
Abstract
Despite the importance of assessing beta diversity to understand the effects of human modifications on biological communities, there are almost no studies that properly addressed how beta diversity varies along anthropogenic gradients. We developed an algorithm to calculate beta diversity among a set of sites included in a moving window along any given environmental gradient. This allowed us to assess beta diversity among sites with similar conditions in terms of human modifications (e.g., land use or instream degradation). We investigated beta diversity using stream fish community data and indicators of human modification quantified at four spatial scales (whole catchment, riparian, local, and instream). Variation in beta diversity was dependent on the scale of human modifications (catchment, riparian, local, instream, and all four scales combined) and on the type of diversity considered (taxonomic or functional). We also found evidence for non-linear responses of both taxonomic and functional beta diversity to human-induced environmental alterations. Therefore, the response of beta diversity was more complex than expected, as it depended on the scale used to quantify human impact and exhibited opposite responses depending on the location along the environmental impact gradient and on whether the response was taxonomic or functional diversity. Anthropogenic modifications can introduce unexpected variability among stream communities, which means that low beta diversity may not necessarily indicate a degraded environmental condition and high beta diversity may not always indicate a reference environmental condition. This has implications for how we should consider beta diversity in environmental assessments.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Changes in the landscape due to human activities, such as agriculture, urbanization, or forestry, have several consequences for riverine ecosystems. Land use influences the input of nonpoint pollutants into streams through surface runoff, augmenting fine sediment, nutrients, and pesticides (Lowrance et al. 1984; Allan 2004). In addition, land conversion may increase stream temperature (Macedo et al. 2013) and modify stream flow and the hydrological regime (Allan 2004). These multiple stressors can be exacerbated depending on the spatial scale of human modifications; for instance, whether they reach the riparian zone (King et al. 2005; Dala-Corte et al. 2016) or the instream physical habitat structure (Sweeney et al. 2004). Also, degradation of the riparian zones can impact freshwater fish communities both in terms of taxonomic and functional composition (Jones et al. 1999; Zeni and Casatti 2014; Giam et al. 2015). Altogether, land use may cause changes in the functioning and services of stream ecosystems.
Studies evaluating the effect of land use on stream fish have detected distinct responses of alpha diversity (e.g., species richness). For instance, the number of stream fish species may decrease in agriculture-disturbed watersheds, mainly because of changes in the quality of instream habitat (Roth et al. 1996; Walser and Bart 1999; Johnson and Angeler 2014). On the other hand, it has been suggested that the environmental changes caused by agriculture may increase species richness at intermediate levels of land use gradient (Dala-Corte et al. 2016). One of the possible mechanisms explaining this latter result is the increase in species richness due to “native invasion” (Scott and Helfman 2001; Lorion and Kennedy 2009), consisting of the colonization of impacted streams by native fish species typical of downstream sections of the drainage network. Whereas the alpha diversity of stream fish can respond either negatively or positively to land use, less is known about whether agricultural land use increases or decreases community variability among streams, i.e., beta diversity.
Habitat alterations might be expected to lead to faunal homogenization, i.e., when biological dissimilarity decreases between communities (Walters et al. 2003; Pool and Olden 2012). For instance, fish beta diversity among streams subjected to agricultural land use and with a high concentration of phosphorus was reduced in relation to a group of reference streams (Johnson and Angeler 2014). Different mechanisms are involved in faunal homogenization (Rahel 2002), such as the invasion or introduction of non-native species, the local extinction of rare/specialist species and the concomitant spread of common/generalist species (Olden and Poff 2004; Hermoso et al. 2012). In riverine systems, the latter mechanism has been recorded in disturbed regions, where widespread fish species present in downstream sections colonize upstream streams (Walters et al. 2003). Increased siltation, temperature, and nutrients can favor the occurrence of tolerant and widespread fish species, usually encountered in warmer, more turbid, sediment- and nutrient-rich streams in the lower portions of watersheds (Scott and Helfman 2001). In this case, agricultural land use could decrease the beta diversity of fish communities both among small streams and among small and large streams.
Changes in fish species composition owing to human impacts can also modify functional beta diversity. For instance, agriculture is expected to reduce benthic specialists due to the input of fine sediment (Jones et al. 1999; Walser and Bart 1999). The impact on benthic fish by siltation may be exacerbated if the riparian zone is impacted (Dala-Corte et al. 2016). Shade reduction owing to the removal or degradation of riparian woody vegetation coupled with increased nutrients from agriculture may cause the proliferation of algae and macrophytes (Lowrance et al. 1997; Burrell et al. 2014) and allows colonization by nektonic species (Dala-Corte et al. 2016). In addition, riparian clearance reduces the input of leaf litter and wood debris—coarse organic particulate matter—into streams (Hyatt and Naiman 2001; Paula et al. 2013) and changes the proportion of autochthonous and allochthonous organic matter input into aquatic food webs (Minshall 1978; Nakano et al. 1999; Majdi et al. 2015). These modifications can benefit the occurrence of nektonic and tolerant fish species to the detriment of benthic species (Casatti et al. 2015; Dala-Corte et al. 2016). Therefore, an important question is whether change in species composition is associated with the loss of functional beta diversity in riverine ecosystems.
Research on beta diversity has historically focused on assessing whether dissimilarity among communities can be explained by or correlated with environmental dissimilarity (usually based on abiotic characteristics) (e.g., Nekola and White 1999; Legendre et al. 2005; Melo et al. 2009). This can be achieved by using methods such as correlation among matrices (e.g., Mantel) or constrained analyses (e.g., RDA, CCA) (Legendre et al. 2005; Tuomisto and Ruokolainen 2006). Another important but understudied facet of beta diversity is how variation in community dissimilarity relates to a particular location along an environmental gradient (see an example using productivity in Andrew et al. 2012). For instance, two pairs of sites may be equally dissimilar (and equally distant in geographical space), but one pair may be located in an undisturbed portion of a gradient, and the other in a disturbed one. If human disturbances cause environmental homogenization, lower beta diversity could be expected between the pair of sites located in the disturbed portion than in the pair located in the undisturbed portion. As an alternative, beta diversity could be high in the disturbed pair if sites differ in the nature of their disturbances (e.g., agriculture and forestry) or if one of the sites in the pair receives a constant influx of downstream migrants (mass effect, Heino et al. 2015). Accordingly, such an approach is particularly pertinent for understanding how anthropic activities affect beta diversity as an ecological indicator and to define thresholds of environment modifications at different spatial scales.
We investigated how the beta diversity of stream fish communities responds to human alterations measured at multiple scales. Specifically, we asked the following: (1) How does the beta diversity of stream fish vary along a land-use gradient? (2) Is the response of beta diversity dependent on the spatial scale (catchment, riparian, local, instream, and all four combined) at which human-induced alterations are measured? (3) Do taxonomic and functional beta diversities respond differently to agricultural land use? To answer these questions, we quantified the beta diversity for sets of similar streams and regressed it against gradients of human impacts at four spatial scales plus geographic distance as a control variable. The aim was to provide insight into the multiple effects of land use on the diversity of freshwater ecosystems. Furthermore, the method employed here can be used to model beta diversity as an ecological indicator of any other type of environmental disturbance or geographic gradient.
Methods
Study area and fish sampling
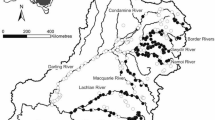
We sampled 54 wadeable streams distributed across a subtropical grassland region in southern Brazil (Fig. 1). The studied area covered a convex hull polygon of 107.497 km2. This area comprises the Pampa lowland grasslands and also the high altitude grasslands of the grassland-forest mosaic situated at the southern limits of the Atlantic Forest biome in southern Brazil. The regional climate type is humid subtropical with hot summers and mild to cool winters, without clear rainy or dry seasons (Cfa type) according to the Köppen-Geiger classification. Although grassland is the dominant vegetation across the landscape, riparian zones commonly develop woody vegetation composed of shrubland and forest.

Location of the 54 streams sampled in the Brazilian Pampa grassland region
Agricultural land use has expanded in south Brazilian grasslands over the last three decades, with a large proportion of native vegetation being converted to agriculture and forestry (mainly soybean, rice, maize and wheat production, or Pinus and Eucalyptus plantations; Overbeck et al. 2007). In addition, cattle and sheep farming have been traditional activities since the seventeenth century, with beef production being widespread (Overbeck et al. 2007). Therefore, streams are susceptible to different levels of environmental disturbances stemming from agricultural land use. To avoid potential confounding factors, all of the sampled streams had less than 1% urban land use in the upland catchment area.
Fish communities were sampled using a standardized sampling effort by electrofishing in a 150 m long reach in each of the 54 studied streams. The sampled reaches always comprised different types of habitats, such as riffles, glides, and pools. Single-pass electrofishing with a mean sampling effort of 3 h was carried out in each stream reach using a 1500 W 150–300/300–600 V DC generator (EFKO GmbH model FEG 1500). Samples were restricted to small wadeable streams (mean wetted width = 4.7 m, SD = 1.8 m) of second (16), third (29), and fourth (9) Strahler orders (Strahler 1952). Field expeditions were carried out during spring and summer between October 2013 and March 2015. Fishes were anesthetized with clove oil and subsequently preserved in 4% formaldehyde in accordance with national ethical guidelines (CEUA-UFRGS #24433) and with pre-approved permits (SISBIO #39672-1).
Functional traits
We estimated the functional diversity of each community by measuring morphological traits of fish species related to habitat preference and to feeding behavior (e.g., Albouy et al. 2011). We expected impacted streams to present altered physical instream habitat and trophic structure, driving responses in functional trait diversity and composition related to habitat preference and feeding behavior. Firstly, we measured 21 morphometric variables of 10 individuals of each fish species, or measured all individuals for species with fewer than 10 captured specimens. Then, we used these morphological measures to calculate 14 functional traits, which consisted of percentage values relative to body or head length or mass. The functional traits included were body compression, body depth, head size, eye position, eye size, mouth position, peduncle length, peduncle compression, pectoral position, pectoral fin area, ventral fin area, caudal fin area, dorsal fin area, and biomass. See Supporting Information (Table S1 and Fig. S2) in Dala-Corte et al. (2016) for details on the morphological measures and a description of the functional traits.
Indicators of human modification
We quantified human modification at four spatial scales and using the combination of all four scales (Fig. 2): (1) percentage of cropland area in the catchment above the sampled reach (catchment cropland); (2) percentage of non-natural riparian vegetation cover, measured along a 50-m wide buffer up to 1 km upstream from the sampling reach (hereafter for simplicity termed upstream riparian deforestation); (3) degradation at the local scale measured on the banks of the sampled reaches and then square root transformed (local impact); (4) instream modification related to substrate composition and homogeneity; and (5) the combination of all these scales by summing the standardized variables (0–1). High values of these variables indicate a high degree of anthropogenic modification (Table 1).

Spatial scales used to assess the effects of land use, including catchment, riparian, local (sample reach), and instream scales. Catchment scale comprised cropland percentage in relation to the total upland area for each sample site. Riparian scale comprised deforestation percentage in a 1 km long 50 m wide buffer along the stream network upstream. Local scale comprised the percentage of alteration by agriculture (visually estimated in situ) immediately adjacent to the sample reach. Instream scale included variables related to substrate modifications
Estimates of percent catchment cropland cover and non-natural upstream riparian vegetation cover were based on a supervised classification of 5-m resolution RapidEye satellite images for 2012 (Geo Catálogo 2015), as detailed in Dala-Corte et al. (2016). The degree of local agricultural effect at the reach scale (local impact) was assessed in situ on the sampled reach margins, during the field samplings (based on Kaufmann et al. 1999).
Local impact was visually estimated as the percentage cover of the riparian zone that was under agricultural use on both stream margins along the 150-m sampled reaches and the density of signals left by livestock use (trampling, manure, and animal visualization) up to 10-m from the stream bank at the 11 cross sections of each sampled reach. Cropland cover percentage and livestock use density were visually classified into four percentage classes: absent (0%), sparse (< 10%), moderate (10–40%), heavy (40–75%), and very heavy (> 75%). The mean value of each percentage class was assigned to each cross section evaluation on both stream margins to calculate mean local impact values for each sampled reach.
Instream habitat modification was estimated using four variables previously known to affect stream fish alpha diversity (Dala-Corte et al. 2016). The amount of coarse particulate organic (CPOM) and substrate grain size composition was quantified following the protocol of Kaufmann et al. (1999). This consisted of visual estimations at 11 cross sections in each 150-m stream segment where we sampled fish. These 11 records of each variable were used to calculate average values per sampled reach. CPOM included the amount of leaf litter and woody debris covering the stream bottom. Substrate percentage cover included the classes bedrock, boulder, cobble, large pebble, small pebble, sand, silt, and hardpan (compacted clay). Detailed descriptions of the method can be found in Dala-Corte et al. (2016). The final instream habitat modification indicator was calculated as the sum of the four instream metrics standardized by range: (1) percent of fine sediment cover, (2) substrate homogenization (inverse of Shannon diversity of substrate classes), (3) instream area without leaf litter, and (4) instream area without woody debris. Therefore, all these instream metrics indicated physical homogenization of the habitat.
Beta diversity along human modification gradients
We developed a new approach to calculate beta diversity along environmental gradients. Beta diversity values for each site comprised the dissimilarity values of the focal site in relation to its two neighboring sites (window size of three) along the gradients of human modifications. Values were calculated separately for each of the five gradients of human modifications (catchment, riparian, local, instream, and all four combined). For this, we arranged sites from the lowest to the highest values of each human modification gradient and calculated the beta diversity of a focal site in relation to its neighboring sites in the gradient. In other words, the beta diversity value in each window was obtained by comparing community composition in the focal site with its two neighboring sites included in a moving window of the gradient under study (Fig. 3). This process allowed us to determine whether sites equally and highly impacted by land use have higher or lower beta diversity than sites with equally low impacts.

Depiction of how beta diversity was calculated along an environmental gradient for each focal site in relation to its neighboring sites. Each number represents a focal site selected sequentially (a–d) along the environmental gradient in each round. In this case, window size was equal to three, as focal sites were compared only with their two neighbors (arrowheads) in the environmental gradient. For this, sites were ordered from lowest to highest values according to the environmental gradient. Note that extreme values in the environmental gradient were never computed as focal sites because of the absence of neighboring sites with lower or higher values
We obtained beta diversity values by coding an algorithm that performed the following steps: (i) sort sites in the sites-by-species matrix (or sites-by-traits matrix when calculating functional diversity) according to a given human modification gradient, (ii) select neighboring sites according to a window size of three (the selected sites were always the one site scored immediately lower and the one site scored immediately higher than the focal site in the environmental gradient), and (iii) calculate beta diversity metrics for each site window (see below). We repeated steps ii and iii for each set of sites in a window to obtain beta diversity values for each focal site. Beta diversity values were only obtained for windows with complete sets of neighboring sites in the window (i.e., not for focal sites in the extremities of the gradient). Thus, a total of 52 beta diversity values were obtained from the 54 sampled sites (Fig. 3).
The moving window approach allowed us to build models using beta diversity as a continuous variable to test against predictors. However, sites selected in a moving window varied according to the gradient under investigation (i.e., the human modification indicators representing each of the five gradients). This is because sites selected in the moving window are dependent on the gradient used to order them. For this reason, we generated models individually for each of the indicators that we hypothesized that could shape beta diversity. This function we coded in the statistical environment R (R Core Team 2017) to obtain beta diversity values is available in the CommEcol package (Melo 2018) and also in the Supplementary Material (Supp. S1).
Beta diversity metrics
Beta diversity assigned to each focal site was obtained by the mean dissimilarity of it in relation to each of its two neighboring sites in the ordered gradient, for both taxonomic and functional composition data. For taxonomic beta diversity, we calculated dissimilarity for both presence-absence and log-transformed abundance data using the Sorensen dissimilarity and its quantitative version, the Bray-Curtis dissimilarity, respectively. For functional beta diversity, we first obtained the community-weighted mean of trait values per site (CWM) with the package FD (Laliberté et al. 2014). We used presence-absence as well as abundance data to weight traits in the CWM. Then, we used the Euclidean distance to calculate functional beta diversity. We employed distinct resemblance metrics because taxonomic and functional data consisted of distinct data types (presence/absence and abundance data for taxonomic matrix and continuous morphometric measures data without zeros for functional matrix). The dissimilarity calculations were carried out using the package vegan (Oksanen et al. 2017).
Effects of human modifications on beta diversity
The responses of taxonomic and functional stream fish beta diversities to human modification gradients were modeled with multiple regression models. We generated separate models for each gradient and for taxonomic and functional beta diversities. In each model, the explanatory variables included were one of the human gradient variables plus geographic distance. Because the effect of human modifications on diversity can be nonlinear, we also included a quadratic term. We used the function poly (R Core Team 2017), which centers both the linear and quadratic terms to remove their collinearity. Geographic distance was included to control for the known relationship of spatial distance decay in similarity (Nekola and White 1999). For instance, sites near to each other in the space are expected to have similar faunas (low beta diversity) or similar disturbance degree, so including geographical distance in the models was important to control for any spatial structure. Geographic distance was calculated using the mean Euclidean distance among focal sites in relation to their two neighbors in each window along the gradients.
We fitted a total of six models for each combination of beta diversity and human modification gradient. Combinations of explanatory variables in these models were (i) a null expectation fitted with an intercept-only model for comparison, (ii) geographic distance, (iii) the linear term of a human modification variable, (iv) the linear and the quadratic form of a human modification variable, (v) the linear human modification variable and the geographic distance, and (vi) the linear and quadratic human modification variable and the geographic distance.
We checked the models for multicollinearity using variance inflation factors analysis—VIF (Fox and Monette 1992), available in the package car (Fox and Weisberg 2011). None of the models presented important multicollinearity, as values of the square root of VIF were inferior to 2, so we kept all variables in the models. Subsequently, we compared the goodness of fit of all six models for each response variable and gradient using the second-order Akaike Information Criterion (AICc). For this, we used the package bbmle (Bolker 2017) for R (R Core Team 2017).
Results
Fish community composition
We sampled a total of 106 fish species at 54 stream sites in the southern Brazilian grasslands. The mean number of species per site was 18.5 (SD 6.7), ranging from 6 to 32 species. Eight species were highly frequent and occurred at more than 50% of the sampled sites (Heptapterus mustelinus, Bryconamericus iheringii, Characidium pterostictum, Rineloricaria stellata, Astyanax laticeps, Rhamdia quelen, Pseudocorynopoma doriae, and Crenicichla lepidota). On the other hand, 67 species were restricted to less than 10% of the sampled sites. Regarding species abundance, only four species had a relative abundance higher than 5% in relation to the total [Bryconamericus iheringii (19.2%), Heptapterus mustelinus (16.4%), Characidium pterostictum (5.9%), and Diapoma alegretensis (5.7%)]. All 106 species found are native to the studied region.
Taxonomic beta diversity
Taxonomic beta diversity using species presence-absence data was best fitted by quadratic models including only catchment cropland or riparian deforestation (ΔAICc > 2.00) (Table 2; Fig. 4a, c). For species abundance data, however, only the quadratic model of catchment cropland was important (Table 3; Fig. 4b). The models showed a hump-shaped effect of catchment cropland or riparian deforestation on taxonomic beta diversity, with the highest beta diversity values at intermediate levels of land use (Fig. 4a–c). There was no support for the effect of human modifications measured at the remaining spatial scales (Tables 2 and 3; Fig. 4).

Effects of human modifications on taxonomic beta diversity based on presence-absence (a, c, e, g, i) and abundance (b, d, f, h, j) data of fish communities in subtropical streams of south Brazil. Explanatory variables were human modifications measured at four spatial scales (Catchment, Riparian, Local, and Instream) and all of them combined into a single index (Index). Fitted lines represent models containing only human modifications selected as the best models using ΔAICc, as shown in Tables 2 and 3
Functional beta diversity
Like taxonomic beta diversity, the quadratic model including only catchment cropland was selected as the most probable for explaining functional beta diversity for both species presence-absence and abundance data (Tables 4 and 5). However, the quadratic fit was positive, indicating a U-shaped effect of catchment cropland on functional beta diversity, with the lowest values at intermediate levels of land use (Fig. 5a, b). The quadratic models including instream modifications were also important and also showed a U-shaped relationship to functional beta diversity (Tables 4 and 5; Fig. 5g, h). Furthermore, local impact had a positive linear effect on functional beta diversity, but only in the model based on species abundance data (Fig. 5f).

Effects of human modifications on functional beta diversity based on presence-absence (a, c, e, g, i) and abundance (b, d, f, h, j) data of fish communities in subtropical streams of south Brazil. Explanatory variables were measured at four spatial scales (Catchment, Riparian, Local, and Instream) and all combined into an index (Index). Fitted lines represent models containing only human modifications selected as the best models using ΔAICc, as shown in Tables 4 and 5
Discussion
Fish community dissimilarity among streams was dependent on the spatial scale at which we measured land use and habitat (catchment, riparian, local, or instream) and on the type of diversity considered (whether taxonomic or functional). The clearest pattern observed was that catchment cropland and riparian deforestation have a hump-shaped effect on taxonomic beta diversity. This means that dissimilarity in fish community composition between streams was highest at intermediate levels of cropland at the catchment scale or deforestation at the riparian scale. This suggests that human modifications of the landscape may cause a subsidy-stress effect on fish community beta diversity (Odum et al. 1979). In addition, functional beta diversity showed a U-shaped response to catchment cropland and to instream modification, indicating that the gain in taxonomic dissimilarity at intermediate levels of modifications is coupled with increased functional redundancy. These results provide insights that help in understanding the complex drivers of stream fish beta diversity at the landscape scale and show a different way of using beta diversity as an indicator of environmental stressors.
The hump-shaped relation of cropland at the catchment scale and deforestation at the riparian scale with taxonomic beta diversity reveals the complexity of the effects of landscape modifications on aquatic environments. Agricultural land use at multiple scales has been acknowledged to change several instream physical, chemical, and biological conditions (Sweeney et al. 2004; Dala-Corte et al. 2016). For example, conversion of the landscape to croplands is associated with nutrient enrichment, water turbidity, stream bottom siltation, reduced shading, and increased temperature (Allan 2004; Niyogi et al. 2007; Macedo et al. 2013; Burrell et al. 2014). Whereas these environmental conditions are stressful for various fish species, such as those found in clear/cold waters and benthic specialists of rocky substrates (Jones et al. 1999; Walser and Bart 1999; Dala-Corte et al. 2016), several slow/warm-water, nektonic, and tolerant fish species may be favored (Casatti et al. 2015; Dala-Corte et al. 2016). Thus, the expanded distribution of these species, which are common in warm and larger streams, may be one of the consequences that alters fish beta diversity in small streams and headwaters (Scott and Helfman 2001; Lorion and Kennedy 2009; Teixeira-de Mello et al. 2015). In fact, we found in a previous study that catchment cropland and deforestation of the riparian zone increased local species richness by the addition of slow-water and nektonic fishes, while lithophilic species were negatively affected (Dala-Corte et al. 2016). Therefore, it is possible that increasing agriculture in the catchment area or deforestation in the riparian buffer increases fish beta diversity at intermediate levels by promoting the random gain of widespread species not common to small streams. However, high levels of alteration (> 40%) act as a stressor and reduce taxonomic beta diversity, as postulated in the subsidy-stress hypothesis (Odum et al. 1979).
Other factors that may explain high taxonomic beta diversity at intermediate levels of cropland in the catchment are the distinct effects of different crop cultures (e.g., soybeans, maize, rice), the spatial location of the farming within the watershed (i.e., spatial configuration), and time since natural cover conversion to agriculture, as well as the management practice adopted by the landowners. All these factors could create variation in environmental conditions along the drainage network and thus generate variation in fish community composition. The length of time since land conversion, for example, may be quite long (~ 1940) or short in the study area (Overbeck et al. 2007), generating regional variation in the fish fauna as well. These factors should be of minor importance in basins that have retained most of the natural vegetation or those in which most of the natural vegetation has been substituted by crop cultures (the extremities of the gradient). Further studies are needed to determine whether these aforementioned processes are responsible for the hump-shaped response observed in fish beta diversity.
Riparian deforestation was important for predicting taxonomic beta diversity based on species presence-absence data, but not on abundance data. This suggests that human impacts in the upstream riparian buffer may have a stronger effect on fish species composition than on species abundance. Therefore, the hump-shaped effect of riparian deforestation on taxonomic beta diversity indicates that low to intermediate levels of deforestation in upstream riparian zones maintain taxonomically distinct fish communities. High levels of degradation of riparian vegetation may cause reduction in beta diversity and loss of several processes essential for freshwater ecosystems. For instance, riparian vegetation provides food, such as fruits and terrestrial insects, that is important to both specialized and generalist fish species that feed on allochthonous items (Allan et al. 2003; Dala-Corte et al. 2017). Submerged tree roots on the stream banks and wood debris provide refuge for several specialized fish species, in addition to creating heterogeneous hydraulic microhabitats (Wright and Flecker 2004). Riparian shading reduces water temperature and also serves as a refuge for other fish species (Macedo et al. 2013). Furthermore, riparian integrity contributes to stream bank stability and reduces stream bottom siltation (Jones et al. 1999; Rabeni and Smale 1995). Thus, our results extend to beta diversity the recommendation of other studies on the importance of upstream riparian integrity for maintaining alpha diversity (Jones et al. 1999; Giam et al. 2015; Dala-Corte et al. 2016).
Unlike the effect on taxonomic beta diversity, we observed a U-shaped effect of catchment cropland and instream modification on functional beta diversity, both for species presence-absence and abundance data. The local impact indicator was an exception, which showed a positive linear (but weak) relation with functional beta diversity. Therefore, streams with intermediate levels of cropland at the catchment scale harbored fish communities with greater taxonomic dissimilarity but higher functional redundancy (low functional beta diversity). Likewise, functional redundancy was higher at intermediate levels of instream modification. Siltation of the streambed and homogenization of the substrate, including reductions in leaf litter and woody debris, may impair mainly the lithophilic and benthic specialists and lead to lower functional diversity (Giam et al. 2015; Dala-Corte et al. 2016). On the other hand, we observed an unexpected result, which was that streams with high values of catchment cropland and instream modification presented high functional beta diversity, similar to streams with low values of these indicators of human modifications. This counterintuitive relationship may be partially explained by low species richness in highly impacted streams (Dala-Corte et al. 2016). These highly impacted streams usually harbor only a few tolerant species, but some of them may be very distinct morphologically, such as the elongated Synbranchus marmoratus and the armored catfishes Callichthys callichthys, Otocinclus flexilis, and Corydoras paleatus. This may explain the U-shaped relation between human modifications and functional beta diversity. In addition, the linear and positive relation between functional beta diversity and local impact may indicate that impacts made locally have a stronger effect in driving functional dissimilarity among streams.
Although low amounts of cropland at the catchment scale may initially cause increases in taxonomic beta diversity, our study suggests that agriculture may disrupt fish beta diversity by changing the fish species composition expected in small streams. For instance, the functional composition of these intermediate impacted streams may be low due to the replacement of morphologically distinct species by a higher number of similar nektonic fish (Dala-Corte et al. 2016). Also, changes in species composition could mean the replacement of endemic species typical of headwaters by a larger number of widespread fish species typical of downstream reaches, or simply by the addition of these latter species to communities of preserved streams (Scott and Helfman 2001). Therefore, other community diversity characteristics not evaluated here, such as unique functions and the proportion of rare or endemic species (which are often found in pristine streams), can be negatively affected even by low levels of landscape modification (e.g., Zeni and Casatti 2014; Casatti et al. 2015; Leitão et al. 2016). Finally, it would be important to investigate response of specific traits separately, as some specific functions may be lost along changes in functional beta diversity.
Conclusions
We addressed the response of beta diversity along gradients of agricultural land cover and found that variation in fish composition among streams may respond in a nonlinear fashion to changes in the landscape. Additionally, the responses may depend on the scale at which environmental modification is quantified and on the diversity metric (taxonomic vs. functional). For instance, intermediate levels of agriculture at the catchment scale and deforestation of upstream riparian zones seem to increase taxonomic beta diversity, but higher levels may cause taxonomic homogenization in landscapes dominated by anthropogenic land cover (> 70% cropland cover). On the other hand, intermediate levels of catchment cropland and instream modification may lead to higher functional redundancy among streams, but extremely high levels of these modifications may increase functional dissimilarity among streams due to the occurrence of some tolerant but morphologically very distinct fish species. In conclusion, the changes in regional diversity patterns of stream fish communities observed in the current study were complex and partially counterintuitive. Anthropogenic modifications can introduce unexpected variability among stream communities, which means that low beta diversity may not always necessarily indicate a degraded environmental condition, and that high beta diversity may not always indicate a reference environmental condition.
References
Albouy, C., Guilhaumon, F., Villéger, S., Mouchet, M., Mercier, L., Culioli, J. M., Tomasini, J. A., Le Loc’h, F., & Mouillot, D. (2011). Predicting trophic guild and diet overlap from functional traits: Statistics, opportunities and limitations for marine ecology. Marine Ecology Progress Series, 436, 17–28.
Allan, J. D. (2004). Landscapes and riverscapes: The influence of land-use on stream ecosystems. Annual Review of Ecology, Evolution, and Systematics, 35, 257–284.
Allan, J. D., Wipfli, M. S., Caouette, J. P., Prussian, A., & Rodgers, J. (2003). Influence of streamside vegetation on inputs of terrestrial invertebrates to salmonid food webs. Canadian Journal of Fisheries and Aquatic Sciences, 60(3), 309–320.
Andrew, M. E., Wulder, M. A., Coops, N. C., & Baillargeon, G. (2012). Beta diversity gradients of butterflies along productivity axes. Global Ecology and Biogeography, 21, 352–364.
Bolker, B., 2017. bbmle: Tools for general maximum likelihood estimation. R package version 1.0.20.
Burrell, T. K., O'Brien, J. M., Graham, S. E., Simon, K. S., Harding, J. S., & McIntosh, A. R. (2014). Riparian shading mitigates stream eutrophication in agricultural catchments. Freshwater Science, 33(1), 73–84.
Casatti, L., Teresa, F. B., Zeni, J. O., Ribeiro, M. D., Brejão, G. L., & Ceneviva-Bastos, M. (2015). More of the same: High functional redundancy in stream fish assemblages from tropical agroecosystems. Environmental Management, 55(6), 1300–1314.
Dala-Corte, R. B., Giam, X., Olden, J. D., Becker, F. G., Guimarães, T. F., & Melo, A. S. (2016). Revealing the pathways by which agricultural land-use affects stream fish communities in South Brazilian grasslands. Freshwater Biology, 61(11), 1921–1934.
Dala-Corte, R. B., Becker, F. G., & Melo, A. S. (2017). Riparian integrity affects diet and intestinal length of a generalist fish species. Marine and Freshwater Research, 68(7), 1272–1281.
Fox, J., & Monette, G. (1992). Generalized collinearity diagnostics. Journal of the American Statistical Association, 87(417), 178–183.
Fox, J., & Weisberg, S. (2011). An {R} Companion to Applied Regression (2nd ed.). Thousand oaks CA: Sage URL: http://socserv.socsci.mcmaster.ca/jfox/Books/Companion.
Geo Catálogo, M. M. A. (2015). Geo Catálogo, Brazil. Ministério do Meio Ambiente (MMA).
Giam, X., Hadiaty, R. K., Tan, H. H., Parenti, L. R., Wowor, D., Sauri, S., Chong, K. Y., Yeo, D. C. J., & Wilcove, D. S. (2015). Mitigating the impact of oil-palm monoculture on freshwater fishes in Southeast Asia. Conservation Biology, 29(5), 1357–1367.
Heino, J., Melo, A. S., Siqueira, T., Soininen, J., Valanko, S., & Bini, L. M. (2015). Metacommunity organisation, spatial extent and dispersal in aquatic systems: Patterns, processes and prospects. Freshwater Biology, 60(5), 845–869.
Hermoso, V., Clavero, M., & Kennard, M. J. (2012). Determinants of fine-scale homogenization and differentiation of native freshwater fish faunas in a Mediterranean Basin: Implications for conservation. Diversity Distributions, 18(3), 236–247.
Hyatt, T. L., & Naiman, R. J. (2001). The residence time of large woody debris in the Queets River, Washington, USA. Ecological Applications, 11(1), 191–202.
Johnson, R. K., & Angeler, D. G. (2014). Effects of agricultural land use on stream assemblages: Taxon-specific responses of alpha and beta diversity. Ecological Indicators, 45, 386–393.
Jones, E. B., Helfman, G. S., Harper, J. O., & Bolstad, P. V. (1999). Effects of riparian forest removal on fish assemblages in southern Appalachian streams. Conservation Biology, 13(6), 1454–1465.
Kaufmann, P. R., Levine, P., Peck, D. V., Robison, E. G., & Seeliger, C. (1999). Quantifying physical habitat in wadeable streams. EPA/620/R-99/003. Washington, D.C: U.S. Environmental Protection Agency.
King, R. S., Baker, M. E., Whigham, D. F., Weller, D. E., Jordan, T. E., Kazyak, P. F., & Hurd, M. K. (2005). Spatial considerations for linking watershed land cover to ecological indicators in streams. Ecological Applications, 15(1), 137–153.
Laliberté, E., Legendre, P., & Shipley, B. (2014). FD: measuring functional diversity from multiple traits, and other tools for functional ecology. R package version 1.0–12.
Legendre, P., Borcard, D., & Peres-Neto, P. R. (2005). Analyzing beta diversity: Partitioning the spatial variation of community composition data. Ecological Monographs, 75(4), 435–450.
Leitão, R. P., Zuanon, J., Villéger, S., Williams, S. E., Baraloto, C., Fortunel, C., Mendonça, F. P., & Mouillot, D. (2016). Rare species contribute disproportionately to the functional structure of species assemblages. Proceedings of the Royal Society B, 283, 20160084.
Lorion, C. M., & Kennedy, B. P. (2009). Riparian forest buffers mitigate the effects of deforestation on fish assemblages in tropical headwater streams. Ecological Applications, 19(2), 468–479.
Lowrance, R., Todd, R., Fail, J., Hendrickson, O., Leonard, R., & Asmussen, L. (1984). Riparian forests as nutrient filters in agricultural watersheds. BioScience, 34(6), 374–377.
Lowrance, R., Altier, L. S., Newbold, J. D., Schnabel, R. R., Groffman, P. M., Denver, J. M., Correll, D. L., Gilliam, J. W., Robinson, J. L., Brinsfield, R. B., Staver, K. W., Lucas, W., & Todd, A. H. (1997). Water quality functions of riparian forest buffers in Chesapeake bay watersheds. Environmental Management, 21(5), 687–712.
Macedo, M. N., Coe, M. T., DeFries, R., Uriarte, M., Brando, P. M., Neill, C., & Walker, W. S. (2013). Land-use-driven stream warming in southeastern Amazonia. Philosophical transactions of the Royal Society of London. Series B, 368(1619), 20120153.
Majdi, N., Boiché, A., Traunspurger, W., & Lecerf, A. (2015). Community patterns and ecosystem processes in forested headwater streams along a gradient of riparian canopy openness. Fundamental and Applied Limnology, 187(1), 63–78.
Melo, A. S. (2018). CommEcol: Community ecology analyses: R package version 1.6.5.
Melo, A. S., Rangel, T. F., & Diniz-Filho, J. A. (2009). Environmental drivers of beta-diversity patterns in New-World birds and mammals. Ecography, 32(2), 226–236.
Minshall, G. W. (1978). Autotrophy in stream ecosystems. BioScience, 28(12), 767–771.
Nakano, S., Miyasaka, H., & Kuhara, N. (1999). Terrestrial-aquatic linkages: Riparian arthropod inputs alter trophic cascades in a stream food web. Ecology, 80(7), 2435–2441.
Nekola, J. C., & White, P. S. (1999). The distance decay of similarity in biogeography and ecology. Journal of Biogeography, 26(4), 867–878.
Niyogi, D. K., Koren, M., Arbuckle, C. J., & Townsend, C. R. (2007). Stream communities along a catchment land-use gradient: Subsidy-stress responses to pastoral development. Environmental Management, 39(2), 213–225.
Odum, E. P., Finn, J. T., & Franz, E. H. (1979). Perturbation theory and the subsidy-stress gradient. BioScience, 29(6), 349–352.
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al. (2017). vegan: Community Ecology Package. R package version 2.4–8.
Olden, J. D., & Poff, N. L. (2004). Ecological processes driving biotic homogenization: Testing a mechanistic model using fish faunas. Ecology, 85(7), 1867–1875.
Overbeck, G. E., Müller, S. C., Fidelis, A., Pfadenhauer, J., Pillar, V. P., Blanco, C. C., et al. (2007). Brazil's neglected biome: The south Brazilian Campos. Perspective in Plant Ecology, Evolution and Systematics, 9(2), 101–116.
Paula, F. R., Gerhard, P., Wenger, S. J., Ferreira, A., Vettorazzi, C. A., & Ferraz, S. F. (2013). Influence of forest cover on instream large wood in an agriculture landscape of southeastern Brazil: A multi-scale analysis. Landscape Ecology, 28(1), 13–27.
Pool, T. K., & Olden, J. D. (2012). Taxonomic and functional homogenization of an endemic desert fish fauna. Diversity and Distributions, 18(4), 366–376.
R Core Team. (2017). R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
Rabeni, C. F., & Smale, M. A. (1995). Effects of siltation on stream fishes and the potential mitigating role of the buffering riparian zone. Hydrobiologia, 303(1–3), 211–219.
Rahel, F. J. (2002). Homogenization of freshwater faunas. Annual Review of Ecology and Systematics, 33, 291–315.
Roth, N. E., Allan, J. D., & Erickson, D. L. (1996). Landscape influences on stream biotic integrity assessed at multiple spatial scales. Landscape Ecology, 11(3), 141–156.
Scott, M. C., & Helfman, G. S. (2001). Native invasions, homogenization, and the mismeasure of integrity of fish assemblages. Fisheries, 26(11), 6–15.
Strahler, A. N. (1952). Hypsometric (area-altitude) analysis of erosional topography. Geological Society of America Bulletin, 63(11), 1117–1142.
Sweeney, B. W., Bott, T. L., Jackson, J. K., Kaplan, L. A., Newbold, J. D., Standley, L. J., Hession, W. C., & Horwitz, R. J. (2004). Riparian deforestation, stream narrowing, and loss of stream ecosystem services. Proceedings of the National Academy of Sciences, 101(39), 14132–14137.
Teixeira-de Mello, F., Meerhoff, M., González-Bergonzoni, I., Kristensen, E. A., Baattrup-Pedersen, A., & Jeppesen, E. (2015). Influence of riparian forests on fish assemblages in temperate lowland streams. Environmental Biology of Fishes, 99(1), 133–144.
Tuomisto, H., & Ruokolainen, K. (2006). Analyzing or explaining beta diversity? Understanding the targets of different methods of analysis. Ecology, 87(11), 2697–2708.
Walser, C. A., & Bart, H. L. (1999). Influence of agriculture on in-stream habitat and fish community structure in Piedmont watersheds of the Chattahoochee River system. Ecology of Freshwater Fish, 8(4), 237–246.
Walters, D. M., Leigh, D. S., & Bearden, A. B. (2003). Urbanization, sedimentation, and the homogenization of fish assemblages in the Etowah River Basin, USA. Hydrobiologia, 494(1–3), 5–10.
Wright, J. P., & Flecker, A. S. (2004). Deforesting the riverscape: The effects of wood on fish diversity in a Venezuelan piedmont stream. Biological Conservation, 120(3), 439–447.
Zeni, J. O., & Casatti, L. (2014). The influence of habitat homogenization on the trophic structure of fish fauna in tropical streams. Hydrobiologia, 726(1), 259–270.
Acknowledgements
We are grateful to C Hartmann, M Camana, M Dalmolin, L de Fries, BA Meneses, LR Podgaiski, V Bastazini, V Lampert, M Santos, KO Bonato, and RA Silveira for their fieldwork assistance. We also thank J Ferrer, T Carvalho, and LR Malabarba (Laboratório de Ictiologia of the UFRGS) for assistance with species identification. This research was funded by CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico; 457503/2012-2). This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001 (doctorate scholarships to RB Dala-Corte and LF Sgarbi). AS Melo received a research fellowship from CNPq (proc. 309412/2014-5).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Dala-Corte, R.B., Sgarbi, L.F., Becker, F.G. et al. Beta diversity of stream fish communities along anthropogenic environmental gradients at multiple spatial scales. Environ Monit Assess 191, 288 (2019). https://doi.org/10.1007/s10661-019-7448-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10661-019-7448-6