
Abstract
The primary mechanism of biocontrol by Pseudomonas fluorescens strains HC1–07 and HC9–07 is production of a cyclic lipopeptide (CLP) and phenazine-1-carboxylic acid, respectively. We introduced the seven-gene operon for the synthesis of phenazine-1-carboxylic acid (PCA) from P. synxantha 2–79 into Pseudomonas fluorescens HC1–07rif to determine if the biocontrol activity of the recombinant strain HC1–07PHZ increased against Gaeumannomyces graminis var. tritici, causal agent of take-all disease of wheat. In vitro inhibition assays showed that strain HC1–07PHZ consistently inhibited the hyphal growth of three isolates of the take-all pathogen on plates of both PDA and KMB, and the recombinant consistently inhibited G. graminis. var. tritici to a greater extent than the wild type. Strain HC1–07PHZ applied at a dose of 102 CFU seed−1 suppressed take-all better than strains HC1–07rif and HC9–07rif applied either individually or in combination. When the dose of the bacteria was increased to 104 CFU seed−1, the strain combination HC1–07rif + HC9–07rif showed significantly better disease suppression than did HC1–07rif, HC9–07rif or HC1–07PHZ applied individually. However, when the bacterial dose was increased to 107 CFU seed−1, strains HC1–07rif and HC1–07PHZ showed significantly better disease suppression than HC9–07rif and the combination HC1–07rif + HC9–07rif.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Take all, caused by Gaeumannomyces graminis var. tritici (Sacc.) von Arx & Olivier var. tritici Walker (G. graminis var. tritici), is considered one of the most important root diseases of wheat in the USA and worldwide (Ramsey 2001; Cook 2003; Freeman and Ward 2004; Kwak et al. 2009; Weller 2015). Crops including barley, rye and triticale are also susceptible to G. graminis var. tritici. Take-all is controlled by crop rotation and tillage, but the modern farming practices of using less tillage for erosion control, and growing several crops of wheat or barley before a break, greatly exacerbate the incidence and severity of the disease. Take-all can also be controlled by take-all decline (TAD), which is the spontaneous decrease in the incidence and severity of take-all that occurs with monoculture of wheat or barley after a severe outbreak of the disease (Weller 2015; Weller et al. 2002). TAD fields owe their suppressiveness to a build-up of pseudomonads in the Pseudomonas fluorescens group (Loper et al. 2012) that produce the antibiotic 2,4-diacetylphloroglucinol (DAPG) (Raaijmakers et al. 1997; Weller et al. 2002; Weller 2015).
Beneficial bacteria have been studied intensively as biocontrol agents against soilborne diseases (Weller 2007; Weller and Thomashow 2015), and fluorescent Pseudomonas spp. are one of the most extensively investigated groups (Weller 2007). Pseudomonads are well adapted to colonizing roots and the rhizosphere, and they suppress disease by a wide variety of mechanisms (Pieterse et al. 2014; Weller 2007) but especially important are the production of antimicrobial metabolites. Antibiotics such as DAPG, pyrrolnitrin, pyoluteorin, phenazines and lipopeptides (LPs) are commonly produced by some of the most effective biocontrol pseudomonads both in culture (Raaijmakers et al. 2002; Raaijmakers et al. 2009; Raaijmakers et al. 2010; Haas and Defago 2005; D’aes et al. 2010; Wang et al. 2014) and in the rhizosphere (Kwak et al. 2012; Mavrodi et al. 2012). Phenazines are a diverse and colorful group of secondary metabolites of bacterial origin that have a broad spectrum of activity and function in the biocontrol of a wide range of plant pathogens including Rhizoctonia root rot of bean, take-all of wheat, Fusarium wilt of flax, Pythium myriotylum on cocoyam, and stem rot disease of groundnut (Tambong and Höfte 2001; Chin-A-Woeng et al. 2003; Weller et al. 2007; Mazurier et al. 2009; D'aes et al. 2011; Mavrodi et al. 2010, 2013; Le et al. 2012). One of the most widely studied phenazine producers is strain P. synxantha 2–79 (formerly known as P. fluorescens 2–79) (Parejko et al. 2012, 2013). Strain 2–79 produces the simplest of the phenazines, phenazine-1-carboxylic acid (PCA), which provides suppression of take-all (Thomashow and Weller 1988). PCA-producing strains are widely distributed on the roots of dryland wheat in the Pacific Northwest of the USA and the antibiotic accumulates on the roots (Mavrodi et al. 2012). Yang et al. (2011) conducted a survey of pseudomonads in Chinese wheat fields and unexpectedly found that PCA-producing Pseudomonas spp. exemplified by P. fluorescens HC9–07 were abundant on wheat, were phylogenetically similar to 2–79, produced PCA in the rhizosphere, and controlled take-all with PCA providing the suppression.
Cyclic lipopeptides (CLPs) are versatile molecules with antimicrobial, cytotoxic, and surfactant properties produced by a variety of bacterial genera including Pseudomonas spp. associated with plants (Raaijmakers et al. 2010). Many studies have demonstrated the involvement of biosurfactants in biological control of plant pathogens (Tambong and Höfte 2001; Tran et al. 2007; D'aes et al. 2011; Le et al. 2012; Olorunleke et al. 2015; Nam et al. 2016). Yang et al. (2014) showed that a viscosin-like CLP was the primary mechanism of suppression in the biocontrol of take all on wheat by P. fluorescens strains HC1–07, however the viscosin-like CLP appeared to have less of a role in the control of Rhizoctonia root rot. This strain was isolated from the same field from which the PCA producer HC9–07 was isolated. Perneel et al. (2008) demonstrated that both phenazines and biosurfactants in Pseudomonas aeruginosa PNA1 were important in the biocontrol of Pythium splendens on bean and Pythium myriotylum on cocoyam. A biosurfactant-deficient mutant and a phenazine-deficient mutant of PNA1 applied individually did not suppress disease, but when the two mutants were concurrently introduced in the soil, the biocontrol activity was restored to wild-type levels. Xu et al. (2011) demonstrated the involvement of CLPs during Pseudomonas CMR12a-mediated biocontrol of Rhizoctonia root rot of bean. Olorunleke et al. (2015) further investigated the interplay of phenazines and CLPs (orfamides and sessilins) produced by CMR12a in two pathosystems: damping-off disease on Chinese cabbage (Brassica chinensis) and root rot disease on bean (Phaseolus vulgaris L.). They found that on cabbage, phenazines alone were sufficient to suppress Rhizoctonia damping-off, whereas the co-production of the two CLPs was required for disease suppression in the absence of phenazines. In contrast, in the Rhizoctonia root rot of bean pathosystem, the co-production of phenazines, sessilins and orfamides was important for effective disease suppression. These results suggest that for suppression of some diseases and on some crops, CLPs and phenazines operate in concert to promote disease control.
Our observations that both CLP- and PCA-producing pseudomonads are common in Chinese wheat fields (Yang et al. 2011, 2014; Yang 2011) and that these metabolites play a key role in the biocontrol of take-all by the Chinese strains HC1–07 and HC9–07, prompted us to determine whether combining the two biocontrol traits into a single strain of Pseudomonas would enhance the biocontrol activity of the recombinant against G. graminis var. tritici. We stably introduced phenazine biosynthetic operon from P. synxantha 2–79 into HC1–07 and showed a significant increase in biocontrol activity against take-all as compared to the wild-type strain. However, the activity was affected by the bacterial dose applied to the wheat seed.
Material and methods
Bacterial strains, plasmids and fungi
Strains and plasmids used in this study are listed in Table 1. Escherichia coli strains were routinely grown at 37 °C on Luria-Bertani (LB) agar or broth (Ausubel et al. 2002). The spontaneous rifampicin-resistant derivative of P. fluorescens HC1–07 and HC9–07 with growth characteristics similar to those of wild-type strains were used through this study. Pseudomonas strains were cultured on King’s medium B (KMB) or M9 media at 27 °C (King et al. 1954; Ausubel et al. 2002). When needed, antibiotics were used at the following concentrations: chloramphenicol (13 μg ml−1), tetracycline (25 μg ml−1), ampicillin (40 μg ml−1), cycloheximide (100 μg ml−1), kanamycin (100 μg ml−1) and rifampicin (100 μg ml−1) (Table 1). All bacteria were stored at −80 °C in LB broth supplemented with 40% (vol/vol) glycerol. G. graminis var. tritici isolates LD5, ARS-A1, and R3-111a-1 (Kwak et al. 2012) were grown on commercial or home-made one-fifth-strength potato dextrose agar (1/5 PDA) (Yang et al. 2011). Briefly, the commercial PDA was made from potato dextrose broth (Becton Dickinson, Sparks, MD) with agar (15 g/L) and adjusted to pH 6.5. To prepare homemade PDA, diced potato (200 g) was cooked in water in a microwave for 15 min, the resultant broth was strained and brought to 1 l, then supplemented with dextrose 20 g, and agar 15 g. The pH was adjusted to 6.5, then the medium was autoclaved. Fungi were stored at 4°Con 1/5-strength PDA and amended with rifampicin (100 μg ml−1) (Kwak et al. 2009).
Construction of recombinant phenazine-producing derivatives of P. fluorescens HC1–07rif
The phenazine biosynthetic operon phzABCDEFG from P. synxantha 2–79 was cloned into pUC18Sfi (Mavrodi et al. 1998), and then moved into the SfiI site of a mini-Tn5Km1, yielding pUTKm-phz. This plasmid was transformed into E. coli S17–1, and mobilized into P. fluorescens HC1–07rif by using a biparental filter mating technique (Huang et al. 2004). Transformants were selected on M9 minimal medium supplemented with kanamycin, and isolates containing presumptive transposed phz genes in genomic DNA were transferred ten times on King’s Medium B agar to assure the stability of kanamycin resistance in the isolates. Isolates also were compared morphologically to the wild type. The insertion of phz genes in each of the recombinant clones was confirmed by PCR with phenazine-gene primers Ps_up1 and Ps_low1 (Mavrodi et al. 2001) and phenazine production was confirmed by thin layer chromatography (TLC) plates. Growth kinetics of wild-type HC1–07 and its PCA-producing derivatives were compared in M9 and KMB broth in vitro (Huang et al. 2004).
In vitro inhibition of fungal pathogens
Inhibition of the growth of G. graminis var. tritici isolates LD5, ARS-A1, and R3-111a-1 (Kwak et al. 2012) by HC1–07rif, HC9–07rif and the HC1–07 PCA-producing derivative HC1–07PHZ were tested on homemade potato dextrose agar (PDA) and KMB agar plates (Yang et al. 2011). Briefly, a 0.5-cm plug from the leading edge of a culture of G. graminis var. tritici grown for 5 days at 24 °C on 1/5-strength PDA was placed in the center of the plate. After that, 5-μl drops from an overnight culture of each bacterium grown in KMB broth were spotted 1 cm from the edge of a Petri dish and allowed to soak into the agar (four spots per plate, each spot represents one isolate). Control plates were inoculated only with G. graminis var. tritici isolates. Plates were incubated at 24 °C and scored after 7 days by measuring the distance between the edges of the bacterial colony and the fungal mycelium. Each bacterial treatment was replicated three times and a single plate served as a replicate (Yang et al. 2011).
Isolation and detection of phenazine-1-carboxylic acid (PCA) in vitro
Extraction of PCA from P. fluorescens HC9–07 and the PCA-producing recombinant of HC1–07 (HC1–07PHZ) was conducted as described by Yang et al. (2011). Briefly, HC9–07 and HC1–07PHZ were grown with shaking in test tubes containing 5 ml of LB broth supplemented with 0.2% glucose or KMB broth at 28 °C for 48 h. Cells were removed by centrifugation (5000 rpm for 5 min) and the culture supernatants were acidified with 45 μl of 10% trifluoroacetic acid. PCA was extracted twice with 10 ml of ethyl acetate. Organic phases were pooled and evaporated to dryness, and residues were suspended in methanol. PCA was detected by thin-layer chromatography (TLC). Twenty μl of methanol was added to the residue, the tubes were shaken to dissolve it, and 2 μl of each sample was loaded onto a Uniplate Silica Gel GHLF plate (Analtech, Inc., Newark, DE, USA). The plate was then placed in a glass tank containing a mix of benzene/acetic acid (95:5 by volume) and allowed to develop for 1 h. PCA was visualized by UV irradiation at 256 nm and migration on the plate was compared to a standard of pure PCA.
Preparation of oat-kernel inoculum of G. graminis Var. tritici
Inoculum of G. graminis var. tritici was prepared as described by Yang et al. (2011). Briefly, 250 ml of treated whole oat grains plus 350 ml of water were combined in a 1-l flask and autoclaved for 90 min twice on two successive days. Sterile oat grains were inoculated with agar disks of G. graminis var. tritici strain R3-111a-1. After 3–4 weeks of growth at room temperature, the contents of each flask were removed, air dried on paper in a laminar flow hood, and stored at 4 °C.
Bacterial treatment of wheat seed
For biocontrol tests in the greenhouse, wheat seeds (cv. Penawawa) were coated with bacteria at doses of approximately 102, 104, or 107 CFU/seed as described previously (Yang et al. 2011). Briefly, KMB plates were spread-inoculated with the test bacterium and incubated for 48 h at room temperature. Wheat seeds were thoroughly mixed with an appropriately-diluted cell suspension in 1% methylcellulose (MC) in a tube, and air-dried for several hours in a laminar flow hood. The final density of bacteria was checked at each inoculum density. Since, in previous experiments we found that control treatments of seeds coated with or without MC did not differ, we used non-MC coated seeds in this study.
Take-all biocontrol assay
A tube assay previously described by Yang et al. (2011) was used to determine the biocontrol activity of the bacterial strains against take-all. Briefly, plastic tubes (2.5 cm diam., 16.5 cm long), each with a hole at the bottom, were supported in a hanging position in racks (200 cones per rack). A cotton ball was used to plug the hole and then the tubes were filled with a 6.5-cm-thick column of sterile vermiculite followed by 10 g of air-dried and sieved Quincy Virgin soil (Shano sandy loam from a non-cropped virgin site near Quincy, WA). G. graminis var. tritici R3-111a-1 was blended into the soil as colonized oat kernels that had been pulverized in a blender and sieved into fractions of known particle sizes. The fraction 0.25–0.5 mm in size was added to the soil at 0.7% (w/w). Three wheat seeds treated with bacteria as described above were placed in each tube and covered with a 1.5-cm-thick topping of vermiculite. Each tube received 10 ml of water and then tubes were covered with plastic. The rack of tubes was incubated at 15–18 °C in a dark/light cycle of 12 h. After the plastic was removed, each cone received 10 ml of water and diluted (1:3, v/v) Hoagland’s solution (macroelements only) twice and once per week, respectively. As a control, we used non-MC coated seeds in this study. Each replicate of a treatment consisted of five tubes and each treatment was replicated at least three times for a total of 15 tubes. Treatments were arranged in a randomized complete block design and the experiment was repeated three times. After 3–4 weeks, plants were removed from the tubes, the roots were washed free of soil, and plants were evaluated for disease severity on a scale of 0–8 as previously described (Pierson and Weller 1994; Huang et al. 2004), where 0 = no disease evident, 8 = plants dead or nearly so.
Rhizosphere colonization assays
Strains HC1–07rif, HC9–07rif, HC1–07PHZ and a combination of strains HC1–07rif and HC9–07rif (1:1) were tested for the ability to colonize the rhizosphere of wheat grown under controlled conditions. Bacterial inocula were prepared and suspended in a solution of 1% methyl cellulose and then added to Quincy virgin soil (Shano sandy loam) as previously described (Landa et al. 2002; Yang et al. 2011) to obtain 104 CFU g−1 of soil. Control treatments consisted of soil amended only with a 1% methylcellulose suspension. The density of each introduced strain was determined by assaying 0.5 g of inoculated soil as described below. Spring wheat (cv. Penawawa) seeds were pre-germinated on moistened sterile filter paper in Petri dishes for 24 h in the dark, and were sown (six seeds per pot) in square pots (height, 6.5 cm; width 7 cm) containing 200 g of the Quincy virgin soil inoculated with one or two bacterial strains (Landa et al. 2002). Each treatment was replicated six times and each pot served as a replicate. Wheat was grown in a controlled environment chamber at 15 °C with a 12-h photoperiod. After 3 weeks of growth, the roots were harvested and immediately processed, and the population density of each bacterial strain was determined by the dilution-endpoint assay as described by Mavrodi et al. (2006). Briefly, for each replicate pot, the roots plus adhering rhizosphere soil from three plants were excised, individually placed in 10 ml of sterile water, vortexed for 1 min and then sonicated in an ultrasonic bath (Bransonic 521; Branson, Shelton, CT) for 1 min. Each plant was processed separately. The root washings were serially diluted in 96-well microtiter plates prefilled with 200 μl of sterile water per well and then 50 μl was transferred into 96-well microtiter plates containing 1/3-strength KMB broth supplemented with rifampin, cycloheximide, ampicillin, and chloramphenicol. Bacterial growth was assessed after 72 h of incubation at room temperature with a microplate spectrophotometer (MR5000; Dynatech Laboratories, Burlington, MA) and an OD600 of ≥0.1 was scored as positive (McSpadden Gardener et al. 2001). In mixed inoculations the two different populations were identified by PCR with Ps_up1/Ps_low1 (Mavrodi et al. 2001) and viscB1/viscB2 primers (Yang et al. 2014) for detecting HC9–07rif and HC1–07rif, respectively. Population densities of total culturable aerobic heterotrophic bacteria were determined by performing the same assay in one-tenth-strength TS broth (1/10× TSB) supplemented with cycloheximide (McSpadden Gardener et al. 2001). The experiment was repeated three times with similar results.
Statistical analyses
Plant heights and rating of diseased roots were compared with STATISTIX (version 8, Analytical Software, St. Paul, MN). Differences among treatments were determined at each bacterial dilution by standard one-way analysis of variance (ANOVA), and mean comparisons among treatments were performed by using Fishers protected least significant difference (LSD) test at P = 0.05 or Kruskal Wallis test at P = 0.05. Bacterial population densities were log transformed before statistical analysis.
Results
Selection and characterization of recombinant strains
Recombinant colonies resulting from the mating of P. fluorescens HC1–07rif with E. coli S17–1(λpir)(pUTKm-phz) were selected on M9 agar containing kanamycin. Twenty colonies were initially isolated and the clone designated HC1–07PHZ was selected for further study. It remained kanamycin-resistant after 10 successive transfers on KMB agar and did not differ morphologically from the wild type strain. It also maintained the same growth rate as the wild type. The presence of the introduced phenazine biosynthesis genes was confirmed by PCR with the phenazine-specific primers PHZ1 and PHZ2 (Delaney et al. 2001). The HC1–07PHZ recombinant strain produced both CLP and PCA in vitro as confirmed by the drop collapse test (Yang et al. 2014) and by thin-layer chromatography (TLC), respectively (results not shown).
In vitro inhibition of fungal pathogens
In vitro inhibition analyses of G. graminis var. tritici isolates LD5, ARS-A1 and R3-111a-1 were conducted on KMB and PDA media. The recombinant strain HC1–07PHZ was more inhibitory to the three G. graminis var. tritici isolates on both media than was strain HC1–07, except that both strains inhibited isolate R3-111a-1 on PDA to the same extent (Table 2). Also strain HC1–07PHZ produced larger zones of inhibition on PDA medium against the three pathogens than did the natural PCA-producing strain HC9–07 (Table 2). In the control treatment with no bacteria, G. graminis var. tritici grew normally and reached the edge of the Petri plate (4.5 cm) after 7 days.
Suppression of take-all disease
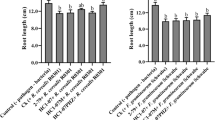
All strains and strain combinations (HC1–07rif, HC1–07PHZ, HC9–07rif and HC1–07rif + HC9–07) at all doses (102, 104, and 107 CFU/seed) suppressed take-all on wheat as compared to the non-treated control (Fig. 1). At a dose of 102 CFU per seed the recombinant strain HC1–07PHZ suppressed G. graminis var. tritici significantly better than did the wild type strain HC1–07rif, but at doses of 104 and 107 CFU/seed the wild-type and recombinant strains were not different. At a dose of 104 CFU/seed, the combination of strains HC1–07rif + HC9–07rif reduced take-all significantly better than the other strains applied individually. At a dose of 107 CFU/seed, strains HC1–07rif and HC1–07PHZ applied individually showed significantly better disease suppression than the treatments HC9–07rif and HC1–07rif + HC9–07. Biocontrol activity, as determined by measurements of shoot length, was in general agreement with those of disease ratings (Fig. 2). Overall, better disease suppression increased with the dose of the bacteria (Figs. 1 and 2).

Suppression of take-all by HC1–07rif, HC9–07rif, the combination HC1–07rif + HC9–07rif, and HC1–07PHZ. Strains were applied at a concentration of 102, 104, 107 CFU/seed in 1% methylcellulose. Wheat seeds of the Control treatment were not inoculated with bacteria (0 Bacterial dose). All bars at the 0 dose represent the same control. Disease rating was evaluated 3 weeks after planting. Bars with the same letters at the same dose are not significantly different by Kruskal-Wallis all-pairwise comparison test (P = 0.05)

Shoot length of wheat treated with HC1–07rif, HC9–07rif, the combination HC1–07rif + HC9–07rif, and HC1–07PHZ at a concentration of 102, 104, 107 CFU/seed in 1% methylcellulose and grown for 3 weeks. Wheat seeds of the Control treatment were not inoculated with bacteria (0 bacterial dose). All bars at the 0 dose represent the same Control. Bars with the same letters at the same dose are not significantly different according to Fisher’s protected least-significant-difference test (P = 0.05) or by Kruskal-Wallis all-pairwise comparison test (P = 0.05)
Bacterial population densities in the rhizosphere of wheat
Strains HC1–07rif, HC9–07rif, HC1–07PHZ, and the combination of HC1–07rif + HC9–07rif were applied to the soil at a concentration of 104 CFU/g soil (Table 3). Three weeks after planting, the population density of HC1–07rif reached log 7.4 CFU g−1 root, a level that was significantly higher than the populations of the other treatments, which ranged from log 6.6 to log 6.9 CFU g−1 root, but were not significantly different. When HC1–07rif and HC9–07rif were introduced together, the population densities of each strain on the roots did not differ significantly (Table 3). The population densities of the total culturable aerobic bacteria in the rhizosphere ranged from log 8.5 to log 8.9 CFU g−1 root but did not differ significantly among any of the treatments (Table 3).
Discussion
During the last four decades, thousands of biocontrol agents (BCAs) have been tested on hundreds of plant diseases. Although the use of biocontrol technology still remains a small fraction of that of chemical pesticides, the number of new BCAs, their performance, and acceptance by farmers continues to increase. However, there are several chronic problems that need to be overcome before the use of biocontrol technology can reach its full potential as an integral component of modern agriculture. The first problem is inconsistent performance. Why a BCA suppresses a disease in one field or year but not in the next is an important unanswered question. A second problem is the narrow spectrum of activity of most BCAs. An agent may be effective against a single disease, but often a disease complex must be controlled. Thirdly, BCAs usually operate over a narrower range of environmental conditions and thus are more sensitive to environmental extremes than chemicals (Weller and Thomashow 2015).
Genetic engineering offers an approach to enhance the consistency of performance, spectrum of activity and colonizing ability of BCAs. All mechanisms of biocontrol have been targeted for improvement (Weller and Thomashow 2015). Antibiotics produced by Pseudomonsas spp. are not only important mechanisms of biocontrol (Haas and Defago 2005; Weller et al. 2007), but their biosynthesis genes have been the ones most frequently used to generate recombinant agents (Weller and Thomashow 2015). In our previous study, Yang et al. (2011) collected hundreds of bacterial isolates from wheat grown in fields in the Chinese provinces of Hebei and Jiangsu. Wheat grown in these fields historically had been rotated with either corn or rice but nothing was known about the phytobiomes of the crops in these fields. The viscosin-like CLP producer HC1–07 and PCA-producer HC9–07 are representatives of the fluorescent pseudomonads isolated from these Chinese fields, and are effective biocontrol agents of take-all and Rhizoctonia root rot (Yang et al. 2011, 2014). CLP and PCA-deficient mutants, respectively, were significantly less suppressive against take-all than the wild-type strains (Yang et al. 2011, 2014). Some CLPs and phenazines can work in concert to promote biocontrol of certain diseases (Perneel et al. 2008; D'aes et al. 2011; Olorunleke et al. 2015). For example, Perneel et al. (2008) demonstrated that both phenazines and biosurfactants produced by P. aeruginosa PNA1 were important in the biological control of Pythium splendens on bean and Pythium myriotylum on cocoyam. When biosurfactant-deficient and phenazine-deficient mutants of PNA1 were applied individually, no disease-suppression occurred, indicating the synergetic interaction of the metabolites. They hypothesized that biosurfactants improve the physical contact between hyphae and phenazines, and thus increase entry of phenazines into the fungal cell.
We hypothesized that introducing the PCA biosynthesis locus into HC1–07rif would substantially improve the biocontrol activity of the recombinant strain. The operon phzABCDEFG from strain 2–79 was selected for insertion into HC1–07rif because strain HC9–07 and other Chinese PCA-producing strains clustered closely with strain 2–79 based upon both analysis of sequences within phzF and comparisons of partial 16S rDNA sequences (Yang et al. 2011). PCA produced by 2–79 is not only the mechanism of suppression of take-all but also contributes to the ecological competence of the strain in the wheat rhizosphere (Mazzola et al. 1992; Weller et al. 2007). The disarmed Tn5 vector (pUT::Ptac-phzABCDEFG), originally constructed by Thomashow and colleagues, has been used extensively to stably introduce a single copy of the PCA biosynthesis genes under the control of a P tac promotor into Pseudomonas spp. from sources worldwide to improve biocontrol activity (Weller and Thomashow 2015). For example, the phz operon was introduced into P. brassicacearum (formerly P. fluorescens) Q8r1–96 (Loper et al. 2012), a strain that naturally produces the antibiotic DAPG and suppresses take-all. Several recombinants of Q8r1–96 were selected (Z30–97, Z32–97, Z33–97 and Z34–97) and all produced both DAPG and PCA. Surprisingly, these transgenic strains were no more suppressive of take-all and Pythium root rot on wheat than the wild-type strain Q8r1–96; however they showed remarkable suppression of Rhizoctonia root rot at a dose of only 100 CFU seed−1, which was 100 to 1000 times less than the dose required for similar disease control by the wild-type Q8r1–96 (Huang et al. 2004). In a similar study, P. fluorescens SBW25 was transformed with the same phz gene construction and the transgenic strains gained enhanced ability to suppress Pythium ultimum damping-off disease of pea when compared to the wild-type strains SBW25 (Timms-Wilson et al. 2000).
In our current study, strain HC1–07PHZ produced both CLP and PCA, and was more inhibitory of G. graminis in vitro than was HC1–07; however, the recombinant colonized the roots slightly less than did the wild type, yet not enough to impact biocontrol activity. Strain HC1–07PHZ also showed greater biocontrol activity against take-all than did the wild type based on both root disease ratings and plant height measurements, but only at a dose of 102 CFU per seed. Our results differ from those of Huang et al. (2004), who saw no improvement in the biocontrol activity against take-all by PCA-producing recombinants of Q8r1–96 transformed with the same construction that we used. Unexpectedly, however, we also found that at doses of 104 or 107 CFU per seed the wild-type strain and the recombinant HC1–07PHZ did not differ. This phenomenon may be explained by the observation that antibiotics produced in natural environments can have multiple roles depending on the amount that accumulates in a niche (Davies et al. 2006; Mavrodi et al. 2012). For example, many Pseudomonas antibiotics that inhibit pathogens also can be phytotoxic (Brazelton et al. 2008; Kwak et al. 2012) if the dose is too high (Bull et al. 1991; M. Yang and D. M. Weller, unpublished data). Thus, antibiotics must be produced by an agent in sufficient quantities to suppress the pathogen, but too much production can damage the roots and negate the beneficial biocontrol effects. For example, the phenazine pyocyanin produced by P. aeruginosa 7NSK2 actually increased the severity of sheath blight on rice caused by R. solani, even though the pathogen is highly sensitive to the antibiotic (De Vleesschauwer et al. 2006). An alternative explanation for the lack of improvement in HC1–07PHZ at the higher doses may simply be that the additional amount of CLP produced by higher populations provided all of the control possible at that time and PCA would have added little to the suppression. This was an unexpected finding and thus is a question that needs to be studied in greater depth.
It was equally surprising to find that the effectiveness of each strain used individually (HC1–07rif, HC9–07rif or HC1-09PHZ) or in combination (HC1–07rif + HC9–07rif) depended on the bacterial dose applied to the seed. Indeed at a dose of 104 the combination treatment provided the best biocontrol of take-all, but at a dose of 107, HC1–07rif and HC1-07PHZ were the best treatments. Combinations of Pseudomonas strains have been tested to improve biocontrol activity against a wide range of pathogens (including G. graminis var. tritici) with mixed success (Pierson and Weller 1994; Weller 2007). For example, Weller and Cook (1983) showed that the combination of P. synxantha 2–79 and P. fluorescens13–79 increased wheat yields and reduced take-all in field plots inoculated with G. graminis var. tritici as compared to the same strains applied individually. However, Guetsky et al. (2002) showed that when two biocontrol agents were applied together in a detached strawberry leaf test, biocontrol efficacy was reduced. Xu et al. (2011) also found that combination of two agents, each with a single main mechanism of action, did not result in appreciable improvement of biocontrol over applications of individual BCAs.
In summary, for the first time to our knowledge, we determined the impact of combining PCA and a viscosin-like CLP as mechanisms in the biocontrol of take-all. Whether the metabolites were produced in a single recombinant strain (HC1–07PHZ) or in two separate strains used in combination (HC1–07rif + HC9–07rif), the combination of metabolites provided a significant improvement in biocontrol activity. Thus, further strain mixtures or recombinants containing these two metabolites should be tested against a wider range of pathogens and diseases. Finally, our results support a growing body of literature that a major benefit of recombinant strains is a reduction in the dose of a BCA needed to provide disease suppression, which would significantly reduce the cost of production of an agent (Weller and Thomashow 2015).
References
Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A., & Struhl, K. (2002). Short protocols in molecular biology (5th ed.). New York, NY: John Wiley & Sons.
Brazelton, J. N., Pfeufer, E. E., Sweat, T. A., Gardener, B. B., & Coenen, C. (2008). 2,4-diacetylphloroglucinol alters plant root development. Molecular Plant-Microbe Interactions, 21, 1349–1358.
Bull, C. T., Weller, D. M., & Thomashow, L. S. (1991). Relationship between root colonization and suppression of Gaeumannomyces graminis var. tritici by Pseudomonas fluorescens strain 2-79. Phytopathology, 81, 954–959.
Chin-A-Woeng, T. F. C., Bloemberg, G. V., & Lugtenberg, B. J. J. (2003). Phenazines and their role in biocontrol by Pseudomonas bacteria. New Phytologist, 157, 503–523.
Cook, R. J. (2003). Take-all of wheat. Physiological and Molecular Plant Pathology, 62, 73–86.
D’aes, J., Maeyer, K. D., Pauwelyn, E., & Höfte, M. (2010). Biosurfactants in plant Pseudomonas interactions and their importance to biocontrol. Environmental Microbiology Reports, 2, 539–572.
D'aes, J., Hua, G. K., De Maeyer, K., Pannecoucque, J., Forrez, I., Ongena, M., Dietrich, L. E., Thomashow, L. S., Mavrodi, D. V., & Höfte, M. (2011). Biological control of Rhizoctonia root rot on bean by phenazine and cyclic lipopeptide producing Pseudomonas CMR12a. Phytopathology, 101, 996–1004.
Davies, J., Spiegelman, G. B., & Yim, G. (2006). The world of subinhibitory antibiotic concentrations. Current Opinion in Microbiology, 9, 445–453.
De Vleesschauwer, D., Cornelis, P., & Höfte, M. (2006). Redox-active pyocyanin secreted by Pseudomonas aeruginosa 7NSK2 triggers systemic resistance to Magnaporthe grisea but enhances Rhizoctonia solani susceptibility in rice. Molecular Plant-Microbe Interactions, 19, 1406–1409.
Delaney, S. M., Mavrodi, D. V., Bonsall, R. F., & Thomashow, L. S. (2001). phzO, a gene for biosynthesis of 2-hydroxylated phenazine compounds in Pseudomonas aureofaciens 30-84. Journal of Bacteriology, 183, 318–327.
Freeman, J., & Ward, E. (2004). Gaeumannomyces graminis, the take-all fungus and its relatives. Molecular Plant Pathology, 5, 235–252.
Guetsky, R., Shtienberg, D., Elad, Y., Fischer, E., & Dinoor, A. (2002). Improving biological control by combining biocontrol agents each with several mechanisms of disease suppression. Phytopathology, 92, 976–985.
Haas, D., & Defago, G. (2005). Biological control of soilborne pathogens by fluorescent pseudomonads. Nature Reviews Microbiology, 3, 307–319.
Huang, Z. Y., Bonsall, R. F., Mavrodi, D. V., Weller, D. M., & Thomashow, L. S. (2004). Transformation of Pseudomonas fluorescens with genes for biosynthesis of phenazine-1-carboxylic acid improves biocontrol of rhizoctonia root rot and in situ antibiotic production. FEMS Microbiology Ecology, 49, 243–251.
King, E. O., Ward, M. K., & Raney, D. E. (1954). Two simple media for the demonstration of pyocyanin and fluorescein. Journal of Laboratory and Clinical Medicine, 44, 302–307.
Kwak, Y. S., Bakker, P. A. H. M., Glandorf, D. C. M., Rice, J. T., Paulitz, T. C., & Weller, D. M. (2009). Diversity, virulence and 2,4-diacetylphloroglucinol sensitivity of Gaeumannomyces graminis var. tritici isolates from Washington state. Phytopathology, 99, 472–479.
Kwak, Y. S., Bonsall, R. F., Okubara, P. A., Paulitz, T. C., Thomashow, L. S., & Weller, D. M. (2012). Factors impacting the activity of 2,4-diacetylphloroglucinol-producing Pseudomonas fluorescens against take-all of wheat. Soil Biology and Biochemistry, 54, 48–56.
Landa, B. B., Mavrodi, O. V., Raaijmakers, J. M., McSpadden Gardener, B. B., Thomashow, L. S., & Weller, D. M. (2002). Differential ability of genotypes of 2,4-diacetylphloroglucinol-producing Pseudomonas fluorescens strains to colonize the roots of pea plants. Applied and Environmental Microbiology, 68, 3226–3237.
Le, C. N., Kruijt, M., & Raaijmakers, J. M. (2012). Involvement of phenazines and lipopeptides in interactions between Pseudomonas species and Sclerotium rolfsii, causal agent of stem rot disease on groundnut. Journal of Applied Microbiology, 112, 390–403.
Loper, J. E., Hassan, K. A., Mavrodi, D. V., Davis, E. W., Lim, C. K., Shaffer, B. T., et al. (2012). Comparative genomics of plant-associated Pseudomonas spp: Insights into diversity and inheritance of traits involved in multitrophic interactions. PLoS Genetics, 8(7), e1002784.
Mavrodi, D. V., Ksenzenko, V. N., Bonsall, R. F., Cook, R. J., Boronin, A. M., & Thomashow, L. S. (1998). A seven-gene locus for synthesis is of phenazine-1-carboxylic acid by Pseudomonas fluorescens 2-79. Journal of Bacteriology, 180, 2541–2548.
Mavrodi, D. V., Bonsall, R. F., Delaney, S. M., Soule, M. J., Phillips, G., & Thomashow, L. S. (2001). Functional analysis of genes for biosynthesis of pyocyanin and phenazine-1-carboxamide from Pseudomonas aeruginosa PAO1. Journal of Bacteriology, 183, 6454–6465.
Mavrodi, O. V., Mavrodi, D. V., Park, A. A., Weller, D. M., & Thomashow, L. S. (2006). The role of dsbA in colonization of the wheat rhizosphere by Pseudomonas fluorescens Q8r1-96. Microbiology, 152, 863–872.
Mavrodi, D. V., Peever, T. L., Mavrodi, O. V., Parejko, J. A., Raaijmakers, J. M., Lemanceau, P., Mazurier, S., Heide, L., Blankenfeldt, W., Weller, D. M., & Thomashow, L. S. (2010). Diversity and evolution of the phenazine biosynthesis pathway. Applied and Environmental Microbiology, 76, 866–879.
Mavrodi, D. V., Mavrodi, O. V., Parejko, J. A., Bonsall, R. F., Kwak, Y.-S., Paulitz, T. C., Thomashow, L. S., & Weller, D. M. (2012). Accumulation of the antibiotic phenazine-1-carboxylic acid in the rhizosphere of dryland cereals. Applied and Environmental Microbiology, 78, 804–812.
Mavrodi, D. V., Parejko, J. A., Mavrodi, O. V., Kwak, Y. S., Weller, D. M., Blankenfeldt, W., & Thomashow, L. S. (2013). Recent insights into the diversity, frequency and ecological roles of phenazines in fluorescent Pseudomonas spp. Environmental Microbiology, 15, 675–686.
Mazurier, S., Corberand, T., Lemanceau, P., & Raaijmakers, J. M. (2009). Phenazine antibiotics produced by fluorescent pseudomonads contribute to natural soil suppressiveness to Fusarium wilt. ISME Journal, 3, 977–991.
Mazzola, M., Cook, R. J., Thomashow, L. S., Weller, D. M., & Pierson, L. S. (1992). Contribution of phenazine antibiotic biosynthesis to the ecological competence of fluorescent pseudomonads in soil habitats. Applied and Environmental Microbiology, 58, 2616–2624.
McSpadden Gardener, B. B., Mavrodi, D. V., Thomashow, L. S., & Weller, D. M. (2001). A rapid polymerase chain reaction-based assay characterizing rhizosphere populations of 2,4-diacetylphloroglucinol producing bacteria. Phytopathology, 91, 44–54.
Nam, H. S., Yang, H. J., Oh, B. J., Anderson, A. J., & Kim, Y. C. (2016). Biological control potential of Bacillus amyloliquefaciens KB3 isolated from the feces of Allomyrina dichotoma larvae. Plant Pathology Journal, 32(3), 273–280.
Olorunleke, F. E., Hua, G. K. H., Kieu, N. P., Ma, Z., & Höfte, M. (2015). Interplay between orfamides, sessilins and phenazines in the control of Rhizoctonia diseases by Pseudomonas sp. CMR12a. Environmental Microbiology Reports, 7, 774–781.
Parejko, J. A., Mavrodi, D. V., Mavrodi, O. V., Weller, D. M., & Thomashow, L. S. (2012). Population structure and diversity of phenazine-1-carboxylic acid producing fluorescent Pseudomonas spp. from dryland cereal fields of central Washington state (USA). Microbial Ecology, 64, 226–241.
Parejko, J. A., Mavrodi, D. V., Mavrodi, O. V., Weller, D. M., & Thomashow, L. S. (2013). Taxonomy and distribution of phenazine-producing Pseudomonas spp. in the dryland agroecosystem of the inland Pacific Northwest, United States. Applied and Environmental Microbiology, 79(12), 3887–3891.
Perneel, M., D’hondt, L., De Maeyer, K., Adiobo, A., Rabaey, K., & Höfte, M. (2008). Phenazines and biosurfactants interact in the biological control of soil-borne diseases caused by Pythium spp. Environmental Microbiology, 10, 778–788.
Pierson, E. A., & Weller, D. M. (1994). Use of mixtures of fluorescent pseudomonads to suppress take-all and improve the growth of wheat. Phytopathology, 84, 940–947.
Pieterse, C. M. J., Zamioudis, C., Berendsen, R. L., Weller, D. M., Van Wees, S. C. M., & Bakker, P. H. A. M. (2014). Induced systemic resistance by beneficial microbes. Annual Review of Phytopathology, 52, 347–375.
Raaijmakers, J. M., Weller, D. M., & Thomashow, L. S. (1997). Frequency of antibiotic-producing Pseudomonas spp. in natural environments. Applied and Environmental Microbiology, 63, 881–887.
Raaijmakers, J. M., Vlami, M., & de Souza, J. T. (2002). Antibiotic production by bacterial biocontrol agents. Antonie Van Leeuwenhoek, l81, 537-547.
Raaijmakers, J. M., Paulitz, T., Steinberg, C., Alabouvette, C., & Moe¨nne-Loccoz, Y. (2009). The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant and Soil, 321, 341–361.
Raaijmakers, J. M., De Bruijn, I., Nybroe, O., & Ongena, M. (2010). Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiology Reviews, 34, 1037–1062.
Ramsey, N. E. (2001). Occurrence of take-all on wheat in Pacific Northwest cropping systems, M.S. thesis, department of plant pathology. Pullman: Washington State University.
Tambong, J. T., & Höfte, M. (2001). Phenazines are involved in biocontrol of Pythium myriotylum on cocoyam by Pseudomonas aeruginosa PNA1. European Journal of Plant Pathology, 107, 511–521.
Thomashow, L. S., & Weller, D. M. (1988). Role of a phenazine antibiotic from Pseudomonas fluorescens in biological control of Gaeumannomyces graminis var. tritici. Journal of Bacteriology, 170, 3499–3508.
Timms-Wilson, T. M., Ellis, R. J., Renwick, A., Rhodes, D. J., Mavrodi, D. V., Weller, D. M., Thomashow, L. S., & Bailey, M. J. (2000). Chromosomal insertion of phenazine-1-carboxylic acid biosynthetic pathway enhances efficacy of damping-off disease control by Pseudomonas fluorescens. Molecular Plant-Microbe Interactions, 13, 1293–1300.
Tran, H., Ficke, A., Asiimwe, T., Höfte, M., & Raaijmakers, J. M. (2007). Role of the cyclic lipopeptide massetolide a in biological control of Phytophthora infestans and in colonization of tomato plants by Pseudomonas fluorescens. New Phytologist, 175, 731–742.
Wang, X. F., Mavrodi, D. V., Ke, L. F., Mavrodi, O. V., Yang, M. M., Thomashow, L. S., Zheng, N., Weller, D. M., & Zhang, J. B. (2014). Biocontrol and growth-promoting activity of rhizobacteria from Chinese fields with contaminated soils. Microbial Biotechnology, 1(1), 1–15.
Weller, D. M. (2007). Pseudomonas biocontrol agents of soilborne pathogens: Looking back over 30 years. Phytopathology, 97, 250–256.
Weller, D. M. (2015). Take-all decline and beneficial pseudomonads. (pp. 363–370). In B. Lugtenberg (Ed.), Principles of plant-microbe interactions-microbes for sustainable agriculture. Switzerland: Springer.
Weller, D. M., & Cook, R. J. (1983). Suppression of take-all of wheat by seed treatments with fluorescent pseudomonads. Phytopathology, 73, 463–469.
Weller, D. M., & Thomashow, L. S. (2015). Phytosanitation and the development of transgenic biocontrol agents. Pp 35-43, in: OECD biosafety and the environmental uses of micro-organisms: Conference proceedings, OECD publishing, Paris. 220 p.
Weller, D. M., Raaijmakers, J. M., McSpadden Gardener, B. B., & Thomashow, L. S. (2002). Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annual Review of Phytopathology, 40, 309–348.
Weller, D. M., Landa, B. B., Mavrodi, O. V., Schroeder, K. L., De La Fuente, L., Blouin Bankhead, S., Allende Molar, R., Bonsall, R. F., Mavrodi, D. V., & Thomashow, L. S. (2007). Role of 2,4-diacetylphloroglucinol-producing fluorescent Pseudomonas spp. in the defense of plant roots. Plant Biology, 1, 4–20.
Xu, X. M., Jeffries, P., Pautasso, M., & Jeger, M. J. (2011). Combined use of biocontrol agents to manage plant diseases in theory and practice: A review. Phytopathology, 101, 1024–1031.
Yang, M. M. (2011). Biological control and mechanisms of suppression of take-all by fluorescent Pseudomonas spp. Ph.D. Thesis. Nanjing Agricultural University, Nanjing, China.
Yang, M. M., Mavrodi, D. V., Mavrodi, O. V., Bonsall, R. F., Parejko, J. A., Paulitz, T. C., Thomashow, L. S., Yang, H. T., Weller, D. M., & Guo, J. H. (2011). Biological control of take-all by fluorescent Pseudomonas spp. from Chinese wheat fields. Phytopathology, 101, 1481–1491.
Yang, M. M., Wen, S. S., Mavrodi, D. V., Mavrodi, O. V., Thomashow, L. S., Guo, J. H., & Weller, D. V. (2014). Biological control of wheat root diseases by the CLP-producing strain Pseudomonas fluorescens HC1-07. Phytopathology, 104, 248–256.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Yang, M., Mavrodi, D.V., Mavrodi, O.V. et al. Construction of a recombinant strain of Pseudomonas fluorescens producing both phenazine-1-carboxylic acid and cyclic lipopeptide for the biocontrol of take-all disease of wheat. Eur J Plant Pathol 149, 683–694 (2017). https://doi.org/10.1007/s10658-017-1217-6
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10658-017-1217-6