
Abstract
Grassland ecosystems are affected by the increasing frequency and intensity of extreme climate events (e.g., droughts). Understanding how grassland ecosystems maintain their functioning, resistance, and resilience under climatic perturbations is a topic of current concern. Resistance is the capacity of an ecosystem to withstand change against extreme climate, while resilience is the ability of an ecosystem to return to its original state after a perturbation. Using the growing season Normalized Difference Vegetation Index (NDVIgs, an index of vegetation growth) and the Standardized Precipitation Evapotranspiration Index (a drought index), we evaluated the response, resistance, and resilience of vegetation to climatic conditions for alpine grassland, grass-dominated steppe, hay meadow, arid steppe, and semi-arid steppe in northern China for the period 1982–2012. The results show that NDVIgs varied significantly across these grasslands, with the highest (lowest) NDVIgs values in alpine grassland (semi-arid steppe). We found increasing trends of greenness in alpine grassland, grass-dominated steppe, and hay meadow, while there were no detectable changes of NDVIgs in arid and semi-arid steppes. NDVIgs decreased with increasing dryness from extreme wet to extreme dry. Alpine and steppe grasslands exhibited higher resistance to and lower resilience after extreme wet, while lower resistance to and higher resilience after extreme dry conditions. No significant differences in resistance and resilience of hay meadow under climatic conditions suggest the stability of this grassland under climatic perturbations. This study concludes that highly resistant grasslands under conditions of water surplus are low resilient, but low resistant ecosystems under conditions of water shortage are highly resilient.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Understanding how plant communities respond to extreme climate events (e.g., heatwaves, periods of heavy rain, and droughts) is a global challenge (Jentsch & Beierkuhnlein, 2008; Seneviratne et al., 2012; Vicente-Serrano et al., 2013). Shifting precipitation patterns are of major concern (Cook et al., 2015). Extreme wet and dry events are becoming more intense and frequent due to climate change (Fischer et al., 2013; Singh et al., 2022; Sreeparvathy & Srinivas, 2022). This applies particularly to continental regions. The increase of extreme climate has recently been documented for China (Chen & Sun, 2021), where extreme climate is likely to become even more pronounced in the near future (Gao et al., 2020; Li et al., 2013; Meng et al., 2020; Zhu et al., 2017).
Extreme climate events are widely acknowledged to have an impact on the dynamics, functioning, and stability of terrestrial ecosystems (Beloiu et al., 2022; Hossain & Li, 2021a; Komatsu et al., 2019; Li et al., 2019). Numerous recent studies have provided convincing evidence that grasslands are particularly vulnerable to extreme dry and wet climatic conditions (Craven et al., 2016; Hossain & Li, 2021b; Liu et al., 2021; Zhang et al., 2017). The functioning and stability of grasslands in many regions, especially alpine, hay meadow, and steppe grasslands (Lu et al., 2019; Lei et al., 2020; Chen et al., 2022; Doležal et al., 2022), are likely to be impacted by the rising frequency and severity of extreme climate events (Bellard et al., 2012; Cardinale et al., 2012; Hossain et al., 2022; Loreau & de Mazancourt, 2013). In a global assessment of drought impacts, grasslands in Central Asia, Northeastern China, and the Mongolian Plateau are considered to represent ecosystems that are most susceptible to water stress (Liu et al., 2021). For large parts of China, there is clear evidence for changes in the frequency and intensity of climate extremes (Chen et al., 2019; Cui et al., 2017), and this is likely to become even more pronounced in the near future (Chen & Sun, 2015; Li et al., 2016, 2020a, b; Sui et al., 2018; Zhu et al., 2018). Specifically, the number of consecutive dry and wet days is predicted to increase in the coming decades (Meng et al., 2021). Considering the emerging threats to ecosystem functioning through climate extremes, our current understanding of how the vegetation of different grassland types is responding to different intensities of extreme climate remains unsatisfactory.
There has been a dispute in ecological studies about the effect of extreme climate on the functioning of ecosystems (Hegerl et al., 2011; Isbell et al., 2015). For instance, some researchers have demonstrated increased grassland productivity during drought (Quan et al., 2020) and wet conditions (Hossain & Li, 2021c; Wilcox et al., 2017). While some other studies reported that the productivity of grasslands was negatively impacted by drought and wet periods (Lei et al., 2020; Padilla et al., 2019). Other studies revealed that grassland performance was unexpectedly steady during extreme climate conditions (Jentsch et al., 2011; Kreyling et al., 2008; Zhang et al., 2019). These divergent findings may be due to variations in the classification and intensity of extreme climate events (Barnes et al., 2016; Isbell et al., 2015; Kreyling et al., 2017; Lei et al., 2020; Li et al., 2021). Instead of using an experimentally controlled climatic condition classification (e.g., Kreyling et al., 2017; Li et al., 2021), the application of a drought index classification, which explains the natural climatic conditions (Isbell et al., 2015) to evaluate the effects of extreme climate on grassland productivity is of great significance given the non-consistent classifications of drought to characterize extreme climate events. This approach will help shed light on how to manage grassland ecosystems sustainably and ensure a steady flow of ecosystem goods and services (Fraser et al., 2015).
Ecosystem stability refers to the capacity of an ecosystem to maintain a balanced and functional state, which is necessary for the steady supply of an ecosystem’s services (Pimm, 1984). No agreement has been reached on whether an ecosystem’s stability is related to its resilience after perturbations, its resistance to disturbances, or both (Bai et al., 2004; Hossain et al., 2022; Isbell et al., 2015). Ecological resistance and resilience are two crucial components that are used to assess and track an ecosystem’s functionality and changes in response to shifting climatic conditions (Biggs et al., 2012). The ability of an ecosystem to tolerate change in a harsh environment is known as ecosystem resistance (Pimm, 1984). Resilience is the ecosystem’s ability to bounce back to its original state after a disturbance (Tilman & Downing, 1994).
Despite extensive research on ecosystem resistance and resilience, earlier attempts yielded varied results (Hossain, 2022; Hossain et al., 2022; Isbell et al., 2015). For instance, many studies have shown that species-rich communities are more resistant to and resilient towards extreme climate than species-poor communities (Craine et al., 2013; Kreyling et al., 2017; Vogel et al., 2012); in contrast, other studies have shown the opposite (Pennekamp et al., 2018; Pfisterer & Schmid, 2002). According to a recent study across three ecoregions (temperate dry steppe, humid temperate, and cold steppe), productive ecosystems exhibit lower resistance to but higher resilience towards dry events and higher resistance to but lower resilience towards wet events (Hossain & Li, 2021c). Although this study (Hossain & Li, 2021c) offered evidence of an inverse pattern of resistance and resilience of vegetation encompassing forests, shrubs, and grasslands, much of the current ambiguity surrounding ecosystem stability may be caused by variability in ecosystem types (e.g., forests, shrubs, and grasslands), differences in grassland types (e.g., alpine and steppe grasslands), and disparities in the spatial and temporal scales of the experiments (Piao et al., 2006; Wang et al., 2021). For example, using the Normalized Difference Vegetation Index (NDVI) values of four grassland types, Tong et al. (2017) reported a higher NDVI value in grass-dominated steppe than that of hay meadow, arid steppe, and semi-arid steppe in Inner Mongolia. Likewise, the spatial heterogeneity of NDVI values was reported by Piao et al. (2006), which showed a higher NDVI value in hay meadows, compared to arid and semi-arid steppes. We may have the best chance to increase our understanding of the resistance and resilience of grassland ecosystems to the predicted rising frequency and magnitude of extreme climate events by applying satellite-based long-term climatic and distinct ecological data (Ding et al., 2020; Hossain & Li, 2021c; Lei et al., 2020; Lu et al., 2019).
Remote sensing data enables the monitoring of ecosystem performance at varying spatial and temporal scales (Hossain et al., 2021; Huete, 2016; Tarantino et al., 2021). The successful utilization of satellite-derived NDVI data has enabled the assessment of vegetation dynamics in response to climatic conditions across various ecosystems (Chu et al., 2019; Cui et al., 2021; Hossain & Li, 2021c; Huang et al., 2021; Xu et al., 2017). Because different grassland types have specific properties, early attempts showed that the relationships between NDVI and climate variability are complex (Bao et al., 2015; Hossain & Li, 2021c; Li et al., 2019; Piao et al., 2011).
Grasslands comprise one-third of China’s land area (Zhou et al., 2014). In particular, grasslands in Inner Mongolia, Gansu, and Qinghai provinces are of great importance because of their high biodiversity and related ecosystem services. Maintaining these grasslands is a substantial contribution towards fulfilling the requirements of the Convention on Biological Diversity (CBD). The large area that is covered by grasslands in these regions and the outstanding economic role (food and fiber production) that grasslands play for the population living in these areas require in-depth knowledge about grassland functioning and threats. Here, we develop a research approach that is based on the current state of knowledge (Chen et al., 2022; Lu et al., 2019; Wang et al., 2019a, 2020) that considers varying degrees of human land use intensity in grasslands, from hay meadows to natural steppe and alpine grasslands, across a broad range of climatic conditions (Hossain et al., 2021; Lei et al., 2020; Wang et al., 2019a).
In this study, using both NDVI and SPEI data, we assessed the functioning and stability of the selected sites within five distinct grassland types (alpine grassland, grass-dominated steppe, hay meadow, arid steppe, and semi-arid steppe) in northern China to extreme climate events over the period 1982–2012. Accordingly, the objectives of this study are (i) to assess the differences and temporal patterns of NDVI of five grassland types, (ii) to examine the responses, resistance, and resilience of grasslands to different types of climatic extremes, and (iii) to conclude on the resistance and resilience of grassland ecosystems to extreme climate events.
Materials and methods
Study area
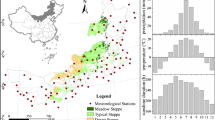
The study area encompasses 5 grassland types (alpine grassland, grass-dominated steppe, hay meadow, arid steppe, and semi-arid steppe), comprising 50 sites (10 sites in each grassland type) in grassland ecosystems in northern China (Fig. 1, Table S1). We selected these study sites from the previously published works (Wang et al., 2019b; Yang et al., 2021) to cover the large gradient in climatic conditions as well as in vegetation structures of Chinese grasslands.

Locations of the study sites in five continental grassland types (alpine grassland, grass-dominated steppe, hay meadow, arid steppe, and semi-arid steppe) in Qinghai, Gansu, and Inner Mongolia (China). Detailed information for site locations, including references, is documented in supplementary information Table S1
The climate in alpine grassland is extremely cold and dry. The mean annual temperature ranges from − 1.8 to 4.2 °C, and annual precipitation varies between 70 and 750 mm, with 90% of annual precipitation occurring from May to September (Zhang et al., 2020). Vegetation in alpine grassland on the Qinghai-Tibet Plateau in northern China is composed of highly productive perennial forbs, compared to grass-dominated meadow and steppes in northern China (Dai et al., 2019). Grasslands in the grass-dominated steppe, hay meadow, semi-arid steppe, and arid steppe are located in central Inner Mongolia in northern China (Hang et al., 2014). Annual precipitation ranges from 300 to 380 mm, and the annual average temperature varies between 0 and 3 °C. Precipitation decreases while temperature increases from the east (hay meadow, grass-dominated steppe) to the west (semi-arid steppe and arid steppe) (Tong et al., 2017). Vegetation in hay meadow and grass-dominated steppe is highly productive, while grasslands in arid and semi-arid steppe are restricted by seasonal climate and less productive than the grass-dominated steppe.
Data sources
In this study, the growing season NDVI of the selected sites in five grassland types and SPEI data were obtained from two sources (Guay et al., 2015 for growing season NDVI and Vicente-Serrano et al., 2010 for SPEI). Over a long time, NDVI has been extensively employed to examine the dynamics, stability, and functioning of ecosystems (Chu et al., 2019; Hossain & Li, 2021c). The Oak Ridge National Laboratory Distributed Active Archive Center provided the growing season NDVI data from 1982 to 2012 used in this investigation (Guay et al., 2015). Growing season (June–August) NDVI data with an 8-km spatial resolution are available in the third-generation Global Inventory Modeling and Mapping Services (GIMMS 3 g) dataset generated from the NOAA Advanced Very High Resolution Radiometer (AVHRR) (Guay et al., 2015). SPEI values indicate the water balance of a particular area based on month-by-month variations in climate since January 1901. SPEI is a site-specific drought index that is employed to depict dry or wet conditions on a global scale. The SPEI dataset was obtained from the SPEIbase v2.5 dataset, which was created based on the CRU 3.24.01 precipitation and potential evapotranspiration to detect and classify climatic conditions (Vicente-Serrano et al., 2010).
Data processing
The growing season NDVI of each grassland type over 31 years (1982–2012) was extracted based on longitude and latitude using statistical software R version 4.0.3 (R Core Team, 2020). We selected point locations in grassland types from the given coordinates of the known grassland types (Wang et al., 2019b; Yang et al., 2021). For example, the coordinates of alpine grassland ranged between 37.48° and 37.76° N and 101.20° and 101.57° E (Yang et al., 2021), and we selected a fixed interval of 0.1° latitude and longitude (e.g., 37.48° N and 101.20° E and 37.58° N and 101.30° E) for extracting the growing season NDVI of 10 sites in this grassland. In addition, for grassland types where the coordinate range is not given, we extracted the growing season NDVI of the given sites. For example, we extracted the growing season NDVI of the given coordinates in the hay meadow for two sites (Yang et al., 2021). For each grassland type, 10 sites were selected (Table S1, Wang et al., 2019b; Yang et al., 2021) and thus the growing season NDVI of 50 sites in 5 grassland types were extracted for this study (Fig. 1). Based on the different types of grasslands, the retrieved growing season NDVI data were grouped.
The normality of growing season NDVI values was tested using both the visual method and a formal statistical test. First, we checked the normality of the dataset by using quantile-quantile (Q-Q) plots for the respective grassland types. We found that data points did not follow the straight diagonal line in the Q-Q plot. Second, as significance test provides more accuracy than visual method, we also tested the normality of the data using the Shapiro-Wilk test and found a p value < 0.05 (Gargano et al., 2022). According to this test, if p > 0.05, then the data is assumed to be normally distributed. We applied the log-transformation (natural logarithms) to normalize the data in the statistical software R (R Core Team, 2020). After the log-transformation, we confirmed the normality of the data using both the Q-Q plot (Fig. S1) and the Shapiro-Wilk test (Table S2; p > 0.05 for all grasslands). The log-transformed growing season NDVI is denoted by NDVIgs, and we used this log-transformed data in the analysis.
Many drought indices, such as the SPEI, Standardized Precipitation Index (SPI), Standardized Terrestrial Water Storage Index (STI), and Palmer Drought Severity Index (PDSI) have been applied to characterize climatic conditions (Cui et al., 2021; Isbell et al., 2015; Wang et al., 2020; Zhang et al., 2012, 2019). Due to its capability to detect water shortages and surpluses over a range of time periods (e.g., 1, 2, 3, …, 48 months), SPEI has been found to be more suitable than other indices (Vicente-Serrano et al., 2012). We extracted a 3-month SPEI value of each August, which represents the wet/dry conditions of the growing season (June–August), as the growing season climate has been proven to be the most important determinant in explaining the functioning of grassland ecosystems (Grant et al., 2017). These 3-month SPEI values represent the different climatic conditions (wet, normal, and dry) during the growing season. Following Isbell et al. (2015), SPEI values were then categorized into five classes (SPEI values for extreme dry: ≤ − 1.28, moderate dry: > − 1.28 to < − 0.67, normal: − 0.67 to < 0.67, moderate wet: 0.67 to < 1.28, and extreme wet: ≥ 1.28). Each class represents a particular intensity of the climatic conditions. This classification is broadly used to detect and quantify the intensity and direction of climatic conditions (Hossain et al., 2021, 2022; Isbell et al., 2015). A value of SPEI-3 ≤ − 1.28 (≥ 1.28) for August 2000 at a specific place corresponds to a degree of seasonal extreme dry (extreme wet) climate that has historically occurred at that site once per decade from June to August. Similarly, SPEI-3 values < − 0.67 (≥ 0.67) for August 2000 at a particular location correspond to a degree of seasonal moderate dry (moderate wet) climate that has happened once every 4 years at that location from June to August (Isbell et al., 2015).
Defining and calculating resistance and resilience
Two crucial components determine the health and stability of an ecosystem (i.e., resistance and resilience, Pimm, 1984). Resistance denotes an ecosystem’s power to tolerate a disturbance (such as droughts), while resilience represents an ecosystem’s capacity and speed of returning to its pre-disturbance state (i.e., normal climatic conditions) after a disturbance. Based on the following equations, NDVIgs resistance and resilience were computed (Hossain et al., 2022; Isbell et al., 2015):
where \(\overline{{Y}_{n}}\) is the NDVIgs values during normal years (i.e., SPEI values are between > − 0.67 and < 0.67) for the period 1982–2012; Ye is the NDVIgs values during an extreme climate event (i.e., SPEI values are between ≥ 0.67 and ≤ − 0.67); and Ye + 1 is the NDVIgs values during a normal year after an extreme climate event.
Because resistance and resilience have no dimensions and are symmetric (i.e., directly comparable between wet and dry climatic conditions), they can be directly compared across grasslands that have varying levels of productivity (i.e., lower or higher NDVIgs values; Hossain & Li, 2021c) as well as across different climatic conditions (Hossain et al., 2022; Isbell et al., 2015). For example, if the NDVIgs during an extreme dry condition decline by 25% and 50% from its normal climatic condition, then Ω = 4 and Ω = 2, respectively. A higher Ω value indicates greater resistance of the vegetation in respective grasslands. Quantifying ∆ necessitates additional information (Ye + 1) than quantifying Ω. In the event of 2 (3) successive extreme climatic conditions (e.g., moderate dry climate in 2010 and extreme dry climate in 2011), we determined the level of ∆ using Ye + 2 (Ye + 3) instead of Ye + 1 in Eq. (ii). If during Ye + 1 NDVIgs value recovers from 0.30 in Ye to 0.50, then ∆ = 1.4. The higher (lower) ∆ values than 1 indicates higher (lower) vegetation resilience.
Data analysis
Assessing the differences and temporal patterns of NDVIgs
The extracted log-transformed NDVIgs values of five grassland types were plotted using boxplots. The differences in the mean NDVIgs among the 5 grassland types were first tested using a one-way ANOVA (Kaufmann & Schering, 2014). Second, as the difference in the mean NDVIgs of the studied grasslands was found statistically significant (p < 0.05), a post-hoc Tukey’s honest significance difference (HSD) test was used to assess the pairwise comparisons of NDVIgs between grassland types (e.g., between alpine grassland and hay meadow and between arid steppe and semi-arid steppe) (Tukey, 1949). Finally, using the “ggplot” package in the statistical software R, temporal patterns in NDVIgs for each type of grassland were displayed (R Core Team, 2020). We applied the Mann-Kendall test (Mann, 1945) to detect statistically significant trends in NDVIgs in respective grassland types over 31 years (Ran et al., 2019). Here, the year was the independent variable and NDVIgs was the dependent variable.
Assessing the correlation between NDVIgs and SPEI
We assessed the autocorrelation of the residuals by the ACF function (Fig. S2) and Durbin-Watson test (Table S3) to confirm that the residuals of the model satisfy the assumption of no autocorrelation (Durbin & Watson, 1992). The Pearson’s correlation (r) was used to analyze the correlation between NDVIgs and SPEI at < 0.05 significance (p) level (Freedman et al., 2007). Here, the dependent variable was NDVIgs and the independent variable was SPEI.
Response of NDVIgs to climatic conditions
As we categorized SPEI values into 5 climatic conditions in order to examine the response of NDVIgs to a particular climate condition, using “ggplot,” we plotted the number of climatic conditions for 31 years in each grassland type. The differences in the mean NDVIgs among five climatic conditions were determined using a one-way ANOVA. When differences in the mean NDVIgs among the five climatic conditions were significant (ANOVA p < 0.05), we compared NDVIgs between climatic conditions using pairwise comparisons. To analyze the pairwise comparisons of NDVIgs between the climatic conditions and between the NDVIgs of a particular climatic condition and their base-mean, a post hoc Tukey’s HSD test was used. Here, base-mean NDVIgs is the mean NDVIgs of all climatic conditions. The pairwise comparisons between the NDVIgs of a climatic condition and the base-mean NDVIgs explain whether the NDVIgs in a given climatic condition is significantly higher, lower, or not significant, compared to the base-mean NDVIgs.
Resistance and resilience of NDVIgs to extreme climate
Like the analysis of NDVIgs and climatic conditions, using a one-way ANOVA and post hoc tests, we also examined the resistance and resilience of all grasslands for four extreme climate events (i.e., extreme dry, moderate dry, moderate wet, and extreme wet). As the resistance and resilience of NDVIgs in hay meadow did not significantly differ (ANOVA p > 0.05) among the four climatic conditions, a post hoc test was not applied for assessing the pairwise comparisons of resistance and resilience between climatic conditions in this grassland type.
Results
Differences and temporal patterns of NDVIgs
The NDVIgs showed obvious differences in five grassland types, and the mean NDVIgs differed significantly among each other (Fig. 2, p < 0.001). The mean NDVIgs was the lowest in semi-arid steppe (0.18) and the highest in alpine grassland (0.42), followed by grass-dominated steppe (0.39), hay meadow (0.25), and arid steppe (0.19) (Fig. 2).

Differences in NDVIgs in five grassland types during the period 1982–2012. One-way ANOVA is used to analyze the differences in mean NDVIgs among the various grassland types. In the post hoc Tukey’s HSD test, different letters (a–e) at the top of boxes indicate statistically significant differences in NDVIgs between grassland types at p < 0.05. The significance of the difference between the NDVIgs of each grassland and the base-mean (mean NDVIgs of five grasslands) is indicated by the asterisk (*) beneath the boxes. The 25th percentile and the 75th percentile are used as the lower and upper quartiles, respectively, to show the interquartile range in boxes. The median (50th percentile) and mean are indicated by the horizontal line and circle close to the center of the box, respectively
The temporal patterns of NDVIgs showed a large variability (Fig. 3). This variability resulted from the differences in site, as we considered 10 sites in each grassland. When all sites in respective grassland assessed together, we found that the mean NDVIgs in alpine grassland, grass-dominated steppe, and hay meadow showed a significant increasing trend (albeit low R value), while the temporal trends of mean NDVIgs in arid and semi-arid steppes were not significant (Fig. 3). The annual increasing rate of mean NDVIgs in alpine grassland was 0.0001 (from 0.420 in 1982 to 0.423 in 2012, p < 0.001), in grass-dominated steppe was 0.0042 (from 0.334 in 1982 to 0.473 in 2012, p < 0.05), and in hay meadow was 0.0039 (from 0.226 in 1982 to 0.339 in 2012, p < 0.001, Fig. 3).

Inter-annual variability in NDVIgs in five grassland types. Solid black lines represent the trends of NDVIgs over the period 1982–2012, detected by the Mann–Kendall test. Confidence intervals of 95% of changes in NDVIgs are represented by bands near the solid lines. The solid points indicate the mean NDVIgs values of all sites in the respective year, and the transparent points represent the site-level NDVIgs values. Asterisks (*, **, and ***) indicate the significance of the changes in NDVIgs at p < 0.05, < 0.01, and < 0.001. “NS” indicates that the trends of inter-annual changes in NDVIgs in arid steppe and semi-arid steppe are not significant
Correlation between NDVIgs and SPEI
Correlation analysis of SPEI and NDVIgs exhibited that NDVIgs in all grassland types decreased with increasing intensity of dry climates (alpine grassland: r = − 0.21, p < 0.001, grass-dominated steppe: r = − 0.43, p < 0.001, hay meadow: r = − 0.28, p < 0.001, arid steppe: r = − 0.49, p < 0.001, and semi-arid steppe: r = − 0.45, p < 0.001) (Fig. 4).

Correlation between SPEI and NDVIgs in five grassland types. The 95% confidence intervals of the correlations between the SPEI and NDVIgs are represented by bands close to the solid lines. The Pearson’s correlation coefficient between SPEI and NDVIgs is displayed with an R and significance level (*** for p < 0.001)
NDVIgs responses to extreme climate events
In order to assess the impact of different climatic conditions on NDVIgs, we categorized the SPEI values into five classes (i.e., extreme dry, moderate dry, normal, moderate wet, and extreme wet). Overall, across all grassland types, every 5 years there was one extreme dry event, of which semi-arid steppe grasslands encountered frequent extreme dry events (every 4 years) and grass-dominated steppe grasslands experienced a lower number of extreme dry events (every 7.4 years) (Fig. 5). Alpine grassland and hay meadow exhibited higher extreme wet events (every 6.2 years), compared with all grassland types (every 7.3 years). For moderate dry climates across all grasslands, a higher number of events were observed in grass-dominated steppe (every 3.1 years) and lower in alpine grassland (every 20 years). Considering moderate wet events across all grassland types, arid and semi-arid steppes experienced a lower number of moderate wet events (every 10 years), while alpine grassland encountered more cases of moderate wet events (every 3.4 years) (Fig. 5).

Observations of climatic conditions (i.e., extreme wet, moderate wet, normal, moderate dry, and extreme dry) in five grassland types over the period 1982–2012
Irrespective of grassland types, wet conditions (moderate and extreme) enhanced NDVIgs, while dry conditions reduced NDVIgs, compared with regular (normal) climatic conditions (Fig. 6). NDVIgs significantly varied among the climatic conditions in respective grassland types (Fig. 6, all p < 0.01). In alpine grassland, compared with the base-mean (i.e., mean NDVIgs for all climatic conditions), extreme dry condition significantly reduced the NDVIgs. Pairwise comparisons revealed that NDVIgs during extreme and moderate wet conditions was significantly higher than that in normal climatic conditions in alpine grassland (Fig. 6a, all p < 0.05). NDVIgs in grass-dominated steppe showed higher sensitivity to a drier climate, as we found NDVIgs during extreme and moderate dry conditions were significantly lower than the base-mean and normal climatic conditions (Fig. 6b, all p < 0.05). In contrast, extreme wet condition had significant positive effects on NDVIgs in comparison with the normal climatic condition and the base-mean (Fig. 6b, all p < 0.001). In hay meadow, NDVIgs in extreme wet condition was significantly higher than NDVIgs in extreme dry, moderate dry, normal climatic conditions and their base-mean (Fig. 6c, all p < 0.001). Neither moderate dry nor extreme dry conditions had significant effects on NDVIgs in hay meadow (Fig. 6c, all p > 0.05). Like grass-dominated steppe, compared with the normal climatic condition and base-mean, significant gains and losses of NDVIgs were observed in extreme wet and extreme dry conditions, respectively, in arid steppe (Fig. 6d, all p < 0.001) and semi-arid steppe (Fig. 6e, all p < 0.001). Notably, none of these two grasslands showed significant changes in NDVIgs between normal and moderate dry climatic conditions (Fig. 6 d and e, all p > 0.05).

The response of NDVIgs to climatic conditions (extreme dry, moderate dry, normal, moderate wet, and extreme wet) in five grassland types: a alpine grassland, b grass-dominated steppe, c hay meadow, d arid steppe, and e semi-arid steppe. The p values indicate the significant differences in NDVIgs between the five intensities of climatic conditions in the specific grassland types. The significance of the difference between the NDVIgs of each climatic condition and the base-mean is indicated by the asterisk (*) beneath the boxes. In the post hoc Tukey’s test, different letters (a–d) at the top of each box denote statistically significant differences in NDVIgs between climatic conditions at p < 0.05. The 25th percentile and the 75th percentile are used as the lower and upper quartiles, respectively, to show the interquartile range in boxes. The median (50th percentile) and mean are indicated by the horizontal line and circle close to the center of the box, respectively. “NS” beneath the boxes indicate the differences between the NDVIgs in a climatic condition and the NDVIgs in all climatic conditions are not significant
Resistance and resilience of grasslands to extreme climate events
Resistance and resilience of five grassland types were assessed for four extreme climate intensities (i.e., extreme dry, moderate dry, moderate wet, and extreme wet) (Figs. 7 and 8). Resistance and resilience of NDVIgs showed large variations across extreme climate intensities, of which alpine grassland and all steppes (grass-dominated, arid, and semi-arid steppes) showed significant differences among four intensities (Figs. 7 and 8, all p < 0.05), while no significant differences among the extreme climate intensities were observed for hay meadow (Figs. 7c and 8c, all p > 0.05).

Vegetation resistance against extreme climate in five grassland types: a alpine grassland, b grass-dominated steppe, c hay meadow, d arid steppe, and e semi-arid steppe. A one-way ANOVA was used to analyze the significance of the differences in the mean grassland NDVIg resistance among the four extreme climate event intensities. In the post hoc Tukey’s HSD test, different letters on the top of boxes indicate significant differences in vegetation resistance between the extreme climate event intensities at p < 0.05. The 25th percentile and the 75th percentile are used as the lower and upper quartiles, respectively, to show the interquartile range in boxes. The median (50th percentile) and mean are indicated by the horizontal line and circle close to the center of the box, respectively

Vegetation resilience towards extreme climate events in five grassland types: a alpine grassland, b grass-dominated steppe, c hay meadow, d arid steppe, and e semi-arid steppe. The statistical details of the figure are the same as those in Fig. 7
Alpine grassland and grass-dominated steppe showed higher (lower) resistance against extreme wet (dry) climate (Fig. 7a and b), while lower (higher) resilience towards extreme wet (dry) climate (Fig. 8a and b). In arid and semi-arid steppes, vegetation showed higher resistance against moderate dry climate, but resistance decreased with increasing drought intensity (i.e., from moderate to extreme dry conditions) in these grasslands (Fig. 7d and e). Conversely, vegetation in these two grassland types showed higher resilience towards increasing drought intensity from moderate to extreme dry climate (Fig. 8d and e). Vegetation resistance and resilience of arid and semi-arid steppes increased with increasing intensity of wet climate (i.e., from moderate to extreme wet) (Fig. 8d and e). No detectable patterns were observed for the resistance and resilience of hay meadow across extreme climate event intensities (Figs. 7c and 8c).
Discussion
Satellite-derived NDVI, a proxy of vegetation vitality, is utilized to assess the functioning and stability of ecosystems under climate stress. In this study, using the SPEI and NDVIgs, grassland response, resistance, and resilience were examined for alpine grassland, grass-dominated steppe, hay meadow, and arid and semi-arid steppes in northern China for the period 1982–2012. The study findings are important to assess the future development of grassland ecosystems and the implementation of nature-based adaptation measures (Beierkuhnlein, 2021).
NDVIgs trends and its relationships with SPEI
For several grassland types, the mean NDVIgs values exhibited substantial differences. Alpine grassland showed a maximum NDVIgs value of 0.42, whereas semi-arid steppe featured a minimum NDVIgs value of 0.18 (Fig. 2). The differences of the mean NDVIgs showed an increasing order of semi-arid steppe (0.18) > arid steppe (0.19) > hay meadow (0.25) > grass-dominated steppe (0.39) > alpine grassland (0.42). The spatial variation of the mean NDVIgs for different grassland types was consistent with those of previous studies (Liu et al., 2017; Piao et al., 2006; Tong et al., 2017).
The results indicate that despite the large inter-annual variability in NDVIgs in alpine grassland, grass-dominated steppe, and hay meadow, a slight greening tendency has been observed in these three grasslands during the investigation period (Fig. 3). This finding is consistent with Shen et al. (2015), which reported that the NDVIgs in the Tibetan Plateau grassland increased over the period 1982–2010. This seems to be a general trend for East Asian grasslands as also other studies across different grassland types in northern China reported an increasing trend of NDVI (alpine grasslands: Piao et al., 2006, Inner Mongolian grasslands: Lu et al., 2019). Enhanced growing season precipitation and air temperature may account for the increasing NDVIgs in productive grassland types (i.e., alpine grassland, grass-dominated steppe, and hay meadow) (Sun et al., 2021). Favorable soil moisture and temperature in the early and peak growing seasons promote plant growth (Gang et al., 2014; Sun et al., 2021). Increased temperature can boost soil nutrients by exacerbating the decomposition and mineralization of litter and roots (Melillo 2002).
However, we did not find any major changes in the greenness or brownness of the vegetation in arid and semi-arid steppes. This fact confirms the findings of Piao et al. (2011), who indicated that the NDVIgs variations for the period 1997–2006 in the Eurasian steppe grasslands were not significant. Very likely, this static behavior in the face of warming is related to the ongoing shortage of moisture in these ecosystems. Additionally, the significant decline of the NDVIgs with increasing intensity of drought (i.e., decreasing SPEI values) across grassland types (Fig. 4) suggests that, as expected, drought events with deficient soil moisture and high potential evapotranspiration set limits to vegetation functioning (tissue die-back, reduced enzymatic activity, impaired photosynthesis) and plant recruitment (Zhong et al., 2010; Craine et al., 2013; Backhaus et al., 2014; Wang et al., 2019a).
Response of grasslands to climatic conditions
Compared with a normal climate, the significant reduction of the NDVIgs during the extreme dry climate in grass-dominated, arid and semi-arid steppes indicates that excessive stress declines vegetation productivity as expected. The reduced NDVIgs during the extreme dry climate can be explained by the weakening of plant photosynthesis caused by a decrease in soil water availability and an increase in evapotranspiration (De Boeck et al., 2011), which leads to an increase in mortality of plants and a decrease in germination of seeds (Chuai et al., 2013; Piao et al., 2014; Zhao et al., 2020).
It is expected in a water-limited environment that the increased NDVIgs during extreme wet condition in all grassland types indicates that precipitation is a key determinant of vegetation growth (Chu et al., 2019). The lower (higher) NDVIgs in extreme dry (extreme wet) conditions, compared to other climatic conditions in our study suggest the translocation of resources to roots in periods of drought and to shoots when other belowground resources are plentiful (Hossain & Li, 2021b).
However, the insignificant differences between the NDVIgs of normal and extreme dry climatic conditions in alpine grassland and hay meadow (Fig. 6a and c) indicate possible complementarity effects among dominant and sub-ordinate species and functional groups. No effect of moderate dry climates on grassland NDVIgs of all grassland types, except grass-dominated steppe suggests that vegetation in these grasslands is able to cope with a certain degree of drought (Hossain & Li, 2020; Li et al., 2019). Our study confirmed that the intensity of the drought is clearly correlated with the impact on grassland ecosystems, which is coherent with previous findings in other grassland ecosystems (Ciais et al., 2005; Hossain et al., 2022), and Southwest China (Li et al., 2019).
Resistance and resilience of grasslands to extreme climate events
Resistance and resilience of the five studied grasslands showed clearly specific patterns in their responses to extreme climate events. For instance, alpine grassland and grass-dominated steppe featured lower resilience but higher resistance to extreme wet climates and higher resilience but lower resistance to extreme dry climates, compared to other climatic conditions. Conversely, arid and semi-arid steppes showed higher resistance (resilience) but lower resilience (resistance) to moderate (extreme) dry climates (Figs. 7 and 8). Hay meadows proved to be stable under climatic fluctuations, as no significant differences in their resistance and resilience were observed among the four climatic conditions. This illustrates that differences in grassland vegetation structure and management are translating into specific responses to climate change.
Ecosystems adapt and respond to climatic fluctuations differently, depending on their species richness (Hossain et al., 2022) and their functional traits (Jentsch et al., 2011). Species richness has been reported as the key determinant in ecosystem resistance (Hossain et al., 2022) and resilience (Vogel et al., 2012) under climatic perturbations. In this study, the apparent lower resistance of vegetation in alpine grassland and grass-dominated steppe against extreme dry climatic conditions could imply that drought-induced stress exerts resource competition among species (Pennekamp et al., 2018). This could be related to the specific sensitivity of certain plant functional types (e.g., perennial grasses that are lacking substantial storage organs below-ground or woody organs) towards short-term climatic extremes. Since alpine grassland and grass-dominated steppe have higher species richness (4–26 species/m−2 in alpine grassland and 2–28 species/m−2 in grass-dominated steppe; Li et al., 2020a, b), compared to the arid and semi-arid steppes (2–11 species/m−2; Li et al., 2020a, b), it is expected that vegetation resistance in alpine grassland and grass-dominated steppe is not affected under moderate dry climate, but that intense drought (i.e., extreme dry) reduced resistance of these grasslands, which is consistent with our findings (Fig. 7a and b).
Examination of the resilience of these two (alpine and grass-dominated) grasslands showed opposite patterns, that is, higher resilience towards dry climatic conditions. This is in line with Isbell et al. (2015) but other studies showed reduced resilience of ecosystems towards dry climatic conditions because of higher resource competition after drought and drought memory effects (De Keersmaecker et al., 2016; Liu et al., 2018).
The apparent discrepancy in the resilience of alpine grassland and grass-dominated steppe might be attributed to variations in plant communities (Craine et al., 2013), the gradient of species richness (Isbell et al., 2015), the existence of stress-tolerance and stress-sensitive species (Fischer et al., 2013), plant functional groups (Kreyling et al., 2017), and functional types (Hossain & Li, 2021a). It requires further field research to scrutinize in detail which processes and traits are effective. As these grassland types are species-rich (Li et al., 2020a, b), several other mechanisms (e.g., plant-plant, plant-soil, plant-environment, and plant-soil-environment interactions) can play a role in strengthening resilience towards dry climatic conditions. For instance, after dry conditions, plants in these two grasslands showed a complementarity effect between dominant and subordinate species, between shallow- and deep-rooted species, and between species belonging to different functional groups.
Grasslands in arid and semi-arid steppes have developed adaptive approaches to deal with drought-induced stresses (Volder et al., 2010). However, severe drought can weaken the capacity of the ecosystem to absorb the water-induced stress. This is evident in our study for the examination of the resistance of arid and semi-arid steppes against moderate and extreme dry climatic conditions. The observed higher resistance of vegetation in these two grasslands against moderate dry climate highlights that plants in arid and semi-arid grasslands can absorb shocks of a certain degree by stimulating fine roots, but a prolonged drought has a severe impact on their functioning due to higher evapotranspiration and lower photosynthesis (De Boeck et al., 2011). In severe water-stress conditions, species show higher resource competition belowground, while in normal climatic conditions plants allocate more resources to shoot (Roy et al., 2016), which is evident in our arid and semi-arid steppes (i.e., lower resistance to but higher resilience towards extreme dry climatic conditions).
Given the increasing frequency and intensity of climate extremes, the sustainable management of grassland ecosystems is crucial for sustaining ecosystem health and human well-being. The resistance and resilience of hay meadows did not differ significantly among the four climatic conditions, which indicate the stability of this grassland type under perturbations. The stability of hay meadows in response to dry and wet climatic conditions suggests that these grasslands are less sensitive to short-term water shortage and logging, compared to the other four grassland types evaluated. This stability can be a reflection of the fact that plants in these meadows are adapted to loose above-ground biomass through regular mowing (Wang et al., 2022; Zhou et al., 2019). Complementary responses of both important functional groups (grasses and forbs) were observed that supported the stability of the entire ecosystems. For instance, grasses responded negatively to precipitation but positively to temperature, and forbs responded positively to climatic variability (Wang et al., 2022). In these ecosystems, grasses responded less sensitively to precipitation and were very drought-resistant (Ma et al., 2017). Moreover, forbs in hay meadows showed adaptive strategies in relation to other functional groups under resource-constrained conditions (Zhou et al., 2019).
The observed lower resistance and higher resilience of alpine grassland and grass-dominated steppe, but higher resistance and lower resilience of arid and semi-arid steppes confirm that vegetation resistance and resilience of these grasslands are inversely correlated. This supports the notion that strongly resistant ecosystems can exhibit low resilience, while less resistant ecosystems can feature high resilience. Comparable results were yielded in grassland studies on other continents (van Ruijven & Berendse, 2010; Vogel et al., 2012; Kreyling et al., 2017; Pennekamp et al., 2018). The stability of hay meadows highlights that these grasslands are less vulnerable to environmental changes in comparison with other grassland types. This can also result from the mowing regime, that is selecting species can cope with disturbances. Given the expected increases in magnitude and duration of extreme climate events in the future, our study may help to identify grasslands that are most susceptible to losing ecosystem functions. To mitigate the impacts of climate change on these vulnerable grasslands in northern China, adapted management and conservation strategies are imperative.
Conclusion
To gain deeper insight into the long-term functioning and stability of grassland ecosystems, we conducted an evaluation of differences and temporal patterns of NDVI and their resistance to and resilience towards climate extremes across five grassland types in northern China. We draw four broad insights from our study, which are as follows:
-
The productive grasslands (i.e., alpine grassland, grass-dominated steppe, and hay meadow) showed a positive trend towards increased greenness over the 31 years.
-
Water surplus led to significant gains in NDVIgs, while water shortage resulted in significant losses in NDVIgs across all grasslands.
-
Alpine grassland and grass-dominated steppe showed lower resistance but higher resilience towards dry climatic conditions.
-
Arid and semi-arid steppes exhibited higher resistance but lower resilience towards moderate dry conditions.
We conclude that plants in arid and semi-arid steppes are highly resistant to moderate dry climatic condition. However, as the intensity of dryness increases, the capacity of plants in these grasslands to absorb excessive stress decreases, resulting in reduced resistance against extreme dry climatic condition. Alpine grassland and grass-dominated steppe showed a decline in resistance and an increase in resilience from extreme wet to extreme dry climatic conditions. Thus, we conclude that because of their high productivity (reflected from higher NDVIgs values), alpine grassland and grass-dominated steppe are highly drought sensitive but have ability to recover faster after extreme dry climatic condition.
This study’s findings imply that highly resistant ecosystems during water surplus conditions are low resilient, while low resistant ecosystems during water shortage conditions are highly resilient. Our study advances the intriguing possibility that resilience in function, rather than resistance controls the stability of productive grasslands. Our research may assist land managers in identifying grasslands that are vulnerable to losing ecosystem functions due to projected extreme climate events. Knowledge of this study provides important information to the decision-makers that the grasslands in Gansu, Qinghai and Inner Mongolia require adapted management and conservation efforts for achieving stable delivery of ecosystem goods and services.
Data availability
The datasets generated and/or analyzed during the current study are available in Oak Ridge National Laboratory Distributed Active Archive Center (Guay et al., 2015; (https://daac.ornl.gov/VEGETATION/guides/GIMMS3g_NDVI_Trends.html) and SPEI SPEIbase v2.5 (Vicente-Serrano et al., 2010; http://spei.csic.es/database.html).
References
Backhaus, S., Kreyling, J., Grant, K., Beierkuhnlein, C., Walter, J., & Jentsch, A. (2014). Recurrent mild drought events increase resistance toward extreme drought stress. Ecosystems, 17(6), 1068–1081. https://www.jstor.org/stable/43677656
Bai, Y., Han, X., Wu, J., Chen, Z., & Li, L. (2004). Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature, 431, 181–184. https://doi.org/10.1038/nature02850
Bao, G., Bao, Y. H., Sanjjava, A., Qin, Z. H., Zhou, Y., & Xu, G. (2015). NDVI-indicated long-term vegetation dynamics in Mongolia and their response to climate change at biome scale. International Journal of Climatology, 35, 4293–4306. https://doi.org/10.1002/joc.4286
Barnes, M. L., Moran, M. S., Scott, R. L., Kolb, T. E., Ponce-Campos, G. E., Moore, D. J. P., Ross, M. A., Mitra, B., & Dore, S. (2016). Vegetation productivity responds to sub-annual climate conditions across semiarid biomes. Ecosphere, 7(5), e01339. https://doi.org/10.1002/ecs2.1339
Beierkuhnlein, C. (2021). Nature-based solutions must be realized - not just proclaimed - in face of climatic extremes. Erdkunde, 75(3), 225–244. https://doi.org/10.3112/erdkunde.2021.03.06
Bellard, C., Bertelsmeier, C., Leadley, P., Thuiller, W., & Courchamp, F. (2012). Impact of climate change on the future of biodiversity. Ecology Letters, 15, 365–377. https://doi.org/10.1111/j.1461-0248.2011.01736.x
Beloiu, M., Stahlmann, R., & Beierkuhnlein, C. (2022). Drought impacts in forest canopy and deciduous tree saplings in Central European forests. Forest Ecology and Management, 509, 120075. https://doi.org/10.1016/j.foreco.2022.120075
Biggs, R., Schluter, M., Biggs, D., Bohensky, E. L., Silver, S. B., Cundill, G., Dakos, G., Daw, T. M., Evans, L. S., Kotschy, K., Leitch, A. M., Meek, C., Quinlan, A., Raudsepp-Hearne, C., Robards, M. D., Schoon, M. L., Schultz, L., & West, P. C. (2012). Toward principles for enhancing the resilience of ecosystem services. Annual Review of Environment and Resources, 37, 421–448. https://doi.org/10.1146/annurev-environ-051211-123836
Cardinale, B. J., Duffy, J. E., Gonzalez, A., Hooper, D. U., Perrings, C., Venail, P., Narwani, A., Mace, G. M., Tilman, D., Wardle, D. A., & Kinzig, A. P. (2012). Biodiversity loss and its impact on humanity. Nature, 486(7401), 59–67. https://doi.org/10.1038/nature11148
Chen, H., & Sun, J. (2015). Changes in climate extreme events in China associated with warming. International Journal of Climatology, 135, 2735–2751. https://doi.org/10.1002/joc.4168
Chen, H. P., & Sun, J. Q. (2021). Anthropogenic influence has increased climate extreme occurrence over China. Sci Bull, 66(8), 749–752. https://doi.org/10.1016/j.scib.2020.12.001
Chen, L. T., Chen, X. H., Cheng, L. Y., Zhou, P., & Liu, Z. Y. (2019). Compound hot droughts over China: Identification, risk patterns and variations. Atmospheric Research, 227, 210–219. https://doi.org/10.1016/j.atmosres.2019.05.009
Chen, J., Dong, G., Chen, J., Jiang, S., Qu, L., Legesse, T. G., Zhao, F., Tong, Q., Shao, C., & Han, X. (2022). Energy balance and partitioning over grasslands on the Mongolian Plateau. Ecological Indicators, 135, 108560. https://doi.org/10.1016/j.ecolind.2022.108560
Chu, H., Venevsky, S., Wu, C., & Wang, M. (2019). NDVI-based vegetation dynamics and its response to climate changes at Amur-Heilongjiang River Basin from 1982 to 2015. Science of the Total Environment, 650, 2051–2062. https://doi.org/10.1016/j.scitotenv.2018.09.115
Chuai, X., Huang, X., Wang, W., & Bao, G. (2013). NDVI, temperature and precipitation changes and their relationships with different vegetation types during 1998–2007 in Inner Mongolia, China. International Journal of Climatology, 33, 1696–1706. https://doi.org/10.1002/joc.3543
Ciais, P., Reichstein, M., Viovy, N., Granier, A., Og´ee, J., Allard, V., Aubinet, M., Buchmann, N., Bernhofer, C., Carrara, A., Chevallier, F., De Noblet, N., Friend, A. D., Friedlingstein, P., Grünwald, T., Heinesch, B., Keronen, P., Knohl, A., Krinner, G., … Valentini, R. (2005). Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature, 437, 529–533. https://doi.org/10.1038/nature03972
Cook, B. I., Ault, T. R., & Smerdon, J. E. (2015). Unprecedented 21st century drought risk in the American Southwest and Central Plains. Science Advances, 1, e1400082. https://doi.org/10.1126/sciadv.1400082
Craine, J. M., Ocheltree, T. W., Nippert, J. B., Towne, E. G., Skibbe, A. M., Kembel, S. W., & Fargione, J. E. (2013). Global diversity of drought tolerance and grassland climate-change resilience. Nature Climate Change, 3, 63–67. https://doi.org/10.1038/nclimate1634
Craven, D., Isbell, F., Manning, P., Connolly, J., Bruelheide, H., Ebeling, A., Roscher, C., van Ruijven, J., Weigelt, A., Wilsey, B., Beierkuhnlein, C., de Luca, E., Griffin, J. N., Hautier, Y., Hector, A., Jentsch, A., Kreyling, J., Lanta, V., Loreau, M., … Eisenhauer, N. (2016). Plant diversity effects on grassland productivity are robust to both nutrient enrichment and drought. Philosophical Transactions of the Royal Society B: Biological Sciences B, 371, 20150277. https://doi.org/10.1098/rstb.2015.0277
Cui, A., Li, J., Zhou, Q., Zhu, R., Liu, H., Wu, G., & Li, Q. (2021). Use of a multiscalar GRACE-based standardized terrestrial water storage index for assessing global hydrological droughts. Journal of Hydrology, 603, 126871. https://doi.org/10.1016/j.jhydrol.2021.126871
Cui, L. L., Shi, J., Du, H. Q., Wen, J. M. (2017). Characteristics and trends of climate extremes in China during 1959–2014. Journal of Tropical Meteorology, 23(4), 368-379. https://www.cnki.net/kcms/doi/10.16555/j.1006-8775.2017.04.003.html. Accessed 12 Jun 2022
Dai, L., Ke, X., Guo, X., Du, Y., Zhang, F., Li, Y., Li, Q., Peng, C., Shu, K., & Cao, G. (2019). Responses of biomass allocation across two vegetation types to climate fluctuations in the northern Qinghai-Tibet Plateau. Ecology and Evolution, 9, 6105–6115. https://doi.org/10.1002/ece3.5194
De Boeck, H. J., Dreesen, F. E., Janssens, I. A., & Nijs, I. (2011). Whole system responses of experimental plant communities to climate extremes imposed in different seasons. New Phytologist, 89(3), 806–817. https://doi.org/10.1111/j.1469-8137.2010.03515.x
De Keersmaecker, W., Van Rooijen, N., Lhermitte, S., Tits, L., Schaminee, J., Coppin, P., Honnay, O., & Somers, B. (2016). Species-rich semi-natural grasslands have a higher resistance but a lower resilience than intensively managed agricultural grasslands in response to climate anomalies. Journal of Applied Ecology, 53(2), 430–439. https://doi.org/10.1111/1365-2664.12595
Ding, Y., Li, Z., & Peng, S. (2020). Global analysis of time-lag and -accumulation effects of climate on vegetation growth. The International Journal of Applied Earth Observation and Geoinformation, 92, 102179. https://doi.org/10.1016/j.jag.2020.102179
Doležal, J., Altman, J., Jandová, V., Chytrý, M., Conti, L., Méndez-Castro, F. E., Klimešová, J., Zelený, D., & Ottaviani, G. (2022). Climate warming and extended droughts drive establishment and growth dynamics in temperate grassland plants. Agricultural and Forest Meteorology, 313, 108762. https://doi.org/10.1016/j.agrformet.2021.108762
Durbin, J., & Watson, G. S. (1992). Testing for serial correlation in least squares regression. II (pp. 260–266). New York: Springer.
Fischer, E. M., Beyerle, U., & Knutti, R. (2013). Robust spatially aggregated projections of climate extremes. Nat Clim Change, 3, 1033–1038. https://doi.org/10.1038/nclimate2051
Fraser, L. H., Pither, J., Jentsch, A., Sternberg, M., Zobel, M., Askarizadeh, D., Bartha, S., Beierkuhnlein, C., Bennett, J., Bittel, A., Boldgiv, B., Boldrini, I., Bork, E., Brown, L., Cabido, M., Cahill, J. F., Carlyle, C. N., Campetella, G., Chelli, S., … Zupo, T. (2015). Worldwide evidence of a unimodal relationship between productivity and plant species richness. Science, 349(6245), 302–305. https://doi.org/10.1126/science.aab3916
Freedman, D., Pisani, R., & Purves, R. (2007). Statistics (international student edition). In R. Pisani, R. Purves (Ed.), 4th edn. New York: WW Norton & Company. p. 720
Gang, C., Zhou, W., Chen, Y., Wang, Z., Sun, Z., Li, J., Qi, J., & Odeh, I. (2014). Quantitative assessment of the contributions of climate change and human activities on global grassland degradation. Environment and Earth Science, 72, 4273–4282. https://doi.org/10.1007/s12665-014-3322-6
Gao, J., Liu, L., & Wu, S. (2020). Hazards of extreme events in China under different global warming targets. Big Earth Data, 4(2), 153–174. https://doi.org/10.1080/20964471.2020.1769254
Gargano, D., Fenu, G., Passalacqua, N. G., Schettino, A., & Bernardo, L. (2022). Relating trait variation to species and community productivity in contrasting Oro-Mediterranean pastures: A 7-years study in the Pollino National Park (S-Italy). Plants, 11(19), 2471. https://doi.org/10.3390/plants11192471
Grant, K., Kreyling, J., Beierkuhnlein, C., & Jentsch, A. (2017). Importance of seasonality for the response of a mesic temperate grassland to increased precipitation variability and warming. Ecosystems, 20(8), 1454–1467. https://doi.org/10.1007/s10021-017-0122-3
Guay, K. C., Beck, P. S. A., & Goetz, S. J. (2015). Long-term arctic growing season NDVI trends from GIMMS 3g, 1982-2012. Data set. Available on-line (http://daac.ornl.gov) from Oak Ridge National Laboratory Distributed Active Archive Center, Oak Ridge, Tennessee, USA. https://doi.org/10.3334/ORNLDAAC/1275. Accessed 18 Jul 2021
Hang, Y. L., Bao, G., Bao, Y. H., & Burenjirigala, A. D. (2014). Spatiotemporal changes of vegetation coverage in Xilingol grassland and its responses to climate change during 2000–2010. Acta Agrestia Sinica, 22(6), 1194–1204.
Hegerl, G., Hanlon, H., & Beierkuhnlein, C. (2011). Elusive Extremes. Nature Geoscience, 4, 142–143. https://doi.org/10.1038/ngeo1090
Hossain, M. L., & Beierkuhnlein, C. (2018). Enhanced aboveground biomass by increased precipitation in a central European grassland. Ecological Processes, 7, 37. https://doi.org/10.1186/s13717-018-0149-1
Hossain, M. L., & Li, J. (2020). Effects of long-term climatic variability and harvest frequency on grassland productivity across five ecoregions. Global Ecology and Conservation, 23, e01154. https://doi.org/10.1016/j.gecco.2020.e01154
Hossain, M. L., & Li, J. (2021). Disentangling the effects of climatic variability and climate extremes on the belowground biomass of C3- and C4-dominated grassland across five ecoregions. Science of the Total Environment, 760, 143894. https://doi.org/10.1016/j.scitotenv.2020.143894
Hossain, M. L., & Li, J. (2021). Biomass partitioning of C3- and C4-dominated grasslands in response to climatic variability and climate extremes. Environmental Research Letters, 16, 074016. https://doi.org/10.1088/1748-9326/ac027a
Hossain, M. L., & Li, J. (2021). NDVI-based vegetation dynamics and its resistance and resilience to different intensities of climatic events. Global Ecology and Conservation, 30, e01768. https://doi.org/10.1016/j.gecco.2021.e01768
Hossain, M. L., Kabir, M. H., Nila, M. U. S., & Rubaiyat, A. (2021). Response of grassland net primary productivity to dry and wet climatic events in four grassland types in Inner Mongolia. Plant-Environ Interact, 2, 250–262. https://doi.org/10.1002/pei3.10064
Hossain, M. L., Li, J., Hoffmann, S., & Beierkuhnlein, C. (2022). Biodiversity showed positive effects on resistance but mixed effects on resilience to climatic extremes in a long-term grassland experiment. Science of the Total Environment, 827, 154322. https://doi.org/10.1016/j.scitotenv.2022.154322
Hossain, M. L. (2022) Grassland ecosystems functioning and stability in response to climatic variability and climate extremes. PhD Dissertation. Hong Kong Baptist University. https://scholars.hkbu.edu.hk/ws/portalfiles/portal/59830046/G22THFL-032373T.pdf. Accessed 9 Feb 2023
Huang, J., Yu, H., Guan, X., Wang, G., & Guo, R. (2016). Accelerated dryland expansion under climate change. Nature Climate Change, 6, 166–171. https://doi.org/10.1038/nclimate2837
Huang, S., Tang, L., Hupy, J. P., Wang, Y., & Shao, G. (2021). A commentary review on the use of normalized difference vegetation index (NDVI) in the era of popular remote sensing. Journal of Forest Research, 32, 1–6. https://doi.org/10.1007/s11676-020-01155-1
Huete, A. (2016). Ecology vegetation’s responses to climate variability. Nature, 531, 181–182. https://doi.org/10.1038/nature17301
Isbell, F., Craven, D., Connolly, J., Loreau, M., Schmid, B., Beierkuhnlein, C., Bezemer, T. M., Bonin, C., Bruelheide, H., de Luca, E., Ebeling, A., Griffin, J. N., Guo, Q., Hautier, Y., Hector, A., Jentsch, A., Kreyling, J., Lanta, V., Manning, P., … Eisenhauer, N. (2015). Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature, 526, 574–577. https://doi.org/10.1038/nature15374
Jentsch, A., & Beierkuhnlein, C. (2008). Research frontiers in climate change: Effects of extreme meteorological events on ecosystems. CR Geoscience, 340, 621–628. https://doi.org/10.1016/j.crte.2008.07.002
Jentsch, A., & White, P. (2019). The theory of pulse dynamics. Ecology, 100(7), e02734. https://doi.org/10.1002/ecy.2734
Jentsch, A., Kreyling, J., Elmer, M., Gellesch, E., Glaser, B., Grant, K., Hein, R., Lara, M., Mirzae, H., Nadler, S. E., Nagy, L., Otieno, D., Pritsch, K., Rascher, U., Schädler, M., Schloter, M., Singh, B. K., Stadler, J., Walter, J., … Beierkuhnlein, C. (2011). Climate extremes initiate ecosystem regulating functions while maintaining productivity. Journal of Ecology, 99, 689–702. https://doi.org/10.1111/j.1365-2745.2011.01817.x
Kaufmann, J., Schering, A. G. (2014). Analysis of variance ANOVA, 1st edn. In N. Balakrishnan, T. Colton, B. Everitt, W. Piegorsch, F. Ruggeri & J. L. Teugels (Eds.) (pp. 1–12). John Wiley & Sons. https://doi.org/10.1002/9781118445112.stat06938
Kendall, M. (1938). A new measure of rank correlation. Biometrika, 30(1–2), 81–89. https://doi.org/10.2307/2332226
Komatsu, K. J., Avolio, M. L., Lemoine, N., Isbell, F., Grman, E., Houseman, G. R., Koerner, S. E., Johnson, D., Wilcox, K. R., Alatalo, J. M., Anderson, J. P., Aerts, R., Baer, S. G., Baldwin, A. H., Bates, J., Beierkuhnlein, C., Belote, R. T., Blair, J., Bloor, J., … Zhang, Y. (2019). Global change effects on plant communities are magnified by time and the number of global change factors imposed. Proceedings of the National Academy of Sciences of the United States of America, 116(36), 17867–17873. https://doi.org/10.1073/pnas.1819027116
Kreyling, J., Beierkuhnlein, C., Elmer, M., Pritsch, K., Radovski, M., Schloter, M., Wollecke, J., & Jentsch, A. (2008). Soil biotic processes remain remarkably stable after 100-year extreme weather events in experimental grassland and heath. Plant and Soil, 308, 175. https://doi.org/10.1007/s11104-008-9617-1
Kreyling, J., Dengler, J., Walter, J., Velev, N., Ugurlu, E., Sopotlieva, D., Ransijn, J., Picon-Cochard, C., Nijs, I., Hernandez, P., Guler, B., von Gillhaussen, P., De Boeck, H. J., Bloor, J. M. G., Berwaers, S., Beierkuhnlein, C., Arfin Khan, M. A. S., Apostolova, I., Altan, Y., … Jentsch, A. (2017). Species richness effects on grassland recovery from drought depend on community productivity in a multisite experiment. Ecology Letters, 20(11), 1405–1413. https://doi.org/10.1111/ele.12848
Lei, T., Feng, J., Lv, J., Wang, J., Song, H., Song, W., & Gao, X. (2020). Net primary productivity loss under different drought levels in different grassland ecosystems. The Journal of Environmental Management, 274, 111144. https://doi.org/10.1016/j.jenvman.2020.111144
Li, J., Zhang, Q., Chen, Y. D., & Singh, V. P. (2013). GCMs-based spatiotemporal evolution of climate extremes during the 21st century in China. Journal of Geophysical Research: Atmospheres, 118(19), 11–017. https://doi.org/10.1002/jgrd.50851
Li, J., Chen, Y. D., Zhang, L., Zhang, Q., & Chiew, F. H. (2016). Future changes in floods and water availability across China: Linkage with changing climate and uncertainties. Journal of Hydrometeorology, 17(4), 1295–1314. https://doi.org/10.1175/JHM-D-15-0074.1
Li, X., Li, Y., Chen, A., Gao, M., Slette, I. J., & Piao, S. (2019). The impact of the 2009/2010 drought on vegetation growth and terrestrial carbon balance in Southwest China. Agricultural and Forest Meteorology, 269–270, 239–248. https://doi.org/10.1016/j.agrformet.2019.01.036
Li, J., Gan, T. Y., Chen, Y. D., Gu, X., Hu, Z., Zhou, Q., & Lai, Y. (2020). Tackling resolution mismatch of precipitation extremes from gridded GCMs and site-scale observations: Implication to assessment and future projection. Atmospheric Research, 239, 104908. https://doi.org/10.1016/j.atmosres.2020.104908
Li, M., Zhang, X., Niu, B., He, Y., Wang, X., & Wu, J. (2020). Changes in plant species richness distribution in Tibetan alpine grasslands under different precipitation scenarios. Global Ecology and Conservation, 21, e00848. https://doi.org/10.1016/j.gecco.2019.e00848
Li, L., Zheng, Z., Biederman, J. A., Qian, R., Ran, Q., Zhang, B., Xu, C., Wang, F., Zhou, S., Che, R., Dong, J., Xu, Z., Cui, X., Hao, Y., & Wang, Y. (2021). Drought and heat wave impacts on grassland carbon cycling across hierarchical levels. Plant, Cell and Environment, 44(7), 2402–2413. https://doi.org/10.1111/pce.13767
Liu, S., Cheng, F., Dong, S., Zhao, H., Hou, X., & Wu, X. (2017). Spatiotemporal dynamics of grassland aboveground biomass on the Qinghai-Tibet Plateau based on validated MODIS NDVI. Science and Reports, 7, 4182. https://doi.org/10.1038/s41598-017-04038-4
Liu, L., Zhang, Y., Wu, S., Li, S., & Qin, D. (2018). Water memory effects and their impacts on global vegetation productivity and resilience. Science and Reports, 8, 2962. https://doi.org/10.1038/s41598-018-21339-4
Liu, H., Lin, L., Wang, H., Zhang, Z., Shangguan, Z., Feng, X., & He, J.-S. (2021). Simulating warmer and drier climate increases root production but decreases root decomposition in an alpine grassland on the Tibetan plateau. Plant and Soil, 458, 59–73. https://doi.org/10.1007/s11104-020-04551-y
Loreau, M., & de Mazancourt, C. (2013). Biodiversity and ecosystem stability: A synthesis of underlying mechanisms. Ecology Letters, 16, 106–115. https://doi.org/10.1111/ele.12073
Lu, Q., Zhao, D., Wu, S., Dai, E., & Gao, J. (2019). Using the NDVI to analyze trends and stability of grassland vegetation cover in Inner Mongolia. Theoretical and Applied Climatology, 135, 1629–1640. https://doi.org/10.1007/s00704-018-2614-2
Ma, Z., Liu, H., Mi, Z., Zhang, Z., Wang, Y., Xu, W., Jiang, L., & He, J. S. (2017). Climate warming reduces the temporal stability of plant community biomass production. Nature Communications, 8(1), 1–7. https://doi.org/10.1038/ncomms15378
Mann, H. B. (1945). Nonparametric tests against trend. Econometrica, 13, 245–259. https://doi.org/10.2307/1907187
McDowell, N., Pockman, W. T., Allen, C. D., Breshears, D. D., Cobb, N., Kolb, T., Plaut, J., Sperry, J., West, A., Williams, D. G., & Yepez, E. A. (2008). Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytologist, 178, 719–739. https://doi.org/10.1111/j.1469-8137.2008.02436.x
Melillo, J. M. (2002). Soil warming and carbon-cycle feedbacks to the climate system. Science, 298(5601), 2173–2176. https://doi.org/10.1126/science.1074153
Meng, C., Xu, Y., Li, Q., Ma, Y., Feng, Q., Ma, W., Pan, J., & Li, K. (2020). Analyses of observed features and future trend of extreme temperature events in Inner Mongolia of China. Theoretical and Applied Climatology, 139(1), 577–597. https://doi.org/10.1007/s00704-019-02969-8
Meng, C., Zhang, L., Gou, P., Huang, Q., Ma, Y., Miao, S., Ma, W., & Xu, Y. (2021). Assessments of future climate extremes in China by using high-resolution PRECIS 2.0 simulations. Theoretical and Applied Climatology, 145, 295–311. https://doi.org/10.1007/s00704-021-03618-9
National Research Council. (1992). Grasslands and grassland sciences in northern China. Washington: The National Academies Press. https://doi.org/10.17226/1942
Padilla, F. M., Mommer, L., de Caluwe, H., Smit-Tiekstra, A. E., Visser, E. J. W., & de Kroon, H. (2019). Effects of extreme rainfall events are independent of plant species richness in an experimental grassland community. Oecologia, 191, 177–190. https://doi.org/10.1007/s00442-019-04476-z
Pennekamp, F., Pontarp, M., Tabi, A., Altermatt, F., Alther, R., Choffat, Y., Fronhofer, E. A., Ganesanandamoorthy, P., Garnier, A., Griffiths, J. I., Greene, S., Horgan, K., Massie, T. M., Machler, E., Palamara, G. M., Seymour, M., & Petchey, O. L. (2018). Biodiversity increases and decreases ecosystem stability. Nature, 563(7729), 109–112. https://doi.org/10.1038/s41586-018-0627-8
Pfisterer, A. B., & Schmid, B. (2002). Diversity-dependent production can decrease the stability of ecosystem functioning. Nature, 416(6876), 84–86. https://doi.org/10.1038/416084a
Piao, S., Mohammat, A., Fang, J., Cai, Q., & Feng, J. (2006). NDVI-based increase in growth of temperate grasslands and its responses to climate changes in China. Global Environmental Change, 16(4), 340–348. https://doi.org/10.1016/j.gloenvcha.2006.02.002
Piao, S., Nan, H., Huntingford, C., Ciais, P., Friedlingstein, P., Sitch, S., Peng, S., Ahlstrom, A., Canadell, J. G., Cong, N., Levis, S., Levy, P. E., Liu, L., Lomas, M. R., Mao, J., Myneni, R. B., Peylin, P., Poulter, B., Shi, X., … Chen, A. (2014). Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nature Communications, 5, 5018. https://doi.org/10.1038/ncomms6018
Piao, S., Wang, X., Ciais, P., Zhu, B., Wang, T., & Liu, J. (2011). Changes in satellite-derived vegetation growth trend in temperate and boreal Eurasia from 1982 to 2006. Global Change Biology, 17, 3228–3239. https://doi.org/10.1111/j.1365-2486.2011.02419.x
Pimm, S. L. (1984). The complexity and stability of ecosystems. Nature, 307, 321–326. https://doi.org/10.1038/307321a0
Quan, Q., Zhang, F., Meng, C., Ma, F., Zhou, Q., Sun, F., & Niu, S. (2020). Shifting biomass allocation determines community water use efficiency under climate warming. Environmental Research Letters, 15, 094041. https://doi.org/10.1088/1748-9326/aba472
R Core Team. (2020). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing. https://www.R-project.org/. Accessed 16 Apr 2021
Ran, Q., Hao, Y., Xia, A., Liu, W., Hu, R., Cui, X., Xue, K., Song, X., Xu, C., Ding, B., & Wang, Y. (2019). Quantitative assessment of the impact of physical and anthropogenic factors on vegetation spatial-temporal variation in Northern Tibet. Remote Sens, 11(10), 1183. https://doi.org/10.3390/rs11101183
Roy, J., Picon-Cochard, C., Augusti, A., Benot, M. L., Thiery, L., Darsonville, O., Landais, D., Piel, C., Defossez, M., Devidal, S., & Escape, C. (2016). Elevated CO2 maintains grassland net carbon uptake under a future heat and drought extreme. Proceedings of the National Academy of Sciences of the United States of America, 113(22), 6224–6229. https://doi.org/10.1073/pnas.1524527113
Seneviratne, S., Nicholls, N., Easterling, D., Goodess, C., Kanae, S., Kossin, J., Luo, Y., Marengo, J., McInnes, K., Rahimi, M., Reichstein, M., Sorteberg, A., Vara, C., Zhang, X., Alexander, L. V., Allen, S., Benito, G., Cavazos, T., Clague, J., … Zwiers, F. W. (2012). Changes in climate extremes and their impacts on the natural physical environment. In C. B. Field, V. Barros, T. F. Stocker, D. Qin, D. J. Dokken, K. L. Ebi, M. D. Mastrandrea, K. J. Mach, G.-K. Plattner, S. K. Allen, M. Tignor, & P. M. Midgley (Eds.), IPCC WGI/WGII special report on managing the risks of extreme events and disasters to advance climate change adaptation (SREX) (pp. 109–230). Cambridge: Cambridge University Press.
Shen, M., Piao, S., Jeong, S. J., Zhou, L., Zeng, Z., Ciais, P., Chen, D., Huang, M., Jin, C. S., Li, L. Z. X., Li, Y., Myneni, R. B., Yang, K., Zhang, G., Zhang, Y., & Yao, T. (2015). Evaporative cooling over the Tibetan Plateau induced by vegetation growth. Proceedings of the National Academy of Sciences of the United States of America, 112, 9299–9304. https://doi.org/10.1073/pnas.1504418112
Singh, J., Ashfaq, M., Skinner, C. B., Andershon, W. B., Mishra, V., & Singh, D. (2022). Enhanced risk of concurrent regional droughts with increased ENSO variability and warming. Nature Climate Change, 12, 163–170. https://doi.org/10.1038/s41558-021-01276-3
Sreeparvathy, V., & Srinivas, V. V. (2022). Global assessment of spatiotemporal variability of wet, normal and dry conditions using multiscale entropy-based approach. Science and Reports, 12(1), 1–18. https://doi.org/10.1038/s41598-022-13830-w
Sui, Y., Lang, X., & Jiang, D. (2018). Projected signals in climate extremes over China associated with a 20 °C global warming under two RCP scenarios. International Journal of Climatology, 38, e678–e697. https://doi.org/10.1002/joc.5399
Sun, H., Wang, J., Xiong, J., Bian, J., Jin, H., Cheng, W., & Li, A. (2021). Vegetation change and its response to climate change in Yunnan Province, China. Advances in Meteorology, 2021(1–20), 8857589. https://doi.org/10.1155/2021/8857589
Tarantino, C., Forte, L., Blonda, P., Vicario, S., Tomaselli, V., Beierkuhnlein, C., & Adamo, M. (2021). Intra-annual sentinel-2 time-Series supporting grassland habitat discrimination. Remote Sens, 13, 277. https://doi.org/10.3390/rs13020277
Tilman, D., & Downing, J. A. (1994). Biodiversity and stability in grasslands. Nature, 367, 363–365. https://doi.org/10.1038/367363a0
Tilman, D., Reich, P. B., Knops, J., Wedin, D., Mielke, T., & Lehman, C. (2001). Diversity and productivity in a long-term grassland experiment. Science, 294, 843–845. https://doi.org/10.1126/science.1060391
Tong, S., Bao, Y., Te, R., Ma, Q., Ha, S., & Lusi, A. (2017). Analysis of drought characteristics in Xilingol grassland of northern China based on SPEI and its impact on vegetation. Math Prob Eng, 5209173, 1–11. https://doi.org/10.1155/2017/5209173
Tukey, J. (1949). Comparing individual means in the analysis of variance. Biometrics, 5(2), 99–114. https://doi.org/10.2307/3001913
van Ruijven, J., & Berendse, F. (2010). Diversity enhances community recovery, but not resistance, after drought. Journal of Ecology, 98(1), 81–86. https://doi.org/10.1111/j.1365-2745.2009.01603.x
Vicente-Serrano, S. M., Beguerıa, S., Lorenzo-Lacruz, J., Camarero, J. J., Lopez-Moreno, J. I., Azorin-Molina, C., Revuelto, J., Moran-Tejeda, E., & Sanchez-Lorenzo, A. (2012). Performance of drought ındices for ecological, agricultural and hydrological applications. Earth Interactions, 16(10), 1–27. https://doi.org/10.1175/2012EI000434.1
Vicente-Serrano, S. M., Gouveia, C., Camarero, J. J., Beguería, S., Trigo, R., Lopez- Moreno, J. I., Azorín-Molina, C., Pasho, E., Lorenzo-Lacruz, J., Revuelto, J., Moran-Tejeda, E., & Sanchez-Lorenzo, A. (2013). Response of vegetation to drought time-scales across global land biomes. Proceedings of the National Academy of Sciences of the United States of America, 110(1), 52–57. https://doi.org/10.1073/pnas.1207068110
Vicente-Serrano, S. M., Beguería, S., López-Moreno, J. I. (2010). A multi-scalar drought index sensitive to global warming: the Standardized Precipitation Evapotranspiration Index – SPEI. Journal of Climatology, 23(7), 1696–1718. https://www.jstor.org/stable/26189715. Accessed 26 Nov 2018
Vogel, A., Scherer-Lorenzen, M., & Weigelt, A. (2012). Grassland resistance and resilience after drought depends on management intensity and species richness. PLoS ONE, 7(5), e36992. https://doi.org/10.1371/journal.pone.0036992
Volder, A., Tjoelker, M. G., & Briske, D. D. (2010). Contrasting physiological responsiveness of establishing trees and a C4 grass to rainfall event, intensified summer drought and warming in oak savanna. Global Change Biology, 16(12), 3349–3362. https://doi.org/10.1111/j.1365-2486.2009.02152.x
Wang, C., Wang, J., Zhang, F., Yang, Y., Luo, F., Li, Y., & Li, J. (2022). Stability response of alpine meadow communities to temperature and precipitation changes on the Northern Tibetan Plateau. Ecology and Evolution, 12(2), e8592. https://doi.org/10.1002/ece3.8592
Wang, Q., Yang, Y., Liu, Y., Tong, L., Zhang, Q. P., & Li, J. (2019a). Assessing the impacts of drought on grassland net primary production at the global scale. Science and Reports, 9, 14041. https://doi.org/10.1038/s41598-019-50584-4
Wang, S., Loreau, M., De Mazancourt, C., Isbell, F., Beierkuhnlein, C., Connolly, J., Doležal, J., Eisenhauer, N., Jentsch, A., Kreyling, J., Lanta, V., Leps, J., Polley, H. W., Reich, P. B., van Ruijven, J., Tilman, D., Wilsey, B., & Craven, D. (2021). Biotic homogenization destabilizes ecosystem functioning by decreasing spatial asynchrony. Ecology, 102(6), e03332. https://doi.org/10.1002/ecy.3332
Wang, X., Dong, J., Baoyin, T., & Bao, Y. (2019b). Estimation and climate factor contribution of aboveground biomass in Inner Mongolia’s Typical/Desert Steppes. Sustainability, 11(23), 6559. https://doi.org/10.3390/su11236559
Wang, Y., Zhang, C., Meng, F.-R., Bourque, C.P.-A., & Zhang, C. (2020). Evaluation of the suitability of six drought indices in naturally growing, transitional vegetation zones in Inner Mongolia (China). PLoS ONE, 15(5), e0233525. https://doi.org/10.1371/journal.pone.0233525
Wilcox, K. R., Shi, Z., Gherardi, L. A., Lemoine, N. P., Koerner, S. E., Hoover, D. L., Bork, E., Byrne, K. M., Cahill, J., Jr., Collins, S. L., Evans, S., Gilgen, A. K., Holub, P., Jiang, L., Knapp, A. K., LeCain, D., Liang, J., Garcia-Palacios, P., Penuelas, J., … Luo, Y. (2017). Asymmetric responses of primary productivity to precipitation extremes: A synthesis of grassland precipitation manipulation experiments. Global Change Biology, 23(10), 4376–4385. https://doi.org/10.1111/gcb.13706
Xu, H. J., Wang, X. P., & Yang, T. B. (2017). Trend shifts in satellite-derived vegetation growth in Central Eurasia, 1982–2013. Science of the Total Environment, 579, 1658–1674. https://doi.org/10.1016/j.scitotenv.2016.11.182
Yang, Z., Collins, S. L., Bixby, R. J., Song, H., Wang, D., & Xiao, R. (2021). A meta-analysis of primary productivity and rain use efficiency in terrestrial grassland ecosystems. Land Degradation & Development, 32(2), 842–850. https://doi.org/10.1002/ldr.3715
Zhang, Q., Li, J., Singh, V. P., & Bai, Y. (2012). SPI-based evaluation of drought events in Xinjiang, China. Nat Hazards, 64(1), 481–492. https://doi.org/10.1007/s11069-012-0251-0
Zhang, Q., Kong, D. D., Singh, V. P., & Shi, P. J. (2017). Response of vegetation to different time-scales drought across China, spatiotemporal patterns, causes and implications. Global and Planetary Change, 152, 1–11. https://doi.org/10.1016/j.gloplacha.2017.02.008
Zhang, L., Ameca, E. I., Cowlishaw, G., Pettorelli, N., Foden, W., & Mace, G. M. (2019). Global assessment of primate vulnerability to extreme climatic events. Nature Climate Change, 9, 554–561. https://doi.org/10.1038/s41558-019-0508-7
Zhang, X., Du, X., & Zhu, Z. (2020). Effects of precipitation and temperature on precipitation use efficiency of alpine grassland in Northern Tibet, China. Scientific Reports, 10, 20309. https://doi.org/10.1038/s41598-020-77208-6
Zhong, L., Ma, Y., Salama, M. S., & Su, Z. (2010). Assessment of vegetation dynamics and their response to variations in precipitation and temperature in the Tibetan Plateau. Climate Change, 103, 519–535. https://doi.org/10.1007/s10584-009-9787-8
Zhao, A., Yu, Q., Feng, L., Zhang, A., Pei, T., et al. (2020). Evaluating the cumulative and time-lag effects of drought on grassland vegetation: a case study in the Chinese Loess Plateau. Journal of Environment Management, 261, 110214. https://doi.org/10.1016/j.jenvman.2020.110214
Zhou, W., Gang, C., Li, J., Zhang, C., Mu, S., & Sun, Z. (2014). Spatial-temporal dynamics of grassland coverage and its response to climate change in China during 1982–2010. Acta Geographica Sinica, 69, 15–30. https://doi.org/10.11821/dlxb201401002
Zhou, B., Li, S., Li, F. U., Dong, S., Ma, F., Zhu, S., Zhou, H., & Stufkens, P. (2019). Plant functional groups asynchrony keep the community biomass stability along with the climate change-a 20-year experimental observation of alpine meadow in eastern Qinghai-Tibet Plateau. Agriculture, Ecosystems & Environment, 282, 49–57. https://doi.org/10.1016/j.agee.2019.06.002
Zhu, J., Huang, G., Wang, X., & Cheng, G. (2017). Investigation of changes in extreme temperature and humidity over China through a dynamical downscaling approach. Earth’s Future, 5(11), 1136–1155. https://doi.org/10.1002/2017EF000678
Zhu, J., Huang, G., Wang, X., Cheng, G., & Wu, Y. (2018). High-resolution projections of mean and extreme precipitations over China through PRECIS under RCPs. Climate Dynamics, 50(11–12), 4037–4060. https://doi.org/10.1007/s00382-017-3860-1
Acknowledgements
The authors do acknowledge the two sources for providing datasets. The satellite-derived NDVI data of 1982–2012 used in this study were obtained from the Oak Ridge National Laboratory Distributed Active Archive Center (Guay et al., 2015, https://daac.ornl.gov/VEGETATION/guides/GIMMS3g_NDVI_Trends.html). We extracted SPEI SPEIbase v2.5 dataset developed by Vicente-Serrano et al. (2010) based on the CRU 3.24.01 precipitation and potential evapotranspiration (http://spei.csic.es/database.html).
Funding
This work was supported by the research grants from the Research Grants Council of the Hong Kong Special Administrative Region, China [project number HKBU12302518] and the National Key R&D Program of China [project number 2019YFC1510400]. Parts of this study were supported by the EU Horizon 2020 Project e-shape under grant no. 820852.
Author information
Authors and Affiliations
Contributions
M. L. Hossain: conceptualization, methodology, software, formal analysis, writing—original draft, and writing—review and editing. J. Li: methodology, writing-review and editing, supervision, funding acquisition, and project administration. Y. Lai: methodology. C. Beierkuhnlein: conceptualization, methodology, writing–review and editing, and supervision.
Corresponding author
Ethics declarations
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Hossain, M.L., Li, J., Lai, Y. et al. Long-term evidence of differential resistance and resilience of grassland ecosystems to extreme climate events. Environ Monit Assess 195, 734 (2023). https://doi.org/10.1007/s10661-023-11269-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10661-023-11269-8