
Abstract
Reliable predictions of future distribution ranges of ecologically important species in response to climate change are required for developing effective management strategies. Here we used an ensemble modelling approach to predict the distribution of three important species of Abies namely, Abies pindrow, Abies spectabilis and Abies densa in the Hindu Kush Himalayan region under the current and two shared socioeconomic pathways (SSP245 and SSP585) and time periods of 2050 and 2090s. A correlative ensemble model using presence/absence data of the three Abies species and 22 environmental variables, including 19 bioclimatic variables and 3 topographic variables, from known distributions was built to predict the potential current and future distribution of these species. The individual models used to build the final ensemble performed well and provided reliable results for both the current and future distribution of all three species. For A. pindrow, precipitation of the driest month (Bio14) was the most important environmental variable with 83.3% contribution to model output while temperature seasonality (Bio4) and annual mean diurnal range (Bio2) were the most important variables for A. spectabilis and A. densa with 48.4% and 46.1% contribution to final model output, respectively. Under current climatic conditions, the ensemble models projected a total suitable habitat of about 433,003 km2, 790,837 km2 and 676,918 km2 for A. pindrow, A. spectabilis and A. densa, respectively, which is approximately 10.36%, 18.91% and 16.91% of the total area of Hindu Kush Himalayan region. Projections of habitat suitability under future climate scenarios for all the shared socioeconomic pathways showed a reduction in potentially suitable habitats with a maximum overall loss of approximately 14% of the total suitable area of A. pindrow under SSP 8.5 by 2090. A decline in total suitable habitat is predicted to be 9.6% in A. spectabilis by 2090 under the SSP585 scenario while in A. densa 6.67% loss in the suitable area is expected by 2050 under the SSP585 scenario. Furthermore, there is no elevational change predicted in the case of A. pindrow while A. spectabilis is expected to show an upward shift by about 29 m per decade and A. densa is showing a downward shift at a rate of 11 m per decade. The results are interesting, and intriguing given the occurrence of these species across the Hindu Kush Himalayan region. Thus, our study underscores the need for consideration of unexpected responses of species to climate change and formulation of strategies for better forest management and conservation of important conifer species, such as A. pindrow, A. spectabilis and A. densa.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Abies, the second largest genus in the family Pinaceae consisting of about 52 species worldwide, is the dominant species in the North Hemisphere coniferous forests (Xiang et al., 2007). The vertical distribution of this genus is largely concentrated in two elevational ranges with 15 species found between 1000 and 2000 m and 13 species found between 2500 and 4000 m elevation (Xiang et al., 2007). The Sino-Himalaya region is the major centre of diversity in Abies, an area where 18 species occur (Farjon, 2017). However, three species from west to east, A. pindrow, A. spectabilis and A. densa, are well defined and broadly accepted in the main Himalayan chain, beginning from Afghanistan’s Hindu Kush eastward as far as Bhutan. Each of these blue-coned firs has an area of discreet distribution but A. spectabilis overlaps with both A. pindrow and A. densa to some extent at its two extremes (Farjon, 2017). Abies spectabilis and A. densa are more closely related to each other than either is to A. pindrow. These plants are valuable forest resources largely used for construction, furniture and other industrial materials. A. pindrow is widely distributed across Hindu Kush Himalaya (HKH) in eastern Afghanistan, western Nepal, north India and Pakistan (Xiang et al., 2007) and has an elevation distribution range between 2000 and 3300 m (Singh, 2018). Similarly, A. spectabilis is distributed in Xizang (China), Afghanistan, north India and Nepal (Xiang et al., 2007) across the HKH with an elevation range from 2500 to 4000 m (Vidakovic, 1993) and A. densa occurs in South Xizang (China), Bhutan, North-east India and Nepal (Xiang et al., 2007) with elevation range between 2450 and 4000 m (Farjon, 2010). Generally, A. pindrow and A. spectabilis occur towards the west while A. densa occurs towards the east of HKH. Since these species form a dominant part of the canopy in the Himalayan forests, they play an immense role in carbon sequestration and, hence, in the mitigation of climate change. Carbon stored in forests forms a dominant part of terrestrial carbon stocks and thus management of forest ecosystems is crucial because of their strong mitigation potential (Fargione et al., 2018; Griscom et al., 2017). Any change in suitable habitat for dominant tree species will have a direct impact on the carbon sequestration capacity of the forest ecosystems.
However, the HKH mountains and the mountainous systems that inhabit these and other species are continuously degrading (Chakraborty et al., 2018) because of both direct (land conversion, exploitation of forest resources (Pandit, 2017) and indirect stressors, such as global climatic change, soil erosion (Salesa & Cerdà, 2020) and the spread of invasive alien species (Lamsal et al., 2018; McDougall et al., 2011) that may disproportionately affect mountains in a long run (Halofsky et al., 2018; Luce, 2018) because of being rich in geodiversity (Gordon, 2018), biodiversity (Antonelli, 2015; Noroozi et al., 2018), sources of major drainage systems (Immerzeel et al., 2020; Viviroli & Weingartner, 2008), home to about twenty per cent of the world’s human population (Körner et al., 2017) and for providing valuable resources, goods and services (Sharma et al., 2019a) which are of critical importance locally, regionally (Gao et al., 2016) and globally (Fang et al., 2018). Mountains support approximately 25% of all terrestrial biodiversity (Sharma et al., 2019a). This richness of biodiversity in the mountains is a result of many local niches created due to steep elevation gradients and associated climatic variations that support a diverse assemblage of species (Körner & Spehn, 2019).
The Hindu Kush Himalaya (HKH), one of the several mountain systems in the world, is also rich in biodiversity (Xu et al., 2019), has the largest glacierized area outside the North and South pole and is home to ten large Asian water systems (Kotru et al., 2020). But this mountain system is experiencing rapid and elevation-dependent warming, along with an increase in extreme climate events, especially since the 1980s (Ren et al., 2017; Sun et al., 2020; Zhan et al., 2017). From 1901 to 2014, the mean annual temperature has increased by approximately 0.1 °C/decade (Ren et al., 2017) and it is predicted to increase by about 1–2 °C from 2021 to 2050 compared to 1961–1990 (IPCC, 2021). This warming has resulted in the loss/retreat of glacier mass, accelerated snowmelt and degradation of permafrost (Chug et al., 2020; Sabin et al., 2020; IPCC Report, 2021), alteration in the distribution of plant communities (Manish et al., 2016) and elevational shift in high-altitude species (Hamid et al., 2020; Telwala et al., 2013). Climate change is of particular concern because it leads to changes in assemblages of species (Parmesan & Yohe, 2003), distribution of species (Naimi et al., 2014) and phenology (Ovaskainen et al., 2013) and various evolutionary changes (Hoffmann & Sgrò, 2011). For these reasons, the studies documenting the effects of climate change on vegetation growth and distribution have been accumulating globally (Baumbach et al., 2021).
Changes in the distribution of species in response to climate change have been studied by employing a species distribution modelling approach. Recently, Bobrowski et al. (2021) reviewed 157 studies published between 2010 and 2021 on modelling species distribution in the Himalaya and most of these studies have addressed the question of potential range changes in species in future under various climate change scenarios. Although many of these studies were carried out on tree species (27%), no such study is available on A. pindrow, A. spectabilis and A. densa which are the dominant conifers in the Hindu Kush Himalayan forests. Since these species grow over a wide geographical range in the Hindu Kush Himalaya, we hypothesized that they may extend their range under the changing climate. To this end, we used an ensemble modelling approach for predicting the potential distribution of A. pindrow, A. spectabilis and A. densa under current and two future shared socioeconomic pathway (SSP) scenarios using the vastly improved Coupled Model Intercomparison Project (CMIP6) model. The shared socioeconomic pathway (SSP) is the latest iteration of scenarios, used for CMIP6 (2016–2021) and IPCC Sixth Assessment Report (AR6) (2021) and its use for the first time in predicting the distribution of three important Abies species was aimed to address the following two main research questions: (i) which environmental/topographic variables explain the distribution of three Abies species in the HKH region; (ii) how would the predicted climate change impact the distribution of three Abies species in the Hindu Kush Himalaya under various climate change scenarios. Since being dominant constituents of the coniferous forests in the Hindu Kush Himalaya, any change in the distribution of these species will impact the structural organization and functional integrity of the forest ecosystem with consequences for the marginalized communities in the Hindu Kush Himalaya.
Materials and methods
Study area
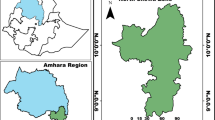
We used an ensemble modelling approach to explore the current distribution of three Abies species under current climatic conditions and predict their future distribution in the HKH region (Fig. 1). The total geographic area of HKH is approximately 4.2 million km2 (Bajracharya & Shrestha, 2011; Bajracharya et al., 2015; Sharma et al., 2019b). Abies pindrow (Royle ex. D.Don) Royle is one of the dominant conifer species in the Hindu Kush Himalayan (HKH) region and is native to Afghanistan, Nepal and West Himalaya (Farjon, 2010). Abies spectabilis (D. Don) Mirb. is native to Afghanistan, Nepal, Pakistan, Tibet and West Himalaya (Turland & Figueiredo, 1999). Similarly, Abies densa Griff. is native to Assam, East Himalaya, Nepal and Tibet (Farjon, 2010).

Map of the study area (the Hindu Kush Himalayan region) with occurrence points of three Abies species used for modelling
Species data
The current occurrence data with correctly geo-referenced coordinates were gathered from our field studies, published records in books, journals, reports and other literature and herbarium records mostly of Kashmir University Herbarium (KASH). Additionally, we also retrieved occurrence records of the three species from the Global Biodiversity Information Facility (https://www.gbif.org) accessed from R 4.0.5 (R Core Team, 2020) using the ‘rgbif’ package (Chamberlain, 2017) and these occurrence records were cleaned and records with missing geo-coordinates or with any dubious coordinate issues were deleted. To ensure the authenticity of occurrence records, the coordinates of the records were compared with the country/region of the site as given in the records (Hijmans et al., 1999). The occurrence points were spatially thinned with ‘spThin’ package (Aiello-Lammens et al., 2015) to keep only one occurrence point per cell of 5 km × 5 km dimension, resulting in 279, 292 and 196 occurrence points for A. pindrow, A. spectabilis and A. densa, respectively. The package gives a spatially thinned occurrence data set in which all occurrence data points are at least 5 km apart (user-defined thinning distance) which helps to reduce the impact of uneven/biased occurrence records on model output (Aiello-Lammens et al., 2015).
Environmental data
Data for 19 bioclimatic variables at a spatial resolution of 2.5 arc minute (~ 5 km) to represent current and future climate scenarios were extracted from the WorldClim 2.1 dataset (Fick & Hijmans, 2017). Such a spatial resolution was selected because of the non-availability of future climatic data at lower resolution (~ 1 km). These bioclimatic variables are derived from monthly temperature and precipitation records from 1970 to 2000 and represent a combination of means, extremes, variability and seasonality of temperature and precipitation data (Ahmad et al., 2020; Zhang et al., 2019). These bioclimatic variables are preferentially used in species distribution modelling studies across taxa since they provide biologically useful climatic information than individual temperature and precipitation (Borzée et al., 2019; Fournier et al., 2017; Kumar, 2012; Root et al., 2003). In addition to bioclimatic variables, three topographic variables aspect, slope and elevation were extracted from DEM at the same resolution to perform robust modelling. The DEM data was derived from the Shuttle Radar Topography Mission (SRTM; Farr et al., 2007).
Future climatic data derived from the BCC-CSM2-HR global circulation model (Beijing Climate Centre, China Meteorological Administration, China) was used. These data have been previously used with great accuracy in the Hindu Kush Himalayan region for the prediction of vegetation distribution in response to future climate change (Ahmad et al., 2020; Dakhil et al., 2019; Shi et al., 2018; Zhan et al., 2017). We thus obtained the predicted values of each bioclimatic variable for two future time periods, near 2050 (2041–2060) and far 2090 (2081–2100) under two shared socioeconomic pathways of 4.5 and 8.5 (Fick & Hijmans, 2017).
Species distribution modelling
We used the ensemble modelling approach for modelling the potential distribution of three Abies species under current and future climatic conditions. Ensemble modelling procedure helps in producing more reliable habitat suitability maps than individual species distribution models. We used nine modelling algorithms available in ‘sdm’ package (Naimi & Araújo, 2016) in R 4.0.5 (R Core Team, 2020), for final ensemble building. The individual modelling methods used for the ensemble included generalized linear model (GLM), generalized additive model (GAM), boosted regression tree (BRT), flexible discriminant analysis (FDA) multivariate adaptive regression spline (MARS), maximum entropy (Maxent), radial basis function (RBF) random forest (RF) and support vector machine (SVM).
We generated 500 pseudo absences randomly for each species within the study region for the smooth running of individual models. Seventy percent of data points were used for model building and 30% were set aside for testing the individual models (Elith & Leathwick, 2009; Souza & Prevedello, 2021). Each modelling method was replicated six times resulting in fifty-four different statistical models which were averaged by calculating the weighted mean to produce the final ensemble.
Selection of predictor variables
Variance inflation factor (VIF) was used for testing multicollinearity in environmental variables (Naimi et al. 2014) and the variables that were highly correlated (r > 0.7) were excluded.
Individual models were evaluated by calculating the area under the curve (AUC) separately for each modelling method and iteration. AUC measures the model capacity to differentiate sites with species presence from sites where the species is absent (Swets, 1988). The values of AUC range from 0 to 1 with 0 being the least perfect and 1 being the most perfect whereas 0.5 indicates that the model prediction is not better than a random guess (Chakraborty et al., 2016; Fielding & Bell, 1997; Phillips & Dudík, 2008; Swets, 1988). Models with AUC scores of more than 0.9 are generally considered the best models (Lobo et al., 2008).
Analysis of ensemble output and habitat change
The continuous suitability maps of resultant ensembles were converted to binary maps using the maximum training specificity and sensitivity threshold method commonly used for the assessment of impacts of climate change on species (Liu et al., 2005, 2013; Souza & Prevedello, 2021).
Potential habitat under the current climate was overlayed with individual future climate change scenarios for assessment of changes in the distribution range of the species. Areas suitable/unsuitable in both current and future climate scenarios were classified as ‘suitable/unsuitable (no change)’ whereas areas which are suitable in the current climate but unsuitable in future climatic conditions were classified as ‘loss’ and areas which are unsuitable in the current climate but were suitable in future climatic conditions were classified as ‘gain’. The total area under different habitat suitability categories as well as habitat change was calculated using the ‘geosphere’ package (Hijmans et al., 2019) in R 4.0.5 (R Core Team, 2020).
The degree of niche overlap between the species was measured by Warren’s similarity static (I) (Warren et al., 2008). The similarity static ‘I’ compares the two habitat suitability predictions, and the output is a value representing the similarity between the ecological niches of the species. The value of ‘I’ ranges between 0 and 1 with 0 being no overlap and 1 being complete overlap (identical niches).
Results
Predictor environmental variables
After checking for multicollinearity, seven environmental variables out of twenty-two were used for modelling the distribution of each species (Fig. 2). Two temperature-related variables, namely mean annual diurnal range (Bio2) and mean temperature of wettest quarter (Bio8), and three precipitation-related variables, namely precipitation of driest month (Bio14), precipitation seasonality (Bio15) and precipitation of warmest quarter (Bio18) and two topographic variables aspect and slope were found to be useful in the prediction of the distribution of A. pindrow. These variables helped in building a robust ensemble model to produce habitat suitability maps for A. pindrow in each 2.5 arc minute grid cell under current climatic conditions in the HKH region. Bioclimatic variables are the most important predictors influencing the distribution of A. pindrow in the HKH region. Precipitation of driest month (Bio14) was the most important variable with variable importance of 83.3% followed by precipitation of warmest quarter (Bio18) and precipitation seasonality (Bio15) with variable importance of 7.4% and 4.9%, respectively (Table 1).

Climatic variables used for modelling the potential distribution of A. pindrow, A. spectabilis and A. densa in the HKH region
In the case of A. spectabilis, three temperature-related variables Bio2, Bio4 and Bio9 and two precipitation-related variables Bio14 and Bio15 along with two topographic variables aspect and slope were related to the distribution of the species. Bio4 was the most important variable with a 48.4% contribution to the final model output while Bio14, Bio9 and slope contributed 16.8%, 16% and 8.5% to the final model output, respectively (Table 1).
For A. densa, two temperature-related variables Bio2 and Bio3 and two precipitation-related Bio14 and Bio15 and three topographic variables aspect, slope and elevation were the important environmental variables explaining the current distribution of species in the HKH region. Bio2 was the most important environmental variable with a 46.1% contribution to the final model output. Bio3 and elevation respectively contributed 27.4% and 12.9% to the final model output (Table 1). Interestingly, temperature-related variables are more important in explaining suitable habitat for A. densa.
Individual response curves for the potential distribution of A. pindrow under the current climate showed that precipitation in the driest month (Bio14) positively influenced the probability of species occurrence, with the highest suitability at total monthly precipitation of approximately 30 mm and unsuitable below 15 mm precipitation in the driest month (Fig. 3). For A. spectabilis, temperature seasonality (Bio4) of approximately 400% resulted in the highest suitability whereas it decreases at higher percentages with areas with more than 600% temperature seasonality being unsuitable for the species (Fig. 3). For A. densa, the annual mean diurnal range (Bio2) of 7.5 degrees resulted in the highest suitability and the suitability decreased at higher mean diurnal ranges with suitability approaching zero after 15 degrees (Fig. 3).

Response curves representing the relationship between key predictor variables and the probability of occurrence of A. pindrow, A. spectabilis and A. densa
Similarly, the suitability of A. pindrow decreased with an increase in precipitation of the warmest quarter (Bio18) while the suitability of A. spectabilis and A. densa increased with an increase in the precipitation of driest month (Bio14) and isothermality (Bio3), respectively.
Model performance
The individual models for all the three species performed very well as indicated by an AUC of 0.96 ± 0.01, 0.95 ± 0.03 and 0.93 ± 0.01 for A. pindrow, A. spectabilis and A. densa, respectively. We then calculated the weighted mean of all the models to produce separate maps of predicted habitat suitability for current and future climatic conditions for the three species of Abies.
Model projections
Based on the current habitat suitability map of A. pindrow, suitability areas are found in northern Pakistan; Indian states of Jammu and Kashmir, Uttarakhand and Himachal Pradesh; Nepal; and Sichuan and Yunnan provinces of China (Fig. 4). A. spectabilis suitable areas lie in the Indian states of Himachal Pradesh, Uttarakhand and Arunachal Pradesh, Nepal, Bhutan and northern parts of Yunnan Province in China (Fig. 4). Likewise, northern parts of Arunachal Pradesh, Meghalaya, Mizoram, Manipur and Nagaland in India, north of Nepal, Bhutan and Xicheng in China is suitable for A. densa under current climatic conditions (Fig. 4).

Potential habitat suitability map of A. pindrow, A. spectabilis and A. densa under current and future climate scenarios
Under the current climatic conditions, the model projections highlight regions with different probability of occurrence of Abies spp. in the HKH region. The total suitable habitat for A. pindrow, A. spectabilis and A. densa is spread over 433,003 km2, 790,837 km2 and 676,918 km2 comprising 10.36%, 18.91% and 16.19% of the total area of the HKH region, respectively (Fig. 5).

Range contraction (loss) and range expansion (gain) under future climate scenarios compared to current climatic conditions in different time periods for A. pindrow, A. spectabilis and A. densa
In HKH, under current climatic conditions, the total area which is suitable for at least one of the Abies species is 1,231,889 km2 (29.46%) (Fig. 6). The area suitable for both A. spectabilis and A densa is 9.94% of the total area while the area suitable for both A. pindrow and A. spectabilis is 6.05% and the area suitable for both A. pindrow and A. densa is 1.6% of the total HKH area. Of the total area of HKH, 1.59% is suitable for all three species while 4.3%, 4.52% and 6.24% areas are exclusively suitable for A. pindrow, A. spectabilis and A. densa, respectively (Fig. 6).

a Map showing overlap between three Abies species in the HKH region. NS = not suitable for Abies, AP = A. pindrow (179,811 km2), AS = A. spectabilis (188,884 km2), AD = A. densa (260,982 km2), AP + AS = overlap between A. pindrow and A. spectabilis (186,277 km2), AP + AD = overlap between A. pindrow and A. densa (260 km2), AS + AD = overlap between A. spectabilis and A. densa (349,021 km2), ALL = three species overlap (66,655 km.2). b Venn diagram showing the overlap in potential suitable area for A. pindrow, A. spectabilis and A. densa under the current climate in the HKH region
The results from the niche overlap test revealed that the realized niches of the three Abies species are quite identical. The similarity static ‘I’ value between potentially suitable habitats for A. pindrow and A. spectabilis was 0.91, while between A. pindrow and A. densa was 0.82. The niche overlap was highest between A. spectabilis and A. densa (0.96).
Predicted habitat suitability in current climatic conditions was projected onto future climatic conditions under selected shared socioeconomic pathways in 2050 and 2090 and it showed a reduction in potentially suitable habitats for all three species.
For A. pindrow, the total suitable area showed a decrease of 9.39% and 10.25% by 2050 under SSP245 and SSP585 climate scenarios, respectively. The loss in the total suitable area is predicted to be more, 11.19% and 14% by 2090 under SSP245 and SSP585 scenarios, respectively (Table 2).
In the case of A. spectabilis, the decrease in the total suitable area by 2050 is 3.36% and 3.11% under SSP245 and SSP585 scenarios, respectively, while the decrease is 5.59% and 9.6% by 2090 (Table 2).
A. densa showed an increase in the total suitable area of 2.82% by 2050 under the SSP245 scenario while there is a decrease of 5.17% under the SSP585 scenario. By 2090, the decrease is 6.67% under SSP245 scenarios and no change in the SSP245 scenario (Table 2).
Range expansion andcontraction
Abies pindrow
Area with favourable climatic conditions for A. pindrow showed an overall decrease under all SSP scenarios in all the time periods (Fig. 5).
By 2050, the currently suitable area is expected to decline by approximately 1.17% and 1.65% under SSP245 and SSP585 scenarios, respectively (Table 3). The species is expected to expand its range in currently unsuitable areas from 0.33 to 0.44% under future climate scenarios by 2050. The area suitable in both current and future climate scenarios ranges from 8.85 to 9.33%. The decline in the currently suitable area ranges from 1.63 to 2.21% under different future scenarios by the year 2090 while there is expansion expected to be between 0.41 and 0.91% by the end of this century. The area suitable under both scenarios is between 8.29 to 8.87% (Table 3).
Abies spectabilis
The range of contraction is predicted to be from 1.36 to 1.60% by 2050 and that of expansion ranges between 0.54 and 0.73% (Table 3). The suitable (no change) area ranges from 17.32 to 17.55%. By the end of this century, the species is expected to lose its habitat by 1.5 to 3.13% while its suitability habitat is expected to increase between 0.91 and 1.31%. 15.79 and 17.42% area is suitable in both scenarios (no change).
Abies densa
The species is expected to lose range from 0.87 to 2.6% while there is predicted range expansion by 1.33 to 1.52% by the middle of this century (Table 3). The no-change suitable area ranged from 13.59 to 15.31%. The range contraction is expected to be between 1.73 and 2.15% by 2090 while the range expansion is predicted to be between 0.9 and 2.15%. The area suitable in both the current and future climate scenario ranges from 14.04 to 14.46%.
Elevation shift
The mean elevation of A. pindrow under current climatic conditions is calculated as 2871 m which shows a decrease by the year 2050 being 2743 m and 2738 m under SSP245 and SSP585 scenarios while by 2090 the mean elevation is shown to decrease to 2730 m in SSP245 and increase to 2933 m under SSP585 scenario (Fig. 7). Overall, there is no drastic change in the mean elevation of A. pindrow in future climate scenarios. For A. spectabilis, the mean elevation under current climatic conditions is 2626 m while it is predicted to increase in all the future climatic scenarios between 2558 and 2733 m in 2050 and 2740 and 2921 m in 2090. Overall, there is approximately a 29 m upward shift per decade for A. spectabilis (Fig. 7). The mean elevation for A. densa under the current climate is 2733 m which was predicted to decline under all future climate scenarios. It indicates a shift towards lower elevations to 2459–2668 m by 2050 and 2575–2641 m by 2090. The average shift downwards is approximately 11 m per decade (Fig. 7).

Plot showing mean elevation of A. pindrow, A. spectabilis and A. densa and its variation with time
Discussion
The present study brings out that the most important environmental variables to the model output were precipitation of the driest month for A. pindrow, temperature seasonality for A. spectabilis and mean annual diurnal range for A. densa. Furthermore, our study predicts that each species is likely to lose potentially suitable habitat area under future climatic change scenarios: with a maximum suitable habitat loss of 14% in A. pindrow, 9.6% in A. spectabilis and 6.7% in A. densa. Also, the species-specific elevational shift is predicted in response to climate change with an upward shift in A. spectabilis, and a downward shift in A. densa but no elevational shift is predicted in A. pindrow. Thus, each species has a unique response to climate change, despite taxonomic and ecological relatedness. For the prediction of future potential distribution and possible range shifts in these Himalayan conifers in response to climate change, a proper mechanistic understanding of the impacts of key environmental and topographic factors on the growth and distribution of these species is greatly needed (Bobrowski et al., 2017). Among the bioclimatic variables, we found that precipitation during the driest month had an overriding influence on the distribution of A. pindrow in the Hindu Kush Himalaya explaining about 83% of its occurrence probability. It is very well known that the growth, development and distribution of plants are significantly influenced by climatic factors, especially temperature and precipitation (Bista et al., 2021). In natural forests, A. pindrow is a shade-tolerant species (Rushforth, 1991), prefers cooler and moist habitats and is thus largely present on the North-East-facing slopes (Sharma et al., 2010). Jha et al. (1984) also recorded the cool/moist habitat preference of A. pindrow. Dendrochronological studies on A. pindrow have also revealed a strong association between radial growth and spring precipitation in dry regions (Gaire et al., 2017; Kharal et al. 2017; Sigdel et al., 2018) but the growing season temperature impacts growth at higher elevations in wet regions ( Gaire et al., 2017; Malik & Sukumar, 2021; Shrestha et al., 2017).
Model projections under various climate change scenarios and time periods revealed a decrease in the distribution range of A. pindrow in the Hindu Kush Himalaya. Such declines in the potential distribution ranges of A. pindrow have been reported earlier by Ali et al. (2014) in the Swat district of northern Pakistan and by Naudiyal et al. (2021) in the Sichuan province of China as well. It could be correlated with the declining summer and annual precipitation in the central-eastern HKH noticed from 1979 to 2010 possibly due to a weakening of the South Asian monsoon (Palazzi et al., 2013; Roxy et al., 2015; Yao et al., 2012). The precipitation trend over the western HKH, which receives a substantial amount of its precipitation from western disturbances during winter (Meher et al., 2018; Palazzi et al., 2013; Singh et al., 2016), is unclear (Hunt et al., 2019; Krishnan et al., 2019; Kumar et al., 2015). CMIP6 model projections, under SSP 8.5 future climate scenario, over the western Himalaya predict an increase in winter precipitation by the end of the current century (Almazroui et al., 2020). The mean precipitation of the driest month shows a decline compared to current in all future scenarios except in the SSP585 (2090) scenario. This decreasing trend in the precipitation is very well indicated in the future distribution changes of the species. A decrease in precipitation or change in its seasonal or annual pattern may result in long dry periods during the growing season thus altering tree water potential (ψ), modify tree physiology and limit tree species distribution (Poudyal et al., 2004; Zobel et al., 2001; Zobel & Singh, 1995). Global warming might also increase aridity through increased vapour pressure deficit which can result in aggravated soil moisture deficit (Giorgi & Lionello, 2008; Piñol et al., 1998).
Drought-induced range contraction in A. pindrow is reported both from dry (Dorman et al., 2013; Sarris et al., 2007) and mesic sites (Castagneri et al., 2014; Jump et al., 2006; Linares & Camarero, 2012) of the Mediterranean forests. The drought caused a widespread decline in tree growth and forest die-off. In Central Europe also, the occurrence of A. pindrow in the past century has declined partly due to climate change and partly due to acid rain and other anthropogenic factors (Mátyás et al., 2021). However, some suggest that the species may benefit from winter warming (van der Maaten-Theunissen et al. 2013), while others reason that it is likely to suffer from increasing summer droughts (Lebourgeois et al., 2010; Thabeet et al. 2009). Although A. pindrow is known to survive under stress and is considered to be highly resilient to environmental changes (Garzón et al., 2019), our results predict its range contraction under future climate scenarios. It needs to be emphasized that long-term growth responses of trees to climate are highly contingent on local climatic site conditions that may buffer the effects of warming trends and changing precipitation regimes.
In A. spectabilis, the temperature is the main driving factor for tree growth while precipitation, especially in the early growing season, in the central Himalayas aids in tree growth by providing sufficient soil moisture for radial growth (Rai et al., 2020). The decline in suitable habitats may be linked with an increase in mean temperatures in future as predicted under different SSP scenarios. An increase in mean temperature enhances evapotranspiration which results in soil moisture deficiency thereby leading to less suitability for the species. To cope with the increased temperature, the species is expected to shift upwards to higher elevations. The rate of upward shift predicted in this study agrees with other studies on the species (Gaire et al., 2014). Previous studies in this region also showed upward elevational shift under future climate scenarios (Chhetri et al., 2018; Liang et al., 2018; Mohapatra et al., 2019; Zomer et al., 2014). Zomer et al. (2014) predicted a significant upward elevational shift of the subalpine coniferous forest zone in future climate scenarios. Likewise, a large upward shift in average elevation was predicted in A. spectabilis, Betula utilis and Pinus wallichiana under future climate scenarios (Chhetri et al., 2018). Schickhoff et al. (2015) also predicted an upward shift of B. utilis under future climate scenarios.
Temperature is again the most important climatic factor for the distribution of A. densa in the HKH region. The areas suitable for the species receive good annual rainfall which ensures ambient water availability throughout the growing season. The species is expected to show downward movement in future climate conditions. There are reports of a declining number of this species near the treeline with dry climate being one of the possible reasons (Ciesla & Donaubauer, 1994). A similar modelling study for the assessment of climate change impacts on Rhododendrons in the eastern Himalaya predicted a significant loss of suitable habitat for this high-altitude species in future climate scenarios (Kumar, 2012).
Conclusion
This study corroborates the general influence of climate change on the potential distribution of plant species, including the dominant Abies species in the Hindu Kush Himalayan region. Compared to Abies pindrow and Abies spectabilis, Abies densa is expected to retain the maximum suitable area under various climate change scenarios over a large period of time. Furthermore, there is an upward elevation shift expected in A. spectabilis and a downward shift in A. densa while there is no change in elevation range in the case of A. pindrow under future climate change scenarios. The key environmental variables influencing habitat suitability varied with species. While precipitation of the wettest month was the key environmental variable explaining the potential habitat suitability for A. pindrow, it was temperature seasonality and annual mean diurnal range in the case of A. spectabilis and A. densa, respectively. These species show varied responses to climate as depicted in the key environmental variables and contrasting shifts in mean elevational shift under future climate scenarios. This highlights the fact that not all species respond to the climate in a similar fashion. Given the predicted forest cover loss in future climate scenarios, detrimental impacts on several vital ecosystem services such as carbon sequestration are expected in future.
By using a robust ensemble modelling approach, it is concluded that predicted climate change is likely to influence the distribution and abundance of key coniferous species in the Hindu Kush Himalaya and appropriate adaptation and mitigation strategies need to be developed to combat the negative impacts of climate change, particularly in the ecologically fragile Hindu Kush Himalaya.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Ahmad, S., Yang, L., Khan, T. U., Wanghe, K., Li, M., & Luan, X. (2020). Using an ensemble modelling approach to predict the potential distribution of Himalayan gray goral (Naemorhedus goral bedfordi) in Pakistan. Global Ecology and Conservation, 21, e00845. https://doi.org/10.1016/j.gecco.2019.e00845
Aiello-Lammens, M. E., Boria, R. A., Radosavljevic, A., Vilela, B., & Anderson, R. P. (2015). spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography, 38(5), 541–545. https://doi.org/10.1111/ecog.01132
Ali, K., Ahmad, H., Khan, N., & Jury, S. (2014). Future of Abies pindrow in Swat district, northern Pakistan. Journal of Forestry Research, 25(1), 211–214. https://doi.org/10.1007/s11676-014-0446-1
Almazroui, M., Islam, M. N., Saeed, S., Saeed, F., & Ismail, M. (2020). Future changes in climate over the Arabian Peninsula based on CMIP6 multimodel simulations. Earth Systems and Environment 2020 4:4, 4(4), 611–630. https://doi.org/10.1007/S41748-020-00183-5
Antonelli, A. (2015). Multiple origins of mountain life. Nature, 524(7565), 300–301. https://doi.org/10.1038/nature14645
Bajracharya, S. R., Maharjan, S. B., Shrestha, F., Guo, W., Liu, S., Immerzeel, W., & Shrestha, B. (2015). The glaciers of the Hindu Kush Himalayas: Current status and observed changes from the 1980s to 2010. International Journal of Water Resources Development, 31(2), 161–173. https://doi.org/10.1080/07900627.2015.1005731
Bajracharya, S., & Shrestha, B. (2011). The status of glaciers in the Hindu Kush-Himalayan region. Kathmandu, Nepal.: ICIMOD. http://lib.riskreductionafrica.org/bitstream/handle/123456789/1124/thestatusofglaciersinthehindu.pdf?sequence=1. Accessed 12 September 2021
Baumbach, L., Warren, D. L., Yousefpour, R., & Hanewinkel, M. (2021). Climate change may induce connectivity loss and mountaintop extinction in Central American forests. Communications Biology, 4(1), 869. https://doi.org/10.1038/s42003-021-02359-9
Bista, R., Chhetri, P. K., Johnson, J. S., Sinha, A., & Shrestha, K. B. (2021). Climate-driven differences in growth performance of cohabitant fir and birch in a subalpine forest in Dhorpatan Nepal. Forests, 12(9), 1137. https://doi.org/10.3390/f12091137
Bobrowski, M., Gerlitz, L., & Schickhoff, U. (2017). Modelling the potential distribution of Betula utilis in the Himalaya. Global Ecology and Conservation, 11, 69–83. https://doi.org/10.1016/j.gecco.2017.04.003
Bobrowski, M., Weidinger, J., & Schickhoff, U. (2021). Is new always better? Frontiers in global climate datasets for modeling treeline species in the Himalayas. Atmosphere, 12(5), 543. https://doi.org/10.3390/atmos12050543
Borzée, A., Andersen, D., Groffen, J., Kim, H.-T., Bae, Y., & Jang, Y. (2019). Climate change-based models predict range shifts in the distribution of the only Asian plethodontid salamander: Karsenia koreana. Scientific Reports, 9(1), 11838. https://doi.org/10.1038/s41598-019-48310-1
Castagneri, D., Nola, P., Motta, R., & Carrer, M. (2014). Summer climate variability over the last 250 years differently affected tree species radial growth in a mesic Fagus–Abies–Picea old-growth forest. Forest Ecology and Management, 320, 21–29. https://doi.org/10.1016/j.foreco.2014.02.023
Chakraborty, A., Joshi, P. K., & Sachdeva, K. (2016). Predicting distribution of major forest tree species to potential impacts of climate change in the central Himalayan region. Ecological Engineering, 97, 593–609. https://doi.org/10.1016/j.ecoleng.2016.10.006
Chakraborty, A., Saha, S., Sachdeva, K., & Joshi, P. K. (2018). Vulnerability of forests in the Himalayan region to climate change impacts and anthropogenic disturbances: A systematic review. Regional Environmental Change, 18(6), 1783–1799. https://doi.org/10.1007/s10113-018-1309-7.
Chamberlain, S. (2017). rgbif: Interface to the global ‘biodiversity’ information facility ‘API’. R Package Version 0.9., 8. https://ropensci.org/. Accessed 21 August 2021
Chhetri, P. K., Gaddis, K. D., & Cairns, D. M. (2018). Predicting the suitable habitat of treeline species in the Nepalese Himalayas under climate change. Mountain Research and Development, 38(2), 153–163. https://doi.org/10.1659/MRD-JOURNAL-D-17-00071.1
Chug, D., Pathak, A., Indu, J., Jain, S. K., Jain, S. K., & Dimri, A. P., et al. (2020). Observed evidence for steep rise in the extreme flow of Western Himalayan Rivers. Geophysical Research Letters, 47(15), e2020GL087815. https://doi.org/10.1029/2020GL087815
Ciesla, W. M., & Donaubauer, E. (1994). Decline and dieback of trees and forests gIoia overview Decline and dieback of trees and forests A global overview.
Dakhil, M. A., Xiong, Q., Farahat, E. A., Zhang, L., Pan, K., Pandey, B., et al. (2019). Past and future climatic indicators for distribution patterns and conservation planning of temperate coniferous forests in southwestern China. Ecological Indicators, 107, 105559. https://doi.org/10.1016/j.ecolind.2019.105559
Dorman, M., Svoray, T., Perevolotsky, A., & Sarris, D. (2013). Forest performance during two consecutive drought periods: Diverging long-term trends and short-term responses along a climatic gradient. Forest Ecology and Management, 310, 1–9. https://doi.org/10.1016/j.foreco.2013.08.009
Elith, J., & Leathwick, J. R. (2009). Species distribution models: Ecological explanation and prediction across space and time. Annual Review of Ecology, Evolution, and Systematics, 40(1), 677–697. https://doi.org/10.1146/annurev.ecolsys.110308.120159
Fang, J., Yu, G., Liu, L., Hu, S., & Chapin, F. S. (2018). Climate change, human impacts, and carbon sequestration in China. Proceedings of the National Academy of Sciences, 115(16), 4015–4020. https://doi.org/10.1073/pnas.1700304115
Fargione, J. E., Bassett, S., Boucher, T., Bridgham, S. D., Conant, R. T., & Cook-Patton, S. C., et al. (2018). Natural climate solutions for the United States. Science Advances, 4(11). https://doi.org/10.1126/sciadv.aat1869
Farjon, A. (2010). A Handbook of the World’s Conifers. A Handbook of the World’s Conifers. Brill. https://doi.org/10.1163/9789004324510_001
Farjon, A. (2017). A Handbook of the World’s Conifers (2nd edition). Brill. https://doi.org/10.1163/9789004324510_001
Farr, T. G., Rosen, P. A., Caro, E., Crippen, R., Duren, R., & Hensley, S., et al. (2007). The shuttle radar topography mission. Reviews of Geophysics, 45(2), RG2004. https://doi.org/10.1029/2005RG000183
Fick, S. E., & Hijmans, R. J. (2017). WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. International Journal of Climatology, 37(12), 4302–4315. https://doi.org/10.1002/joc.5086
Fielding, A. H., & Bell, J. F. (1997). A review of methods for the assessment of prediction errors in conservation presence/absence models. Environmental Conservation, 24(1), 38–49. https://doi.org/10.1017/S0376892997000088
Fournier, A., Barbet-Massin, M., Rome, Q., & Courchamp, F. (2017). Predicting species distribution combining multi-scale drivers. Global Ecology and Conservation, 12, 215–226. https://doi.org/10.1016/j.gecco.2017.11.002
Gaire, N. P., Koirala, M., Bhuju, D. R., & Borgaonkar, H. P. (2014). Treeline dynamics with climate change at the central Nepal Himalaya. Climate of the past, 10(4), 1277–1290. https://doi.org/10.5194/cp-10-1277-2014
Gaire, N. P., Bhuju, D. R., Koirala, M., Shah, S. K., Carrer, M., & Timilsena, R. (2017). Tree-ring based spring precipitation reconstruction in Western Nepal Himalaya since AD 1840. Dendrochronologia, 42, 21–30. https://doi.org/10.1016/J.DENDRO.2016.12.004
Gao, C., Knorr, K.-H., Yu, Z., He, J., Zhang, S., Lu, X., & Wang, G. (2016). Black carbon deposition and storage in peat soils of the Changbai Mountain, China. Geoderma, 273, 98–105. https://doi.org/10.1016/j.geoderma.2016.03.021
Garzón, M. B., Robson, T. M., & Hampe, A. (2019). ΔTraitSDMs: Species distribution models that account for local adaptation and phenotypic plasticity. New Phytologist, 222(4), 1757–1765. https://doi.org/10.1111/NPH.15716
Giorgi, F., & Lionello, P. (2008). Climate change projections for the Mediterranean region. Global and Planetary Change, 63(2–3), 90–104. https://doi.org/10.1016/j.gloplacha.2007.09.005
Gordon, J. E. (2018). Mountain geodiversity: characteristics, values and climate change. in C Hoorn, A Perrigo & A Antonelli (eds), Mountains, Climate and Biodiversity. . Wiley-Blackwell, Chichester, pp. 137–154.
Griscom, B. W., Adams, J., Ellis, P. W., Houghton, R. A., Lomax, G., Miteva, D. A., et al. (2017). Natural climate solutions. Proceedings of the National Academy of Sciences of the United States of America, 114(44), 11645–11650. https://doi.org/10.1073/PNAS.1710465114/-/DCSUPPLEMENTAL
Halofsky, J. E., Peterson, D. L., Karen Dante-Wood, S., & Hoang, L. (2018). Toward climate-smart resource management in the Northern Rockies. In Advances in Global Change Research (Vol. 63, pp. 221–228). Springer International Publishing. https://doi.org/10.1007/978-3-319-56928-4_12
Hamid, M., Khuroo, A. A., Malik, A. H., Ahmad, R., Singh, C. P., Dolezal, J., & Haq, S. M. (2020). Early evidence of shifts in alpine summit vegetation: A case study from Kashmir Himalaya. Frontiers in Plant Science, 11, 421. https://doi.org/10.3389/fpls.2020.00421
Hijmans, R. J., Schreuder, M., De La Cruz, J., & Guarino, L. (1999). Using GIS to check co-ordinates of genebank accessions. Genetic Resources and Crop Evolution, 46(3), 291–296. https://doi.org/10.1023/A:1008628005016
Hijmans, R. J., Williams, E., & Vennes, C. (2019). Spherical trigonometry for geographic applications [R Package ’geosphere’].
Hoffmann, A. A., & Sgrò, C. M. (2011). Climate change and evolutionary adaptation. Nature, 470(7335), 479–485. https://doi.org/10.1038/nature09670
Hunt, K. M. R., Turner, A. G., & Shaffrey, L. C. (2019). Falling trend of western disturbances in future climate simulations. Journal of Climate, 32(16), 5037–5051. https://doi.org/10.1175/JCLI-D-18-0601.1
Immerzeel, W. W., Lutz, A. F., Andrade, M., Bahl, A., Biemans, H., Bolch, T., et al. (2020). Importance and vulnerability of the world’s water towers. Nature, 577(7790), 364–369. https://doi.org/10.1038/s41586-019-1822-y
IPCC. (2021). Climate change 2021: The physical science basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Masson-Delmotte, V. Zhai, P. Pirani, A. Connors, S. L. Péan, C. Berger, S. Caud, N. Chen, Y. . (V. Masson-Delmotte, P. Zhai, A. Pirani, S. L. Connors, C. Péan, S. Berger, et al., Eds.). Cambridge University Press. In Press. https://www.ipcc.ch/. Accessed 12 September 2021
Jha, M. N., Rathore, R. K., & Pande, P. (1984). Soil factor affecting the natural regeneration of silver fir and spruce in Himachal Pradesh. Indian Forester, 110(3), 293–298. http://ischolar.info/index.php/indianforester/article/view/10475. Accessed 10 September 2021
Jump, A. S., Hunt, J. M., & Penuelas, J. (2006). Rapid climate change-related growth decline at the southern range edge of Fagus sylvatica. Global Change Biology, 12(11), 2163–2174. https://doi.org/10.1111/j.1365-2486.2006.01250.x
Kharal, D. K., Thapa, U. K., & St. George, S., Meilby, H., Rayamajhi, S., & Bhuju, D. R. (2017). Tree-climate relations along an elevational transect in Manang Valley, central Nepal. Dendrochronologia, 41, 57–64. https://doi.org/10.1016/J.DENDRO.2016.04.004
Körner, C., Jetz, W., Paulsen, J., Payne, D., Rudmann-Maurer, K., Spehn, M., & E. (2017). A global inventory of mountains for bio-geographical applications. Alpine Botany, 127(1), 1–15. https://doi.org/10.1007/s00035-016-0182-6
Körner, C., & Spehn, E. (2019). A Humboldtian View of Mountains. Science, 365(6458), 1061. https://doi.org/10.1126/SCIENCE.AAZ4161
Kotru, R. K., Shakya, B., Joshi, S., Gurung, J., Ali, G., Amatya, S., & Pant, B. (2020). Biodiversity conservation and management in the Hindu Kush Himalayan Region: Are transboundary landscapes a promising solution? Mountain Research and Development, 40(2), A15. https://doi.org/10.1659/MRD-JOURNAL-D-19-00053.1
Krishnan, R., Sabin, T. P., Madhura, R. K., Vellore, R. K., Mujumdar, M., Sanjay, J., et al. (2019). Non-monsoonal precipitation response over the Western Himalayas to climate change. Climate Dynamics, 52(7–8), 4091–4109. https://doi.org/10.1007/s00382-018-4357-2
Kumar, N., Yadav, B. P., Gahlot, S., & Singh, M. (2015). Winter frequency of western disturbances and precipitation indices over Himachal Pradesh, India: 1977–2007. Atmósfera, 28(1), 63–70. https://doi.org/10.1016/S0187-6236(15)72160-0
Kumar, P. (2012). Assessment of impact of climate change on Rhododendrons in Sikkim Himalayas using Maxent modelling: Limitations and challenges. Biodiversity and Conservation, 21(5), 1251–1266. https://doi.org/10.1007/s10531-012-0279-1
Lamsal, P., Kumar, L., Aryal, A., & Atreya, K. (2018). Invasive alien plant species dynamics in the Himalayan region under climate change. Ambio, 47(6), 697–710. https://doi.org/10.1007/s13280-018-1017-z
Lebourgeois, F., Rathgeber, C. B. K., & Ulrich, E. (2010). Sensitivity of French temperate coniferous forests to climate variability and extreme events (Abies alba, Picea abies and Pinus sylvestris ). Journal of Vegetation Science, 21(2), 364–376. https://doi.org/10.1111/j.1654-1103.2009.01148.x
Liang, Q., Xu, X., Mao, K., Wang, M., Wang, K., Xi, Z., & Liu, J. (2018). Shifts in plant distributions in response to climate warming in a biodiversity hotspot, the Hengduan Mountains. Journal of Biogeography, 45(6), 1334–1344. https://doi.org/10.1111/jbi.13229
Linares, J. C., & Camarero, J. J. (2012). Growth patterns and sensitivity to climate predict silver fir decline in the Spanish Pyrenees. European Journal of Forest Research, 131(4), 1001–1012. https://doi.org/10.1007/s10342-011-0572-7
Liu, C., Berry, P. M., Dawson, T. P., & Pearson, R. G. (2005). Selecting thresholds of occurrence in the prediction of species distributions. Ecography, 28(3), 385–393. https://doi.org/10.1111/j.0906-7590.2005.03957.x
Liu, C., White, M., & Newell, G. (2013). Selecting thresholds for the prediction of species occurrence with presence-only data. Journal of Biogeography, 40(4), 778–789. https://doi.org/10.1111/jbi.12058
Lobo, J. M., Jiménez-Valverde, A., & Real, R. (2008). AUC: A misleading measure of the performance of predictive distribution models. Global Ecology and Biogeography, 17(2), 145–151. https://doi.org/10.1111/j.1466-8238.2007.00358.x
Luce, C. H. (2018). Effects of climate change on snowpack, glaciers, and water resources in the Northern Rockies. In Advances in Global Change Research (Vol. 63, pp. 25–36). Springer, Cham. https://doi.org/10.1007/978-3-319-56928-4_3
Malik, R., & Sukumar, R. (2021). June–July temperature reconstruction of Kashmir Valley from tree rings of Himalayan pindrow fir. Atmosphere, 12(3), 410. https://doi.org/10.3390/atmos12030410
Manish, K., Telwala, Y., Nautiyal, D. C., et al. (2016). Modelling the impacts of future climate change on plant communities in the Himalaya: a case study from Eastern Himalaya, India. Modeling Earth Systems and Environment, 2, 92. https://doi.org/10.1007/s40808-016-0163-1
Mátyás, C., Beran, F., Dostál, J., Čáp, J., Fulín, M., & Vejpustková, M., et al. (2021). Surprising drought tolerance of fir (Abies) species between past climatic adaptation and future projections reveals new chances for adaptive forest management. Forests 2021, Vol. 12, Page 821, 12(7), 821. https://doi.org/10.3390/F12070821
McDougall, K. L., Khuroo, A. A., Loope, L. L., Parks, C. G., Pauchard, A., Reshi, Z. A., et al. (2011). Plant invasions in mountains: Global lessons for better management. Mountain Research and Development, 31(4), 380–387. https://doi.org/10.1659/MRD-JOURNAL-D-11-00082.1
Meher, J. K., Das, L., Benestad, R. E., & Mezghani, A. (2018). Analysis of winter rainfall change statistics over the Western Himalaya: The influence of internal variability and topography. International Journal of Climatology, 38, e475–e496. https://doi.org/10.1002/joc.5385
Mohapatra, J., Singh, C. P., Hamid, M., Verma, A., Semwal, S. C., Gajmer, B., et al. (2019). Modelling Betula utilis distribution in response to climate-warming scenarios in Hindu-Kush Himalaya using random forest. Biodiversity and Conservation, 28(8–9), 2295–2317. https://doi.org/10.1007/s10531-019-01731-w
Naimi, B., & Araújo, M. B. (2016). sdm: A reproducible and extensible R platform for species distribution modelling. Ecography, 39(4), 368–375. https://doi.org/10.1111/ecog.01881
Naimi, B., Hamm, N. A. S., Groen, T. A., Skidmore, A. K., & Toxopeus, A. G. (2014). Where is positional uncertainty a problem for species distribution modelling? Ecography, 37(2), 191–203. https://doi.org/10.1111/j.1600-0587.2013.00205.x
Naudiyal, N., Wang, J., Ning, W., Gaire, N. P., Peili, S., & Yanqiang, W., et al. (2021). Potential distribution of Abies, Picea, and Juniperus species in the sub-alpine forest of Minjiang headwater region under current and future climate scenarios and its implications on ecosystem services supply. Ecological Indicators, 121(November 2020), 107131. https://doi.org/10.1016/j.ecolind.2020.107131
Noroozi, J., Talebi, A., Doostmohammadi, M., Rumpf, S. B., Linder, H. P., & Schneeweiss, G. M. (2018). Hotspots within a global biodiversity hotspot - areas of endemism are associated with high mountain ranges. Scientific Reports, 8(1), 10345. https://doi.org/10.1038/s41598-018-28504-9
Ovaskainen, O., Skorokhodova, S., Yakovleva, M., Sukhov, A., Kutenkov, A., Kutenkova, N., et al. (2013). Community-level phenological response to climate change. Proceedings of the National Academy of Sciences, 110(33), 13434–13439. https://doi.org/10.1073/PNAS.1305533110
Palazzi, E., von Hardenberg, J., & Provenzale, A. (2013). Precipitation in the Hindu-Kush Karakoram Himalaya: Observations and future scenarios. Journal of Geophysical Research: Atmospheres, 118(1), 85–100. https://doi.org/10.1029/2012JD018697
Pandit, M. K. (2017). Life in the Himalaya: An ecosystem at risk. Harvard University Press.
Parmesan, C., & Yohe, G. (2003). A globally coherent fingerprint of climate change impacts across natural systems. Nature, 421(6918), 37–42. https://doi.org/10.1038/nature01286
Phillips, S. J., & Dudík, M. (2008). Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography, 31(2), 161–175. https://doi.org/10.1111/j.0906-7590.2008.5203.x
Piñol, J., Terradas, J., & Lloret, F. (1998). Climate warming, wildfire hazard, and wildfire occurrence in coastal eastern Spain. Climatic Change, 38(3), 345–357. https://doi.org/10.1023/A:1005316632105
Poudyal, K., Jha, P. K., Zobel, D. B., & Thapa, C. B. (2004). Patterns of leaf conductance and water potential of five Himalayan tree species. Tree Physiology, 24(6), 689–699. https://doi.org/10.1093/TREEPHYS/24.6.689
R Core Team. (2020). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/. https://www.r-project.org/
Rai, S., Dawadi, B., Wang, Y., Lu, X., Ru, H., & Sigdel, S. R. (2020). Growth response of Abies spectabilis to climate along an elevation gradient of the Manang valley in the central Himalayas. Journal of Forestry Research, 31(6), 2245–2254. https://doi.org/10.1007/S11676-019-01011-X
Ren, Y.-Y., Ren, G.-Y., Sun, X.-B., Shrestha, A. B., You, Q.-L., Zhan, Y.-J., et al. (2017). Observed changes in surface air temperature and precipitation in the Hindu Kush Himalayan region over the last 100-plus years. Advances in Climate Change Research, 8(3), 148–156. https://doi.org/10.1016/j.accre.2017.08.001
Root, T. L., Price, J. T., Hall, K. R., Schneider, S. H., Rosenzweig, C., & Pounds, J. A. (2003). Fingerprints of global warming on wild animals and plants. Nature, 421(6918), 57–60. https://doi.org/10.1038/nature01333
Roxy, M. K., Ritika, K., Terray, P., Murtugudde, R., Ashok, K., & Goswami, B. N. (2015). Drying of Indian subcontinent by rapid Indian Ocean warming and a weakening land-sea thermal gradient. Nature Communications, 6(1), 7423. https://doi.org/10.1038/ncomms8423
Rushforth, K. (1991). CONIFERS. Batsford Ltd.
Sabin, T. P., Krishnan, R., Vellore, R., Priya, P., Borgaonkar, H. P., Singh, B. B., & Sagar, A. (2020). Climate change over the Himalayas. In Assessment of Climate Change over the Indian Region (pp. 207–222). Singapore: Springer Singapore. https://doi.org/10.1007/978-981-15-4327-2_11
Salesa, D., & Cerdà, A. (2020). Soil erosion on mountain trails as a consequence of recreational activities. A comprehensive review of the scientific literature. Journal of Environmental Management, 271, 110990. https://doi.org/10.1016/j.jenvman.2020.110990
Sarris, D., Christodoulakis, D., & Korner, C. (2007). Recent decline in precipitation and tree growth in the eastern Mediterranean. Global Change Biology, 13(6), 1187–1200. https://doi.org/10.1111/j.1365-2486.2007.01348.x
Schickhoff, U., Bobrowski, M., Böhner, J., Bürzle, B., Chaudhary, R. P., Gerlitz, L., et al. (2015). Do Himalayan treelines respond to recent climate change? An evaluation of sensitivity indicators. Earth System Dynamics, 6(1), 245–265.
Sharma, C. M., Suyal, S., Ghildiyal, S. K., & Gairola, S. (2010). Role of physiographic factors in distribution of Abies pindrow (Silver Fir) along an altitudinal gradient in Himalayan temperate forests. The Environmentalist, 30(1), 76–84. https://doi.org/10.1007/s10669-009-9245-1
Sharma, E., Molden, D., Rahman, A., Khatiwada, Y. R., Zhang, L., & Singh, S. P., et al. (2019a). Introduction to the Hindu Kush Himalaya assessment. In The Hindu Kush Himalaya Assessment (pp. 1–16). Cham: Springer International Publishing. https://doi.org/10.1007/978-3-319-92288-1_1
Sharma, E., Molden, D., Rahman, A., Khatiwada, Y. R., Zhang, L., & Singh, S. P., et al. (2019b). Introduction to the Hindu Kush Himalaya assessment. In The Hindu Kush Himalaya Assessment (pp. 1–16). Cham: Springer International Publishing. https://doi.org/10.1007/978-3-319-92288-1_1
Shi, Y., Zhang, D.-F., Xu, Y., & Zhou, B.-T. (2018). Changes of heating and cooling degree days over China in response to global warming of 1.5 °C, 2 °C, 3 °C and 4 °C. Advances in Climate Change Research, 9(3), 192–200. https://doi.org/10.1016/j.accre.2018.06.003
Shrestha, K. B., Chhetri, P. K., Bista R. (2017). Growth responses of Abies spectabilis to climate variations along an elevational gradient in Langtang National Park in the central Himalaya Nepal. Journal of Forest Research 1–8 https://doi.org/10.1080/13416979.2017.1351508
Sigdel, S., Dawadi, B., Camarero, J., Liang, E., & Leavitt, S. (2018). Moisture-limited tree growth for a subtropical Himalayan conifer forest in Western Nepal. Forests, 9(6), 340. https://doi.org/10.3390/f9060340
Singh, S., Ghosh, S., Sahana, A. S., Vittal, H., & Karmakar, S. (2016). Do dynamic regional models add value to the global model projections of Indian monsoon? Climate Dynamics 2016 48:3, 48(3), 1375–1397. https://doi.org/10.1007/S00382-016-3147-Y
Singh, S. P. (2018). Research on Indian Himalayan Treeline Ecotone: An overview. Tropical Ecology, 59(2), 163–176. www.tropecol.com. Accessed 11 September 2021
de Souza, A. C., & Prevedello, J. A. (2021). Climate change and biological invasion as additional threats to an imperiled palm. Perspectives in Ecology and Conservation, 19(2), 216–224. https://doi.org/10.1016/j.pecon.2021.02.003
Sun, L., Luo, J., Qian, L., Deng, T., & Sun, H. (2020). The relationship between elevation and seed-plant species richness in the Mt. Namjagbarwa region (Eastern Himalayas) and its underlying determinants. Global Ecology and Conservation, 23, e01053. https://doi.org/10.1016/J.GECCO.2020.E01053
Swets, J. (1988). Measuring the accuracy of diagnostic systems. Science, 240(4857), 1285–1293. https://doi.org/10.1126/science.3287615
Telwala, Y., Brook, B. W., Manish, K., & Pandit, M. K. (2013). Climate-induced elevational range shifts and increase in plant species richness in a Himalayan biodiversity epicentre. PLoS ONE, 8(2), e57103. https://doi.org/10.1371/journal.pone.0057103
Thabeet, A., Vennetier, M., Gadbin-Henry, C., Denelle, N., Roux, M., Caraglio, Y., & Vila, B. (2009). Response of Pinus sylvestris L. to recent climatic events in the French Mediterranean region. Trees, 23(4), 843–853. https://doi.org/10.1007/s00468-009-0326-z
Turland, N., & Figueiredo, E. (1999). World checklist and bibliography of conifers; mistletoes of Africa. Botanical Journal of the Linnean Society, 130(2), 183–184. https://doi.org/10.1111/J.1095-8339.1999.TB00518.X
van der Maaten-Theunissen, M., Kahle, H.-P., & van der Maaten, E. (2013). Drought sensitivity of Norway spruce is higher than that of silver fir along an altitudinal gradient in southwestern Germany. Annals of Forest Science, 70(2), 185–193. https://doi.org/10.1007/s13595-012-0241-0
Vidakovic, M. (1993). Conifers: morphology and variation. Tree Physiology, 12(4), 319–319. https://doi.org/10.1093/treephys/12.4.319a
Viviroli, D., & Weingartner, R. (2008). “Water towers”—A global view of the hydrological importance of mountains. Advances in Global Change Research, 31, 15–20. https://doi.org/10.1007/978-1-4020-6748-8_2
Warren, D. L., Glor, R. E., & Turelli, M. (2008). Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution, 62(11), 2868–2883. https://doi.org/10.1111/j.1558-5646.2008.00482.x
Xiang, X., Cao, M., & Zhou, Z. (2007). Fossil history and modern distribution of the genus Abies (Pinaceae). Frontiers of Forestry in China, 2(4), 355–365. https://doi.org/10.1007/s11461-007-0058-4
Xu, J., Badola, R., Chettri, N., Chaudhary, R. P., Zomer, R., & Pokhrel, B., et al. (2019). Sustaining biodiversity and ecosystem services in the Hindu Kush Himalaya. In The Hindu Kush Himalaya Assessment (pp. 127–165). Cham: Springer International Publishing. https://doi.org/10.1007/978-3-319-92288-1_5
Yao, F., Huang, Y., Zhang, Y., Dong, Y., Ma, H., Deng, C., et al. (2012). Subclinical endothelial dysfunction and low-grade inflammation play roles in the development of erectile dysfunction in young men with low risk of coronary heart disease. International Journal of Andrology, 35(5), 653–659. https://doi.org/10.1111/J.1365-2605.2012.01273.X
Zhan, Y.-J., Ren, G.-Y., Shrestha, A. B., Rajbhandari, R., Ren, Y.-Y., Sanjay, J., et al. (2017). Changes in extreme precipitation events over the Hindu Kush Himalayan region during 1961–2012. Advances in Climate Change Research, 8(3), 166–175. https://doi.org/10.1016/j.accre.2017.08.002
Zhang, K., Zhang, Y., Zhou, C., Meng, J., Sun, J., Zhou, T., & Tao, J. (2019). Impact of climate factors on future distributions of Paeonia ostii across China estimated by MaxEnt. Ecological Informatics, 50, 62–67. https://doi.org/10.1016/j.ecoinf.2019.01.004
Zobel, D. B., & Singh, S. P. (1995). Tree water relations along the vegetational gradient in the Himalayas. Current Science, 68(7), 742–745. https://www.jstor.org/stable/24096660?seq=1#metadata_info_tab_contents. Accessed 10 September 2021
Zobel, D., Garkoti, S., Singh, S. P., Tewari, A., & Negi, C. (2001). Patterns of water potential among forest types of the central Himalaya. Current Science, 80(6), 774–779.
Zomer, R. J., Trabucco, A., Metzger, M. J., Wang, M., Oli, K. P., & Xu, J. (2014). Projected climate change impacts on spatial distribution of bioclimatic zones and ecoregions within the Kailash Sacred Landscape of China, India. Nepal. Climatic Change, 125(3–4), 445–460. https://doi.org/10.1007/s10584-014-1176-2
Acknowledgements
We thank the Head, Department of Botany, University of Kashmir for providing laboratory facilities. Support under the Centre with Potential for Excellence in Particular Areas (CPEPA) scheme by the University Grants Commission (UGC), New Delhi to the University of Kashmir is also gratefully acknowledged which helped in the conduct of present work as well.
Author information
Authors and Affiliations
Contributions
RAM, ZAR and IR conceived the research idea. IR collected the field data; RAM performed the species distribution modelling and ZAR and RAM wrote the paper. SPS gave valuable inputs in the introduction and discussion part of the manuscript.
Corresponding author
Ethics declarations
Ethical approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Malik, R.A., Reshi, Z.A., Rafiq, I. et al. Decline in the suitable habitat of dominant Abies species in response to climate change in the Hindu Kush Himalayan region: insights from species distribution modelling. Environ Monit Assess 194, 596 (2022). https://doi.org/10.1007/s10661-022-10245-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10661-022-10245-y