
Abstract
Bitter rot (BR) and glomerella leaf spot (GLS) occur in all the Brazilian South orchards with different species frequency per region. The main species found in Brazil belong to the Colletotrichum acutatum complex (Cac) and to the C. gloeosporioides complex (Cgc). The objectives of this study were to evaluate the favorable conditions for the germination and growth of five Colletotrichum species and, to compare pathogenicity and aggressiveness among species and between two apple cultivars in fruit, causing BR, and in leaves, causing GLS. The highest germination of conidia (above 90%) was achieved between 15 and 30 °C for Cgc species while for Cac species the highest germination (mean of 71%) was at 25 °C. The optimal temperature for mycelial growth did not differ between complexes and ranged from 23 to 27 °C. Althought, Cgc had a higher estimated maximum temperature then Cac. C. siamense cause BR symptoms regardless of cultivar or type of inoculation. C. nymphaeae and C. melonis caused symptoms in wounded and unwounded fruit but for C. paranaense and C. fructicola the symptoms were observed only in wounded fruit. Leaf symptoms of GLS was more severe in stressed leaves inoculated on the abaxial side compared to non-stressed leaves inoculated on the adaxial side. C. fructicola and C. nymphaeae were able to cause GLS symptoms in leaves with severity varying from 17.5 to 25% (cv. Gala). C. fructicola was the most severe causing GLS symptoms in cv. Gala (severity around 20.5%) and, in cv. Eva (severity around 11%). We discuss the roles that each species plays as a pathogen to better understand BR and GLS in different orchards conditions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Colletotrichum is one of the most critical plant pathogens that cause anthracnose in fruit and flowers of a wide range of hosts, mainly in tropical and subtropical regions (Sutton et al. 1992; Cannon et al. 2012). On apples (Malus domestica Borkh.), species of Colletotrichum can cause bitter rot (BR) and glomerella leaf spot (GLS) (Gonzalez et al. 2006; Moreira et al. 2019).
BR is a disease that occurs in fruit and is present in most countries where apples are grown and may result in losses of up to 50% in the pre- and post-harvest periods (Sutton et al. 1992). Typical symptoms in fruit include light brown lesions that grow in diameter over time and become dark brown and depressed (Taylor 1971). All apple cultivars are susceptible to BR, particularly those belonging to the late harvest group such as Granny Smith, Pink Lady, and Fuji (Denardi et al. 2003).
GLS is a disease that causes symptoms mainly on leaves but also can cause small lesions on fruit. It was first reported in 1970 in the USA (Taylor 1971). In Brazil, it was reported in 1983, and its presence has progressively increased and occurs being one of the most severe disease in all commercial orchards in South Brazil (Sutton and Sanhueza 1998; Moreira et al. 2019), causing severe damage to the Gala cultivar and its clones, which correspond to 60% of the production area (Petri et al. 2011). Symptoms of GLS appear as purple-red spots 2 days after infection and then coalesce and grow into irregular necrosis (Leite et al. 1988; Moreira et al. 2019). In fruits, it can cause small brown lesions (1–3 mm) with a slight depression that does not increase in size over time (Sutton and Sanhueza 1998). Golden delicious descendant cultivars, such as the Gala cultivar, are highly susceptible to GLS (Sutton and Sanhueza 1998; Gonzalez and Sutton 1999). Additionally, symptoms on cultivar Eva, a resistant cultivar, were observed and the pathogen was isolated and confirmed as pathogenic (Moreira et al. 2019). This cultivar was reported to be resistant to the pathogen (Furlan et al. 2010), a fact that has been worrying producers from warmer regions of Brazil as São Paulo and Paraná State, where this cultivar is grown.
Through morphological and molecular methodologies, the Colletotrichum genus was reclassified (Damm et al. 2012; Weir et al. 2012) and separated into groups: C. acutatum complex (Cac), C. gloeosporioides complex (Cgc), and C. boninense complex (Cbc). Therefore, new species have been reported to cause BR; C. fioriniae, C. clavatum, C. godetiae, C. nymphaeae, C. melonis, and C. paranaense (Bragança et al. 2016; Munir et al. 2016; Velho et al. 2015; Kou et al. 2014; Baroncelli et al. 2014) belonging to Cac; and C. theobromicola, C. siamense, C. fragariae, and C. fructicola (Alaniz et al. 2012; Velho et al. 2015) belonging to Cgc. Similarly, new species have been reported as causal agents of GLS, such as C. nymphaeae, C. paranaense, and C. melonis, belonging to Cac; C. fructicola, C. siamense, and C. theobromicola, belonging to Cgc; and C. karstii, belonging to Cbc (Bragança et al. 2016; Velho et al. 2015; Moreira et al. 2019). Species of Colletotrichum from apple show small intraspecific variation and isolates belonging to the same species have similar incubation period, latent period and type of GLS symptoms (Moreira et al. 2019).
Frequent incidence of BR and GLS is observed in subtropical regions where rain and warm weather favour conidia spread and development of the disease. Although both diseases often occur simultaneously in orchards, their relationship with causal agents is not fully understood. The importance of infection from flowers as a source of primary inoculum was recently reported (Hamada et al. 2020). In this work the authors prove that Colletotrichum isolated from flower can cause symptoms of GLS on leaves and fruit and also can cause BR.
Several factors can affect the pathogen-host relationship, such as temperature, host-related characteristics, and the environment in which the host is grown. Temperature differences have significant implications in the management of the disease based on chemical control, in genetic resistance of cultivars, and in harvest strategies (Phoulivong et al. 2012). This information has a large impact when you consider that apple trees are grown in several regions of Brazil, in which the planted crops, rainfall, and climate are different. Infection on the upper or lower surface of the tissues, as well as age and maturation of the plants tissue can influence the appearance of symptoms (Santos et al. 1999). Another fact is the environment, which can leave the plants more predisposed to the pathogen attack depending on the level of stress it provides (Bedendo et al. 2018). Plant growth and pathogen survival are influenced by several factors, including abiotic and biotic stresses. Plants respond to these factors by inducing their defence mechanisms, which include the expression of several effectors, receptors, signalling, and protective molecules (Jain and Kumar 2015).
The identification of new species of Colletotrichum causing both BR and GLS symptoms, and the differences in frequency of Colletotrichum species within the apple regions of Brazil (Moreira et al. 2019) suggest that temperature and infections process can vary and play an important factor to pathogenicity and aggressivity of the two diseases. Thus, the objectives of this study were: (i) to compare the effect of temperature on germination and on mycelium growth of different species of Colletotrichum, (ii) to quantify the incubation period, the latent period and the severity of these species on fruit and leaves of two cultivars with different levels of resistance (‘Eva’ and ‘Gala’), and (iii) to investigate the differences in the infection and colonization processes of Colletotrichum species on leaves obtained from non-stressed and stressed plants.
Material and methods
The selected isolates were obtained from symptomatic apple leaves collected in different producing regions in Brazil (Table 1). Five species that belongs to different complexes following the new classification of the genus Colletotrichum were chosen (Damm et al. 2012; Weir et al. 2012). They are: C. nymphaeae, C. paranaense and C. melonis, belonging to Cac and; C. fructicola and C. siamense, belonging to Cgc (Bragança et al. 2016). One isolate of each species of Colletotrichum was chosen due to the small intraspecific variation reported by Moreira et al. 2019. During the tests the isolates were kept in Potato-Dextrose-Agar (PDA) media (39 g L−1, Himedia®, Curitiba, Brazil) and were later stored on filter paper.
Effect of temperature on the conidia germination of Colletotrichum species in vitro
Spore suspensions from single-spore isolates were prepared and adjusted to 104 conidia ml−1. An aliquot of 40 μL of each suspension was transferred and spread into a fresh Petri dishes containing agar-agar culture medium (16 g L−1 agar-agar, 1000 mL distilled water). The plates were incubated at temperatures of 10, 15, 20, 25 and 30 ± 1 °C and with 12 h light regime. After 24 h of incubation, the process of germination was stopped by adding 20 μL of lactoglycerol over the suspension. The experimental design was completely randomized with three replicates per treatment. For the evaluation, the first 50 conidia found on each plate were counted with the aid of a light microscope at 40x magnification. The conidia were considered germinated when presenting a germ tube size equal to or greater than its length.
Germination and appressorium formation in leaf and fruit surface of cultivar Gala
Detached leaves (technique adapted from Liu et al. 2007) and apple fruits of Gala cultivar were inoculated with conidial suspensions adjusted to 104 conidia mL−1 of each Colletotrichum species. An aliquot of 40 μL of each suspension was deposited on a pre-defined area on the surface of wounded (w) and unwounded (nw) fruits, in the adaxial and abaxial sides of detached leaves. Leaves and fruits treated with sterile distilled water were used as control. The experimental design was completely randomized with three replicates per treatment, where each experimental plot consisted in a moisturized chamber containing a leaf/fruit. The experiment was maintained at 24 °C in two wetting periods (6 and 24 h). The inoculation was done with five Colletotrichum species on both sides of the leaf and with and without wounding on fruit. The essays were done with three repetitions and the samples were evaluated in two wetting periods, in total 72 leaf samples and 72 fruit samples.
Before cutting the samples, they were fixed by depositing an aliquot of 50 μL of Karnovsky solution (2.5% glutaraldehyde and 2.5% of formaldehyde solution in sodium-cacodilate buffer 0.05 M – pH 7.2 and CaCl2 0.001 M) (Karnovsky 1965) over inoculated area. One sample (approximately 1 cm2) of each repetition were excised from the inoculated regions of the leaves/fruits and post-fixed in 2% (w/v) osmium tetroxide (OsO4) vapour for 12 h (Kim 2008; Baggio et al. 2015), mounted on aluminum stubs using double-sided carbon tape and transferred into a container with silica gel present to dry for 48 h. The fragments were sputter coated (SCD 050 Bal Tec) and examined with a LEO 435 VP scanning electron microscope (Leo Elektronenmikroskopie GmbH) at an accelerating voltage of 20 kV and 12.0 mm working distance.
Effect of temperature on the mycelial growth of Colletotrichum species in vitro
For each isolate, mycelial plugs of 5 mm diameter were removed from the edge of actively growing, 8-day-old colonies and transferred to the center of Petri dishes containing PDA (39 g L−1, Himedia®, Curitiba, Brazil). The isolates were incubated at temperatures of 5, 10, 15, 20, 25 and 30 ± 1 °C, and with 12 h light regime. The experimental design was completely randomized with four replicates per treatment. The evaluation of the experiment was performed daily for 7 days by measuring perpendicular diameters of the colonies.
Pathogenicity of Colletotrichum species on apple fruits of cultivars Gala and Eva
Fruit surface was disinfested by immersion in 70% alcohol for 1 min, and then in 0.5% sodium hypochlorite for 2 min, and thereafter washed with sterile distilled water and dried at room temperature for 4 h.
Fruits were placed individually in disinfested plastic containers containing moistened filter paper with sterilized distilled water. Inoculation was performed with and without wound through two methodologies: (i) 40 μL of conidial suspension at a concentration of 104 conidia mL−1 in the equatorial region of the fruit, and (ii) with mycelial plugs (5 mm) obtained from the edges of 8-day-old colonies. Fruit inoculated only with distilled water, following the method described above, were used as control.
After inoculation, the fruits were incubated in growth chambers (Conviron®) at 25 °C and in a photoperiod of 12 h light. The incubation and latent periods, the incidence of fruit with symptoms and the diameter of each lesion were evaluated daily until the first lesion reached the diameter of the fruit. The experimental design was completely randomized with six replicates per treatment, each experimental unit represented by a fruit.
Pathogenicity of Colletotrichum species on detached apple leaves of cultivars Gala obtained from non-stressed and from stressed plants
The evaluation of the GLS development was first carried out in the Gala cultivar, comparing the aggressiveness of the symptoms on leaves inoculated on the abaxial and adaxial surfaces and also comparing the aggressiveness of the symptoms on leaves of non-stressed plants and of stressed plants. The leaf samples of non - stressed plants were obtained from an experimental orchard with pest and weed control and with nutritional management. The sampled leaves of stressed plants were from potted plants growing in a greenhouse with a high level of woolly apple aphid infestation.
For inoculation of the detached leaves, the conidial suspensions of each of the Colletotrichum species were prepared in water with tween (20%) and adjusted to 104 conidia mL−1. The control treatment was represented by mixing water with tween (20%) without pathogen structures. An aliquot of 100 μL of each suspension was deposited on a pre-defined area on the surface of each detached leaf (technique adapted from Liu et al. 2007). Each leaf was packed in a Gerbox, containing cotton and filter paper moistened, and incubated at 25 °C for 7 days. In each experiment a completely randomized design was used, with treatments in the factorial scheme, with six treatments (five Colletotrichum species and control) and two types of inoculations (abaxial and adaxial), with four replicates, in which each Gerbox was an experimental unit containing one leaf.
The development of the disease was evaluated daily for 7 days under a stereoscopic microscope and the incubation period was determined for each species and for each environment from which the leaves were obtained. The incubation period was determined when the symptom under a stereoscopic microscope was observed. This parameter was only validated when at least 50% of the repetitions presented a symptom.
Uninoculated stressed and non-stressed leaves were prepared for anatomical observation under light microscopy, totaling 10 samples per leaf type (stressed or non-stressed). For the preparation of the material, first a smaller sample of each leaf was cut out, with the aid of cutting blades. These regions were sectioned in the middle, and the sections were fixed in Karnowski’s solution (Karnowski 1965) and kept refrigerated in Eppendorfs. After this step, the cuts were dehydrated in ethyl alcohol of 30, 50, 70, 90 and 3x in 100% with duration of 30 min in each solution. After the last wash the samples were infiltrated in a solution of 100% ethyl alcohol (1:1), leaving to rest for 24 h in the refrigerator. After this period, the samples were transferred to pure historesin solution (Leica® historesin Embeeding Kit), which remained in the refrigerator for another 24 h. The embedding was then performed in a solution of resin + activator powder + hardener. Aliquots of 400 μL of this solution were deposited in smaller blocks in a silicone form. Each cut was inserted into a block while the solution hardened. After hardening, the material was uninformed and glued on wooden cubes. Slices of 7 μm were made on a rotating microtome and the samples were stained with toluidine blue pH 4. Each sample (10 samples per leaf type) was then observed and photographed under a light microscope to compare the anatomic differences between stressed and non-stressed leaves. The methodology for microscopy sample preparation was adapted from O’Brien and Mc Cully (1981).
Incubation period and severity of GLS caused by Colletotrichum species on apple leaves of cultivars Gala and Eva obtained from the commercial orchard
The evaluation of the development of GLS symptoms was also performed in detached leaves of Gala and Eva cultivars obtained from commercial orchard plants 20 days after sprouting, washed in sterile distilled water and kept in Petri dishes containing agar-agar medium. The leaves were inoculated on the adaxial side with the different Colletotrichum species by conidia spray at a concentration of 104 conidia mL−1 with the aid of a manual atomizer. After inoculation the plates containing the leaves were incubated in BOD chambers at 24 °C and a photoperiod of 12 h light. The experimental design was completely randomized with five replicates per treatment; each experimental unit was represented by a plate containing a leaf. The incubation period was evaluated by daily leaf observation until symptom onset and the severity (%) of GLS was evaluated 20 days after inoculation using the scale described by Kowata et al. (2010).
Data analysis
The conidial germination data at different temperatures were used to calculate the percentage of germination of each isolate and mean germination percentage were compared at each tested temperature. Mycelial growth data were used to compare the isolates after 7 days of incubation. The averages of mycelial growth were compared at 25 °C. For the analysis of pathogenicity on fruits, were used the diameter data 2–3 weeks after inoculation for fruits with and without wound, respectively. The means were compared between inoculated isolates by the 2 methodologies in fruits of Gala and Eva cultivars.
The homogeneity of variance was verified by Bartlett test, then the data were submitted to analysis of variance (ANOVA), verifying the effect of treatments by F test for all tests. When the test was significant the means were compared by Scott-Knott test at 5% of probability. There were no transformation data for statistical analysis in any of these experiments.
For mycelial growth data were also determined minimum, optimum and maximum temperatures considering the area under the curve of mycelial growth (AUCMG) of each isolate. For this, the data was adjusted to generalized beta function by Bassanezi et al. (1998):
where T is the temperature, Y is the component (AUCMG), Tmin is the minimum temperature, Topt is the optimum temperature, Tmax is the maximum temperature and B3 is a parameter which influences the temperature around the optimum. The means of Tmim, Topt and Tmax were compared between Colletotrichum complexes by t test 5% of probability.
All tests were performed twice. As no significant difference was observed between the trials the statistical analysis was performed with the mean of the treatments in the two trials of each experiment. All analyzes were performed using the software R (R Core Team).
Results
Effect of temperature on the germination of conidia of Colletotrichum species in vitro
The conidial germination of the Cgc was significantly higher than the conidial germination of the Cac at all temperatures tested, except at 25 °C. At 10 °C all isolates still have germination with the exception of C. melonis. At 30 °C all isolates were germinated, and the germination percentage was higher than 90% for C. melonis, C. fructicola and C. siamense. The specie C. paranaense got lower germination at all temperatures and did not exceed 37% (Table 2).
Effect of temperature on the mycelial growth of Colletotrichum species in vitro
The minimum temperature for the development of colonies in PDA medium of the species of the Cac was significantly lower than the minimum temperature for the development of colonies of the Cgc. The maximum temperature for the development of colonies on PDA medium to the species belonging to Cac was also significantly lower than the maximum temperature for the development of Cgc colonies. The optimum temperature for the development of species belonging to the Cac was between 22 and 24 °C and between 25 and 26 °C for species belonging to the Cgc. The average optimum temperature was 23.8 °C for species belonging to the Cac and 25.6 °C for species belonging to the Cgc. The best temperature range for the development of colonies belonging to the Cac was 23.3 to 24.3 °C and to the Cgc was 24.5 to 26.8 °C (Table 3).
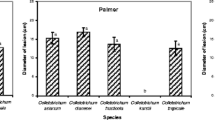
The species of the Cgc developed significantly larger colonies on PDA medium after 7 days at 25 °C than the species of the Cac. The maximum average diameter was 5.34 cm for species belonging to the Cgc and 3.51 cm for species belonging to Cac (Fig. 1).

Colonies of Colletotrichum species (C. nymphaeae, C. paranaense and C. melonis belonging to C. acutatum species complex and C. siamense and C. fructicola belonging to C. gloeosporioides species complex) growing in PDA medium after 7 days at 25 °C. The value above of the colonies is the mycelial growth in centimeters. Original data without processing; means followed by different letters differ significantly by the Scott-Knott test at 5% probability
Germination and appressorium formation in leaf and fruit surface of cultivar Gala
The germination of conidia 6 h after inoculation (data not shown) and the formation of appressoria 24 h after inoculation (Fig. 2a-e) were observed on abaxial and on adaxial sides of the leaves. In wounded fruits, 6 h after inoculation, C. nymphaeae and C. paranaense had germinated conidia (data not shown), and 24 h after inoculation all five species showed apressorium formation (Fig. 2f-j). In unwounded fruits it was not observed conidial germination of any of the five species 24 h after inoculation (data not shown).

Scanning electron microscopy of apple (Malus domestica) leaves of cultivar Gala inoculated with Colletotrichum nymphaeae (a), C. paranaense (b), C. melonis (c), C. fructicola (d) and C. siamense (e) in the adaxial side after 24 h and, scanning electron microscopy of wounded ‘Gala’ fruits inoculated with C. nymphaeae (f), C. paranaense (g), C. melonis (h), C. fructicola (i) and C. siamense (j) after 24 h. es = epicuticular striae; c = cuticle; s = spore; ns = non-germinated spore; ap = apressorium. Bars scale 10 μm (c, d), 20 μm (a, e, f, g, h, i, j), 30 μm (b)
Pathogenicity of Colletotrichum species on apple fruits of cultivars Gala and Eva
All species of Colletotrichum inoculated with spore suspension or mycelial plug in apple fruits of cultivars Gala and Eva caused BR symptom in wounded fruits. In general, in wounded fruit the BR lesions were larger in fruits inoculated with C. nymphaeae. The mean size of the lesions was significantly different between species when the fruits of Gala and Eva cultivars were inoculated with mycelial plug and when the ‘Gala’ fruits were inoculated with conidial suspension. However, on Eva there was no significant difference between the species inoculated with conidia. In unwounded fruits of cultivars Gala and Eva the average lesion sizes of BR after three weeks showed no significant difference between the species when inoculated with mycelial plug and suspension of conidia, respectively. Despite the large difference in mean values, the variation in lesion size between repetitions of the same treatment was very large in unwounded fruits and, consequently, the mean comparison test could not detect the difference between treatments. The averages were significantly different when the unwounded fruits of the cultivars Gala and Eva were inoculated with a suspension of conidia and mycelial plug, respectively. The highest average lesion was found in fruits inoculated with C. siamense (Table 4).
The incubation period (IP) of BR on wounded fruits of the cultivars Gala and Eva inoculated with conidia suspension or mycelial plug ranged from 2 to 6 days. On unwounded Gala fruits, the IP ranged from 15 to 25 days. On unwounded Eva fruits, the IP ranged from 5 to 17 days. Some species did not cause BR symptoms or symptoms occurred on less than 50% of the fruits (Table 4).
In wounded fruits, only lesions caused by C. nymphaeae and C. paranaense sporulated in fruits of cultivars Gala and Eva, and the latent period (LP) ranged from 6 to 21 days. In unwounded fruits the lesions caused by C. nymphaeae sporulated in cultivar Eva and the latent period was 20 days, and the BR lesions caused by C. paranaense sporulated in Gala and the latent period was 33 days. The other species did not sporulate over the lesions or sporulation was on less than 50% of the fruits (Table 4).
Pathogenicity of Colletotrichum species on apple leaves of cultivars Gala obtained from non-stressed and from stressed plants
Light microscopy images of the cuts made on stressed and non-stressed leaves showed differences in the anatomical structure. In the case of non-stressed leaves, the palisade parenchyma (Pp) tissue contained three strata with a tiny intercellular space, while the Pp of the stressed leaves contained only two strata (Fig. 3).

Cross sections of non-stressed leaves (a) and stressed leaves (b) without inoculation. ep: upper epidermis. pp.: palisade parenchyma. Bars scale 50 μm (a, b)
In the daily evaluation of the development of GLS symptoms the C. nymphaeae species showed no symptoms when inoculated on the adaxial side of both stressed and non-stressed leaves. In the abaxial side of non-stressed leaves, small lesions were observed under a stereoscopic microscope, but were not visible to the naked eye. On the abaxial side of stressed leaves the symptoms were visible to the naked eye (Table 5; Fig. 4).

Detached leaves of cultivar Gala (Malus domestica) inoculated with Colletotrichum nymphaeae (Cn), C. paranaense (Cp), C. melonis (Cm), C. fructicola (Cf) and C. siamense (Cs) and observed through stereoscopic microscope after 7 days of the inoculation. Bars scale 500, 200, 1000, 1000 μm (Cn), 500, 200, 1000, 500 μm (Cp), 500, 200, 200, 1000 μm (Cm), 1000, 2500, 1000, 2500 μm (Cf), 500, 200, 200, 1000 μm (Cs)
The C. paranaense species caused small lesions visible only in a stereoscopic microscope on the adaxial side of both stressed leaves and non-stressed leaves and on the abaxial side of non-stressed leaves. In the abaxial side of stressed leaves there was a great manifestation of visible symptoms to the naked eye (Table 5; Fig. 4). The C. melonis species when inoculated on the adaxial side of non-stressed leaves did not present any manifestations of GLS symptoms. In the adaxial side of stressed leaves and in the abaxial side of both stressed leaves and non-stressed leaves, visible lesions were observed only in stereoscopic microscope (Table 5; Fig. 4).The C. fructicola species showed symptoms visible to the naked eye in all situations evaluated, both on the adaxial and abaxial sides of the leaves, and also on both non-stressed leaves and stressed leaves (Table 5; Fig. 4).The C. siamense species when inoculated on the adaxial side of non-stressed leaves did not cause symptoms and on the stressed leaves few lesions were observed visible only in a stereoscopic microscope. In the abaxial side, visible lesions were observed only in stereoscopic microscopes in both non-stressed and stressed leaves (Table 5; Fig. 4). The incubation period ranged from two to three days in non-stressed leaves inoculated on the adaxial side, from three to six days in stressed leaves inoculated on the adaxial side, from two to four days in non-stressed leaves inoculated on the abaxial side and was two days for all Colletotrichum species inoculated in stressed leaves on the abaxial side (Table 5).
Incubation period and severity of GLS caused by Colletotrichum species on apple leaves of cultivars Gala and Eva obtained from the commercial orchard
The only species of Colletotrichum that caused symptoms of GLS on ‘Gala’ leaves from commercial orchard were C. fructicola and C. nymphaeae, and the only specie of Colletotrichum that caused symptoms of GLS on Eva leaves from commercial orchard was C. fructicola (Fig. 5). In ‘Gala’ leaves inoculated with C. fructicola the average severity of the disease was 17.5% in experiment 1 and 20.5% in experiment 2 and the incubation period was two days in both experiments. In Gala leaves inoculated with C. nymphaeae the average severity of the disease was 22.0% in experiment 1 and 25.0% in experiment 2 and the incubation period was three days in both experiments. In ‘Eva’ leaves inoculated with C. fructicola the average severity of the disease was 10.5% in experiment 1 and 11.0% in experiment 2, and the incubation period was three days in both experiments (Table S1). During the assessment period of experiments (20 days) and conditions used pathogen sporulation on the surface of the lesions was not observed.

Glomerella Leaf Spot symptoms 20 days after inoculation with Colletotrichum fructicola in detached leaves of cultivar Gala (a) and cultivar Eva (b). Glomerella Leaf Spot symptoms 20 days after inoculation with C. nymphaeae in detached leaves of cultivar Gala (c)
Discussion
C. nymphaeae, C. paranaense, and C. melonis species from apple had optimal germination of the conidia at temperatures between 20 and 25 °C, while C. fructicola and C. siamense had wider intervals (15–30 °C). This difference can be related to the prevalence of each species in the apple production regions of Brazil (Moreira et al. 2019).
On the leaves, although all species have formed appressorium on the abaxial and adaxial sides of the leaves; in some inoculations, there are no symptoms of GLS. C. fructicola and C. siamense caused symptoms observable by the naked eye in detached leaves of the Gala cultivar, and C. fructicola also caused symptoms observable by the naked eye in leaves of the Eva cultivar, which was known to be resistant to the pathogen (Furlan et al. 2010). Most Uruguayan isolates of C. fructicola from apple did not cause typical GLS symptoms on apple leaves; a few of them could infect plants and provoke spots resembling those seen in the initial stages of disease and with a longer incubation period, but seldom resulted in leaf tissue necrotization (Velho et al. 2015). This observation suggests that the Uruguayan leaf-infecting isolates of C. fructicola and some Colletotrichum species studied in the present work could be adapted. McDonald and Linde (2002) stated that some Colletotrichum isolates can be in the transitional evolutionary phase because they are not well adapted to leaf tissue colonization.
Colletotrichum species exhibit different growth rates depending on the incubation temperature. C. fructicola and C. siamense developed significantly larger colonies at 25 °C on PDA medium than C. nymphaeae, C. paranaense, and C. melonis. In tests conducted by Velho et al. (2015), C. fructicola (Cgc) isolates grew faster at 25 °C compared with other species. Moreover, in general, colonies of C. melonis and C. nymphaeae that belong to the Cac grew less rapidly than C. fructicola and C. theobromicola that belong to the Cgc. In fact, the growth rates of Cac isolates from different fruits, including apples, are generally slower at all temperatures than Cgc isolates (Wharton and Dieguez-Uribeondo 2004).
The optimum temperatures for the development of colonies in the PDA medium of C. nymphaeae, C. paranaense, and C. melonis were between 22 and 24 °C, and between 25 and 26 °C for C. fructicola and C. siamense. The average optimum temperature was 23.8 °C for C. nymphaeae, C. paranaense and C. melonis and 25.6 °C for C. fructicola and C. siamense. A similar result was obtained by Grahovac et al. (2012), who reported that optimal growth temperatures for the isolates belonging to the Cac and Cgc were between 23 and 28 °C. Even so, differences in growth rate and optimum growth temperature are not always a safe criterion for identifying the species (Freeman et al. 1998).
The five species of Colletotrichum formed appressorium on the surface of wounded fruits 24 h after inoculation, and all species caused BR symptoms on the fruit of Gala and Eva cultivars when inoculated with conidia suspension or mycelial plug. Despite a much longer period of incubation, most species also cause symptoms of BR on unwounded fruits. Velho et al. (2015) inoculated the fruits of cultivar Gala’ with five species of Colletotrichum, including C. melonis, C. fructicola, C. nymphaeae, C. karstii, and C. theobromicola, and all were pathogenic to wounded fruits, producing typical symptoms of BR. Wounds are known to increase the success of Colletotrichum infection and disease severity (Denardi et al. 2003; Phoulivong et al. 2012). In addition, wounds can increase the infectivity of C. fructicola in fruit of pear (Jingjing et al. 2014), and peach (Moreira et al. 2020). However, some fungi produce a variety of enzymes that degrade the cuticle and the cell wall and can infect plants without wounds (Liao et al. 2012). Isolates that are unable to infect unwounded fruits are often deficient in these enzymes. Velho et al. (2018) reported that C. fructicola BR-isolate showed significantly higher extracellular enzyme activity when compared to C. fructicola GLS-isolate. However, during C. fructicola leaf infection, no significant differences between isolates were detected for all enzymes tested. Therefore, the study of the enzymatic activity in the infection (ex vivo and in vivo) by different Colletotrichum species on apple leaves and fruit (wounded and unwounded) is a challenge for future research.
According to Velho et al. (2015), symptoms appear after 3–4 days in wounded fruit and after five-six days in unwounded fruits. Similar results were found in this study, where the BR incubation period in cultivars Gala and Eva ranged from two to six days in fruits inoculated with wounds. However, in unwounded fruits, the incubation period was much higher, being of 15 to 25 days in ‘Gala’ fruits and five to 17 days in ‘Eva’ fruit.
In the present study, only C. nymphaeae and C. paranaense (both Cac) sporulated on BR lesions, with latent period ranging from six to 21 days in wounded fruits and from 20 to 33 days in unwounded fruits. Munir et al. (2016) also observed more spores in fruits inoculated with Cac than in fruits inoculated with Cgc. In addition, Velho et al. (2015) observed sporulation in fruits inoculated with C. nymphaeae, C. melonis (both Cac), and C. fructicola (Cgc). Cgc seems to have a disadvantage in sporulation compared to Cac, and this could be a concern for disease chemical control during harvest and postharvest.
The incubation period of GLS caused by C. fructicola was two days for both cultivars, and a similar result was observed by Velho et al. (2015), who reported symptoms in apple leaves of cultivar Gala two-three days after inoculation. These authors also reported symptoms of GLS only when inoculated with C. fructicola, which differs from the data obtained in the present study, where Gala leaves showed severe GLS symptoms when inoculated with C. nymphaeae (IP = three days). According to Lakshmi et al. (2011), Colletotrichum species were more pathogenic in fruits or leaves due to the types of enzymes produced. According to Gonzalez et al. (2006), only a limited number of genotypes can cause GLS. This may indicate that different parts of the plant can act selectively on the specialization of organs as regards resistance to pathogens (Doyle et al. 2013). Expanding the capacity of dual infection may provide an opportunity for the pathogen to survive during adverse periods in orchards (Lakshmi et al. 2011).
Therefore, according to the results obtained in this work, for the management of the BR and the GLS in a specific orchard, it is important to know the frequency of each species within each species complex, since different species have different abilities to develop symptoms of BR and GLS. Similar results were obtained by Munir et al. (2016), who reported the importance of accurate identification of Colletotrichum species within each species complex, since they exhibit differences in pathogenicity. In addition, Moreira et al. (2019) observed a difference in Colletotrichum species occurring in apple orchards in southern Brazil. In Paraná State, the predominant species is C. nymphaeae and in Santa Catarina and Rio Grande do Sul States, the predominant species is C. fructicola. This difference can be attributed to the fungicide management, the microclimatic differences of each region, and the plant stress level.
Stressed and non-stressed leaves, when inoculated on the adaxial side with the species C. nymphaeae, C. paranaense, C. melonis, and C. siamense showed no symptoms of GLS visible to the naked eye, but only few lesions visible under a stereoscopic microscope. In these cases, the incubation period ranged from two to six days, contrasting with the regularity of the disease. Normally, under conditions of temperature above 20 °C and leaf wetness of 10 h, as in the study conditions, the incubation period for the disease is 48 h (Moreira et al. 2019). In inoculations with Cgc isolates collected before 2010, the GLS severity was directly related to the wetness period in the temperature range between 12 °C and 26 °C and, with progressive increase of lesions ranging from tiny necrotic points to total leaf necrosis (Katsurayama and Boneti 2010).
The presence of symptoms, even if only visible in a stereoscopic microscope for these species, shows that the pathogen infected the host, but its development was interrupted, presenting a quiescent infection and that, even in stressed leaves, did not have the capacity to overcome the defence reactions of the plant. For the genus Colletotrichum, it is common to develop quiescent infections (Peres et al. 2005). In addition, the PR-10 protein class has been one of the most studied proteins in apple in response to pathogen infection (Ziadi et al. 2001; Liu and Ekramoddoullah 2006; Jain and Kumar 2015). PR-10 expression was increased by infection with C. fructicola in susceptible apple plants (Rockenbach et al. 2015). Thus, research aimed at identifying the gene(s) involved in blocking Colletotrichum species infection in apple plants is of prime importance.
In the abaxial side of non-stressed leaves, for the same species discussed previously, the manifestation of symptoms of GLS visible via a stereoscopic microscope was greater than that on the adaxial side, presenting an incubation period of two to four days. In stressed leaves inoculated on the abaxial side, there were intense symptoms visible to the naked eye, where only C. melonis and C. siamense species caused symptoms visible only via a stereoscopic microscope. The incubation period for this treatment was 2 days, indicating a more rapid penetration of the host by the pathogen.
In young apple leaves it has been reported that splashed rain droplets carrying conidia of Colletotrichum spp. tend to deposit on the anticlinal cell walls due to the highly hydrophobic and wavy nature of their surfaces (Sutton and Shane 1983; Bringe et al. 2006). In addition, it has been demonstrated that epidermal cells of young apple leaves exhibit a distinctive curvature of the periclinal cell walls that results in an undulated surface of the cuticle. However, during ontogenesis, the adaxial surface significantly reduces its surface undulance and hydrophobicity (Bringe et al. 2006), thus decreasing the chances of infection on this side of the leaf. This explanation corroborates the less severe symptoms of GLS on the adaxial side of older leaves.
It was observed that under light microscopy the inferior epidermis of the leaves is different and morphologically thinner when compared to the upper epidermis. The importance of the epidermis in the development of diseases was reported by Zanão Junior (2007), who indicated that when the epidermis is less thick, the incubation period may be shorter. Infection on the abaxial side of leaves is also important for other fungi, such as Phytophthora capsici in rubber tree. In young leaflets, the infection can occur by the abaxial side in both the stomata and directly by the cuticle. In older leaves, infection, if it occurs, is only possible on the abaxial side, since on the adaxial side there is a large cuticle deposition (Santos et al. 1999). In the present study it was possible to observe that the appressorium formation does not occur on stomata. The evaluation of the cuticle thickness in the abaxial and adaxial sides and in different cultivars inoculated with Colletotrichum spp. would be an interesting study for future works.
In stressed leaves observed in light microscopy the parenchyma presented fewer layers than in non-stressed leaves. Differences such as density, compaction and cell wall thickness in the parenchyma layer may result in differences in resistance to microorganisms (Prabhpreet et al. 2000).
In the field, infection on the abaxial side can be very important, since the main form of dissemination of the genus Colletotrichum is by water splashes (Timmer and Agostini 1994). Thus, there is a large movement of air and water, and droplets containing spores can reach the leaf tissue at the underside of the leaves. In addition, there is a practical implication in the management of the disease because when the spore is deposited on the lower surface of the leaves, they become difficult to reach by conventional spraying, especially in the case of contact products (Raetano and Bauer 2000). The main fungicides used to control GLS belong to the dithiocarbamate group, especially mancozeb (Katsurayama and Boneti 2009), which does not translocate through the plant. Ideally, in this case, it would be necessary to use equipment that would provide uniform coverage of the fungicides on both the adaxial and abaxial surfaces (Raetano and Bauer 2000).
The latent period for the species inoculated in leaves in the present study was not determined until 7 days after inoculation, but needs to be determined in future studies to better infer the importance of these species, since this period is a fundamental monocyclic component in the understanding of epidemics, as it influences their speed and duration (Amorim and Bergamin Filho 2018). In addition, epidemiological field studies and also molecular and biochemical aspects are important to discover why Colletotrichum isolates have different abilities to cause symptoms of BR and GLS.
Another relevant aspect that must be taken into account in future studies is that, in general, Colletotrichum species show different lifestyles that vary between groups of species, with most species being able to sequentially switch between lifestyles. The lifestyle patterns found in Colletotrichum species can be broadly categorized as necrotrophic, hemibiotrophic, latent, or quiescent and endophytic. Evidence suggests that the interaction between the host plant and the endophytic fungus can sometimes switch from mutualistic to antagonistic or pathogenic depending on the plant’s physiological condition, host genotype, and environmental conditions (De Silva et al. 2017).
In most cases, differences in host resistance to Colletotrichum appear only during colonization of the infective hyphae in epidermal and parenchyma cells (Bentes and Matsuoka 2002; Gomes et al. 2009; Wharton et al. 2001; Wharton and Dieguez-Uribeondo 2004). For Colletotrichum species, two basic colonization strategies have been reported: intracellular hemibiotrophy (IHB) and subcuticular-intramural necrotrophy (SIN). In the IHB strategy, the penetration of infective hyphae occurs in the epidermal cell, whereas in the SIN colonization, the fungus grows beneath the cuticle, within the periclinal and anticlinal walls of epidermal cells. Although species of Colletotrichum usually colonize the host through the IHB, the SIN pattern or the use of the two strategies of colonization has been reported for some species (Araujo et al. 2014; Diéguez-Uribeondo et al. 2005; Mendgen and Hahn 2002; Peres et al. 2005; Wharton and Dieguez-Uribeondo 2004). In the SIN strategy, the interaction is largely necrotrophic and the biotrophic phase is too short, or even non-existent (Wharton and Dieguez-Uribeondo 2004). Accordingly, plants of apples infected with C. gloeosporioides exhibit disease symptoms as early as 45 h after inoculation (Becker et al. 2000), indicating that the fungus follows the SIN-strategy of tissue colonization.
Unravelling the apple-Colletotrichum interaction and the different lifestyles can help to understand the roles that species plays as pathogen, and consequently to better manage the disease in the field. The present work presents an important step in the initial study of infection and lesion formation in apple leaves and fruit, mainly because it involves the study of 5 different species of Colletotrichum isolated from apple. This work highlighted important aspects about the species’ aggressiveness and raised new questions for future work.
References
Alaniz, S., Hernandez, L., Damasco, D., & Mondino, P. (2012). First report of Colletotrichum acutatum and C. fragariae causing bitter rot of apple in Uruguay. Plant Disease, 96, 458.
Amorim, L., & Bergamin Filho, A. (2018). Epidemiologia: análises temporal e espacial. In Lilian Amorim, Jorge Alberto Marques Rezende, Armando Bergamin Filho. (Org.). Manual de Fitopatologia: Princípios e Conceitos. 5ed. Ouro Fino: Agronômica Ceres Ltda, v. 1, p. 520-530.
Araujo, L., Gonçalves, A. E., & Stadnik, M. J. (2014). Ulvan effect on conidial germination and appressoria formation of Colletotrichum gloeosporioides. Phytoparasitica, 42, 631–640.
Baggio, J., Gonçalves, F. P., Lourenço, S. A., Tanaka, F. A. O., Pascholati, S. F., & Amorim, L. (2015). Direct penetration of Rhyzopus stolonifer into stone fruits causing Rhyzopus rot. Plant Pathology, 65, 633–642.
Baroncelli, R., Sreenivasaprasad, S., Thon, M. R., & Sukno, S. A. (2014). First report of apple bitter rot caused by Colletotrichum godetiae in the United Kingdom. Plant Disease, 98, 1000.
Bassanezi, R. B., Amorim, L., Bergamin Filho, A., & Hau, B. (1998). Effects of bean line pattern mosaic virus on the monocyclic components of rust and angular leaf spot of Phaseolus bean at different temperatures. Plant Pathology, 47, 289–298.
Becker, W. F., Katsurayama, Y., & Boneti, J. I. S. (2000). Mancha foliar da Gala: principal doença de verão da cultura da macieira. Agropecuária Catarinense, 13, 14–20.
Bedendo, I. P., Amorim, L., & Mattos Junior, D. (2018). Ambiente e Doença. In: Lilian Amorim, Jorge Alberto Marques Rezende, Armando Bergamin Filho. (Org.). Manual de Fitopatologia: Princípios e Conceitos. 5ed. Ouro Fino: Agronômica Ceres Ltda, v. 1, p. 93-103.
Bentes, J. L. S., & Matsuoka, K. (2002). Histology of Colletotrichum guaranicola and Paullinia cupana var. sorbilis on resistant and susceptible clones. Fitopatologia Brasileira, 27, 71–77.
Bragança, C. A. D., Damm, U., Baroncelli, R., Massola Júnior, N. S., & Crous, P. W. (2016). Species of the Colletotrichum acutatum complex associated with anthracnose diseases of fruit in Brazil. Fungal Biology, 120, 547–561.
Bringe, K., Schumacher, C. F. A., Schmitz-Eiberger, M., Steiner, U., & Oerke, E.-C. (2006). Ontogenetic variation in chemical and physical characteristics of adaxial apple leaf surfaces. Phytochemistry, 67, 161–170.
Cannon, P. F., Damm, U., Johnston, P. R., & Weir, B. S. (2012). Colletotrichum current status and future directions. Studies in Mycology, 73, 181–213.
Damm, U., Cannon, P. F., Woudenberg, J. H. C., & Crous, P. W. (2012). The Colletotrichum acutatum species complex. Studies in Mycology, 73, 37–113.
De Silva, D. D., Crous, P. W., Ades, P. K., Hyde, K. D., & Taylor, P. W. J. (2017). Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biology Reviews, 31, 155–168.
Denardi, F., Berton, O., & Spengler, M. M. (2003). Resistencia genética a podridão amarga em maçã determinadas pela taxa de desenvolvimento da doença em frutos com e sem ferimentos. Revista Brasileira de Fruticultura, 25, 494–497.
Diéguez-Uribeondo, J., Förster, H., Soto-Estrada, A., & Adaskaveg, J. E. (2005). Subcuticular-intracellular hemibiotrophic and intercellular necrotrophic development of Colletotrichum acutatum on almond. Phytopathology, 95, 751–758.
Doyle, V. P., Oudemans, P. V., Rehner, S. A., & Litt, A. (2013). Habitat and host indicate lineage identity in Colletotrichum gloeosporioides from wild and agricultural landscapes in North America. PLoS One, 8, 62–94.
Freeman, S., Katan, T., & Shabi, E. (1998). Characterization of Colletotrichum species responsible for anthracnose diseases of various fruits. Plant Disease, 82, 596–605.
Furlan, C. R. C., Mesquita Dantas, A. C., Denardi, F., Becker, W. F., & Mantovani, A. (2010). Resistência genética dos acessos do banco de germoplasma de macieira da epagri à mancha foliar de glomerella (Colletotrichum gloeosporioides). Revista Brasileira de Fruticultura, 32, 507–514.
Gomes, S., Prieto, P., Martins-Lopes, P., Carvalho, T., Martin, A., & Guedes-Pinto, H. (2009). Development of Colletotrichum acutatum on tolerant and susceptible Olea europaea L. cultivars: A microscopic analysis. Mycopathologia, 168, 203–211.
Gonzalez, E., & Sutton, T. B. (1999). First report of Glomerella leaf spot (Glomerella cingulata) of apple in the United States. Plant Disease, 83, 1074–1074.
Gonzalez, E., Sutton, T. B., & Correl, J. C. (2006). Clarification of the etiology of Glomerella leaf spot and bitter rot of apple caused by Colletotrichum spp. based on morphology and genetic, molecular and pathogenicity tests. Phytopathology, 96, 982–992.
Grahovac, M., Indic, D., Vukovic, S., Hrust, J., Gvozdenac, S., Mihajlovic, M., & Tanovi, B. (2012). Morphological and ecological features as differentiation criteria for Colletotrichum species. Zemdirbyste-Agriculture, 99, 189–195.
Hamada, N. A., Moreira, R. R., Gomes Figueiredo, J. A., & May De Mio, L. L. (2020). Colletotrichum acutatum complex isolated from apple flowers can cause bitter rot and glomerella leaf spot. Bragantia, 79, 399–406.
Jain, S., & Kumar, A. (2015). The pathogenesis related class 10 proteins in plant defense against biotic and abiotic stresses. Advanced Plants Agricultural Research, 2, 305–314.
Jingjing, J., Hongyan, Z., Huannan, L., Zhenhua, W., Yongsen, C., Ni, H., Guoping, W., Gilbert, N., & Wenxing, X. (2014). Identification and characterization of Colletotrichum fructicola causing black spots on young fruits related to bitter rot of pear (Pyrus bretschneideri Rehd.) in China. Crop Protection, 58, 41–48.
Karnovsky, M. J. (1965). A formaldehyde–glutaraldehyde fixative of high osmolality for use in electron microscopy. The Journal Cell Biology, 27, 137–138.
Katsurayama, Y., & Boneti J. I. S. (2009). Manejo das doenças de verão da macieira no sul do Brasil. In: Stadnik, M. J. Manejo integrado de doenças da macieira. Florianópolis-SC: Federal University of Santa Catarina, p. 45–64.
Katsurayama, Y., & Boneti, J. I. S. (2010). Model for forecasting the leaf spot disease in apple trees based on temperature and leaf wetness duration. Agropecuária Catarinense, 23, 82–84.
Kim, K. W. (2008). Vapor fixation of intractable fungal cells for simple and versatile scanning electron microscopy. Journal of Phytopathology, 156, 125–128.
Kou, L., Verneta, G., Luo, Y., & Jurick, W. (2014). First report of Colletotrichum fioriniae causing postharvest decay on ‘Nittany’ apple fruit in the United States. Plant Disease, 98, 993.
Kowata, L. S., Strapasson, M., Challiol, M. A., & May De Mio, L. L. (2010). Glomerella leaf spot in apple: Validation of proposed diagrammatic scale and efficiency of fungicides. Ciência Rural, 40, 1502–1508.
Lakshmi, B. K. M., Reddy, P. N., & Prasad, R. D. (2011). Cross-infection potential of Colletotrichum gloeosporioides Penz. isolates causing anthracnose in subtropical fruit crops. Tropical Agricultural Research, 22, 183–193.
Leite, R. P., Tsuneta, M., & Kishino, A. Y. (1988). Ocorrência de mancha foliar de Glomerella em maicieira no estado do Paraná. Fundação Institito Agronômico do Paraná. Informe de Pesquisa, 81p.
Liao, C. Y., Chen, M. Y., Chen, Y. K., Wang, T. C., Sheu, Z. M., Kuo, K. C., Chang, P. F. L., Chung, K. R., & Lee, M. H. (2012). Characterization of three Colletotrichum acutatum isolates from Capsicum spp. European Journal of Plant Pathology, 133, 599–608.
Liu, J. J., & Ekramoddoullah, A. K. M. (2006). The family 10 of plant pathogenesis-related proteins: Their structure, regulation, and function in response to biotic and abiotic stresses. Physiological and Molecular Plant Pathology, 68, 3–13.
Liu, G., Kennedy, R., Greenshields, D. L., Peng, G., Forseille, L., Selvaraj, G., & Wei, Y. (2007). Detached and attached arabidopsis leaf assays reveal distinctive defense responses against hemibiotrophic Colletotrichum spp. Molecular Plant-Microbe Interactions, 20, 1308–1319.
McDonald, B. A., & Linde, C. (2002). Pathogen population genetics, evolutionary potential and durable resistance. Annual Review of Phytopathology, 40, 349–379.
Mendgen, K., & Hahn, M. (2002). Plant infection and the establishment of fungal biotrophy: Review. Trends in Plant Science, 10, 1–5.
Moreira, R. R., Peres, N. A., & May De Mio, L. L. (2019). Colletotrichum acutatum and C. gloeosporioides species complex associated with apple in Brazil. Plant Disease, 103, 268–275.
Moreira, R. R., Araldi, G. S., & May De Mio, L. L. (2020). Colletotrichum acutatum complex causing anthracnose on peach in Brazil. Australasian Plant Pathology, 49, 179–189.
Munir, M., Amsden, B., Dixon, E., Vaillancourt, L., & Ward Gauthier, N. A. (2016). Characterization of Colletotrichum species causing bitter rot of apples in Kentucky orchards. Plant Disease, 100, 2194–2203.
O’Brien, T. P., & Mc Cully, M. E. (1981). The study of plant structure principles and select methods (45p). Melbourne: Termarcrphi Pty.
Peres, N. A., Timmer, L. W., Adaskaveg, J. E., & Correll, J. C. (2005). Lifestyles of Colletotrichum acutatum. Plant Disease, 89, 784–796.
Petri, J. L., Leite, G. B., Couto, M., & Francescatto, P. (2011). Avanços na cultura da macieira no Brasil (pp. 48–56). Special Issue: Revista Brasileira de Fruticultura.
Phoulivong, S., McKenzie, E. H. C., & Hyde, K. D. (2012). Cross infection of Colletotrichum species; a case study with tropical fruits. Current Research in Environmental & Applied Mycology, 2, 99–111.
Prabhpreet, I., Dhillon, M., Singh, P. P., & Inder, P. (2000). Mesophyll characteristics of muskmelon leaf in relation to resistance/susceptibility to downy mildew. Plant Disease Research, 15, 18–23.
Raetano, C. G., & Bauer, F. C. (2000). Effect of the air speed in air-assisted sprayer on bean crop pesticide. Bragantia, 62, 329–334.
Rockenbach, M. F., Boneti, J. I., Cangahuala-Inocente, G. C., Gavioli-Nascimento, M. C. A., & Guerra, M. P. (2015). Histological and proteomics analysis of apple defense responses to the development of Colletotrichum gloeosporioides on leaves. Physiological and Molecular Plant Pathology, 89, 97–107.
Santos, A. F., Matsuoka, K., Maffia, L. A., & Alfenas, A. C. (1999). Importância da estrutura da superfície foliar ima interação Phytophthora capsici - Hevea brasiliensis. Boletim de Pesquisa Florestal, 39, 67–80.
Sutton, T. B., & Sanhueza, R. M. V. (1998). Necrotic leaf blotch of Golden delicious e Glomerella leaf spot: A resolution of common names. Plant Disease, 82, 267–268.
Sutton, T. B., & Shane, W. W. (1983). Epidemiology of the perfect stage of Glomerella cingulata on apples. Phytopathology, 73, 1179–1183.
Sutton, B.C., Bailey, J.A., & Jeger, M.J. (1992). The genus Glomerella and its anamorph Colletotrichum. In: J.A. Bayley, M.J. Jeger (Eds.), Colletotrichum: Biology, pathology and control (pp. 1-26). Wallingford:CABI.
Taylor, J. (1971). A necrotic leaf blotch and fruit rot of apple caused by a strain of Glomerella cingulata. Phytopathology, 61, 221–224.
Timmer, L. W., & Agostini, J. P. (1994). Population dynamics and survival of strains of Colletotrichum gloeosporioides on citrus in Florida. Phytopathology, 84, 420–425.
Velho, A. C., Alaniz, S., Casanova, L., Mondino, P., & Stadnik, M. J. (2015). New insights into the characterization of Colletotrichum species associated with apple diseases in southern Brazil and Uruguay. Fungal Biology, 119, 229–244.
Velho, A. C., Mondino, P., & Stadnik, M. J. (2018). Extracellular enzymes of Colletotrichum fructicola isolates associated to apple bitter rot and Glomerella leaf spot. Mycology, 9, 145–154.
Weir, B. S., Johnston, P. R., & Damm, U. (2012). The Colletotrichum gloeosporioides species complex. Studies in Mycology, 73, 115–180.
Wharton, P. S., & Dieguez-Uribeondo, J. (2004). The biology of Colletotrichum acutatum. Anales del Jardin Botánico de Madrid, 61, 3–22.
Wharton, P. S., Julian, A. M., & O’Connell, R. J. (2001). Ultrastructure of the infection of Sorghum bicolor by Colletotrichum sublineolum. Phytopathology, 91, 149–158.
Zanão Junior, L. A. (2007). Resistência do arroz à mancha-parda mediada por Silício e Manganês. Viçosa:Federal University of Viçosa, Master thesis.
Ziadi, S., Poupard, P., Brisset, M. N., Paulin, J. P., & Simoneau, P. (2001). Characterization in apple leaves of two subclasses of PR-10 transcripts inducible by acibenzolar-S-methyl, a functional analogue of salicylic acid. Physiological and Molecular Plant Pathology, 59, 33–43.
Acknowledgments
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001, Fundação Araucária (FA), by Conselho Nacional de desenvolvimento Científico e tecnológico (CNPq), and by Casadinho/Procad - Chamada Pública MCT/CNPq/MEC/CAPES - Ação Transversal n°06/2011. Grant numbers: 483566/2013-6, 552334/2011-1, and 552334/2011-1. The authors would also like to thank Professor Dr. Nelson Sidnei Massola Junior, coordinator of the Phytopathogenic Fungi Laboratory of the Escola Superior de Agricultura Luiz de Queiros (ESALQ/USP), for providing the isolates used in this study.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
We declare that our manuscript complies to the all ethical rules applicable for this journal and that there are no potential conflicts of interest or even research involving human participants and/or animals.
Electronic supplementary material
ESM 1
(DOCX 13 kb)
Rights and permissions
About this article

Cite this article
Moreira, R.R., Zielinski, E.C., Castellar, C. et al. Study of infection process of five species of Colletotrichum comparing symptoms of glomerella leaf spot and bitter rot in two apple cultivars. Eur J Plant Pathol 159, 37–53 (2021). https://doi.org/10.1007/s10658-020-02138-y
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10658-020-02138-y