
Abstract
Extra-early quality protein maize (QPM) hybrids with combined tolerance to drought and low soil nitrogen (N) are crucial for reduced food insecurity and malnutrition in sub-Saharan Africa (SSA). Ninety six extra-early QPM single-crosses derived from 24 extra-early QPM inbreds using the North Carolina Design II plus four checks were evaluated under stress (drought and low-N) and non-stress environments in Nigeria, 2012 and 2013. The objectives were to (i) determine the gene action for grain yield and other agronomic traits in the QPM inbreds (ii) identify the best inbred testers across environments (iii) classify the inbreds into heterotic groups and (iv) evaluate the performance and stability of the hybrids. General combining ability (GCA) effects contributed more to the total variation among the hybrids for grain yield and most measured traits under each and across environments suggesting that additive gene effects were more important in the inheritance of the traits. The inbreds were classified into three heterotic groups. Inbreds TZEEQI 7 and TZEEQI 134 were the best testers while TZEEQI 7, TZEEQI 60, TZEEQI 61, TZEEQI 8, TZEEQI 11, TZEEQI 137, TZEEQI 63, TZEEQI 66, TZEEQI 44, and TZEEQI 157 possessed favorable alleles for stress tolerance and could be used for population improvement and development of stress tolerant hybrids. The hybrids TZEEQI 183 × TZEEQI 7, TZEEQI 181 × TZEEQI 7 and TZEEQI 144 × TZEEQI 183 were high yielding and the most stable across environments and should be tested extensively in on-farm trials and commercialized in the sub-region.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Maize (Zea mays L.) is a major staple food crop for rural and urban consumers in sub-Saharan Africa (SSA). Its importance as food and industrial crop in SSA has been increasing steadily over the last few decades (Fakorede et al. 2003). Maize provides about 20 % of the world’s food calories and 15 % of all food proteins, and has potential for reducing the food security challenges posed by the increasing levels of urbanization in SSA countries (Byerlee and Eicher 1997). However, during the last two decades, production and productivity have not matched population growth due to a number factors including poor availability of improved seeds, herbicides, and fertilizers, inadequate agronomic management practices, labour shortage, rising cost of inputs and increased levels of biotic and abiotic constraints. Presently, Striga hermonthica (Del.) Benth parasitism, drought, and low soil nitrogen (low-N) constitute the most important stress factors limiting maize production in SSA (Badu-Apraku et al. 2011a). Of these stress factors, recurrent drought and low-N are the most important.
Considering the ongoing climatic changes caused principally by global warming, the pressure on food production in drought-prone environments will increase in the near future (Curry et al. 1995). Global warming and its associated effects have also changed weather patterns leading to erratic and unreliable amounts and distribution of rainfall which are resulting in several billions of U.S dollars lost annually to drought (Badu-Apraku et al. 2011a). According to Edmeades et al. (1995), 15 % average annual grain yield loss is caused by recurrent drought in SSA. This yield loss may even be as high as 40–90 % when the drought coincides with the flowering and grain filling periods (NeSmith and Ritchie 1992).
Soil nitrogen is important for increased maize production as it promotes vegetative growth, maximizes both kernel initiation and kernel set, and plays a key role in filling the kernel sink (Below et al. 1997). Its deficiency interferes with protein synthesis, induces leaf senescence and therefore reduces the general growth of the plant (Bruns and Abel 2003) thereby limiting yield. In most developing countries, maize production is carried out under low-N conditions (McCown et al. 1992) due to limited use of N fertilizers, reduced N-uptake in drought prone environments, high price ratios between fertilizer and grain yield, non-availability of fertilizer or lack of credit for farmers (Banziger and Lafitte 1997). Annual maize yield losses resulting from low-N varies from 10 to 50 % (Wolfe et al. 1988). Under farmer’s field conditions, drought and soil nutrient deficiencies do occur simultaneously, and the combined effects may be total crop failure (Cechin and Press 1993; Kim and Adetimirin 1997; Badu-Apraku et al. 2011a). Therefore, maize varieties and hybrids targeted to drought prone areas of the sub-region must be tolerant to drought and low-N for increased productivity. The development of the extra-early (80–85 days to physiological maturity) quality protein maize (QPM) germplasm with combined tolerance to drought and low-N is therefore crucial for increased maize productivity in SSA and offers an effective partial solution to enhance maize production and food security among resource poor farmers.
Normal maize has a major nutritional constraint as human food due to its deficiency in two essential amino acids: lysine and tryptophan, even though it contains about 10 % protein. Therefore infants fed on it without balanced protein supplements suffer from malnutrition and develop diseases such as kwashiorkor or pellagra (Badu-Apraku and Lum 2010). QPM has about twice the normal levels of lysine and tryptophan and may supply about 70–73 % of human protein requirements, compared to 46 % from normal maize. The biological value of the protein is 80 % for QPM compared to 40–57 % for normal maize and 86 % for eggs (Bressani 1992). Also, QPM has 90 % relative value (RV) of milk compared to 40 % RV for normal maize. Extra-early QPM cultivars can therefore, be important source of nutrients for the malnourished populations in rural as well as urban areas of SSA.
Combining ability is the ability of genotypes to combine during hybridization such that favourable genes/characters are transmitted to their progenies (Panhwar et al. 2008). Combining ability studies are important in designing plant breeding programs especially in testing procedures when studying and comparing the performance of inbred lines in hybrid combinations. Combining ability can be classified as (i) general combining ability (GCA) which is associated with genes which are additive in their effects and (ii) specific combining ability (SCA) which is attributed to the non-additive gene effects (Rojas and Sprague 1952). The additive gene effects are the predictable portion of the genetic effects and are therefore more useful to plant breeders. In biometrical genetics, North Carolina Design II (Comstock and Robinson 1948) is frequently used by plant breeders to assess the nature of the genetic action influencing quantitative traits. One of the features of this design is that the variation due to hybrids is partitioned into components due to GCA-female and GCA-male (i.e. female, male main effects) and to SCA in terms of male × female interactions (Pixley and Frey 1991).
The International Institute of Tropical Agriculture (IITA) maize improvement program has during the past few years intensified efforts at developing and promoting the adoption of extra-early and early maturing QPM varieties with tolerance/resistance to Striga, drought as well as low soil N. This has resulted in the conversion of several elite normal endosperm extra-early and early cultivars to QPM. Also, extra-early maturing QPM inbreds with increased levels of lysine and tryptophan have been developed. The extra-early QPM inbred lines should be heterotic in hybrid combinations so that they could be successfully utilized for population improvement and for the development of extra-early QPM hybrids which are presently lacking in the sub-region. However, the combining ability and heterotic groups of the extra-early QPM inbreds have not been determined. Furthermore, no extra-early QPM inbred testers are available in SSA to facilitate the development of stress-tolerant hybrids. The identification of extra-early QPM inbred testers and determination of the heterotic groups of the inbreds is crucial for successful extra-early QPM breeding programs in the SSA. The objectives of the present study were to (i) determine the gene action controlling grain yield and other agronomic traits of the inbreds (ii) identify the best QPM inbred testers across stress and non-stress environments (iii) classify the QPM inbreds into heterotic groups and (iv) evaluate the performance and stability of the QPM hybrids across stress and non-stress environments.
Materials and methods
Genetic materials
The extra-early white QPM inbred lines were developed in the IITA Maize Improvement Program from a cross between two extra-early white normal endosperm elite Striga resistant inbred lines and QPM donor source (Pool 15 SR) in an effort to convert them to QPM. The F1 crosses were backcrossed to the recurrent parents to obtain BC1 and BC2. The BC2 was selfed for four cycles to develop 245 S6 inbreds. Based on the results of screening of the 245 inbreds under drought and the analysis of the tryptophan contents, the most promising 18 drought tolerant and six susceptible inbreds with high tryptophan content were selected for the present study (Supplementary Table 1).
Generation of North Carolina Design II crosses
The twenty four drought tolerant and susceptible extra-early white QPM inbred lines were crossed using the North Carolina Design II (NCD II) mating design (Comstock and Robinson 1948) with six sets each consisting of four inbreds. The four inbreds in one set were used as females and crossed with four inbreds in another set used as males. Each inbred line was used as a female parent in one set and as a male parent in another set. The 96 extra-early NCD II crosses plus four checks (two QPM single crosses, one three-way normal endosperm and one commercial normal endosperm hybrid) were evaluated under stress (drought and low-N) and non-stress (well-watered, rainfed and high-N) environments in Nigeria.
Field studies
Two set of trials, inbred trial (24 extra-early white QPM inbred lines used in the NCD II) and hybrid trial (96 extra-early white QPM hybrids plus four checks) were evaluated under drought, low-N, well-watered, rainfed and high-N environments in 2012 and 2013. The inbreds trial was evaluated under terminal drought at Bagauda (lat. 12°00′N, long. 8°22′E, 580 m and 800 mm annual rainfall), a natural drought-prone site during the 2012 rainy season. The inbreds were also evaluated under induced drought stress and well-watered conditions during the dry season of 2012/2013 as well as under rainfed conditions at Ikenne (lat. 3°7′E, long. 6°87′N, elevation 30 m, 1200 mm annual rainfall) during the rainy season of 2013. A randomized complete block design with two replications was used for all the inbred trials at all locations. The experimental units were single-row plots, each 4 m long with inter-row spacing of 0.75 m. Plant spacing within rows was 0.40 m. Three seeds were planted per hill and seedlings later thinned to two per hill at 2 weeks after planting (WAP) to give a final population density of about 66,666 plants ha−1. For the induced drought stress experiment at Ikenne, the plants were irrigated only during the first 3 weeks of growth, using a sprinkler irrigation system which provided 17 mm of water weekly as described by Badu-Apraku and Akinwale (2011) and Badu-Apraku et al. (2011b). The plants were thus subjected to induced drought at the flowering and grain filling periods and relied on the stored moisture in the soil from 21 days after planting (DAP) until maturity. The soil in the experiment station at Ikenne is eutric nitrosol and the experimental area is flat with high water-holding capacity (Soil Survey Staff 1999). The well-watered environment at Ikenne was obtained by supplying irrigation water throughout the growing period. Except for the well-watered treatments, all management practices were the same for both well-watered and drought experiments. Fertilizer was applied at the rate of 60 kg ha−1 each of N, phosphorus and potassium to the well-watered and induced drought experiments at planting. An additional 30 kg ha−1 N was top-dressed at three WAP. However, under terminal drought and rainfed conditions, the fertilizer was applied at 2 and 4 WAP. The trials were kept weed-free with the application of atrazine and gramoxone as pre- and post-emergence herbicides at 5 l/ha each of primextra and paraquat and subsequently, by hand weeding.
The inbred lines were also evaluated under low-N (30 kg ha−1) and high-N (90 kg ha−1) conditions at Mokwa (9°18′N, 5°4′E, 457 m altitude, 1100 mm annual rainfall) and Ile-Ife (7°28′ N, 4°33′ E, and 244 m above sea level, 1200 mm annual rainfall) during the 2012 rainy season. The experimental fields were depleted of nitrogen by continuously planting maize and removing the plant residue after each harvest. Soil samples were taken from 0 to 15 cm depth before planting. The nitrogen content was determined at the IITA soil laboratory at Ibadan, Nigeria according to AOAC (1984) method. The results of the soil analysis revealed that the soil at Mokwa contained 0.085 % of nitrogen, 0.20 % potassium and 6.32 % phosphorus while the soil at Ife contained 0.084 % of nitrogen, 0.358 % of potassium and 2.05 % of phosphorus. Nitrogen fertilizer (Urea) was applied to bring the total available nitrogen to 30 kg ha−1 for the low-N and 90 kg ha−1 for the high-N fields, respectively as revealed by the soil tests. The N fertilizer was applied at two WAP. Single superphosphate and muriate of potash fertilizers were applied to both the low- and high-N blocks to supply 60 kg P2O5 ha−1 and 60 kg K2O ha−1, respectively. The low- and high-N trials were in adjacent blocks, separated by about 10 m alley to minimize the movement of N across blocks. All other crop management practices were the same as described above for the drought and well-watered experiments.
The hybrid trial was evaluated under terminal drought at Bagauda during the 2012 rainy season. The hybrids were also evaluated under induced drought stress during the dry season of 2012/2013 and rainfed conditions during the rainy season of 2013 at Ikenne. In addition, the hybrids were evaluated under low- and high-N conditions at Mokwa and Ile-Ife during the rainy seasons of 2012 and 2013. A 10 × 10 lattice design with two replications was used in all hybrid trials at all locations. The number of rows, planting distance and all other management practices were the same as described for the inbred trials.
Data collection
Data were recorded on drought-stressed, low-N and non-stress (well-watered, rainfed and high-N environments) plots for number of days to 50 % anthesis and silking, plant and ear heights, root and stalk lodging, plant and ear aspects and husk cover. Anthesis-silking interval (ASI) was computed as the difference between number of days to 50 % silking and 50 % anthesis while the number of ears per plant was calculated as the number of harvested ears divided by the number of plants in a plot. In addition, the stay-green characteristic was scored for the drought stressed and low-N experiments at 70 DAP on a scale of 1–9, where 1 represented plants with almost all leaves green and 9, plants with virtually all leaves dead. Shelled grains from cobs harvested from each plot under drought and low-N were weighed. Grain moisture of the shelled grains was measured using Dickey-John moisture meter. Grain yield, adjusted to 15 % moisture content was computed from the shelled grain weight. Under non-stress conditions, cobs harvested from each plot were weighed and representative samples were shelled to determine percent grain moisture content. Grain yield adjusted to 15 % moisture content was computed from cob weight and grain moisture using 80 % shelling percentage.
Statistical analysis
Analysis of variance (ANOVA) were conducted separately for inbred and hybrid experiments on data collected across stress (drought and low-N), non-stress and across stress and the non-stress environments with general linear model (GLM) procedure of statistical analysis system (SAS) software, version 9.3 using a RANDOM statement with the TEST option (SAS Institute Inc 2011). The location-year combinations under each research condition were considered as environments in the combined analysis for each field experiment. In the ANOVA for the hybrid trials, locations, replicates, blocks and hybrids were regarded as random factors under stress, non-stress and across environments. On the other hand, in the ANOVA for the inbred trials, locations, replicates and blocks were considered as random factors while the inbreds were considered as a fixed factor under the contrasting environments. The variation due to hybrids was divided into variation due to male (sets), female (sets), and female × male (sets) interaction. The main effects of male (sets) and female (sets) represented the general combining ability (GCA) while the female × male (sets) interaction represented specific combining ability (SCA) effect (Hallauer and Miranda Filho 1988). The F tests for male (sets), female (sets) and female × male (sets) mean squares were calculated using the mean squares for their respective interaction with environment.
The proportion of GCA-male, GCA-female and SCA for each trait was calculated as the percentage of the sum of squares for the crosses under stress, non-stress and across environments. The GCA-males and GCA-female effects for grain yield and the other traits were computed from the adjusted means using the line × tester approach (Singh and Chaudhary 1985). Standard errors for GCA effects were estimated according to Cox and Frey (1984). Two tailed t tests were used to test the significance of the GCA and SCA effects. In addition, the F test or variance ratio was used to compare the mean squares of the GCA male and female as suggested by Kearsey and Pooni (1996).
To identify the inbred lines or hybrids for combined tolerance to drought and low-N, a base index (I) that integrated increased grain yield across drought and low-N with good plant and ear aspects, delayed leaf senescence, short ASI and increased number of ears per plant was used. The best performing genotypes under stress conditions were identified based on the index values estimated as follows:
where PASP is the plant aspect, EASP the ear aspect, EPP the ears per plant, ASI the anthesis-silking interval and STGR the stay-green characteristic. Each parameter was standardized with a mean of zero and a standard deviation of 1 to minimize the effect different scales. A positive value of I was an indicator of tolerance of the lines or hybrid to stress, while a negative value was an indicator of susceptibility of a line or hybrid (Badu-Apraku et al. 2011a).
Heterotic grouping based on GCA of multiple traits (HGCAMT) proposed by Badu-Apraku et al. (2013, 2015a) was used to group the 24 inbred lines. The grouping was achieved by standardizing the GCA effects (mean of zero and standard deviation of 1) of the traits that had significant means squares for G under each study conditions using the following statistical model:
where Y is HGCAMT, which is the genetic value measuring relationship among genotypes based on the GCA of multiple traits i to n; Yi is the individual GCA effect of genotypes for trait i, \({\rm{\bar{Y}}}\) is the mean of GCA effects across genotypes for trait i, s is the standard deviation of the GCA effects of trait i and εij is the residual of the model associated with the combination of inbred i and trait j.
The traits included grain yield, days to silking and anthesis, ASI, plant and ear heights, ear aspect, plant aspect, ears per plant and the stay-green characteristic. The standardised GCA effects were subsequently subjected to Ward’s minimum variance cluster analysis to construct the dendrogram for grouping of the inbreds using SAS software version 9.3 (SAS Institute Inc 2011).
The means of the grain yield data for the single cross hybrids (best 15 and worst 10) selected using the base index plus the four checks were subjected to the additive main effects and multiplicative interaction (AMMI) analysis to examine the relationships among hybrids (G), environments (E) and G × E Interaction (GEI) and to identify the high yielding and stable hybrids across environments as well as the hybrids adapted to specific environments. The AMMI model used was described in detail by Zobel et al. (1988), Gauch and Zobel (1988) and Crossa (1990).
Results
Significant (P < 0.05) differences were observed among inbreds for grain yield under stress and non-stress environments (Supplementary Table 1). Furthermore, significant (P < 0.05) effects were observed for the G and E mean squares for all measured traits under stress, non-stress and across environments except mean squares of G for plant height and husk cover under stress, ASI, husk cover and EPP under non-stress as well as EPP across environments. The GEI was significant for all measured traits under each and across environments except ASI, plant height and husk cover under stress, days to silking, ASI and ear height under non-stress as well as plant and ear heights across environments (Tables 1, 2). The general combining ability of male (GCAm) and female (GCAf) were significant (P < 0.01) for all measured traits under stress except GCAm for plant height and husk cover. Under non-stress conditions, GCAf and GCAm were significant for all traits except husk cover and EPP for GCAm and ASI for GCAf (Table 1). Across environments, GCAf and GCAm were significant for all traits except GCAm for husk cover (Table 2). The specific combining ability (SCA) was only significant for days to silking and anthesis under stress, grain yield under non-stress conditions (Table 1) and grain yield, days to silking and anthesis and ear aspect across environments (Table 2). Under stress conditions, the GCAm × E interaction were significant for only yield, days to anthesis, ear height and plant aspect while GCAf × E interactions were significant for all measured traits except ASI, plant height and husk cover. The SCA × E interactions were significant for grain yield, days to silking and plant aspect. Under non-stress conditions, GCAm × E interaction effects were significant for days to anthesis, ASI, plant aspect and EPP while the GCAf × E interaction effects were significant for grain yield, husk cover and plant aspect. The SCA × E interaction effect was significant only for plant aspect (Table 1). Across environments, the GCAf × E interaction effects were significant for all traits except days to silking, ASI, husk cover and EPP while GCAm × E interaction mean squares were significant for all traits except ASI, plant height, ear height and husk cover. The SCA × E interaction effects were significant for grain yield, days to silking and plant aspect (Table 2).
Partitioning of hybrid sums of squares into GCA (GCAf + GCAm) and SCA revealed that GCA accounted for more than 50 % (55–87 %) of the total variation among the hybrids for all the measured traits under stress, non-stress and across environments except for plant height under stress environments. There were slight variations in the contributions of the GCAf and GCAm effects to the total sum of squares for all traits under stress and non-stress environments. However, the variance ratio (F test) of the mean squares of GCAm to the GCAf and GCAf to GCAm were not statistically significant (P ≤ 0.05) for the measured traits except ASI with GCAm 3.2 times larger than the GCAf under non-stress environment (Table 3).
The GCA effects of the inbred lines under stress, non-stress and across environments are shown in Table 4. Under stress environments, inbred TZEEQI 183 showed significant and positive GCA effects for grain yield when used as either male or female parent whereas TZEEQI 1 had significant and positive GCA effects for grain yield when used as a male parent while TZEEQI 7, TZEEQI 60 and TZEEQI 134 had significant and positive GCA effects for grain yield when used as female parents. Inbred TZEEQI 109 had significant and negative GCA effects (GCAm and GCAf) for the stay-green characteristic while inbred TZEEQI 102 had significant and negative GCAf for stay-green characteristic. Positive and significant GCAf effects for plant aspect were recorded for inbreds TZEEQI 61 and TZEEQI 145, GCAm for TZEEQI 66 and both GCAm and GCAf for TZEEQI 157. Ear aspect showed significant and positive GCAm for TZEEQI 66 and TZEEQI 157 as well as positive and significant GCAf for TZEEQI 38, TZEEQI 96 and TZEEQI 145. EPP had significant and positive GCAm for inbreds TZEEQI 1, TZEEQI 66 as well as the GCAf for TZEQI 7. Under non-stressconditions, inbreds TZEEQI 7 and TZEEQI 134 had positive and significant (P ≤ 0.5) GCAm for grain yield. TZEEQI 1 and TZEEQI 60 had significant and positive GCA for grain yield only when used as male while TZEEQI 109 and TZEEQI 137 had significant and positive GCA for grain yield when used as female parents. Across stress and non-stressenvironments, inbreds TZEEQI 7, TZEEQI 60, TZEEQI 109, TZEEQI 134 and TZEEQI 183 had significant and positive GCA for grain yield when used as male as well as female parent. However, TZEEQI 1 had significant and positive GCA for grain yield when used as only a male parent.
Heterotic groupings and identification of extra-early white QPM inbreds testers across environments
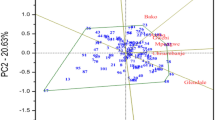
The dendogram constructed based on the HGCAMT revealed three heterotic groups across environments (Fig. 1). Group 1 consisted of TZEEQI 1, TZEEQI 7, TZEEQI 61, TZEEQI 181 and TZEEQI 137; group 2 comprised TZEEQI 60, TZEEQI 109, TZEEQI 134 and TZEEQI 183; while TZEEQI 3, TZEEQI 8, TZEEQI 102, TZEEQI 44, TZEEQI 11, TZEEQI 52, TZEEQI 101, TZEEQI 144, TZEEQI 63, TZEEQI 190, TZEEQI 38, TZEEQI 145, TZEEQI 96, TZEEQI 66 and TZEEQI 157 constituted group 3.

Dendrogram of 24 extra-early white QPM inbreds constructed from GCA effects of grain yield and multiple traits (HGCAMT) using Ward’s minimum variance cluster analysis across environments
According to Pswarayi and Vivek (2008), the choice of a line as a potential tester for classifying other lines into heterotic groups should be based on high and positive GCA effects, classification into a heterotic group and per se grain yield. Based on these criteria, TZEEQI 7 was the highest yielding inbred across environments (Supplementary Table 1), was placed in the first heterotic group, had the highest positive and significant GCA-female and male effects for grain yield and was therefore the best tester for heterotic group 1. Also, TZEEQI 134 recorded high grain yield, had the highest positive GCA-female and male effects for grain yield, was classified into the second heterotic group and was therefore identified as the best tester for heterotic group 2. However, no inbred could be identified as a tester in group 3 because none of the inbreds met the established criteria.
Performance of the extra-early QPM inbreds and hybrids under stress, non-stress and across environments
The mean grain yield of the inbred lines ranged from 523 kg ha−1 for TZEEQI 190–1697 kg ha−1 for TZEEQI 60 with an overall mean of 1129 kg ha−1 and yield reduction of 9–55 % under stress environments (Supplementary Table 1). Under non-stress environments, grain yield ranged from 996 kg ha−1 for TZEEQI 145 to 2435 kg ha−1 for TZEEQI 11 with an overall mean of 1673 kg ha−1. Based on the IITA base index, ten (TZEEQI 7, TZEEQI 60, TZEEQI 61, TZEEQI 8, TZEEQI 11, TZEEQI 137, TZEEQI 63, TZEEQI 66, TZEEQI 44 and TZEEQI 157) out of the 24 inbreds were indentified to have combined tolerance to drought and low-N (Supplementary Table 1). The most stress tolerant (top 15) and most susceptible (worst five) extra-early QPM single cross hybrids and four checks selected under stress environments on one hand and non-stress environments on the other using the base index are presented in Supplementary Table 2. The grain yield of the hybrids under stress environments, ranged from 1177 kg ha−1 for TZEEQI 52 × TZEEQI 63 to 3352 kg ha−1 for the commercial normal endosperm hybrid check-TZEE 21 × TZEEI 29 with an overall mean of 2047 kg ha−1. Under non-stress environments the mean grain yield varied from 1806 kg ha−1 for TZEEQI 52 × TZEEQI 3 to 4580 kg ha−1 for the commercial hybrid check (TZEEI 21 × TZEEI 29) with a mean of 3024 kg ha−1. The highest yielding extra-early normal endosperm hybrid check, TZEEI 21 × TZEEI 29 out-yielded the best QPM hybrid by 25 % under stress and 19 % under non-stress environments. The grain yield reduction ranged from 4–52 % for the hybrids under stress environments. The yield loss was generally associated with increased days to anthesis and ASI, poor ear and plant aspects and fewer ears per plant (Supplementary Table 2).
The AMMI biplot of the grain yield and stability of the 29 extra-early maturing QPM hybrids (top 15, worst 10 plus four checks) selected using the base index under stress environments is presented in Fig. 2. The vertical dotted line of the biplot depicts the grand mean for grain yield, while the horizontal dotted line (y-ordinate) represents the interaction principal component axes 1 (IPCA1) value of zero. Genotypes located close to the horizontal line have small interactions with the environments and are therefore considered to be more stable than those farther away from it while the farther a cultivar is to the right side of the grand mean line, the higher the grain yield. Therefore, extra-early QPM hybrids 5 (TZEEQI 183 × TZEEQI 7), 18 (TZEEQI 181 × TZEEQI 7) and 19 (TZEEQI 144 × TZEEQI 183) were the most stable and relatively high yielding across environments due to their closeness to the zero IPCA1 score and display of grain yield greater than the mean grain yield. Hybrids 2 (TZEEQI 7 × TZEEQI 60), 3 (TZEEQI 7 × TZEEQI 44), 7 (TZEEQI 109 × TZEEQI 134), 8 (TZEEQI 134 × TZEEQI 183) and 16 (TZEEQI 109 × TZEEQI 137) had grain yields greater than the mean grain yield across environments but had a strong positive interaction with the IPCA1 suggesting that they were adapted to high-yield environments (IK13DT, IF12LN, MK13LN, IF12OP and IF13OP). On the contrary, the normal endosperm commercial hybrid check, 28 (TZEEI 21 × TZEEI 29) and the three-way hybrid check 29 ((TZEE 21 × TZEEI 14) × TZEEI 29) were identified as the highest yielding across environments but had a very strong negative interaction with IPCA1 scores indicating that they were adapted to low-yield environments (IK13DT, BG12DT, IF13LN, MK12LN, IK13OP and MK13OP). Of the 100 extra-early hybrids evaluated across drought and low nitrogen, 52 % exhibited varying levels of tolerance to stress (Table not shown).

AMMI biplot of grain yield and the first interaction principal component axis (IPCA 1) of 25 extra-early white QPM hybrids (best 15 and worst 10 based on the base index) and four checks evaluated across 11 environments between 2012 and 2013 in Nigeria. C = check; BG12DT = Bagauda under drought stress, 2012; IK13DT = Ikenne under drought stress, 2012/2013; IF12LN and IF13LN = Ile-Ife under low-N, 2012 and 2013; MK12LN and MO13LN = Mokwa under low-N, 2012 and 2013; IF12HN and IF13HN = Ile-Ife under high-N, 2012 and 2013; K12HN and MO13HN = Mokwa under high-N, 2012 and 2013
Discussion
The significant differences detected among the inbreds and hybrids for grain yield and most measured traits under stress, non-stress and across environments indicated that there was adequate genetic variability among the inbreds and hybrids to allow good progress from selection for improvements of the traits. This result is consistent with the findings of Rosielle and Hamblin (1981), Badu-Apraku et al. (2011b) and Badu-Apraku and Oyekunle (2012). The significant environment and GEI detected for grain yield and most measured traits under stress, non-stress and across environments indicated that the expression of the traits would differ under contrasting environments. This result corroborates the findings of Badu-Apraku et al. (2011a, b) and confirms the important role that environment plays in indentifying tolerant genotypes with favourable response to stress and non-stress conditions.
The significant GCA observed for grain yield and most measured traits under each and across environments indicated that additive gene action was important in the inheritance of grain yield and the other traits. The significant SCA effect detected for grain yield only under non-stress and across environments revealed that non-additive gene action was also important in the inheritance of grain yield under non-stress environments. The preponderance of GCA effects over SCA for grain yield and most measured traits under stress, non-stress and across environments indicated that additive gene action was more important than non-additive gene action for the traits and that GCA was the major contributor to the heritable variation in grain yield and most of the measured traits of the QPM hybrids. The predominance of GCA over SCA effects, indicated that early generation testing may be more effective and that promising hybrids could be identified and selected mainly based on the prediction from GCA effects and that the best hybrids could be obtained by crossing the parents with the highest GCA (Baker 1978; Badu-Apraku et al. 2013). Furthermore, the preponderance of additive gene action in the present study indicated that a great potential existed for the improvement of the traits of interest through recurrent selection methods (Badu-Apraku et al. 2011b) and the identification of suitable parents for direct use or for further breeding (Simmonds 1979). This observation corroborates the findings of other studies on the inheritance of quantitative traits in QPM (Vasal et al. 1993; Bhatnagar et al. 2004). On the other hand, the result disagrees with the findings of Katsantonis et al. (1988) and Betrán et al. (2003) who reported that gene action controlling grain yield varied with the type of stress, with additive gene action more important than non-additive gene action under drought stress while non-additive gene action was more important under low-N. The non-significant SCA mean squares observed for the stay-green characteristic under stress environments indicated that non-additive gene action was not important in the inheritance of stay-green characteristic in this study. This result corroborates the findings of Badu-Apraku et al. (2013) and Ifie et al. (2015). This result however is in disagreement with the findings of Badu-Apraku et al. (2015b) who reported significant GCA and SCA effects for the stay-green characteristic under low-N. The differences in the results of the different studies could be due to the use of different genetic materials. The significant GCAm x environment and GCAf x environment effects obtained for grain yield and most measured traits under stress, non-stress and across environments indicated that the GCA of the parental inbred lines varied in the different environments. This result is consistent with the findings of Badu-Apraku et al. (2013). The authors found significant GCA × E interaction mean squares for most measured traits under different environments. The non-significant SCA for grain yield under non-stress environments and most measured traits under stress, non-stress and across environments revealed that the SCA was consistent in the different environments unlike the GCA. Similar results were reported by Ifie et al. (2015). The larger sum of squares of the GCAm compared to the GCAf for ASI under non-stress environments indicated that paternal inheritance played an important role in the inheritance of ASI under non-stress environments. However, the non-significant differences in the contributions of the GCAf and GCAm sum of squares for grain yield and most measured traits under each and across environments indicated that maternal and paternal effects played similar roles in the inheritance of the traits. Contrary to the results of this study, Derera et al. (2008) reported that maternal effects modified grain yield, ASI and ear aspect under drought as well as ASI and ear aspect under well-watered environments in maize hybrids. Similarly, Oyekunle and Badu-Apraku (2014) showed the importance of maternal effects in the inheritance of grain yield under only well-watered conditions. Also, Adebayo et al. (2014) found paternal effects for ear aspect in late and intermediate maturing single cross hybrids under drought.
Inbred lines displaying significant and positive GCA effects for grain yield are most likely to contribute favorable alleles in a recurrent selection program and could be used as parents to develop synthetic population that could be improved for drought and low-N tolerance. Alternatively, such inbred lines could be used to develop outstanding hybrids for commercialization. The significant positive GCAm effects displayed for the grain yield of inbred TZEEQI 1 and GCAf effects for grain yield of TZEEQI 7, TZEEQI 60 and TZEEQI 134 under stress indicated that the inbreds would contribute favorable alleles for grain yield under stress when used as males and females, respectively. Similarly, the positive and significant GCAm and GCAf effects displayed by TZEEQI 183 suggested that the inbreds could contribute to improved grain yield in their offspring when used either as male or female parent. The negative and significant GCAm and GCAf effects of the stay-green characteristic for TZEEQI 109 suggested that the inbred would contribute alleles that would delay leaf senescence in its progenies when used either as male or female parent. In contrast, the negative and significant GCAf effects exhibited by TZEEQI 102 indicated that the inbred would contribute to delayed leaf senescence in the progeny when used only as a female parent. The significant positive GCAm effects displayed for grain yield by the inbreds TZEEQI 1 and TZEEQI 60 and GCAf effects by TZEEQI 109 as well as TZEEQI 137 under non-stress environment revealed that the inbreds would contribute favorable alleles for grain yield when used either as males or females under non-stress environments. Similarly, the positive and significant GCAm and GCAf effects for grain yield displayed by inbreds TZEEQI 7 and TZEEQI 134 showed that the inbreds could contribute to improved grain yield in their offspring when used either as male or female parents.
Classification of the extra-early QPM inbreds into appropriate heterotic groups is essential to maximize their potential usefulness for the development of productive hybrids and synthetic varieties and for the identification of potential extra-early QPM testers which are lacking completely in SSA. The inbreds classified into the same heterotic group in the present study may be recombined to form heterotic populations that could be improved through recurrent selection. The inbreds TZEEQI 7 and TZEEQI 134 identified as the best testers in the present study could be utilized for classifying QPM inbreds into heterotic groups, to assess the combining ability and to identify superior hybrid combinations in tropical extra-early QPM.
The grain yield reductions of 9–55 % for the extra-early inbreds and 4–52 % for hybrids under stress environments revealed that the levels of stress imposed during the evaluations were severe enough to allow effective discrimination among the inbreds and hybrids. The yield reductions recorded in the present study fall within the range reported by previous workers (NeSmith and Ritchie 1992; Badu-Apraku et al. 2005; Derera et al. 2008 and Badu-Apraku et al. 2011b).
The positive base index values recorded for TZEEQI 7, TZEEQI 60, TZEEQI 61, TZEEQI 8, TZEEQI 11, TZEEQI 137, TZEEQI 63, TZEEQI 66, TZEEQI 44 and TZEEQI 157 under stress environments suggested that these inbreds possess stress tolerant genes. These inbreds may be used as germplasm sources for introgression of genes for stress tolerance into tropical extra-early QPM populations and for the development of hybrids and synthetic varieties with combined tolerance to drought and low-N. Hybrids, TZEEI 21 × TZEEI 29 (commercial check), TZEE 21 × TZEEI 14 × TZEEI 29 (check), TZEEQI-134 × TZEEQI-181 (QPM) and TZEEQI-7 × TZEEQI-60 (QPM) identified as the most outstanding in terms of stress tolerance should be tested extensively in multi-location trials and promoted for adoption and commercialization in the stress-prone areas of SSA.
Presently, there are no adapted extra-early maturing quality protein maize hybrids with high grain yield potential which could be of great benefit to resource poor maize farmers through increased production and improved incomes and nutrition in SSA. In the present study, the highest yielding extra-early normal endosperm hybrid check, TZEEI 21 × TZEEI 29 out-yielded the best QPM hybrid by 25 % under stress and 19 % under non-stress conditions. The lower grain yield recorded for the extra-early QPM hybrids was not surprising because the inbred lines used in the present study were selected based on only their reactions to drought and tryptophan content and had not been grouped into their respective heterotic groups before the study to facilitate the exploitation of maximum heterosis through carefully planned crosses. The results of this study is a clear indication that there is a need for more efforts and resources to be devoted to breeding more yield competitive stress tolerant extra-early QPM maize inbreds and hybrids. Nevertheless, inbred lines identified with positive and significant GCA effects for grain yield (TZEEQI 7, TZEEQI 60, TZEEQI 60, TZEEQI 109, TZEEQI 134 and TZEEQI 183) across environment in the present study could be crossed with other QPM inbred lines from different genetic sources to increase the chances of identifying more productive extra-early QPM hybrids.
The QPM hybrids, TZEEQI 183 × TZEEQI 7, TZEEQI 181 × TZEEQI 7 and TZEEQI 144 × TZEEQI 183 identified by the AMMI biplot as relatively high yielding and most stable across environments should be tested extensively for consistency of their performance and commercialized in the sub-region. However, the hybrids, TZEEQI 7 × TZEEQI 60, TZEEQI 7 × TZEEQI 44, TZEEQI 109 × TZEEQI 134, TZEEQI 134 × TZEEQI 183, TZEEQI 109 × TZEEQI 137, TZEEI 21 × TZEEI 29 and (TZEE 21 × TZEEI 14) × TZEEI 29 identified to be high yielding in specific environment could also be tested extensively in those environments and commercialized.
In conclusion, the GCA effects contributed more than 50 % (55–87 %) of the total variation among the hybrids for grain yield and most measured traits under each and across environments suggesting that additive genetic effects were more important in the control of inheritance of the traits. The inbreds were classified into three heterotic groups based on the HGCAMT. Inbreds, TZEEQI 7 and TZEEQI 134 were identified as the best testers and could be used to group tropical extra-early QPM inbreds. Inbreds TZEEQI 7, TZEEQI 60, TZEEQI 61, TZEEQI 8, TZEEQI 11, TZEEQI 137, TZEEQI 63, TZEEQI 66, TZEEQI 44 and TZEEQI 157 possess genes for stress tolerance. These inbreds may be used as germplasm sources for introgression of genes for stress tolerance into extra-early QPM populations and for the development of hybrids and synthetic varieties with combined tolerance to drought and low-N. The QPM hybrids TZEEQI 183 × TZEEQI 7, TZEEQI 181 × TZEEQI 7 and TZEEQI 144 × TZEEQI 183 were identified as high yielding and most stable across environments. These hybrids should be tested extensively in on-farm trials and commercialized in the sub-region while efforts continue to develop extra-early QPM hybrids which are competitive with the normal endosperm extra-early hybrids.
References
Adebayo MA, Menkir A, Blay E, Gracen V, Danquah E, Hearne S (2014) Genetic analysis of drought tolerance in adapted x exotic crosses of maize inbred lines under managed stress conditions. Euphytica 196:261–270
AOAC (1984) Official methods of analysis, 14th edn. Association of Official Agricultural Chemists, Washington, DC, pp 9–23
Badu-Apraku B, Akinwale RO (2011) Identification of early-maturing maize inbred lines based on multiple traits under drought and low N environments for hybrid development and population improvement. Can J Plant Sci 91:931–942
Badu-Apraku B, Lum AF (2010) The pattern of grain yield response of normal and quality protein maize cultivars in stressed and non-stressed environments. Agron J 102:381–394
Badu-Apraku B, Oyekunle M (2012) Genetic analysis of grain yield and other traits of extra-early yellow maize inbreds and hybrid performance under contrasting environments. Field Crops Res 129:99–110
Badu-Apraku B, Menkir A, Lum AF (2005) Assessment of genetic diversity in extra-early Striga resistant tropical inbred lines using multivariate analyses of agronomic data. J Genet Breed 59:67–80
Badu-Apraku B, Fakorede MAB, Oyekunle M, Akinwale RO (2011a) Selection of extra-early maize inbreds under low N and drought at flowering and grain-filling for hybrid production. Maydica 56:1721–1735
Badu-Apraku B, Oyekunle M, Akinwale RO, Lum AF (2011b) Combining ability of early-maturing white maize inbreds under stress and nonstress environments. Agron J 103:544–557
Badu-Apraku B, Oyekunle M, Fakorede MAB, Vroh I, Akinwale RO, Aderounmu M (2013) Combining ability, heterotic patterns and genetic diversity of extra-early yellow inbreds under contrasting environments. Euphytica 192:413–433
Badu-Apraku B, Annor B, Oyekunle M, Akinwale RO, Fakorede MAB, Talabi AO, Akaogu IC, Melaku G, Fasanmade TY (2015a) Grouping of early maturing quality protein maize inbreds based on SNP markers and combining ability under multiple environments. Field Crops Res 183:169–183
Badu-Apraku B, Fakorede MAB, Talabi AO, Oyekunle M, Akaogu IC, Akinwale RO, Annor B, Melaku G, Fasanmade Y, Aderounmu M (2015b) Gene action and heterotic groups of early white quality protein maize inbreds under multiple stress environments. Crop Sci 56:183–199
Baker RJ (1978) Issues in diallel analysis. Crop Sci 18:533–536
Banziger M, Lafitte HR (1997) Efficiency of secondary traits for improving maize for low-nitrogen target environments. Crop Sci 37:1110–1117
Below FE, Brandau PS, Lambert RJ, Teyker RH (1997) Combining ability for N use efficiency in maize. In: Edmeades GO, Banziger M, Mickeisen HR, Pena-Valdivia CB (eds) Drought and low-N tolerant maize: Proceedings of a symposium Elbatan, Mexico, 25–29 March 1996. CIMMYT, Mexico City, pp 316–319
Betrán JF, Ribaut JM, Beck DL, Gonzalez de Leon D (2003) Genetic analysis of inbred and hybrid grain yield under stress and non-tress environments in tropical maize. Crop Sci 43:807–817
Bhatnagar S, Betran FJ, Rooney LW (2004) Combining abilities of quality protein maize inbreds. Crop Sci 44:1997–2005
Bressani R (1992) Nutritional value of high-lysine maize in humans. In: Mertz ET (ed) Quality protein maize. American Association of Cereal Chemists, St. Paul, pp 205–225
Bruns HA, Abel CA (2003) Nitrogen fertility effects on Bt Endotoxin and nitrogen concentrations of maize during early growth. Agron J 95:207–211
Byerlee D, Eicher C (1997) Africa’s emerging maize revolution. Lynee Reienner Publishers, Cary
Cechin I, Press MC (1993) The influence of nitrogen on growth and photosynthesis of sorghum infected with Striga from different provenances. Weed Res 3:289–298
Comstock RE, Robinson HF (1948) The components of genetic variance in population of bi-parental progenies and their use in estimating the average degree of dominance. Biometrics 4:254–266
Cox J, Frey KJ (1984) Combining ability and the selection of parents for specific oat mating. Crop Sci 24:963–967
Crossa J (1990) Statistical analyses of multilocation trials. Adv Agron 44:55–85
Curry RB, Jones JW, Boote KJ, Peart RM, Allen LH, Pickering NG (1995) Response of soybean to predicted climate change in the USA. Climate Change and Agriculture: Analysis of Potential International Impacts. ASA Special Publication, Madison, pp 163–183
Derera J, Tongoona P, Vivek BS, Laing MD (2008) Gene action controlling grain yield and secondary traits in southern African maize hybrids under drought and non-drought environments. Euphytica 162:411–422
Edmeades GO, Banzinger M, Chapman SC, Ribaut JM, Bolaños J (1995) Recent advances in breeding for drought tolerance in maize. In: Badu-Apraku B, Akoroda MO, Ouedraogo M, Quin FM (eds) Contributing to food self-sufficiency: maize research and development in West and Central Africa. Proceedings of a Regional Maize Workshop. May 28–June 2, 1995. IITA-Cotonou, Cotonou, pp 24–41
Fakorede MAB, Badu-Apraku B, Kamara AY, Menkir A, Ajala SO (2003) Maize revolution in West and Central Africa: an overview. In: Badu-Apraku B, Fakorede MAB, Ouedraogo M, Carsky RJ, Menkir A (eds) Proceedings of a Regional Maize Workshop. IITA-Cotonou, Cotonou, pp 14–18
Gauch HG, Zobel RW (1988) Predictive and postdictive success of statistical analyses of yield trials. Theor Appl Genet 76:1–10
Hallauer AR, Miranda Filho JB (1988) Quantitative genetics in maize breeding, 2nd edn. Iowa State University Press, Ames. p. 468
Ifie BE, Badu-Apraku B, Gracen V, Danquah EY (2015) Genetic analysis of grain yield of IITA and CIMMYT early-maturing maize inbreds under Striga-infested and low-soil nitrogen environments. Crop Sci 55:610–623
Katsantonis N, Gagianas A, Fotiadis N (1988) Genetic control of nitrogen uptake, reduction and partitioning in maize (Zea mays L.). Maydica 33:99–108
Kearsey MJ, Pooni HS (1996) The genetical analysis of quantitative traits. Chapman and Hall, London
Kim SK, Adetimirin VO (1997) Responses of tolerant and susceptible maize hybrids to timing and rate of nitrogen under Striga hermonthica infestation. Agronomy Journal 89:38–44
McCown RL, Keating BA, Robert RE, Jones RK (1992) Strategies for sustainable crop production in semi-arid Africa. Outlook on Agriculture 21:21–31
NeSmith DS, Ritchie JT (1992) Effects of soil water-deficits during tassel emergence on development and yield components of maize (Zea mays L.). Field Crops Res 28:251–256
Oyekunle M, Badu-Apraku B (2014) Genetic analysis of grain yield and other traits of early-maturinmg maize inbred lines under drought and well-watered conditions. J Agro Crop Sci 200:92–107
Panhwar SA, Baloch MJ, Jatoi WA, Veesar NF, Majeedano MS (2008) Combining ability estimates from line x tester mating design in upland cotton. Proc Pak Acad Sci 45(2):69–74
Pixley KV, Frey KJ (1991) Combining ability for test weight and agronomic traits of oat. Crop Sci 31:1448–1451
Pswarayi A, Vivek BS (2008) Combining ability amongst CIMMYT’s early maturing maize (Zea mays L.) germplasm under stress and non-stress conditions and identification of testers. Euphytica 162:353–362
Rojas BA, Sprague GF (1952) Comparison of variance components in corn yield trials: III. General hybrids of maize. Ind J Genet 62:312–315
Rosielle AA, Hamblin J (1981) Theoretical aspects of selection for yield in stress and non-stress environments. Crop Sci 21:943–946
SAS Institute Inc. (2011) Base SAS_ 9.3 Procedures Guide. SAS Institute Inc., Cary
Simmonds NW (1979) Principles of Crop Improvement. Longman, London
Singh RH, Chaudhary BD (1985) Biometrical methods in quantitative genetic analysis. Kalyani publisher, New Delhi, pp 103–157
Soil Survey Staff (1999) Soil taxonomy: A basic system of soil clas-sification for making and interpreting soil surveys, 2nd edn. USDA–NRCS Agriculture Handbook No. 436. U.S. Gov. Print. Office, Washington, DC. pp. 869
Vasal SK, Srinivasan G, Pandey S, González F, Crossa J, Beck DL (1993) Heterosis and combining ability of CIMMYT’s quality protein maize germplasm: I. Lowland Trop Crop Sci 33:46–51
Wolfe DW, Henderson DW, Hsiao TC, Alvio A (1988) Interactive water and nitrogen effects on maize. II. Photosynthetic decline and longevity of individual leaves. Agron J 80:865–870
Zobel RW, Wright MJ, Gauch HG (1988) Statistical analysis of a yield trial. Agron J 80:388–393
Acknowledgments
The authors appreciate the financial support of the Alliance for a Green Revolution in Africa (AGRA) and the International Institute of Tropical Agriculture (IITA) for this study. We are also grateful to the staff of the IITA Maize Improvement Program in Ibadan, Nigeria, for technical assistance.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Annor, B., Badu-Apraku, B. Gene action controlling grain yield and other agronomic traits of extra-early quality protein maize under stress and non-stress conditions. Euphytica 212, 213–228 (2016). https://doi.org/10.1007/s10681-016-1757-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-016-1757-4