
Abstract
A nationwide monitoring with regard to the presence and distribution of ‘Candidatus Phytoplasma prunorum’ and its vector, Cacopsylla pruni, has been carried out in Germany between 2013 and 2017. In total, 286 sites with cultivated and wild Prunus species were surveyed. 806 plant samples covering 94 administrative districts in all 13 federal area states and one city-state were analysed by PCR. Furthermore, 3108 C. pruni were caught in 63 districts and tested for ‘Ca. P. prunorum’. European stone fruit yellows was found mainly in symptomatic apricot, plum, almond and peach but also for the first time in Germany in sweet cherry. Non-symptomatic infections were detected in 437 randomly sampled P. spinosa at 104 sites without any stone fruit growing nearby representing a natural infection rate of 14%. The vector C. pruni was found on all P. spinosa plants at 62 sites spread across Germany. The infection rate of C. pruni was 1–2% regardless whether the insects were caught in stone fruit growing areas or at wild habitats. Our results strongly indicate that ‘Ca. P. prunorum’ is widespread in all natural habitats in Germany and that wild P. spinosa represent a major source of infection in stone fruit orchards. A representative sample of 1164 individuals of C. pruni captured at 100 sites was molecularly typed: all but one insect were of B-type, the one which is the proven vector of ‘Ca. P. prunorum’. C. pruni A-type was detected for the first time in Germany near the French border.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Several economically important disorders on Prunus are collectively referred to as European stone fruit yellows (ESFY) (Lorenz et al. 1994; Cieślińska 2011; Marcone et al. 2011): apricot chlorotic leaf roll of apricot (Prunus armeniaca), leptonecrosis of Japanese plum (P. salicina) and yellows and decline diseases of peach (P. persica), European plum (P. domestica), almond (P. dulcis) and flowering cherry (P. serrulata). A unique pathogen is associated with ESFY, the phytoplasma ‘Candidatus Phytoplasma prunorum’ (Seemüller and Schneider 2004). The incidence of ‘Ca. P. prunorum’ depends on the region and species of the tree. Particularly susceptible are apricot, peach and Japanese plums. Severe outbreaks were observed since 1977 especially in the Mediterranean basin (Marcone et al. 2010, 2011). Due to its high economic importance ‘Ca. P. prunorum’ is currently regulated as quarantine organism in the EU in Annex IAII of Council Directive 2000/29/EC (Anonymous 2000; Steffek et al. 2012).
Infected trees show numerous symptoms depending on the susceptibility of the plant species, the rootstock and scion cultivar, virulence of the infecting strain, age of the tree and climatic conditions. The typical symptoms which can be observed in most of the infected Prunus species are off-season growth and premature break of leaf buds before flowering. In summer, leaf yellowing or reddening in combination with leaf roll is the common symptom (Marcone et al. 2011). These symptoms are highly correlated with the presence of the phytoplasma as detected by molecular means (Jarausch et al. 1998; Jarausch et al. 2008). Infected trees may also show less specific symptoms like leaf deformation, reduced terminal growth, die-back and decline.
ESFY is known to occur in most southern and central European countries (Cieślińska 2011; Marcone et al. 2010, 2011) but it has also been detected in Asia Minor (Jarausch et al. 2000; Sertkaya et al. 2005; Tedeschi et al. 2013; Allahverdi et al. 2014; Valasevich and Schneider 2016) as well as in northern Africa (Ben Khalifa et al. 2011). Its highest spread is in the Mediterranean basin while its northern border ranges from England (Davies and Adams 2000) via Germany (Jarausch et al. 2007b) to Poland (Cieślińska and Morgaś 2011). In Germany, the first detection of the agent in different Prunus species has already been reported in 1992 by Lederer and Seemüller.
Carraro et al. (1998) identified the psyllid species Cacopsylla pruni (Scopoli) as vector for ‘Ca. P. prunorum’ in Italy and Jarausch et al. (2001) confirmed the vector capacity of this psyllid species in France. In Germany, C. pruni was confirmed as vector in transmission trials conducted by us (Jarausch et al. 2007a). C. pruni is a European and Central Asian species which is known from almost all of Europe (Lauterer 1999). This univoltine psyllid is strictly oligophagous on Prunus spp. and overwinters as an adult on conifers (Jarausch and Jarausch 2016; Gallinger and Gross 2018). At the end of winter/early spring, C. pruni remigrants move from the overwintering plants back to Prunus for oviposition. The insects of the new generation feed on the reproduction hosts until the beginning of July when they leave the stone fruits as adults to move to overwintering hosts (Thébaud et al. 2009). The phytoplasma multiplies during winter in the psyllid making the returning migrants highly infective. The disease spread is regarded to be monocyclic (Thébaud et al. 2009; Jarausch et al. 2013). The natural infection rate of C. pruni varies according to the region: in France, Germany and Bulgaria low infection rates between 1 and 3% were observed (Yvon et al. 2004; Jarausch et al. 2007a, b; Thébaud et al. 2008; Etropolska et al. 2015), whereas from Italy (Carraro et al. 2004) and Austria (Maier et al. 2013) rates of more than 10% were reported.
Nation-wide surveys for the presence of ESFY disease have only been conducted in stone fruit growing areas, e.g. in France (Jarausch et al. 1998), Turkey (Ulubaş Serçe et al. 2006), Spain (Sabaté et al. 2015) or Bulgaria (Etropolska et al. 2015), and confirmed a wide-spread presence of ESFY in orchards of susceptible species like apricots, Japanese plum and peach. During a survey between 2000 and 2006, Jarausch et al. (2007b) could show that ‘Ca. P. prunorum’ as well as its vector, C. pruni, were present on all cultivated Prunus species in several stone fruit growing regions in Southwestern Germany.
The role of wild Prunus for the spread of ESFY was first studied by Carraro et al. (2002) in the heavily ESFY-infected region Friuli-Venezia Giulia in Northeast Italy. They found ‘Ca. P. prunorum’ as well as C. pruni in P. spinosa and P. cerasifera also at sites far from stone-fruit orchards and concluded that the cycle of ESFY can be completed independently from the presence of ESFY-infected cultivated stone-fruit trees. These results were confirmed by Yvon et al. (2004) in a case study in Southeastern France who detected ‘Ca. P. prunorum’-infected P. spinosa and C. pruni in a hedgerow of blackthorn. While Carraro et al. (2002) did not find ESFY-infected wild P. mahaleb, Sabaté et al. (2015) recently reported high infection rates from wild P. mahaleb bushes as well as from C. pruni caught on these plants from Catalonia, Spain. As these authors did not find infected P. spinosa, they concluded that P. mahaleb is a key factor in the local ESFY cycle in Spain.
In the present study, a nationwide monitoring was conducted between 2013 and 2017 including cultivated and wild Prunus spp. in order to elucidate the real presence and distribution of ‘Ca. P. prunorum’ and its vector C. pruni all over Germany. Based on this data it was the final goal to identify the epidemiological traits of ESFY disease and to contribute to the revision of its quarantine status in the EU.
Material & methods
The first part of the study was initiated 2010 by the AlPlanta Institute and comprised some selected fruit growing areas in Germany as continuation to those regions already published in Jarausch et al. (2007b, 2008). A comprehensive nationwide monitoring was started 2013 including plant and insect samples on symptomatic cultivated Prunus spp. from all German stone fruit growing regions in a first step. Between 2014 and 2017 the study was completed by a second part comprising plant and insect samples on wild non-symptomatic Prunus spp. located far from cultivated stone fruit areas all over Germany.
Monitoring of Prunus species
The monitoring of ESFY infection on cultivated Prunus spp. was based on surveying symptomatic plants in fruit growing areas all over Germany. Visual inspections as well as sampling of suspicious plants was basically done by plant health inspectors in their particular zone of responsibility and by farmers in their own orchards during summer before harvest. Branch samples of about 30 cm length were taken directly from symptomatic parts or from different parts all around a suspicious tree and sent to AlPlanta for molecular analyses. Localization of the sampling site was documented via GPS coordinates or exact description of the location and photographs. This way it was possible to establish precise maps of all sample sites in Germany. The monitoring of wild Prunus spp. was less evident as infected P. spinosa or P. cerasifera do not show symptoms. One part of this survey was focused on wild Prunus hedgerows in the vicinity of fruit trees. A second part of sampling included wild Prunus habitats far from fruit growing areas based on the experience of AlPlanta researchers with regard to putative risk pools and agglomeration sites for C. pruni. Branch samples of about 30 cm length were taken from wild Prunus spp. erraneously.
Captures of C. pruni
Based on data on C. pruni population dynamics in Germany (Jarausch et al. 2007b) insect monitoring started by mid-March on wild Prunus spp. – mostly P. spinosa- where the highest population of C. pruni remigrants appear after hibernation. Survey of C. pruni was undertaken on wild Prunus spp. and cultivated Prunus all over Germany from March until July including overwintered remigrants as well as individuals of the new generation (emigrants). Insect samples were taken by researchers of AlPlanta using the beating tray method. Captured psyllids were frozen at −20 °C and C. pruni identification was done using different determination keys (Ossiannilsson 1992; Burckhardt and Jarausch 2007).
Molecular detection of ‘Ca. P. prunorum’ in plant samples
Phloem was prepared from 1 to 3 branches per plant sample and total nucleic acids were extracted with a modified CTAB-based protocol as described by Jarausch et al. (2011). All plant samples were tested with the 16SrX group specific primers fO1/rO1 as published by Lorenz et al. (1995). Identification of ‘Ca. P. prunorum’ was done with specific primers ESFY-f/ESFY-r as described by Yvon et al. (2009). In samples with low phytoplasma concentration which gave only a faint band in direct PCR with primers fO1/rO1 the following nested PCR approach was adopted for ‘Ca. P. prunorum’ identification: a first PCR was performed using forward primer fO1 (Lorenz et al. 1995) and reverse primer P7 (Schneider et al. 1995) in a final volume of 20 μl with 1x Taq polymerase buffer with 1.5 mM MgCl2, 0.125 mM of each dNTP, 1 μM of each primer, 0.5 U of 5prime Taq polymerase (5prime, Germany) and 1 μl of total DNA in cycle conditions of 1 min at 95 °C for initial denaturation followed by 20 cycles at 95 °C for 15 s, 55 °C for 20 s and 72 °C for 1 min 30 s, with a final 4-min extension period at 72 °C. One microliter of the PCR reaction was further amplified for 40 cycles with the specific primers ESFY-f/ESFY-r as described by Yvon et al. (2009).
Molecular detection of ‘Ca. P. prunorum’ in insect samples
DNA was extracted from each psyllid individual with the modified CTAB-based protocol according to Jarausch et al. (2011) or with the TNES protocol, presented by Nicolas Sauvion at the COST FA0807 molecular identification training school in Montpellier, September 2012 (Sauvion 2012). PCR amplification of phytoplasma DNA was achieved with universal ribosomal primers fU5/rP7 (Lorenz et al. 1995; Schneider et al. 1995) as described in Jarausch et al. (2011) or with the 16SrX group specific primers fO1/rO1 as published by Lorenz et al. (1995). Identification of ‘Ca. P. prunorum’ was done as for plant samples with specific primers ESFY-f/ESFY-r as described by Yvon et al. (2009).
Molecular typing of C. pruni
Molecular typing of C. pruni specimen was done according to the protocol of Peccoud et al. (2013): the universal primer Cp480R (5’-TACATCCGAGGGTCGGTATC-3’) was used in triplex PCR together with the group-A specific primer CpA300F (5’- GGCCAGTAGTTAAACCGGACT-3’) and the group-B specific primer CpB120F (3’-TCCACGGGGTCCGCGATA-5’). The PCR was performed in a final volume of 20 μl with 1x Taq polymerase buffer with 2.5 mM MgCl2, 0.125 mM of each dNTP, 0.25 μM of each primer, 0.5 U of 5prime Taq polymerase (5prime, Germany) and 1 μl of total insect DNA. PCR cycles were as follows: initial denaturation at 95 °C for 1 min followed by 35 cycles at 95 °C for 15 s, 65 °C for 20 s and 72 °C for 30 s, with a final 4-min extension period at 72 °C. PCR products were analysed on 2% agarose gel stained with ethidium bromide for visualization under UV light. Specific PCR products of C. pruni type A had a size of 172 bp, whereas PCR products of C. pruni type B had a size of 377 bp. Positive controls for C. pruni types were kindly provided by Nicolas Sauvion (INRA Montpellier).
Results
ESFY detection in plant samples
In a first survey conducted in 2013 to 2014, plant samples were mainly obtained from cultured Prunus in stone fruit growing regions. Special attention was given to plants with typical ESFY symptoms like early bud break or chlorotic leaf roll, but plants with suspicious symptoms like pale leaves or non-chlorotic rolled leaves, were also sampled. In 2014, samples were also taken from wild Prunus which is predominantly Prunus spinosa and less common P. cerasifera in Germany. In particular P. spinosa does not show any ESFY symptoms. Therefore, samples were taken randomly from 5 to 10 plants per site. Wild Prunus were sampled near stone fruit growing areas but mainly in regions without stone fruit growing in order to monitor the spread of ESFY all over Germany. In 2015 to 2017, special surveys were conducted in non-stone fruit growing areas, e.g. in the North and East of Germany. All samples were tested with 16SrX group-specific primers and ESFY-specific primers. Table 1 gives a summary of the results obtained for all 13 federal area states plus the city of Berlin. ESFY infection was detected at 133 out of 258 sites in all states. More details about the sites and the results are given in Tab. S1.
Table 2 shows an analysis of the data with respect to the cultured Prunus species. Apricot (P. armeniaca) is the major stone fruit damaged by ESFY also in Germany. Samples were obtained from eight federal states and showed a relatively high infection rate of 67%. Peach (P. persica) is also very susceptible to ESFY but infection with ‘Ca. P. prunorum’ could only be confirmed in 36% of the samples. The third major ESFY-susceptible stone fruit crop in Germany is European plum (P. domestica). Although plum trees rarely show typical symptoms 43% of the samples tested positive. A special case is almond (P. amygdalus) which is grown in Southwest Germany for ornamental reasons. Almond trees show typical bud break symptoms and, accordingly, 100% of the samples were positive. Cherries are known to be fairly resistant to ESFY (Jarausch et al. 1999). Therefore, these crops were only sampled when suspicious symptoms were observed. These were chlorosis and sudden death of the plant. For the first time in Germany, ESFY infection in sweet cherry (P. avium) could be confirmed by specific PCR. Surprisingly, wild non-symptomatic P. spinosa in the neighborhood of stone fruit orchards were found in 37% of the cases to be infected with ‘Ca. P. prunorum’. This result prompted us to test P. spinosa more systematically in wild habitats where no stone fruit growing areas were nearby.
ESFY detection in insect samples
‘Ca. P. prunorum’ is transmitted only by one vector, the psyllid Cacopsylla pruni. The monitoring included therefore sampling of this vector for two reasons: 1) the distribution of C. pruni in Germany and thus the risk for ESFY spread should be elucidated, and 2) ESFY infection in wild habitats far from stone fruits growing areas should be detected by the presence of ‘Ca. P. prunorum’ in the vector. It is known that the major natural host plant of C. pruni is P. spinosa (Lauterer 1999; Carraro et al. 2002). Therefore, C. pruni was mainly captured on P. spinosa. A detailed list of the sampling sites is given in Tab. S2. Captures of C. pruni were predominantly done in 2014, only additional captures were done between 2015 and 2017. Table 3 gives a summary of the C. pruni samples and the detection of ‘Ca. P. prunorum’ in these individuals. C. pruni was found in all 12 federal area states tested plus Berlin. No sampling could be conducted in Mecklenburg-Vorpommern for technical reasons but the presence of C. pruni in this state is highly probable. Therefore, it can be concluded that C. pruni is well established in every region of Germany as it was found at all 97 sites tested. In total, 3108 insects were captured and individually tested by PCR. By this the natural infection rate could be determined. It ranged between 0.8 and 2.0%. However, infected C. pruni were only found at 30 out of 97 sites. This was probably due to the low number of individuals captured at many sites (s. Tab. S2). In 2014, 2516 remigrants were captured and 45 tested positive (1.8%) whereas only 481 emigrants could be captured from which 6 were positive (1.2%). A striking difference was observed between both sexes in 2014: whereas males encountered only for one third of the captures they had an infection rate of 2.8% (30 positive out of 1077). Females were predominantly captured but only 21 out of 1920 were infected (1.1%).
Sauvion et al. (2007) and Peccoud et al. (2013) reported on two different molecular types of C. pruni which might even be two different species. Therefore, a representative number of C. pruni individuals was molecularly typed as described by Peccoud et al. (2013). Table 4 gives a summary of the results. In total, 1163 individuals representing 100 different sites across Germany were of C. pruni B-type which is regarded to be the C. pruni reference type. However, for the first time in Germany an individual of C. pruni A-type was detected very close to the French border at Saarland.
Distribution of ESFY in Germany
The monitoring included plant and insect samples because it offered also the opportunity to sample at different times during the year and, thus, to sample at more places. Insects were sampled from March to May when they are on their host plant and plant samples were sampled in summer until autumn when symptoms are visible. Additional plant samples were obtained during winter until early spring when bud break symptoms occur. Thus, to show the pervasive distribution of ESFY in Germany, positive plant and insect data were merged. Fig. 1 shows the distribution of positive and negative sampling sites in Germany. It can be seen that ESFY is present all over the country independent of stone fruit growing regions.

Distribution of plant and insect samples tested positive or negative by specific PCR for ‘Ca. P. prunorum’ in Germany
Distribution of ESFY in wild habitats
One unexpected result of the monitoring was the nationwide presence of ESFY in wild habitats. Table 5 shows the data for plant and insect samples obtained from wild habitats separately. It can be seen that ESFY is present in all 13 federal area states in natural environment. This is further demonstrated in Fig. 2 were all data of wild habitats are shown. A total number of 437 asymptomatic P. spinosa samples was analysed for these areas and 14.4% of these samples were positive for ‘Ca. P. prunorum’. Natural infections of P. spinosa were found in every state where a substantial number of samples were collected. This signifies that ESFY is well established in the natural environment in Germany. It has been found at extreme places ranging from Alpine regions to coastal regions (s. Tab. S1). Accordingly, infected C. pruni were also detected in several states. The natural infection rate of C. pruni was 1.4%. In comparison, ‘Ca. P. prunorum’ infection in C. pruni captured on P. spinosa in fruit growing areas in 2014 was 2.0% (27 positive out of 1366). The infection rate of C. pruni from commercial apricot and plum orchards was 1.2% (three out of 243 samples; s. Tab. S2). Additional captures of C. pruni in an experimental apricot orchard with known ESFY spread at Neustadt (Rhineland-Palatinate) revealed an infection rate of remigrants of only 0.5% in 2014 (one out 192 individuals).

Distribution of ‘Ca. P. prunorum’ in wild habitats in Germany as revealed by specific PCR detection in plant and insect samples
Discussion
European stone fruit yellows is a quarantine disease which is regulated in Europe and other countries. It is reported from 11 EU and six non-EU countries (Steffek et al. 2012). None of these countries declares the disease to be ‘widespread’ probably because country-wide monitorings are missing. ESFY was first described in France (Chabrolin 1924) and a survey in France with molecular means showed that it was found in all areas tested (Jarausch et al. 1998). ESFY is well studied in southern countries with important apricot and peach growing where it causes large economic losses (Steffek et al. 2012), however, its distribution in northern countries with only local apricot growing areas is largely unknown. Germany is considered to be at the Northern border of ESFY distribution (Steffek et al. 2012). Therefore, the objective of the present work was to study with regard to the quarantine status of ESFY the nationwide distribution in Germany, in particular if ESFY-free regions exist in Germany or whether the disease is endemic.
Although European stone fruit yellows was first described in Germany in 1992 (Lederer and Seemüller 1992), the distribution of this quarantine disease was largely unknown. We conducted first surveys since 2000 in stone fruit orchards in Southwest Germany and found high infections in apricot orchards. ESFY was also detected in peach and European plum orchards as well as in almond grown for flowering (Jarausch et al. 2007b, 2008). We also confirmed the presence of C. pruni in Germany and proved by transmission trials its vector capacity (Jarausch et al. 2007a, 2008). During these surveys ESFY was detected only in one case also in wild Prunus cerasifera adjacent to stone fruit orchards. Thus, the question whether this quarantine pathogen was introduced into the orchards by latently infected planting material or by natural spread from the wild habitats remained unanswered.
To test for ESFY-free regions, samples were mainly obtained from non-stone fruit growing regions and wild habitats. As in addition cultured Prunus from orchards or planted cultivations like almonds for flowering were tested, the obtained result is a first nationwide overview of the distribution of ESFY in wild and cultured habitats. Our data clearly demonstrate that ESFY is endemic in Germany. It was not only found in major stone fruit growing regions but also in wild habitats far from any stone fruit growing. As the vector, C. pruni, is also widespread in Germany, a natural cycle of ESFY maintenance in wild Prunus is guaranteed. C. pruni is a European and Central Asian species which is known from almost all of Europe (Lauterer 1999). ‘Ca. P. prunorum’ might have been introduced to Europe from Asia along with the susceptible Prunus species like apricot and peach but ESFY has never been reported outside Europe apart from Asia Minor and North Africa. Our data strongly support the hypothesis that ESFY is an endemic European disease with a natural cycle including wild Prunus. As these wild autochtonous European Prunus like P. spinosa and P. cerasifera are tolerant to the disease, a coevolution between pathogen and plant host can be assumed. In Germany, the dominant and most widespread wild Prunus species is P. spinosa. This plant does not show any symptoms of ESFY (Carraro et al. 2002; Jarausch et al. 2008; this work) but can be infected. It has to be regarded as less susceptible. In addition, it is the preferred host plant of C. pruni (Lauterer 1999) and accordingly, we found C. pruni on every P. spinosa tested – sometimes at high population densities. This is supported by data from Carraro et al. (2002), Yvon et al. (2004) or Maier et al. (2013).
ESFY is widespread in wild habitats as we found in our random sampling a mean infection rate of about 14% and a nationwide distribution of infected plants. This is in the range of local infection rates of wild P. spinosa reported from other countries: 25% infections were found in France (Jarausch et al. 1998) and Italy (Carraro et al. 2002), 12% in Austria (Maier et al. 2013) whereas Yvon et al. (2004) reported only 3% infected P. spinosa in Southeastern France. These plants represent a risk for infections of susceptible stone fruit orchards such as apricot and peach. Although the population densities of C. pruni on P. spinosa were quite high the individual infection rate of the insects ranged only between 1 and 2%. This confirms our previous results (Jarausch et al. 2007a, b), only in heavily infected apricot orchards infection rates of up to 5% were found (Jarausch et al. 2007b). Similar low infection rates were observed in France by Jarausch et al. (2001), Yvon et al. (2004) and Thébaud et al. (2008) or in Bulgaria (Etropolska et al. 2015). By contrast, much higher infection rates were reported from Austria (Maier et al. 2013) and Italy (Carraro et al. 2004).
Despite this low infection rate infected remigrants represent a high risk for susceptible stone fruit orchards as the univoltine species overwinters on conifers (Jarausch and Jarausch 2016; Gallinger and Gross 2018) and remigrates to Prunus in early spring (Thébaud et al. 2009). At this time the individuals are highly infectious (Jarausch et al. 2007a; Thébaud et al. 2009) and the dispersal in the orchard is more or less randomly on a regional scale (Thébaud et al. 2006, 2009). Accordingly, we found high infection rates in susceptible stone fruits like apricot and Japanese plum confirming previous data from Southwest Germany (Jarausch et al. 2007b, 2008) as well as from other countries (Marcone et al. 2010, 2011). Our nationwide data confirm also the relatively low infection rate of the highly susceptible peach (Jarausch et al. 1998, 2007a, 2008). A new finding for Germany is ESFY infection of sweet cherry. In general, phytoplasma decline diseases of sweet and sour cherry remain unclear as different phytoplasmas have been found to be associated (Navrátil et al. 2001; Paltrinieri et al. 2008; Cieślińska and Morgaś 2011). An ESFY-related decline in sweet cherry was first observed in the Southwestern part of France and was called ‘Molières disease’ (Bernhard et al. 1977) but turned out by molecular means to be a stolbur type (Jarausch, personal comm.). However, ‘Ca. P. prunorum’ infection of sweet and sour cherry was confirmed in few declining trees in Czech Republic (Navrátil et al. 2001; Ludvíková et al. 2011) and in North-Central Italy (Paltrinieri et al. 2001). In contrast, experimental inoculations of thirteen sweet cherry cultivars with ‘Ca. P. prunorum’ demonstrated a high level of resistance in P. avium (Jarausch et al. 1999). We found two isolated cases of sudden decline in sweet cherry and could confirm infection with ‘Ca. P. prunorum’ with ESFY-specific primers. This indicates rather a hypersensitivity to ‘Ca. P. prunorum’ than a high resistance.
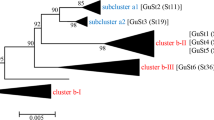
Sauvion et al. (2007) and Peccoud et al. (2013) hypothesised the existence of two species of C. pruni based on genetic analyses. These two C. pruni types A and B can easily be distinguished by a triplex PCR. However, it remains unclear whether both types can transmit ‘Ca. P. prunorum’ as transmission trials have been conducted before the identification of the two types. In Southern France both types coexist and might have been used in transmission trials (Peccoud et al. 2013). We made an exhaustive molecular typing of the C. pruni captured all over Germany and found with one exception only C. pruni B-type. The exception is a recent finding of A-type next to the French border. The A-type was for the first time found in a northern country outside of France. We therefore can conclude that our previous transmission trials have been carried out with the B-type. The B-type is also the dominant type across Europe (Peccoud et al. 2013) and has been recently reported to be the only type found in Bulgaria (Etropolska et al. 2016).
Also the phytoplasma is not homogenous. Multilocus sequence typing (MLST) of the marker genes imp, aceF, pnp and secY revealed at least 34 different haplotypes within the species ‘Ca. P. prunorum’ (Danet et al. 2011). Moreover, differences in strain virulence were described by Kison and Seemüller (2001) and others. Danet et al. (2011) and recently Dermastia et al. (2018) could correlate the aceF type A6 to hypo-virulent strains which induce no or only mild symptoms in Prunus. We contributed 22 German isolates covering 5 stone fruit growing regions with 16 apricot or peach orchards to the MLST analysis of Danet et al. (2011). Eight different haplotypes of ‘Ca. P. prunorum’ were identified. The main haplotype aceF A3-pnp P1-imp I1-secY S1 was found in all regions and was identified by Dermastia et al. (2018) as founder haplotype of ‘Ca. P. prunorum’. The dominant aceF types in Europe (A3 and A8) were also dominant in German isolates. The major imp type I1 in Europe accounted also for 77% of the German isolates. By contrast, the two A6 types identified in the analysis originated both from symptomatic trees indicating that this marker alone is not sufficient to characterize hypo-virulent strains (Dermastia et al. 2018). We conclude that the genetic variability of the German isolates of ‘Ca. P. prunorum’ has no impact on our results as it does not differ from other European regions.
We believe that our results are representative for central European countries. E.g., ESFY has been reported to be widely present in Czech Republic (Navrátil et al. 2001) and Poland (Cieślińska 2011). There is no reason to exclude natural ESFY infections also in northern countries like Scandinavian countries where P. spinosa and C. pruni are present (Steffek et al. 2012). Thus, ESFY should no longer be regarded as a quarantine pest. Our results have also important consequences for the protection of orchards planted with susceptible crops like apricot and peaches. Infection sources are not only infected trees inside the orchard but have to be looked for rather outside in the wild environment. Control of incoming C. pruni remigrants in spring is therefore of paramount importance.
References
Allahverdi, T., Rahimian, H., & Babaeizad, V. (2014). Association of Candidatus Phytoplasma prunorum associated with plum yellow leaf stunt in Iran. Iranian Journal of Plant Pathology, 50(2), 185.
Anonymous. (2000). Council directive 2000/29/EC of 8 May 2000 on protective measures against the introduction into the community of organisms harmful to plants and plant products and against their spread within the community. Official Journal of the European Communities, L 169, 1–112.
Ben Khalifa, M., Aldaghi, M., Hacheche, H., Kummert, J., Marrakchi, M., & Fakhfakh, H. (2011). First report of ‘Candidatus Phytoplasma prunorum’ infecting apricots in Tunisia. Journal of Plant Pathology, 93(2), 517–519.
Bernhard, R., Marénaud, C., Eymet, J., Fos, A., & Moutous, G. (1977). Une maladie complexe de certains Prunus: «Le dépérissement de Molières». Comptes Rendus de l’Académie d’Agriculture de France, Séance, 2(2), 178–189.
Burckhardt, D. & Jarausch, W. (2007). Pictorial key of central European Cacopsylla species associated with Rosaceae.- Online: http://www.psyllidkey.eu. Accessed 10 Jan 2019.
Carraro, L., Osler, R., Loi, N., Ermacora, P., & Refatti, E. (1998). Transmission of European stone fruit yellows phytoplasma by Cacopsylla pruni. Journal of Plant Pathology, 80, 233–239.
Carraro, L., Ferrini, F., Ermacora, P., & Loi, N. (2002). Role of wild Prunus species in the epidemiology of European stone fruit yellows. Plant Pathology, 51, 513–517.
Carraro, L., Ferrini, F., Labonne, G., Ermacora, P., & Loi, N. (2004). Seasonal infectivity of Cacopsylla pruni, vector of European stone fruit yellows phytoplasma. Annals of Applied Biology, 144, 191–195.
Chabrolin, C. (1924). Quelques maladies des arbres fruitiers de la Vallée du Rhône. Annales Epiphytes, 10, 265–333.
Cieślińska, M. (2011). European stone fruit yellows disease and its causal agent ‘Candidatus Phytoplasma prunorum’. Journal of Plant Protection Research, 51(4), 441–447.
Cieślińska, M., & Morgaś, H. (2011). Detection and identification of ‘Candidatus Phytoplasma prunorum’, ‘Candidatus Phytoplasma mali’ and ‘Candidatus Phytoplasma pyri’ in stone fruit trees in Poland. Journal of Phytopathology, 159(4), 217–222.
Danet, J. L., Balakishiyeva, G., Cimerman, A., Sauvion, N., Marie-Jeanne, V., Labonne, G., Laviňa, A., Battle, A., Križanac, I., Škorić, D., Ermacora, P., Ulubaş Serçe, C., Cağlayan, K., Jarausch, W., & Foissac, X. (2011). Multilocus sequence analysis reveals the genetic diversity of European fruit tree phytoplasmas and supports the existence of inter-species recombination. Microbiology, 157(2), 438–450.
Davies, D. L., & Adams, A. N. (2000). European stone fruit yellows phytoplasmas associated with a decline disease of apricot in southern England. Plant Pathology, 49, 635–639.
Dermastia, M., Dolanc, D., Mlinar, P., & Mehle, N. (2018). Molecular diversity of ‘Candidatus Phytoplasma mali’ and ‘Ca. P. prunorum’ in orchards in Slovenia. European Journal of Plant Pathology, 152, 791–800.
Etropolska, A., Jarausch, W., Jarausch, B., & Trenchev, G. (2015). Detection of European fruit tree phytoplasmas and their insect vectors in important fruit-growing regions in Bulgaria. Bulgarian Journal of Agricultural Science, 21, 1248–1253.
Etropolska, A., Jarausch, W., Jarausch, B., & Trenchev, G. (2016). Molecular typing of Bulgarian specimen of the phytoplasma vectors Cacopsylla pruni Scopoli and Cacopsylla melanoneura (Foerster). Bulgarian Journal of Agricultural Science, 22, 98–102.
Gallinger, J., & Gross, J. (2018). Unraveling the host plant alternation of Cacopsylla pruni – Adults but not nymphs can survive on conifers due to phloem/xylem composition. Frontiers in Plant Science, 9, 484. https://doi.org/10.3389/fpls.2018.00484.
Jarausch, W., & Jarausch, B. (2016). A permanent rearing system for Cacopsylla pruni, the vector of ‘Candidatus Phytoplasma prunorum’. Entomologia Experimentalis et Applicata, 159, 112–116.
Jarausch, W., Lansac, M., Saillard, C., Broquaire, J. M., & Dosba, F. (1998). PCR assay for specific detection of European stone fruit yellows phytoplasmas and its use for epidemiological studies in France. European Journal of Plant Pathology, 104(1), 17–27.
Jarausch, W., Eyquard, J. P., Mazy, K., Lansac, M., & Dosba, F. (1999). High level of resistance of sweet cherry (Prunus avium L.) towards European stone fruit yellows phytoplasmas. Advances in Horticultural Science, 13, 108–112.
Jarausch, W., Saillard, C., Broquaire, J. M., Garnier, M., & Dosba, F. (2000). PCR-RFLP and sequence analysis of a non-ribosomal fragment for genetic characterization of European stone fruit yellows phytoplasmas infecting various Prunus species. Molecular and Cellular Probes, 14, 171–179.
Jarausch, W., Danet, J. L., Labonne, G., Dosba, F., Broquaire, J. M., Saillard, C., & Garnier, M. (2001). Mapping the spread of apricot chlorotic leaf roll (ACLR) in southern France and implication of Cacopsylla pruni as a vector of European stone fruit yellows (ESFY) phytoplasmas. Plant Pathology, 50, 782–790.
Jarausch, B., Fuchs, A., Mühlenz, I., Lampe, I., Harzer, U., & Jarausch, W. (2007a). Research on European stone fruit yellows in Germany. Bulletin of Insectology, 60(2), 389–390.
Jarausch, B., Mühlenz, I., Fuchs, A., Lampe, I., Harzer, U., & Jarausch, W. (2007b). Untersuchungen zur Europäischen Steinobstvergilbung (ESFY) in Deutschland. Gesunde Pflanzen, 59, 183–192.
Jarausch, B., Mühlenz, I., Beck, A., Lampe, I., Harzer, U., & Jarausch, W. (2008). Epidemiology of European stone fruit yellows in Germany. Acta Horticulturae, 781, 417–422.
Jarausch, B., Schwind, N., Fuchs, A., & Jarausch, W. (2011). Characteristics of the spread of apple proliferation by its vector Cacopsylla picta. Phytopathology, 101, 1471–1480.
Jarausch, B., Sauvion, N., & Jarausch, W. (2013). Spread of European fruit tree phytoplasma diseases. Phytopathogenic Mollicutes, 3(1), 25–30.
Kison, H., & Seemüller, E. (2001). Differences in strain virulence of the European stone fruit yellows phytoplasma and susceptibility of stone fruit trees on various rootstocks to this pathogen. Journal of Phytopathology, 149(9), 533–541.
Lauterer, P. (1999). Results of investigations on Hemiptera in Moravia, made by Moravian Museum (Psylloidea 2). Acta Musei Moraviae, Scientiae Biologicae (Brno), 84, 71–151.
Lederer, M., & Seemüller, E. (1992). Demonstration of mycoplasmas in Prunus species in Germany. Journal of Phytopathology, 134(2), 89–96.
Lorenz, K.-H., Dosba, F., Poggi-Pollini, C., Llacer, G., & Seemüller, E. (1994). Phytoplasma diseases on Prunus species in Europe are caused by genetically similar organisms. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz, 101, 567–575.
Lorenz, K. H., Schneider, B., Ahrens, U., & Seemüller, E. (1995). Detection of the apple proliferation and pear decline phytoplasmas by PCR amplification of ribosomal and nonribosomal DNA. Phytopathology, 85, 771–776.
Ludvíková, H., Fránová, J., & Jana Suchá, J. (2011). Phytoplasmas in apricot, peach and sour cherry orchards in East Bohemia, Czech Republic. Bulletin of Insectology, 64(Supplement), S67–S68.
Maier, C., Bachinger, K., Mörtel, J., Engel, C., Czipin, L., & Riedle-Bauer, M. (2013). European stone fruit yellows in Austria: Epidemiological observations and a mark and recapture experiment tracking the dispersal of its vector Cacopsylla pruni (Hemiptera: Psyllidae) in a model apricot orchard. Journal of Phytopathology, 161, 713–722.
Marcone, C., Jarausch, B., & Jarausch, W. (2010). Candidatus Phytoplasma prunorum’, the causal agent of European stone fruit yellows: An overview. Journal of Plant Pathology, 92, 19–34.
Marcone, C., Jarausch, B., Jarausch, W., & Dosba, F. (2011). European stone fruit yellows phytoplasma. In A. Hadidi, M. Barba, T. Candresse, & W. Jelkmann (Eds.), Virus and virus-like diseases of pome and stone fruits (pp. 233–241). St. Paul: APS press ISBN 978-0-89054-396-2.
Navrátil, M., Válová, P., Fialová, R., Petrová, K., Poncarová-Voráčková, Z., Fránová, J., Nebesářová, J., & Karešová, R. (2001). Survey for stone fruit phytoplasmas in the Czech Republic. Acta Horticulturae, 550, 377–382.
Ossiannilsson, F. (1992). The Psylloidea (Homoptera) of Fennoscandia and Denmark. Fauna Entomologica Scandinavica, 29, 1–346.
Paltrinieri, S., Martini, M., Stefani, E., Fideghelli, C., Pondrelli, M., & Bertaccini, A. (2001). Phytoplasma infection in peach and cherry in Italy. Acta Horticulturae, 550, 365–369.
Paltrinieri, S., Bertaccini, A., & Lugaresi, C. (2008). Phytoplasmas in declining cherry plants. Acta Horticulturae, 781, 409–416.
Peccoud, J., Labonne, G., & Sauvion, N. (2013). Molecular test to assign individuals within the Cacopsylla pruni complex. PLoS One, 8(8), e72454. https://doi.org/10.1371/journal.pone.0072454.
Sabaté, J., Laviña, A., & Batlle, A. (2015). Incidence and distribution of ‘Candidatus Phytoplasma prunorum’ and its vector Cacopsylla pruni in Spain: An approach to the epidemiology of the disease and the role of wild Prunus. Plant Pathology, 65(5), 837–846.
Sauvion, N. (2012). Minutes of the training school for molecular identification of psyllid vectors, Montpellier. (http://www.costphytoplasma.ipwgnet.org/WG2.htm). Accessed 10 Jan 2019
Sauvion, N., Lachenaud, O., Genson, G., Rasplus, J.-Y., & Labonne, G. (2007). Are there several biotypes of Cacopsylla pruni? Bulletin of Insectology, 60(2), 185–186.
Schneider, B., Seemüller, E., Smart, C. D., & Kirkpatrick, B. C. (1995). Phylogenetic classification of plant pathogenic mycoplasma-like organism or phytoplasmas. In S. Razin & J. G. Tully (Eds.), Molecular and diagnostic procedures in Mycoplasmology (pp. 369–380). San Diego: Academic Press.
Seemüller, E., & Schneider, B. (2004). Candidatus Phytoplasma mali’, ‘Candidatus Phytoplasma pyri’ and Candidatus Phytoplasma prunorum’, the causal agents of apple proliferation, pear decline and European stone fruit yellows, respectively. International Journal of Systematic and Evolutionary Microbiology, 54, 1217–1226.
Sertkaya, G., Martini, M., Ermacora, P., Musetti, R., & Osler, R. (2005). Detection and characterization of phytoplasmas in diseased stone fruits and pear by PCR-RFLP analysis in Turkey. Phytoparasitica, 33(4), 380–390.
Steffek, R., Follak, S., Sauvion, N., Labonne, G., & MacLeod, A. (2012). Distribution of ‘Candidatus Phytoplasma prunorum’ and its vector Cacopsylla pruni in European fruit-growing areas: A review. EPPO Bulletin, 42(2), 191–202.
Tedeschi, R., Jarausch, B., Delic, D., & Weintraub, P. (2013). Actual distribution of fruit tree and grapevine phytoplasma diseases and their vectors in Europe and neighboring regions. Phytopathogenic Mollicutes, 3(1), 3–4.
Thébaud, G., Sauvion, N., Chadoeuf, J., Dufils, A., & Labonne, G. (2006). Identifying risk factors for European stone fruit yellows from a survey. Phytopathology, 9, 890–899.
Thébaud, G., Yvon, M., Labonne, G., & Alary, R. (2008). European stone fruit yellows: Consequences of the life cycle of the vector and of the multiplication of the phytoplasma in the insect on the epidemiology of the disease. Acta Horticulturae, 781, 423–428.
Thébaud, G., Yvon, M., Alary, R., Sauvion, N., & Labonne, G. (2009). Efficient transmission of ‘Candidatus Phytoplasma prunorum’ is delayed by eight months due to a long latency in its host-alternating vector. Phytopathology, 99, 265–273.
Ulubaş Serçe, Ç., Gazel, M., Çaglayan, K., Baş, M., & Son, L. (2006). Phytoplasma diseases of fruit trees in germplasm and commercial orchards in Turkey. Journal of Plant Pathology, 88(2), 179–185.
Valasevich, N., & Schneider, B. (2016). Detection, identification and molecular diversity of ‘Candidatus Phytoplasma prunorum’ in Belarus. Journal of Plant Pathology, 98(3), 625–629.
Yvon, M., Labonne, G., & Thébaud, G. (2004). Survival of European stone fruit yellows phytoplasma outside fruit crop production areas: A case study in southeastern France. Acta Horticulturae, 657, 477–481.
Yvon, M., Thébaud, G., Alary, R., & Labonne, G. (2009). Specific detection and quantification of the phytopathogenic agent ‘Candidatus Phytoplasma prunorum’. Molecular and Cellular Probes, 23(5), 227–234.
Acknowledgements
The nationwide monitoring was supported from 2013 to 2014 by a fund provided by the Julius Kühn-Institute (JKI) for national and international Plant Health. A. Etropolska was supported by a DAAD grant. The production of the distribution maps by Peter Horney (JKI Kleinmachnow) is greatly acknowledged. The authors would like to thank the following staff members of the German Plant Protection Services and the JKI for providing plant and insect samples:
Hans-Dieter Beuschlein, Werner Dahlbender, Uwe Dederichs, Conni Dippel, Paul Epp, Michael Fischbach, Peter Göbel, Uwe Harzer, Manfred Hellmann, Georg Henkel, Günther Hensel, Anne-Kathrin Hentsch, Armin Hofhansel, Stefanie Lapcik, Eveline Maring, Maureen Möwes, Sigrid von Norsinski, Eva Satzl, Elfie Schell, Franz-Josef Scheuer, Manfred Schröder, Bernd Schumacher, Sonja Schurig, Gerhard Steinecke, Silke Steinmöller, Martin Trautmann, Katrin Veit, Elke Weddell, Anne Wilstermann and Simone Wittke.
Funding
This study was supported from 2013 to 2014 by a fund provided by the Julius Kühn-Institute (JKI) for national and international Plant Health. Work conducted from 2015 to 2017 was funded by RLP AgroScience itself. A. Etropolska was supported by a DAAD grant.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The manuscript has not been submitted to another journal for simultaneous consideration.
The manuscript has not been published previously. However, a conference abstract on preliminary data has been published in 2014. This abstract does not contain any concrete data. Thus, the present manuscript contains the entire data which are true and accurate to the knowledge of the authors.
A single study is not split up into several parts to increase the quantity of submissions and submitted to various journals or to one journal over time.
Any work (data, text, or theories) of others besides the authors has been properly acknowledged.
All authors (including their responsible authorities) listed in the title page agreed to be named as authors on this manuscript.
Authors whose names appear on the submission have contributed sufficiently to the scientific work and therefore share collective responsibility and accountability for the results.
Authors ensure the correct author group, corresponding author, and order of authors at submission.
Authors are aware that changes of authorship or in the order of authors are not accepted after acceptance of a manuscript.
Authors are aware that upon request they should be prepared to send relevant documentation or data in order to verify the validity of the results.
Authors are aware of the COPE guidelines.
Conflict of interest
The authors declare that they have no conflict of interest.
Informed consent
Informed consent was obtained from all individual participants included in the study.
Rights and permissions
About this article

Cite this article
Jarausch, W., Jarausch, B., Fritz, M. et al. Epidemiology of European stone fruit yellows in Germany: the role of wild Prunus spinosa. Eur J Plant Pathol 154, 463–476 (2019). https://doi.org/10.1007/s10658-019-01669-3
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10658-019-01669-3