A, Kim D, Kim J, An S, Kumar V, Anuradha G, Yamini KN, Zhang W, Muniswamy S, Kim JS, Penmetsa RV, von Wettberg E, Datta SK (2017) Whole-genome re-sequencing of 292 pigeonpea accessions identifies genomic regions associated with domestication and agronomic traits. Nat Genet 49:1082–1088
Vavilov N, Freier F (1951) Studies on the origin of cultivated plants. Acme Agency, Buenos Aires, pp 1–185
Verma MM, Sandhu PS (1995) Pigeonpea hybrids: historical development, present status and future prospective in Indian context. Hybrid research and development. Indian Society for Seed Technology, IARI, New Delhi, pp 121–137
Wallis ES, Byth DE, Saxena KB (1981) Flowering responses of thirty-seven early maturing lines of pigeonpea. In: International Workshop on Pigeonpeas, Vol 2, 15–19 Dec 1980, ICRISAT, Patancheru, AP, India, pp 143–150
Weller JL, Ortega R (2015) Genetic control of flowering time in legumes. Front Plant Sci 6:207
Williams IH (1977) Behaviour of insects foraging on pigeonpea (Cajanus cajan (L.) Millsp.). Trop Agric 54:353–363
Yang S, Pang W, Ash G, Harper J, Carling J, Wenzl P, Huttner E, Zong X, Kilian A (2006) Low level of genetic diversity in cultivated pigeonpea compared to its wild relatives is revealed by diversity arrays technology. Theor Appl Genet 113:585–595
Yuan L (1986) Hybrid rice in China. China J Rice Sci 1:8–18
Zeven AC, Zhukossky PM (1975) Dictionary of cultivated plants and their centre of diversity. Centre for Agriculture Publication and Documentation, Wageningen
Acknowledgements
Authors are thankful to the Department of Agriculture Cooperation and Farmers Welfare, Ministry of Agriculture and Farmers Welfare, Government of India; United States Agency for International Development (USAID); Crop Trust; Department of Biotechnology, Government of India and ICRISAT for funding various projects related to pigeonpea. This work has been undertaken as part of the CGIAR Research Program on Legumes and Dryland Cereals (GLDC). ICRISAT is a member of CGIAR Consortium.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Saxena, K.B., Saxena, R.K., Sharma, S. et al. The drivers and methodologies for exploiting wild Cajanus genome in pigeonpea breeding. Euphytica 214, 222 (2018). https://doi.org/10.1007/s10681-018-2298-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10681-018-2298-9