
Abstract
Periodic assessments of reference condition wetlands are needed to determine changes over time; however, they are rarely conducted. The vegetation from past assessments, 1998 to 2004, was compared to 2016 assessments of 12 reference wetlands in the Missouri Coteau sub-ecoregion of the Prairie Pothole Region using nonmetric multidimensional scaling and permutational multivariate analysis of variance. Analyses indicated the vegetation in the 2016 assessments trended away from the abundance of native highly conservative species as found during the 1998 to 2004 assessments. Instead, the 2016 plant communities trended towards lower abundance of the same native conservative species and higher abundance of non-native species. Both the average coefficient of conservatism values and floristic quality index values significantly declined, supporting the interpretation that reference wetlands were moving towards plant communities with lower abundance of highly conservative species. The assumption that reference wetlands in the Prairie Pothole Region will change little over time is challenged by these findings. Vegetation in refence wetlands within the Prairie Pothole Region is no longer resembles past monitoring and is trending towards a distinct vegetation composition. Future management will need to consider the potential of reference wetlands’ vegetation composition moving away from a historic baseline and how this may impact future wetland assessment, especially when vegetation is compared to reference conditions.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Reference wetlands, by definition, operate at the highest ecological function and generally provide the highest benchmark for condition and function (Brinson & Rheinhardt, 1996; Hawkins et al., 2010; Herlihy et al., 2019). Most wetland assessments, whether at the landscape, rapid, or in-depth level, are validated through processes that involve comparison to reference condition wetlands in the area (Herlihy et al., 2019; Magee et al., 2019; Stein et al., 2009). Defining reference conditions and identifying corresponding reference wetlands occurs early in the creation of an assessment protocol (Brinson & Rheinhardt, 1996; Otte et al., 2021; Smith et al., 1995).
Wetlands across the world are changing due to a variety of natural and anthropogenic factors including changing hydrology, changes in surrounding land use, and constant pressure from invasive and exotic species (Zedler & Kercher, 2005). Reference wetlands are not immune to degradation, and increases in invasive species and subsequent decreases in species richness have been documented in reference wetlands (DeKeyser et al., 2003; Kentula et al., 2004; Otte et al., 2021). Currently, there is debate as to which baseline reference wetlands should be compared, that is, historic conditions or those that include anthropogenic influences and limitations (Kopf et al., 2015; McNellie et al., 2020; Otte et al., 2021; Stoddard et al., 2006). Documenting the character and extent of changes occurring at wetland reference sites will help inform decisions regarding appropriate baselines to compare a region’s wetlands.
The Prairie Pothole Region (PPR) of central North America is one of the world’s most wetland-rich ecosystems (Doherty et al., 2018). Efforts to protect wetlands in the US’ portion of the PPR began with simply setting aside habitat and evolved through the decades to assess wetland condition and preserve function (Dixon et al., 2019; Hargiss et al., 2017). In recent years, wetland condition assessment has taken a multi-level approach dependent upon the identification of reference wetlands to quantify departure from reference conditions (Mack, 2006; Stoddard et al., 2006; US EPA, 2016; Hargiss et al., 2017; Herlihy et al., 2019; Gallaway et al., 2020). Vegetation is often used in condition assessments because vegetation has proven to be one of the most sensitive and ever-present features available to assess wetland condition (DeKeyser et al., 2003; Herlihy et al., 2019).
The PPR is considered one of the least impacted wetland systems in the world (Otte et al., 2021). Still, periodic wetland assessments are needed in the PPR to help identify and quantify any recent disturbance and trigger conservation efforts (US EPA, 2016), as well as to help evaluate the success of wetland restoration or replacement efforts (Moreno-Mateos et al., 2012; Otte et al., 2021; Paradeis et al., 2010). In order to adequately observe change over time, it is estimated that more than 10 years are needed between assessments (Gong et al., 2010; Wilson & Nilsson, 2009). To date, the authors are unaware of any studies in the PPR that document change over time (10 years +) of reference condition wetlands. The goal of the current project was to assess if reference condition wetlands in the PPR are changing over time. Understanding potential changes in one of the least impacted wetland systems in the world will inform the debate as to whether it is appropriate to assume PPR refence wetlands reflect historic baselines.
Materials and methods
Site selection
The study area for this project was the Missouri Coteau sub-ecoregion of the PPR within the boundaries of the state of North Dakota. The Missouri Coteau is known for having extensive mixed grass prairie, is poorly drained with many depressions occupied by wetlands, and is the most wetland-rich sub-ecoregion within the PPR (Bryce et al., 1996; Stewart & Kantrud, 1971). The dominant land uses in this ecoregion include farming of small grain, corn, soybean, and sunflowers, as well as livestock ranching.
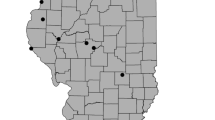
Surveys conducted by the North Dakota Department of Environmental Quality and the Environmental Protection Agency (EPA) between 1998 and 2004 were used as the sample population for this study (DeKeyser et al., 2003; Hargiss et al., 2008). The sample population consisted of 24 reference wetlands within the Missouri Coteau, including 10 seasonal, nine temporary, and five semi-permanent wetlands. The vegetation of the reference wetlands sample population was sampled extensively at least 10 years prior to 2016. Permission was obtained to survey 19 wetlands from the sample population, of which 12 (four temporary, four seasonal, and four semi-permanent) were randomly selected for sampling in 2016 (Fig. 1).

Location of the 12 reference wetlands within the Missouri Coteau sub-ecoregion of the Prairie Pothole Region (shaded area is the Missouri Coteau sub-region within North Dakota and the boundary of the Prairie Pothole Region is the solid black line)
Reference wetlands were located within a wetland complex landscape typical of the Missouri Coteau. Land use surrounding the reference wetlands had not changed since the 1998–2004 sampling and consisted of native prairie managed with periodic fire and grazing that ranged from moderate yearly grazing to periodic light grazing (see supplementals table S1 for land use at each refence wetland and their location). The past (1998–2004) and present (2016) surveys of the reference wetlands were both conducted during dry periods as measured by the Palmer hydrologic drought index (Fig. 2).

Regional monthly Palmer hydraulic drought index values for the past (1998 to 2004) and present (2016) sampling periods
Vegetation sampling methods
Vegetation sampling for past and present assessments occurred between late June and early August. In 2016, the plant community at each site was sampled using a 1-m2 quadrat method, similar to DeKeyser et al. (2003) and Hargiss et al. (2008). Vegetation zones assessed for temporary wetlands consisted of the low prairie and wet meadow zones, adding the shallow marsh zone for seasonal and semi-permanent wetlands and the deep marsh zone for semi-permanent wetlands (Stewart & Kantrud, 1971). A total of eight quadrats were evenly distributed and sampled in the low prairie, seven in the wet meadow, five in the shallow marsh, and five in the deep marsh zones. Within each quadrat, a list of primary species was generated with each species assigned a percent aerial cover. Additionally, species located between quadrats, but not within quadrats were documented as secondary species.
The methods in 2016 were designed to adequately sample each vegetation zone and be compared to the past surveys where a higher number of 1-m2 quadrats was used in each wetland zone. The reduction in the number of quadrats used in the 2016 survey was due to the sample adequacy analysis by DeKeyser (2000) using the species area curve method (Peck, 2016). Results of the sample adequacy analysis using the species area curve method by DeKeyser (2000, p. 28) indicated that the past surveys oversampled the wetland zones, i.e., the past surveys used more quadrats than necessary to establish the number and abundance of species at the reference wetland sites. Thus, using fewer quadrats in 2016 adequately sampled the number of species and their abundance in each of the 12 wetlands and the past and present surveys likely were comparable (see supplemental table S2 for the difference in sampling methods for each reference wetland).
The percent aerial cover for each species was averaged over all quadrats irrespective of wetland zones within each wetland for each sample period. The species aerial cover for each wetland was visualized using Nonmetric Multidimensional Scaling (NMS) in PC-ORD Version 7 (McCune & Mefford, 2011) using the Relative Sorenson distance measure. The NMS was conducted using a procedure based on meeting these conditions: “1) the final stress less than 20, 2) randomization test P ≤ 0.05, and 3) a reduction of at least 5 points of stress with each additional axis” (Peck, 2016). A permutational multivariate analysis of variance (PERMANOVA) of the species percent cover by wetland was used to test if past and present plant communities were different using PRIMER-E (Version 7) and the PERMANOVA + module (Anderson et al., 2008). Time, with two levels past and present, was treated as a fixed effect while sites were treated as random effects. The Relative Sorenson distance measure was also used in the PERMANOVA analysis. All percent data were arc-sine square root transformed before analysis.
There was a slight difference in the treatment of secondary species between the past and present surveys. Each of the 12 present surveys recorded secondary species while four of the past surveys did not. Secondary species were sparsely distributed within each of the wetland zones (secondary species were those that were found between 1-m2 quadrats but not within 1-m2 quadrats at a wetland site.) Because of their sparse distribution and correspondingly low abundance, secondary species are arbitrarily assigned an aerial cover of 0.1% for the plant community analysis. While the inclusion of secondary species does increase the species richness of a wetland site, using the Relative Sorenson distance measure for the NMS and PERMANOVA reduces the influence of sparse species (i.e., secondary species) (Peck, 2016). Using the Relative Sorenson distance measure balances the heterogeneity and sparsity between the comparison datasets with their proportional abundance, thereby placing more influence on the abundance of redundant species and reducing sensitivity to outliers. Thus, not including secondary species in four of the past datasets, but including secondary species in all the present datasets should not unduly influence the analysis.
Species lists generated from past and present surveys were used to calculate the average coefficient of conservatism value (C-value) and floristic quality index (FQI) for reference wetland plant communities in order to examine potential changes over time. The C-values and FQI provide numerical measures reflective of plant community condition useful for the comparison of plant community conditions (NGPFQAP, 2001; Mushet et al., 2002; Freyman et al., 2016). The average C-value and FQI were calculated based on the presence of native plant species within each wetland wherein each native plant species was assigned a C-value between 0 and 10 with higher C-values awarded to conservative native species, while lower C-values are given to “weedy” and opportunistic native species (NGPFQAP, 2001). Non-native species are not given a C-value and are not included in FQI calculations. The FQI is calculated using the following equation: FQI = average C-value × √N, where N = the total number of native species (NGPFQAP, 2001).
The average C-value and FQI for each pair of past and present surveys of reference wetlands were compared using a paired t-test. This test determines if the differences among the 12 paired wetland observations were significantly different from zero. Present values were subtracted from past values to provide a value reflecting change over time. In this analysis, negative values indicate a decrease in average C-value or FQI over time.
Results
The NMS analysis of all wetland zones produced a final result with three dimensions (final stress = 10.73163, randomization test P < 0.05, final instability = 0.00000, number of iterations = 82, I = 0.6875, A = 0.3017) representing 86% of the variation in the dataset. Axis 1 represented most of the of the variation (42.6%), while axis 2 and axis 3 represented similar amounts of variation (axis 2 = 25.2%, axis 3 = 18.1%) (Figs. 3 and 4). The PERMANOVA test indicated that the past and present reference wetland plant communities were significantly different (P < 0.001), and all sites had secessional vectors moving toward the negative end of axes 1 and 2 (Fig. 3). The sites did not trend in any meaningful way in relation to axis 3. Axis 1 showed how sites were different initially, and the successional vectors of the sites moving to the negative end of the axis except for a single site (BT9906) (Fig. 4).

Nonmetric multidimensional scaling (NMS) ordination of all vegetation zones (low prairie, wet meadow, shallow marsh, and deep marsh) showing axes 1 and 2. Each pair of points represents one reference condition wetland site. The arrow connecting the two points represents the change over time. PERMANOVA indicated that past and present reference wetland plant communities were distinct (p < 0.001). The floristic quality index (FQI) for the native species correlated with the positive or negative end of the axes is shown (see Table 1 for species’ correlations with the axes)

Nonmetric multidimensional scaling (NMS) ordination of all vegetation zones (low prairie, wet meadow, shallow marsh, and deep marsh) showing axes 1 and 3. Each pair of points represents one reference condition wetland site. The arrow connecting the two points represents the change over time. PERMANOVA indicated that past and present reference wetland plant communities were distinct (p < 0.001). The floristic quality index (FQI) for the native species correlated with the positive or negative end of the axes is shown (see Table 1 for species’ correlations with the axes)
Species positively correlated with axis 1 had a higher FQI (16.7) in comparison to those correlated with the negative end of axis 1 (FQI = 12.6) (Table 1). The only site that did not have a successional vector moving towards the negative end of axis 1 was BT9906. The movement of 11 of the 12 sites can be interpreted using the r values from the simple regression correlation coefficients which is based on species abundance (aerial coverage) related to values for axes 1, 2, and 3 (Table 1). Here, the present sites are moving from the positive end of axis 1 which is related to a high abundance of conservative native species (positive r values) towards the negative end with a higher abundance of three non-native species (negative r values), three native species with a C-value of 2 or lower, and four native species with a C-value of four and higher. The movement to a lower abundance of species with a C-value of 10 also signals that present wetlands are moving to a plant community with a low abundance of species with the highest conservative value. Species positively correlated with axis 2 had a higher FQI (30.1), when compared to the negative end of the axis (FQI = 8.5) (Table 1). The movement of all successional vectors towards the negative end of axis 2 (Fig. 3) can be interpreted as present sites moving away from a plant community with a high abundance of 42 native species (positive r values) towards a plant community with a higher abundance of two native species and one non-native species (Melilotus officinalis) (negative r values).
Overall, the average C-values significantly decreased at all 12 sites from past to present (P < 0.001) (Table 2). A decline in average C-value over time indicates either a loss of high C-value species or an increase of native species with a C-value < 3. The FQI scores from past to present declined significantly (P = 0.001) (Table 3), paralleling the C-value results, with the exception of a single wetland (Table 3).
Discussion
The reference wetland plant community analyses showed there was an almost universal directional trend over the course of 10 or more years in the Missouri Coteau sub-ecoregion of the PPR. This trend was characterized by the present plant community moving towards lower abundance of a variety of species with high conservative value with an increase in the abundance of both non-native species and a few native species. The decline in species with high conservative value is supported by the decline in average C-values of all present sites and the lower FQI scores observed at 11 of 12 sites.
The PPR, in comparison to other wetland regions in the world, is considered to have wetlands relatively unimpacted by anthropogenic factors (Otte et al., 2021; Smith et al., 1995). The lack of anthropogenic impact means the PPR will have wetlands that fit the definitions of natural, undisturbed, or unimpacted wetlands, which can then serve as reference wetlands for assessment and monitoring protocols. If one assumes these PPR reference wetlands are still representative of a historic baseline, then one would assume there would be no apparent directional change over time because an obvious directional change should be less likely in an unimpacted system. However, the directional trend of reference wetlands observed in this study challenges this assumption. Recent research suggests the majority of temporary and seasonal wetland plant communities on USFWS lands are dominated by non-native invasive species throughout the southern PPR and that native species have declined (Jones, 2021). The increase of non-native species and decrease of conservative species in these reference wetlands suggests a situation wherein the Missouri Coteau sub-ecoregion of the PPR may be undergoing a departure from historic conditions. The uniformity of change indicates that all sites are being subjected to the same pressures acting at a regional level, because local pressures would result in subsets of sites trending in certain directions, appearing more random when observed at a regional scale. Changes in local hydrologic conditions due to increased surface or subsurface drainage (Tangen & Finocchiaro, 2017) would be only experienced by a subset of the reference wetlands and would not likely occur across all of the different reference wetlands.
A potential explanation for the regional change observed in this study is that the PPR is in a period of ecosystem homogenization (McLean et al., 2022a, 2022b). McLean et al. (2022b) developed a conceptual model that suggests a period of substantially increased precipitation increased water levels and resulted in decreased fluctuation of water levels can drive homogenization of PPR wetlands, even in anthropogenically unimpacted sites. According to McLean et al. (2022a), a hydroclimatic period of increased precipitation in the northern section of the PPR started in 1993 and was responsible for wetlands becoming homogenous. In this study, the past sampling period was near the beginning of this period of homogenization while the present sampling period was well into the homogenous period. McLean et al. (2022b) suggest that continued high water levels which result in relatively little water fluctuation would result in non-native species replacing native species and a loss of native conservative species. The homogenization process due to high water levels would support the directional change observed in this current study of reference wetlands in the Missouri Coteau sub-ecoregion of the PPR.
There are several points that would contradict homogenization as an explanation for the directional change. The first is the Palmer hydrologic drought index showed that sampling occurred during dryer periods which is not consistent with the contention that water levels were consistently higher. However, there may have been local conditions that produced continued high water levels. Because the wetlands water levels were not monitored over time, there is no way to tell if water levels were consistently high. Another point is that the majority of wetlands sampled were temporary and seasonal which by definition should have dried out at some point during the typical growing season, unless these wetlands underwent a shift to semi-permanent (which was not evident during sampling). The following alternative explanations which allow for varied water levels would not be subject to these contradictory points as homogenization.
An alternative explanation for the observed directional change is that it could be attributed to yearly natural variation inherent in this system. Comparisons of C-values and FQI values of the current study with those from Euliss and Mushet (2011), wherein C-values and FQI scores were tracked from 2007 to 2010 on 15 wetlands, found that the reported average difference between high and low values for individual wetlands were lower (0.36) than the average difference between past and present values reported in this study (− 0.59). The FQI acted in a similar way with this study having a higher average change of − 7.33 versus 6.24 for the Euliss and Mushet (2011) study. Again, the higher level of change over time for C-values and FQI in this study supports our interpretation that the directional change is higher than intrinsic year to year variability within PPR wetlands.
Another alternative view of the directional change is that it is not signaling a break from historical conditions because PPR wetlands are dynamic systems and, as such, a directional change in the vegetation could reverse over time (Mushet et al., 2020, 2022). Mushet et al. (2020) posit there could be a period in the future when PPR reference wetlands would return to historical conditions. The ability to cycle back to historic conditions is due to the PPR being an inherently unstable system that over a decadal timeline, driven by oscillations in precipitation levels, individual wetlands will go through a series of alternative stable states, and at some point, the individual wetland will revert back to a state similar to the historical conditions documented by past monitoring. This ability to revert to prior conditions is dependent on the surrounding landscape remaining a freshwater ecosystem mosaic (Mushet et al., 2019a). This mosaic will provide a reservoir of diversity at the landscape level (Mushet et al., 2019b) and, as hydraulic conditions and other abiotic and biotic factors cycle over time, these individual reference sites will again resemble past historic monitoring. The reference sites chosen for this study are imbedded in landscapes that would qualify as freshwater mosaics with intact native prairie and a variety of other wetlands types present, combined with a stable land use management that promotes the continued exitance of the mosaic. Thus, following Mushet et al. (2019a), the mosaic conditions were present for these reference wetlands to revert at some point in the future to a condition similar to what was observed during historic monitoring efforts.
Conclusions
This study found the assumption that reference wetlands in the PPR will not experience directional change over time is not a sound assumption, despite the widespread belief that there are areas of the PPR with low anthropogenic impact. Even in this supposedly unimpacted system, there are pressures from factors already present which will likely increase in the future, such as the continued expansion of non-native species and hydroclimatic changes. Whether these reference wetlands will remain in the condition as sampled in 2016 or revert to historic conditions in the future is unknown at this point in time. If the surrounding landscape is preserved as a freshwater ecosystem mosaic, then there is a pathway for the reference wetlands to change to an alternative stable state similar to those observed during monitoring in the past. In contrast, if the surrounding landscapes continue to lose conservative native species due to non-native plant species invasions, the ability to revert to past conditions may be lost. Thus, continued documentation of changes in reference wetlands will be necessary so scientists and managers can plan for the reversion of reference wetlands towards historic baseline conditions or plan for how departures from historical baselines will affect future management. Researchers and managers should consider the potential future scenarios and consider which baseline is most appropriate to guide future management actions and goals (Kopf et al., 2015).
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Anderson, M. J., Gorley, R. N., & Clarke, R. K. (2008). PERMANOVA+ for PRIMER: Guide to software and statistical methods (p. 214). Auckland, New Zealand:PRIMER-e.
Brinson, M. M., & Rheinhardt, R. (1996). The role of reference wetlands in functional assessment and mitigation. Ecological Applications, 6, 69–76. https://doi.org/10.2307/2269553
Bryce, S.A., Omernik, J. M., Pater, D. A., Ulmer, M., Schaar, J., Freeouf, J., Johnson, R., Kuck, P., & Azevedo, S. H. (1996). Ecoregions of North Dakota and South Dakota. Retrieved April 15, 2016, from http://ecologicalregions.info/htm/ndsd_eco.htm
DeKeyser, E. S., Kirby, D. R., & Ell, M. J. (2003). An index of plant community integrity: Development of the methodology for assessing prairie wetland plant communities. Ecological Indicators, 3, 119–133. https://doi.org/10.1016/S1470-160X(03)00015-3
DeKeyser, E. S. (2000). A vegetative classification of seasonal and temporary wetlands across a disturbance gradient using a multimetric approach (p. 124). Fargo, North Dakota: PhD dissertation, North Dakota State University.
Dixon, C., Vacek, S., & Grant, T. (2019). Evolving management paradigms on U.S. Fish and Wildlife Service Lands in the Prairie Pothole Region. Rangelands, 41, 36–43.
Doherty, K. E., Howerter, D. W., Devries, J. H., & Walker, J. (2018). Prairie Pothole Region of North America. In: Finlayson, C. M., Milton, G. R., Prentice, R. C., & Davidson, N. C. (Eds.) The wetland book (pp. 679–688). Springer. https://doi.org/10.1007/978-94-007-4001-3_15.
Euliss, N. H., Jr., & Mushet, D. M. (2011). A multi-year comparison of IPCI scores for prairie pothole wetlands: Implications of temporal and spatial variation. Wetlands, 31(4), 713–723.
Freyman, W. A., Masters, L. A., & Packard, S. (2016). The Universal Floristic Quality Assessment (FQA) calculator: An online tool for ecological assessment and monitoring. Methods in Ecology and Evolution, 7, 380–383.
Gallaway, S., Davis, C., Dvorett, D., & Tramell, B. (2020). Validation of the Oklahoma Rapid Assessment Method (OKRAM) in depressional wetlands using EPA’s three-tiered framework. Wetlands, 40, 925–937.
Gong, P., Niu, Z. G., Cheng, X., Zhao, K. Y., Zhou, D. M., Guo, J. H., Liang, L., Wang, X. F., Li, D. D., Huang, H. B., Wang, Y., Wang, K., Li, W. N., Wang, X. W., Ying, Q., Yang, Z. Z., Ye, Y. F., Li, Z., Zhuang, D. F., … & Yan, J. (2010). China’s wetland change (1990–2000) determined by remote sensing. Science China Earth Science, 53, 1036–1042. https://doi.org/10.1007/s11430-010-4002-3
Hargiss, C. L. M., DeKeyser, E. S., Kirby, D. R., & Ell, M. J. (2008). Regional assessment of wetland plant communities using the index of plant community integrity. Ecological Indicators, 8, 303–307. https://doi.org/10.1016/j.ecolind.2007.03.003
Hargiss, C. L. M., DeKeyser, E. S., Norland, J. E., & Ell, M. J. (2017). Comparing tiers of a multi-tiered wetland assessment in the Prairie Pothole Region. Wetland Ecology and Management, 25, 639–647. https://doi.org/10.1007/s11273-017-9540-4
Hawkins, C. P., Olson, J. R., & Hill, R. A. (2010). The reference condition: Predicting benchmarks for ecological and water-quality assessments. Journal of the North American Benthological Society, 29, 312–343. https://doi.org/10.1899/09-092.1
Herlihy, A. T., Kentula, M. E., Magee, T. K., Lomnicky, G. A., Nahlik, A. M., & Serenbetz, G. (2019). Striving for consistency in the National Wetland Condition Assessment: Developing a reference condition approach for assessing wetlands at a continental scale. Environmental Monitoring and Assessment, 191, 1–20.
Jones, S. (2021). Assessment of Prairie Pothole conditions and plant community composition on FWS fee-title lands (p. 153). Fargo, North Dakota: M.S. Thesis, North Dakota State University.
Kentula, M. E., Gwin, S. E., & Pierson, S. M. (2004). Tracking changes in wetlands with urbanization: Sixteen years of experience in Portland, Oregon, USA. Wetlands, 24, 734–743. https://doi.org/10.1672/0277-5212(2004)024[0734:TCIWWU]2.0.CO;2
Kopf, R. K., Finlayson, C. M., Humphries, P., Sims, N. C., & Hladyz, S. (2015). Anthropocene baselines: Assessing change and managing biodiversity in human-dominated aquatic ecosystems. BioScience, 65, 798–811.
Mack, J. J. (2006). Landscape as a predictor of wetland condition: An evaluation of the Landscape Development Index (LDI) with a large reference wetland dataset from Ohio. Environmental Monitoring and Assessment, 120, 221–241.
Magee, T. K., Blocksom, K. A., & Fennessy, M. S. (2019). A national-scale vegetation multimetric index (VMMI) as an indicator of wetland condition across the conterminous United States. Environmental Monitoring and Assessment, 191, 1–28.
McCune, B., & Mefford, M. J. (2011). Multivariate analysis of ecological data, PC-ORD, version 6 (p. 300). Corvallis, Oregon: MjM Software.
McLean, K., Mushet, D., & Sweetman, J. (2022b). Climate and land use driven ecosystem homogenization in the Prairie Pothole Region. Water, 14(19), 3106.
McLean, K. I., Mushet, D. M., & Sweetman, J. N. (2022a). Temporal coherence patterns of prairie pothole wetlands indicate the importance of landscape linkages and wetland heterogeneity in maintaining biodiversity. Frontiers in Ecology and Evolution, 10, 897872. https://doi.org/10.3389/fevo.2022.897872
McNellie, M. J., Oliver, I., Dorrough, J., Ferrier, S., Newell, G., & Gibbons, P. (2020). Reference state and benchmark concepts for better biodiversity conservation in contemporary ecosystems. Global Change Biology, 26, 6702–6714.
Moreno-Mateos, D., Power, M. E., Comin, F. A., & Yockteng, R. (2012). Structural and functional loss in restored wetland ecosystems. PLOS Biology, 10, e1001247. https://doi.org/10.1371/journal.pbio.1001247
Mushet, D. M., Euliss, N. H., Jr., & Shaffer, T. L. (2002). Floristic quality assessment of one natural and three restored wetland complexes in North Dakota, USA. Wetlands, 22, 126–138.
Mushet, D. M., Solensky, M. J., & Erickson, S. F. (2019b). Temporal gamma-diversity meets spatial alpha-diversity in dynamically varying ecosystems. Biodiversity and Conservation, 28(7), 1783–1797.
Mushet, D. M., McKenna, O. P., & McLean, K. I. (2020). Alternative stable states in inherently unstable systems. Ecology and Evolution, 10(2), 843–850.
Mushet, D. M., Alexander, L. C., Bennett, M., Schofield, K., Christensen, J. R., Ali, G., ... & Lang, M. W. (2019a). Differing modes of biotic connectivity within freshwater ecosystem mosaics. JAWRA Journal of the American Water Resources Association, 55(2), 307–317.
Mushet, D. M., Euliss Jr, N. H., Rosenberry, D. O., LaBaugh, J. W., Bansal, S., Levy, Z. F., ... & Tangen, B. (2022). Lessons learned from wetlands research at the Cottonwood Lake Study Area, Stutsman County, North Dakota, 1967–2021 (No. 1874). US Geological Survey.
Northern Great Plains Floristic Quality Assessment Panel (NGPFQAP). (2001). Coefficients of conservatism for the vascular flora of the Dakotas and adjacent grasslands (p. 32). Reston, VA: U.S. Geological Survey, Biological Resources Division, Information and Technology Report USGS/BRD/ITR-2001–0001. https://doi.org/10.3133/2002366
Otte, M. L., Fang, W. T., & Jiang, M. (2021). A framework for identifying reference wetland conditions in highly altered landscapes. Wetlands, 41, 1–12.
Paradeis, B. L., DeKeyser, E. S., & Kirby, D. R. (2010). Evaluation of restored and native Prairie Pothole Region plant communities following an environmental gradient. Natural Areas Journal, 30, 294–304. https://doi.org/10.3375/043.030.0305
Peck, J. E. (2016). Multivariate analysis for ecologists: Step-by-step (2nd ed., p. 192). MjM Software.
Smith, R. D., Ammann, A., Bartoldus, C., & Brinson, M. M. (1995). An approach for assessing wetland functions using hydrogeomorphic classification, reference wetlands, and functional indices. Technical Report WRP-DE-9 ( p. 79). U.S. Army Corps of Engineers, Waterways Experiment Station, Wetlands Research Program Washington, DC, USA.
Stein, E. D., Brinson, M., Rains, M. C., Kleindl, W., & Hauer, F. R. (2009). Wetland assessment debate. Wetland Science & Practice, 26, 20–24. https://doi.org/10.1672/055.026.0405
Stewart, R. E., & Kantrud, H. A. (1971). Classification of natural ponds and lakes in the glaciated Prairie Region. US Fish and Wildlife Service Resour Publ, 92, 57.
Stoddard, J. L., Larsen, D. P., Hawkins, C. P., Johnson, R. K., & Norris, R. H. (2006). Setting expectations for the ecological condition of streams: The concept of reference condition. Ecological Applications, 16, 1267–1276. https://doi.org/10.1890/10510761(2006)016[1267:SEFTEC]2.0.CO;2
Tangen, B. A., & Finocchiaro, R. G. (2017). A case study examining the efficacy of drainage setbacks for limiting effects to wetlands in the Prairie Pothole Region, USA. Journal of Fish & Wildlife Management, 8, 513–529.
US EPA. (2016). National Wetland Condition Assessment 2011: A collaborative survey of the Nation’s wetlands. EPA-843-R-15-005 (p. 105). Washington, DC: US Environmental Protection Agency.
Wilson, S. D., & Nilsson, C. (2009). Arctic alpine vegetation change over 20 years. Global Change Biology, 15, 1676–1684. https://doi.org/10.1111/j.1365-2486.2009.01896.x
Zedler, J. B., & Kercher, S. (2005). Wetland resources: Status, trends, ecosystem services, and restorability. Annual Review in Environmental Resources, 30, 39–74.
Acknowledgements
Thank you to Jeremy Hackley and Nikki Ellingson for data entry.
Funding
This research was supported by the North Dakota Department of Environmental Quality and United States Department of Agriculture NIFA Project ND02396.
Author information
Authors and Affiliations
Contributions
Christina Hargiss and Edward DeKeyser wrote and received the grant for the project. Savannah Metz and Edward DeKeyser conducted the field work for the project. Christina Hargiss and Deirdre Prischmann-Voldseth co-advised Savannah Metz, the masters student on the project, and assisted in writing and editing. Savannah Metz wrote the results and discussion sections as part of her masters thesis. Jack Norland conducted statistical analysis and assisted in writing of the manuscript. Christina Hargiss wrote and edited the final manuscript. All authors contributed to final study design and coordination. Savannah Metz and Breanna Kobiela contributed to data analyses. Savannah Metz wrote the first draft of the manuscript and all authors reviewed and edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Hargiss, C.L.M., Metz, S.F., DeKeyser, E.S. et al. Vegetation changes over time in Prairie Pothole Region reference wetlands. Environ Monit Assess 195, 855 (2023). https://doi.org/10.1007/s10661-023-11451-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10661-023-11451-y