
Abstract
Widespread soil contamination with oil and the toxicity of petroleum hydrocarbons to soil biota make it extremely important to study microbial responses to oil stress. Soil metabolites reflect the main metabolic pathways in the soil microbial community. The examination of changes in the soil metabolic profile and metabolic function is essential for a better understanding of the nature of the pollution and restoration of the disturbed soils. The present study aimed to assess the long-term effect of oil on the ecological state of the soil, evaluate quantitative and qualitative differences in metabolite composition between soil contaminated with oil and non-contaminated soil, and reveal biologically active metabolites that are related to oil contamination and can be used for contamination assessment. A long-term field experiment was conducted to examine the effects of various oil concentrations on the biochemical properties and metabolic profile of the soil. Podzolic soil contaminated with oil demonstrated the long-term inhibition of soil biological activity and vegetation. Oil affected the metabolic activity of soil fungi increasing the production of toxic metabolites. A metabolomic approach was employed to determine soil metabolites. The metabolite profile was found to vary greatly between oil-contaminated and non-contaminated soils. Carbohydrates had the largest number of metabolites negatively affected by oil, while the content of organic acids, phenolic compounds, and terpenoids was mainly increased in oil-contaminated soil. The evaluation of the long-term impact of oil on microbial metabolism can make a valuable contribution to the assessment of soil quality and the activity of soil microorganisms being under stress from oil pollution. The results contribute to a further understanding of the role of microorganisms in the ecological functions of contaminated soil, which can be useful in the development of rehabilitation strategies for disturbed sites.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Soil contamination with petroleum and its derivative products is a global environmental problem. Petroleum oil enters the environment through different routes including leakages from wellheads, pipelines, and underground storage tanks, accidental spills during transportation, incorrect disposal of petroleum wastes, drilling operations, and many other ways. Increasing the contamination of the natural environment leads to loss of vegetation cover, a decrease in soil fertility, and progressive soil degradation (Albert & Tanee, 2011; Alrumman et al., 2015; Souza et al., 2014; Wyszkowska et al., 2015). To solve the problem of rehabilitation of oil-contaminated soils, it is necessary to determine the critical values of stress factors that can lead to irreversible violations of fundamental biological processes in soil ecosystems (Bakina et al., 2021; Geng et al., 2022; Onwosi et al., 2020). No less important is the qualitative and quantitative assessment of the relationships between the components of the biogeocenoses of disturbed soils. The implementation of these approaches is impossible without understanding the mechanisms of self-purification of oil-contaminated soils and a timely comprehensive assessment of the ecological state of the soil as the main component of biogeocenosis.
Ecological assessment is necessary not only to determine the current suitability of soils for use but also to identify potentially dangerous functional disorders in soil ecosystems that may manifest themselves in the future (Hernandez-Soriano & Jimenez-Lopez, 2014). Soil contamination could cause serious disturbance of its natural metabolic profile and metabolic function. Thus, careful examination of these violations is essential for a better understanding of the nature of environmental pollution (Patil et al., 2016).
One effective way to assess changes in soil function and quality is metabolomic analysis, as soil metabolites reflect important metabolic pathways and complex molecular networks in the soil microbial community (Withers et al., 2020). Soil metabolites have been actively studied in recent years to investigate the effects of environmental stresses on soil biota (Griffin, 2014; Li et al., 2019; Rochfort et al., 2015). Functional shifts at the molecular level provide deep insight into biological responses to stress conditions and can be even more sensitive than traditional indicators (Pagano et al., 2016).
In the functioning of biogeocenosis, a key role is played by soil microorganisms with a genetic potential to degrade anthropogenic pollutants that decompose and mineralize organic substances. Pollutants affect several of the major metabolic pathways in soil microorganisms, including the tricarboxylic acids cycle, glycolysis, and amino acids metabolism, causing a decrease in the production of the associated metabolites (Hernandez-Soriano & Jimenez-Lopez, 2014). Metabolic response to soil contamination can be considered as part of the biological reaction of soil microorganisms to the influence of pollutants (Boer et al., 2013; Tremaroli et al., 2009). Metabolites related to stress response comprise an appropriate group of target compounds for risk assessment. However, there is no work or data reporting the metabolic profile of oil-contaminated soil and the degree to which metabolomic fingerprints of contaminated soil may differ. There is a lack of information about the overall shifts in soil microbial metabolic function, and changing trends in biologically active soil metabolites in response to long-term contamination with oil.
Therefore, in this study, it is hypothesized that (1) long-term soil contamination with oil will affect soil microbial metabolic function depending on the level of contamination, and (2) the combination of metabolomics with classical culture-based microbiological and biochemical methods will provide new insight into the ecological state of the soil. Based on this, the specific objectives of this study were to assess the long-term effect of oil on the ecological state of the soil, to evaluate quantitative and qualitative differences in metabolite composition between soil contaminated with oil and non-contaminated soil, and to reveal biologically active metabolites that are related to oil contamination and can be used for assessment of land recovery following oil drilling, transportation, and refinery activities.
Materials and methods
Soil treatment and sampling
The field experiment was performed for 6 years (2010–2016) at an experimental site near St. Petersburg, Russia (58°49′53″N, 30°7′56″E) (Fig. 1). This area has a humid continental climate with average annual temperature, precipitation, and air humidity of 5.8 °C, 660 mm, and 78%, respectively. Soil moisture in the area is mostly high, since evapotranspiration is low due to the cool climate. The studied soil is sandy podzol (Albic Podzol according to WRB IUSS Working group WRB, 2006) covered with moss and lichen. The soil contains 96.5% sand, 3.1% clay, and 0.4% silt (pHH2O of 4.05). The vegetation on the study plots was allowed to develop naturally throughout the entire research period. Five square plots, measuring 20 m2 each, were established separately in the flattened part of the study areas. The first plot served as an uncontaminated control, and four were treated with crude oil (0.6, 1.2, 3.5, and 8.5 L m−2). The vegetation cover (plants, sod, and litter) was removed from treatment plots prior to the application of oil. The peaty moss-lichen cover was removed from the soil surface to the mineral sandy horizon.

Location of the experimental plots
Crude oil (saturated hydrocarbons—46.9%, aromatics—22.4%, polyaromatic hydrocarbons—9.8%, and asphaltenes—2.1%) was collected from oil refinery “Kirishinefteorgsintez” (Leningrad Region, Russia). The oil was poured evenly across the experimental plots using 0.6, 1.2, 3.5, and 8.5 L per m−2 of soil. After pouring, the upper layer of 0–10 cm was additionally mixed with a rake to achieve a uniform concentration of oil over the entire depth of the polluted layer. Thereafter, vertical migration and distribution of oil and oil products were governed by natural causes (wetting and gravity).
Soil samples were collected one week and three months after contamination, and later on annually in late September–early October (i.e., at the end of the growing season). Each plot was divided into four subplots of 2.5 m2 with a method of randomized repetitions (Dospehov, 1985), and 10 coring sites were chosen within the subplot (One plot ➔ Four subplots ➔ 10 coring sites). Soil samples were taken with a soil corer from a depth of 0–10 cm. All ten soil cores, collected from each subplot, were pooled and sieved (< 2 mm). This combined sample represented one replicate so there were four replications from each plot. The total weight of a combined soil sample was 900 g.
Chemical and physical analyses
The chemical and physical characteristics of the collected samples (pH, sum of exchangeable bases, hydrolytic acidity, base saturation, total organic carbon, N-NH4, available phosphorus, and potassium) were determined according to Arinushkina (1970). Total petroleum hydrocarbons (TPH) were extracted from soil samples with hexane and measured according to protocol PND F 16.1:2.21-98 (2012). TPH levels were determined by UV fluorescence spectroscopy using a FLUORAT-02-3 M spectrofluorometer (Nordinkraft-Sensor, Russia). The fluorescence of the samples was measured at 270 nm excitation and 320 nm emission wavelengths.
Assessment of soil toxicity
The ecotoxicity of contaminated soils was determined by direct contact (solid-phase) bioassay. Solid-phase bioassays were based on the germination and growth of the common wheat Triticum aestivum L. (Kapel’kina et al., 2009). Soil samples were put into 100-mm-diameter Petri dishes with the moisture adjusted to 60% of the maximum water-holding capacity. Twenty seeds with > 90% germination rate were arranged on the soil surface. The test dishes were incubated at 20 °C for 72 h. The measurement endpoints used were seed germination and seedling growth (roots and shoots).
To identify changes in vegetation, the research team measured foliage projective cover for each plot, which is the relative area covered by all the plants in a plot as seen above (Mirkin et al., 2002). The measurements were done annually at the end of the growing season.
Assessment of soil enzyme activities
Four types of soil enzymatic activity (dehydrogenase, catalase, phosphatase, and urease) were determined using the methods described by Haziev (2005) with few modifications. Dehydrogenase activity was analyzed via the reduction of the 2,3,5-triphenyltetrazolium chloride (colorless) to a red-colored 2,3,5-triphenyl formazan (TPF) with a 1% (w/v) glucose solution added as a source of organic carbon. After an incubation period of 24 h at 30 °C, TPF was extracted with methanol, and the optical density of the solution was measured at 484 nm (spectrophotometer Genesys 10 UV, Thermo Spectronic, USA). The latter values served to determine the TPF concentration. Dehydrogenase activity was expressed as mg TPF g−1 day−1.
The activity of catalase was determined following the Johnson and Temple method (1964) by back-titrating residual H2O2 with KMnO4. The mixture of 2 g of dry soil and 0.3% hydrogen peroxide solution was shaken at 120 rpm for 20 min and then 1.5 M H2SO4 was added. Afterward, the solution was filtered, and the liquid was titrated using 0.1 M KMnO4. The result was expressed as ml KMnO4 g−1.
Phosphatase activity was determined with the Galstyan method (Haziev, 2005). The method is based on the colorimetric assaying of the released substrate after the incubation of 1 g of dry soil with a 0.5% p-nitrophenylphosphate solution and a modified universal buffer for 30 min at 30 °C. After incubation, 1 N NaOH was added to the samples followed by filtration. The released p-nitrophenol was determined at 440 nm using a spectrophotometer Genesys 10 UV (Thermo Spectronic, USA). Phosphatase activity was expressed as mg P2O5 g−1 30 min−1.
Urease activity was measured using urea as a substrate (Haziev, 2005). Five grams of dry soil and 1 ml of methylbenzene were mixed in a 150-ml conical flask. After 15 min, 10 ml of 10% urea and 20 ml of citrate buffer (pH 6.7) were added to the sample. The mixture was incubated at 37 °C for 24 h and filtered. One milliliter of the filtrate was transferred into a volumetric flask, mixed with 4 ml of sodium phenoxide, and 3 ml of sodium hypochlorite, and diluted to 50 ml. Absorbance was determined at 578 nm using a spectrophotometer Genesys 10 UV (Thermo Spectronic, USA). The urease activity was expressed as μg NH3-N g−1 h−1 at 37 °C.
Enumeration of soil fungi and assessment of fungal allelopathic activity
Soil heterotrophic fungi were enumerated by the dilution-plate method (Zvyagintsev, 1991) using Czapek agar medium supplemented with streptomycin. The inoculated Petri plates were incubated at 24° for 5–7 days. After incubation, the number of filamentous fungi was counted. The results are given as CFU g−1 (colony forming units of the isolates per gram dry weight of soil).
The allelopathic activity of soil fungi was determined as the total phytotoxicity of fungal metabolites using the microbiological method as previously described (Polyak et al., 2020). Briefly, soil suspension was spread on the surface of Czapek agar medium, supplemented with streptomycin, and fungi were cultured in four replicates, at a constant temperature of 24 °C for 14 days. After incubation, the agar gel with grown fungi was turned over and seeds of plants were distributed on the back side of the agar gel (20 seeds per Petri dish). Common wheat (Triticum aestivum L.) was used for testing. Seed germination and root and shoot length of each seedling were determined three days after seeds were placed on the agar plate.
The allelopathic indices of the soil fungal community were calculated according to the following formulas:
where Ka1 and Ka2 are allelopathic indices calculated by measuring the inhibition of the length of roots and shoots, respectively;
lexp1 and lexp2—the length of roots and shoots, respectively, in experimental plates (Czapek agar gel with grown fungi);
lcontr1 and lcontr2—the length of roots and shoots, respectively, in control plates (sterile Czapek agar gel).
The permissible limit of phytotoxicity calculated using these formulas was 0.3 (Danilova, 2010).
Soil metabolite profiling analysis
The soil metabolite extraction method was modified from Swenson and Northen (2019). Briefly, 2 g soil samples were extracted twice with 8 ml of methanol: H2O = 4:1 (v/v). The soil mixture was sonicated for 10 min followed by centrifugation and the two extracts were combined. Two milliliters of the extract was flash‐frozen in liquid nitrogen, freeze‐dried for 48 h until complete dryness, and stored at − 80 °C prior to gas chromatography–mass spectrometry (GC–MS) analysis.
Metabolites were analyzed using an Agilent 6850 gas chromatograph equipped with a 5975 mass selective detector (Agilent Technologies, USA): column HP-5MS, 30 m × 0.25 mm × 0.25 μm. Chromatography was performed with linear temperature programming, from 70 to 320 °C for 6 °C/min, in the mode of constant carrier gas flow through the column (1 mL/min). The carrier gas was helium. Mass spectra were scanned in the range of 50–800 m/z with a frequency of 1.6 scans/s. Chromatograms of the samples were recorded using the total ion current. The metabolome data have been normalized into relative abundance obtained by dividing the normalized abundance by the sample-level total normalized abundance. The calculation of relative concentration on the dry weight of the sample and concentration of internal standard tricosane (1 μg μl−1) was performed by methods of semi-quantitative analysis. This approach meets minimum reporting standards related to the chemical analysis aspects of metabolomics experiments (including sample preparation, experimental analysis, quality control, metabolite identification, and data preprocessing) of the Chemical Analysis Working Group (CAWG) as part of the Metabolomics Standards Initiative (MSI) (Sumner et al., 2007).
GC–MS data analysis was performed using the PARADISe software (Johnsen et al., 2017) in association with NIST MS Search (National Institute of Standards and Technology (NIST, USA). For additional metabolite identification, the AMDIS (Automated Mass Spectral Deconvolution and Identification System, NIST, USA) was used with mass-spectra libraries: NIST2010, Golm Metabolome Database (GMD) and in-house library of the Resource Centre "The Development of Molecular and Cell Technologies" of the St. Petersburg University. Mass spectra were considered reliably identified if they coincided with the library record with a match factor of at least 80 (800 for the NIST search).
Statistical analyses
Pairwise differences were determined using Tukey's post-hoc multiple comparison test. Differences were considered significant at p < 0.05. Data were normalized and mean-centered prior to principal component analysis (PCA). Statistical analyses were performed using the Statistica software (version 10; Statsoft).
Results
General soil properties
The studied sandy podzolic soil was characterized by a low content of exchangeable bases, a low degree of base saturation, and high acidity (Table 1). The soil was depleted of nutrients and organic matter. Soil contamination in all tested doses did not affect the acid–base properties of the soil. There were no significant differences between unpolluted and polluted soils in any of the controlled parameters (pH, hydrolytic acidity, the sum of exchangeable bases, and the base saturation percentage) for the entire observation period.
The content of available phosphorus (AP) and potassium (AK) was less than 10 mg kg−1 of soil; total organic carbon (TOC) was only 1%. Soil physicochemical characteristics remained unchanged during the research period except for a decrease in the already low concentration of AP and AK in the first two years after soil contamination. After soil contamination with oil at doses of 3.5 l m−2 and above, there was a significant decrease in the content of AP (by 70–75% of the control value) and potassium (by almost 40%). By the end of the first growing season, AP and especially AK were gradually restored, although there was a significant difference compared to unpolluted and slightly polluted soils. The content of AP at maximum oil concentration was 50% compared to the control, AK—75%. By the end of the second growing season, the content of AP and AK was fully restored to the level of the unpolluted soil.
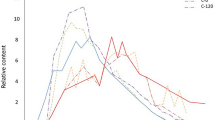
The content of organic carbon in oil-contaminated soils differed from the control uncontaminated soils, and this difference remained almost unchanged over the entire observation period. The rate of oil degradation in contaminated soil was low. The concentration of TPH remained nearly unchanged after two years of contamination (Fig. 2). After the third year, some decontamination occurred but even over a period of 6 years, the initial oil contamination of 0.48–3.75% TPH was reduced to 0.19–1.31%, which corresponded to a decontamination of 40–65%. The highest degradation rate (18% per year) was observed three years after contamination in soil contaminated with 8,5 l of oil per m2.

Degradation of total petroleum hydrocarbons (TPH) in soil contaminated with oil (0.6, 1.2, 3.5, and 8.5 l m−2)
Effect of petroleum hydrocarbons on enzymatic activities of soil
Oil pollution influences the biological activity of soil. Soil enzyme activity might be an informative indicator of the intensity of microbiological processes in contaminated soil (Lee et al., 2020). In contaminated soil, the values of soil enzymatic activities differed from the uncontaminated control (Fig. 3). Soil phosphatase activity showed mean values of 1.5–1.8 times higher compared to control after treatment with 1.2–8.5 l of oil per m2. However, the increased level of phosphatase activity was observed only for two years after the application of oil. Later on, the activity of phosphatase decreased and remained two times lower than phosphatase activity in the control soil even six years after contamination.

Effect of petroleum hydrocarbons on soil enzymatic activities in soil contaminated with oil (0.6, 1.2, 3.5, and 8.5 l m−2)
Urease showed enhanced activity during the first year of the experiment followed by a pronounced decrease compared to unpolluted soil. Plots treated with 3.5 and 8.5 l m−2 of oil demonstrated an increase of 262–284% compared to the control soil. Two years after treatment, urease activity decreased in the oil-treated soil in comparison to the control, for both doses. Six years after treatment, urease activity values were 2.5 times lower in soil treated with 3.5 l m−2 and 4.3 times lower in soil treated with 8.5 l m−2 of oil compared to the control soil.
Catalase activity did not statistically vary among the treatments three months after the addition of oil. Later on, catalase activity was not affected by 0.6–1.2 l m−2 treatments while it was lower in soils treated with 3.5 and 8.5 l m−2 of oil than in the control soil (50% and 35% of the control value, respectively).
The low level of oil contamination (0.6 l m−2) did not affect soil enzymes except dehydrogenase. Dehydrogenase activity was significantly (p < 0.05) increased in slightly polluted soil (by 63% in comparison with the control soil). The activity of dehydrogenase was increased, even more (by 109%), in soils treated with higher doses of oil, although the differences among doses 1.2–3.5 l m−2 were not significant. Dehydrogenase activity remained at an enhanced level in contaminated soils throughout the research study.
Effect of oil on plant growth and toxicity of soil
The low rate of oil biodegradation was manifested in the restoration of vegetation. The impact of oil on plant growth depended on the level of pollution. At plots treated with oil at concentrations of 0.6 and 1.2 l m−2, plant growth was observed a year after the introduction of oil (Fig. 4a). Nevertheless, plant cover at these plots was only 37–54% compared to unpolluted soil (control). After two years, the plant cover decreased even more (20–44% compared to control) and increased only the following year. For the least polluted soil (0.6 l m−2), the difference decreased to 20% after three years, and after six years, there were no statistically significant differences between polluted and unpolluted soils. Oil products remaining in the soil by this time did not have a negative effect on plant growth. Despite the low level of pollution at plots with 1.2 l m−2 of oil, plant growth was inhibited even six years after the introduction of oil. Oil concentrations of 3.5 and 8.5% had an even stronger inhibitory effect on vegetation (Fig. 4a). Vegetation growth was inhibited nearly completely during the whole studied period.

Effect of petroleum hydrocarbons on plant cover (a) and phytotoxicity (b–d) of soil contaminated with oil (0.6, 1.2, 3.5, and 8.5 l m−2)
Direct-contact ecotoxicity testing was conducted to evaluate the level of soil toxicity to plants. The results of a contact bioassay measuring germinating capacity as well as root and shoot length of wheat seedlings (Triticum aestivum L.) are presented in Fig. 4b-d. Seed germination was strongly inhibited after soil treatment with oil. Germinating capacity, root and shoot length were severely affected in soil treated with 1.2–8.5 l m−2 of oil (Fig. 4b). A 30–60% decrease in germinating capacity relative to the control was observed. Two years later, the inhibiting effect decreased to 20–35%. After six years, only plots treated with 8.5 l m−2 of oil demonstrated the inhibition of germinating capacity (19% compared to the control).
Root growth was further inhibited. A decrease in root length (42–60% relative to control) was observed in soil polluted with 3.5 and 8.5 l m−2 of oil. After two years of contamination, the germination of wheat seeds was inhibited even more than after one year (65–86% decrease in the root length of wheat seeds). Four years after the introduction of oil, the toxicity of soil decreased (42–50% inhibition of root length). After six years, the differences in the length of wheat roots between soil polluted with 8.5 l m−2 of oil and unpolluted soil were still significant (p < 0.05). Soil polluted with 3.5 l m−2 of oil had no effect on root growth by that time. Oil at a dose of 0.6 and 1.2 l m−2 affected root growth with a smaller effect (up to 30% inhibition), which disappeared after six years of the experiment.
A similar response pattern was found for shoot growth (Fig. 4d). The highest inhibitory effect (up to 85% decrease in shoot length) was observed in the first growing season. After 6 years of pollution, no significant differences in the length of shoots were found between plots polluted with 0.6–3.5 l m−2 of oil and the control (p > 0.05). Soil polluted with the maximum dose of oil (8.5 l m−2) inhibited shoot growth during the whole observation period.
Effect of oil on the abundance and allelopathic activity of soil microorganisms
Oil-polluted soil was characterized by significant changes in soil microbiota. Six years after treatment, fungal abundance was 6–7 times lower compared to uncontaminated soil in soil treated with 0.6–1.2 l of oil m−2, while in more contaminated soil (3.5–8.5 l m−2), it decreased by 30–50 times (Fig. 5a).

Effect of oil on the abundance of fungi (a) and indices of allelopathic activity of fungi calculated by root (b) and shoot length (c) of common wheat Triticum aestivum L.
One year after contamination, no significant differences in fungal abundance were found between soils exposed to different doses of oil. The differences between oil-contaminated and uncontaminated soil began to appear after two years of contamination. At this stage, all samples of contaminated soil demonstrated a significant decrease in fungal abundance (by 65–79% compared to uncontaminated soil). After four years, the number of fungi in contaminated soil was even lower (3–20%) compared to the control, and it declined to a minimum value of 1–2% after six years in soil contaminated with 3.5–8.5 l of oil per m2. In soil contaminated with 0.6–1.2 l of oil per m2, the number of fungi did not exceed 17% compared with uncontaminated soil.
The changes affected not only the number of fungi but also their metabolic activity. The study of the allelopathic activity of fungal metabolites revealed their increased phytotoxicity. The allelopathic indices of soil fungi were determined using the root and shoot length of common wheat Triticum aestivum L. (Fig. 5b, c). Indices were calculated by inhibition of root length (Ka1) and shoot length (Ka2).
Seed germination, root and shoot length of common wheat responded differently to metabolites of soil fungi. Germination of wheat seeds was not sensitive to these compounds, while roots and shoots were affected by fungal metabolites. Root and shoot length turned out to decrease over time, especially in bioassays with the fungal community of highly contaminated soils.
There were no statistically significant differences (p > 0.05) in the allelopathic indices of fungi that inhabited the control soil and soil contaminated with the lowest dose of oil (0.6 l m−2). The root and shoot length of wheat was the same for both contaminated and uncontaminated soils. The mycobiota of these soils did not have toxic effects on plants during the whole study period. The allelopathic indices, calculated based on root and shoot length, did not exceed the threshold value of 0.3. In contrast, the mycobiota of soil treated with 1.2 l m−2 of oil demonstrated increased allelopathic activity (130–160% compared to the control). In soils with higher doses of oil (3.5 and 8.5 l m−2), fungal metabolites had an even more strong impact on plant growth. Six years after the contamination, the allelopathic activity of fungi was 5–7 times higher compared to the control. Moreover, the indices calculated from both roots and shoots exceeded the threshold value of 0.3.
Microbial metabolite responses to oil contamination in soil
Combining classical biochemical and modern methods of analysis (gas chromatography, high-performance liquid chromatography, and mass spectrometry), it is possible to get more objective and complete information on the biochemical characteristics of the studied soil samples. Soil metabolomic analysis could provide evidence for changes in microbial communities and functions in oil-contaminated soil. The research team detected 118 known metabolites, including organic acids, phosphoric acids, carbohydrates, polyols, fatty acids, sterols, phenolic compounds, flavonoids, terpenoids, and amino acids, and there were some metabolites that were not classified. The contents of many metabolites were significantly affected by contamination (p < 0.05), with an increase in 24 metabolites and a decline in 47 metabolites. The changes in metabolites mainly occurred in phenolic compounds, terpenoids, organic acids, and carbohydrates. In the contaminated soil, the percentage of the phenolic compounds and terpenoids increased by 2.3 and 1.2 times, respectively (Fig. 6). Phenolic compounds had the largest number of metabolites increased by oil. Among phenolic compounds, the content of benzoic acid showed a great increase in contaminated soil (Fig. 7a). Compared to the control, there was a higher content of some other phenolic acids in the oil-contaminated soils, including phenylacetic, vanillic, gallic, and ferulic acids.

The effect of oil contamination on the proportion of soil metabolites


The effect of oil contamination on the proportion of soil phenolic compounds (a), organic acids (b), and carbohydrates (c). Values are reported in relative abundance from peak areas relative to an internal standard peak
At the same time, some metabolites negatively responded to contamination. Compared to the control, the content of fatty acids was decreased, especially in the soil treated with the maximum dose of oil. In addition, the content of carbohydrates in the soil was reduced by 17–25% with oil treatment (Fig. 6). Sugar concentrations in the soil varied across oil treatment, with trends typically toward lower sugar concentrations in the oil-contaminated soils (Fig. 7c). The soil treated with oil showed a significant decrease in glucose, mannose, arabinose, and galactose, by (%) 34.8–68.1, 8.4–69.2, 36.6–64.3, and 40.7–100, respectively, although the differences among the different doses of oil were mostly not significant.
The response of organic acids differed among treatments, with a decrease in maleic and galacturonic acids, and an increase in hydroxypropionic, fumaric, and malic acids (Fig. 7b). In addition, the content of some other acids including glyceric acid was positively correlated with oil treatments. The most increased organic acid in the contaminated soil was malic acid. The varying metabolic compositions among soil samples were further confirmed by the heat map analysis of the top 60 metabolites (Fig. 8).

Heat map showing expression profiles of soil samples based on the most significant known metabolites
In the samples of uncontaminated soil, the content of low molecular weight organic substances was mainly represented by sugars, carboxylic acids, and fatty acids. Samples of oil-contaminated soils, in addition to organic sugars, acids, and fatty acids, contained the main compounds of terpenoids and phenolics. The main terpenoids were dehydroacetic acids and quinic acids. The main presented phenolic compounds were 4-hydroxybenzoic acid and gallic acid.
The diversity of low molecular weight organic compounds in oil-contaminated soils turned out to be significantly lower than in the control soil. For example, some amino acids (glycine, valine), polyols (myo-inositol), and sterols (cholesterol, b-sitosterol) were found only in samples of unpolluted soil. On the other hand, malic acid was found in all samples of oil-contaminated soils and was practically not contained in the samples of the control soil. Ellagic acid was not detected in the samples of the control soil, while it was present in the samples of oil-contaminated soils.
Principal component analysis (PCA) revealed oil concentration as a strong driver of the variation among phenolic compounds, accounting for 41% of the total variability (Fig. 9). Similar patterns were observed for organic acids with 31% and terpenoids with 43% variation on PC1, respectively. PCA identified larger differences along PC1 and PC2 between the control and oil-contaminated soils. The main differential compounds between the non-contaminated and contaminated soils were phenolic acids and terpenoids. The analysis showed that certain low oil concentration was correlated with terpenoids, while higher concentrations were correlated with different phenolic compounds.

Principal component analysis plots of the soil metabolites: oil{1}—0.6 l m−2; oil{2}—1.2 l m−2; oil{3}—3.5 l m−2; oil{4}—8.5 l m−2
Discussion
The widespread soil contamination with oil and the toxicity of petroleum hydrocarbons to soil biota makes it extremely important to study microbial responses to stress. The results obtained in this study showed a decrease in total hydrocarbon concentrations and a metabolic shift of the soil microbial community determined by oil. The rate of oil degradation in soil was low. A significant amount of TPH (35–60%) still remained in soils even six years after contamination. The slow TPH degradation rate is apparently related to the persistence and limited bioavailability of remaining hydrocarbons and the deficiency of nutrients necessary for microbial growth and biodegradation. The concentration of nutrients was initially low in sandy podzolic soil, and it decreased even more after treatment with oil (Table 1).
The main reason for the decrease in the content of nutrients in oil-contaminated soil is the hydrophobicity of soil particles covered with an oil film. The contamination of soil by oil leads to increased soil water repellency (Hewelke et al., 2018). Oil-contaminated soils are much worse when wetted with water and thus hydrophobicity prevents the extraction of mobile compounds. As the oil decomposes, the hydrophobicity of the soil is eliminated. Thus, after a few years, nutrient mobility was restored to the control level.
Soil contamination with petroleum hydrocarbons is known to contribute to a high C:N ratio that has an adverse influence on microbial activity (Shahi et al., 2016). Moreover, the degradation of higher molecular weight hydrocarbons may produce toxic intermediate metabolites that can inhibit hydrocarbon-oxidizing microorganisms. The accumulation of inhibiting metabolites may also slow down the biodegradation process (Stroud et al., 2007).
The long-term impact of contamination on soil quality was observed in vegetation restoration. Six years after treatment, plant growth was strongly inhibited in the soil contaminated with 1.2 l m−2 of oil, and almost complete inhibition of plant growth was observed in the plots treated with higher oil concentrations. The direct-contact ecotoxicity testing confirmed the high level of soil toxicity to plants. The inhibition of plant growth is a typical response to oil contamination, and the duration of restoration is especially long for poor soils characterized by low organic carbon and low biological activity (Chugunova et al., 2022; Salanitro et al., 1997).
Due to the slow rate of hydrocarbon biodegradation, oil pollution has a long-term effect on the biological activity of soil. At the beginning of the experiment, the activity of soil enzymes (phosphatase, urease, and catalase) increased in contaminated soils (Fig. 3). A short-term initial increase in the enzymatic activity may be caused by the biodegradation of the easily available hydrocarbons (Dindar et al., 2015). The activity of soil enzymes decreased afterward and remained at a lower level than enzymatic activity in the control soil even six years after contamination. This can be explained by the fact that later, the residual hydrocarbons are comprised mostly of compounds that are recalcitrant and resistant to biodegradation. Heavy oil fractions are known to block enzyme activity by coating organo-mineral and cell surfaces, thereby preventing soluble substrates from reaching the enzyme molecules (Lee et al., 2020).
A different effect was revealed for dehydrogenase activity exceeding the level of the control soil throughout the experiment. Dehydrogenases play an important role in the biodegradation of petroleum hydrocarbons transporting electrons and hydrogen through a chain of intermediate electron carriers to a final electron acceptor (oxygen) (Tate, 2002; Varjani & Upasani, 2017). Dehydrogenase activity reflects the microbial biomass and degradation activity of soil microbiota. It is often strongly affected by the introduction of oil; both positive and negative correlations with oil concentration were reported under various conditions (Polyak et al., 2018). In the present study, dehydrogenase activity decreased over time, which indicates a decrease in microbial oxidative activities in soil. Soil dehydrogenase is considered a good measure of the change of microbial oxidative activity under environmental pollution (Tan et al., 2017; Wolińska & Stępniewska, 2012).
Nutritional deficiencies may limit the degradation potential of soil microorganisms able to oxidize hydrocarbons (Silva-Castro et al., 2015). It was found that oil pollution influenced soil microbiota, significantly changing the total number of soil fungi. Fungal abundance decreased up to 50 times depending on the concentration of oil. The identified changes were dependent on the duration of contamination. The lowest number of fungi was revealed in the soil contaminated with maximum oil concentrations (3.5–8.5 l per m2) six years after treatment.
Oil contamination is known to inhibit the growth of sensitive microorganisms and increase the number of hydrocarbon-degrading microorganisms (Ławniczak et al., 2020; Xu et al., 2018). Hydrocarbon-degrading fungi become beneficial for contaminated environments as they easily adapt to extreme conditions and contribute to hydrocarbon removal from soil (Borowik et al., 2017; Marchand et al., 2017). Among fungi demonstrating high efficiency in the removal of petroleum products from the soil are species of the genera Aspergillus, Candida, Alternaria, Cladosporium, Paecilomyces, and Trichoderma, Fusarium (Ameen et al., 2016; El-Hanafy et al., 2017; Jiang et al., 2016; Marchand et al., 2017; Silva et al., 2015). Representatives of these genera synthesize many biologically active substances, including enzymes, antibiotics, and various toxins.
Fungal toxins may affect soil organisms: plants, animals, fungi, and bacteria (Battilani et al., 2016). Many hydrocarbon-degrading fungi produce allelopathic substances influencing the growth and reproduction of other organisms (Ismaiel & Papenbrock, 2015; Polyak et al., 2020; Selbmann et al., 2013). The allelopathic effect may be enhanced under stress conditions caused by nutrient deficiency and changes in the soil moisture content (Barazani & Friedman, 2001). The allelopathic activity of fungi in the contaminated soil increased. Fungal metabolites had the strongest impact on plant growth (5–7 times higher compared to the control, in soils treated with high doses of oil). The indices of allelopathic activity calculated for both roots and shoots of common wheat exceeded the threshold value of 0.3. Thus, on the one hand, fungi take part in oil degradation improving soil quality, and on the other hand, hydrocarbons influence the allelopathic activity of fungi increasing soil toxicity.
Depending on the nature of hydrocarbons, they are generally transformed by degrading microorganisms through biochemical processes to more polar organic compounds such as alcohols, organic acids, aldehydes, and ketones (Okoh, 2006). Soil microorganisms exhibit a great diversity of metabolic abilities. The metabolites isolated from soil samples not only reflect the chemical composition of the soil but also depend on the microorganisms present in the soil. Thus, shifts in the composition of soil metabolites can help to evaluate the effect of contaminants on soil microbial processes.
Little research has been carried out on the metabolic profile of soil in response to long-term stress. Earlier, Li et al. (2019) demonstrated an obvious shift in soil microbial function in agricultural soil under fresh polycyclic aromatic hydrocarbons stress. The present study is the first to apply metabolomics approaches to distinguish poor soils that are contaminated and non-contaminated by oil. We used untargeted metabolomics as an unbiased metabolomics analysis that can discover new biomarkers. In contrast to targeted metabolomics, which measures ions from limited known metabolites, an untargeted approach registers all ions in a sample (Vinayavekhin & Saghatelian, 2010).
The changes in metabolites mainly occurred in carbohydrates, organic acids, phenolic compounds, and terpenoids. In general, the most oil-polluted soils were characterized by the least diversity and quantitative content of low molecular weight organic compounds. Carbohydrates had the largest number of metabolites negatively affected by oil. Moreover, sterols and flavonoids were mainly reduced in the contaminated soil, while phenolic acids were mainly increased.
Carbohydrates were significantly changed by oil treatments, which specifically decreased glucose, mannose, arabinose, and galactose inducing changes in soil functioning. These monosaccharides are among the most important primary inputs of organic carbon in the soil. Typically, they represent more than 90% of total hydrolyzable carbohydrates (Chesire, 1977). The ratio greater than 1 of mannose to xylose indicates that a soil carbohydrate material is synthesized mostly by microorganisms in contaminated soils (Hu et al., 1995). Glucose is the dominant hexose among soil sugars. The decrease in the amount of glucose in contaminated soil may be explained by its diverse origins. Glucose is released by plant roots, from the decomposed cellulose of plant residues, and synthesized by microorganisms (Gunina & Kuzyakov, 2015), while both plants and microorganisms are suppressed in contaminated soil.
Meanwhile, the importance of soil carbohydrates as carbon and energy sources for soil microbiota, as well as their effect on the stability of soil particle aggregates, is well known (Coelho et al., 1988; Uzoho & Igbojionu, 2014). Sugar metabolism regulates many pathways and cycles such as glycolysis and the citric acid cycle, which are essential in various activities of soil microbiota (Sun et al., 2022). According to metabolic analysis, the decrease in compounds related to sugar metabolism may significantly contribute to changes in metabolic pathways in the control soil and soils treated with different doses of oil.
The important differences between the metabolic profile types of unpolluted and polluted soil are the higher concentrations of organic acids in the former. These differences may suggest microbial stress response and the relationship between metabolites. An increase in carboxylic and dicarboxylic acids (hydroxypropionic, fumaric, and malic) was observed. Both bacteria and fungi are capable of producing various organic acids, but in terms of productivity, fungi are incomparable (Karaffa et al., 2021). Fungi are known to be the best fumaric acid producers, mainly species from the genera Aspergillus, Rhizopus, Mucor, Cunninghamella, and Cirinella (Jiménez-Quero et al., 2017; Yang et al., 2017). Malic acid can be produced by species of the fungus Aspergillus (West, 2015). In filamentous fungi, there is a significant accumulation of fumaric, malic, and citric acids under specific stress conditions (Goldberg et al., 2006). Thus, after the impact of oil, some fungi accumulate intermediates of the Krebs cycle as end products.
Organic acids are among the major metabolites exhibiting allelopathic potential. The most commonly reported soil allelochemicals are phenolic acids (Latif et al., 2017). The contents of phenolic acids were higher in the contaminated soils with the highest number of fungi. It is known that phenolic acids are synthesized and perform physiological functions in plants (Cosme et al., 2020; Vuolo et al., 2018). They are no less characteristic of fungi (Valanciene et al., 2020). Moreover, phenolic acids, in high concentrations, were found in samples of heavily polluted soils, where vegetation cover was practically absent.
Soil fungi produce antioxidants including phenolic compounds, flavonoids, saponins, steroids, tannins, terpenoids, and alkaloids. The antioxidant effect of phenolic compounds is determined by their high anti-radical activity, which is due to hydroxyl groups forming a conjugated system with double bonds of the benzene ring. High antioxidant activity was demonstrated by soil isolates Penicillium citrinum and P. granulatum, Aspergillus wentii (Arora & Chandra, 2011). Among fungi with high biochemical activity are A. phoenicis, A. proliferans, A. niger, and P. chrysogenum (Ameen et al., 2022). Fungal responses to petroleum hydrocarbons can be reflected by differences in metabolic responses, enzyme induction and synthesis, and extracellular metabolite production.
Phenolic acids are classified as hydroxybenzoic and hydroxycinnamic acids. Hydroxybenzoic acids are composed of a C6-C1 skeleton and include benzoic acid, p-hydroxybenzoic acid, vanillic acid, gallic acid, protocatechuic acid, syringic acid, gentisic acid, veratric acid, and salicylic acid. Hydroxycinnamic acids have a C6-C3 skeleton and include cinnamic acid, coumaric acid, ferulic acid, sinapic acid, and caffeic acid (Pratyusha, 2022).
The contents of 4-hydroxybenzoic acid and gallic acid showed the greatest increase by contamination, inferring that the contents of these phenolic acids, which are metabolites with high biological activity, clearly positively responded to oil stress. 4-hydroxybenzoic acid is known as a product of fungal benzoate catabolism (Wright, 1993). In most fungi, benzoic acid is hydroxylated to 4-hydroxybenzoic acid, which is then also hydroxylated to protocatechuic acid, resulting in ring cleavage and the b-ketoadipate pathway (Mäkelä et al., 2015).
Earlier, high concentration of phenolic compounds, especially gallic acid, was shown to contribute to the biological activity of Penicillium flavigenum (Tavares et al., 2018). In addition, Aspergillus niger and A. fischeri are capable of producing gallic acid during the assimilation and catabolism of organic compounds (Bajpai & Patil, 2008; Valanciene et al., 2020). Robledo et al. (2008) reported the high production of ellagic acid, a dimeric gallic acid derivative, by Aspergillus niger as a result of the hydrolysis of ellagitannins. In the present study, ellagic acid was found in the oil-contaminated soil and was practically not contained in the control soil.
The proportion of ellagic and quinic acids also changed significantly in the contaminated soil. Ellagic acid is a powerful bioactive compound with antimicrobial and antioxidant properties. These molecules exhibit high antioxidant capacity due to the stability of free radicals (Evtyugin et al., 2020). They can be produced by A. niger and A. fumigatus. Quinic acid is efficiently produced by A. niger, A. awamori, A. sojae, P. chrysogenum, Mucorales mucor, Mucorales rhizopus, and Monascus purpureus as a metabolic intermediate of the shikimic acid pathway (Patent, 2009).
The concentration of vanillin and vanillic acid increased significantly in the oil-contaminated soil. The aromatic metabolism of fungi includes the conversion of ferulic acid into vanillic acid, which is then converted to vanillin. This pathway was observed for A. niger, Botrytis, Cephalosporium, Penicillium, Trichoderma, and Verticillium species (Lesage-Meessen et al., 1996; Mäkelä et al., 2015). A demethylation pathway of ferulic acid to caffeic acid was described for a Penicillium species (Tillett & Walker, 1990).
The results obtained in this study indicate that the production of phenolic acids may be enhanced under stress conditions caused by soil contamination with oil. Biologically active substances of this group are accumulated in soil and inhibit the germination of seeds and plant growth. Taking into account the particular role of phenolic compounds in soil ecosystems, their increased content causes a significant allelopathic tension in phytocenoses (Polyak & Sukcharevich, 2019).
PCA revealed that the first two principal components, (PC1 and PC2), explained 76.38% of the total variance and soil samples treated with different doses of oil obviously differed from each other. PCA gave a preliminary indication of a relationship between the allelopathic activity of fungi isolated from soil and soil contamination. Metabolomics helps to identify biomarkers indicative of specific environmental conditions. The significant loadings of phenolic compounds, terpenoids, and organic acids toward contaminated soils indicate that these metabolites are specific to these soils, implying the potential role of fungi in soil properties. Identification of such metabolites may help to evaluate microbial succession and functioning pathways. The metabolomic approach provides an opportunity to identify the features of the development and biochemical activity of microbial communities in oil-polluted soil, which can be useful in the development of remediation strategies for contaminated environments (Sharma et al., 2022).
Conclusions
The obtained results revealed the long influence of oil on the biological activity of soil and vegetation. The degradation potential of soil microorganisms was limited by the nutritional deficiency of poor sandy podzolic soil. The effect of oil pollution on soil microbiota was observed in changes in the abundance and allelopathic activity of soil fungi increasing the production of toxic metabolites. Untargeted metabolomics was employed to determine soil metabolites, as well as their changing trends in oil-contaminated soil. The concentration and profile of metabolites were found to vary greatly between oil-contaminated and non-contaminated soils. The changes in metabolites mainly occurred in carbohydrates, organic acids, phenolic compounds, and terpenoids. Some metabolite compounds, which changed under stress conditions, may influence soil quality and plants. It can be concluded that oil contamination can alter the composition and activity of the soil microbial community toward a higher abundance of fungi-producing bioactive compounds that inhibit the growth of plants. The evaluation of the long-term impact of oil on microbial metabolism can make a valuable contribution to the assessment of soil quality and the activity of soil microorganisms being under stress from oil pollution. The results contribute to a further understanding of the role of microorganisms in the ecological functions of disturbed soil, which can be useful in the development of rehabilitation strategies for post-industrial lands. More work is needed to explore the metabolomic responses of different types of soil to identify overall shifts in soil microbial metabolic function and specific biomarkers of oil pollution.
References
Albert, E., & Tanee, F. (2011). A laboratory trial of bioaugmentation for removal of total petroleum hydrocarbon (TPH) in Niger Delta soil using Oscillatoria bornettia. Journal of Microbiology and Biotechnology, 1, 147–168.
Alrumman, S. A., Standing, D. B., & Paton, G. I. (2015). Effects of hydrocarbon contamination on soil microbial community and enzyme activity. Journal of King Saud University, 27, 31–41. https://doi.org/10.1016/j.jksus.2014.10.001
Ameen, F., AlNAdhari, S., Yassin, M. A., Al-Sabri, A., Almansob, A., Alqahtani, N., & Stephenson, S. (2022). Desert soil fungi isolated from Saudi Arabia: Cultivable fungal community and biochemical production. Saudi Journal of Biological Sciences, 29(4), 2409–2420. https://doi.org/10.1016/j.sjbs.2021.12.011
Ameen, F., Moslem, M., Hadi, S., & Al-Sabri, A. E. (2016). Biodegradationof diesel fuel hydrocarbons by mangrove fungi from Red Sea Coast of Saudi Arabia. Saudi Journal of Biological Sciences, 23, 211–218. https://doi.org/10.1016/j.sjbs.2015.04.005
Arinushkina, E. V. (1970). Manual on chemical analysis of soils. Publishing House of Moscow State University. In Russian.
Arora, D. S., & Chandra, P. (2011). In vitro antioxidant potential of some soil fungi: Screening of functional compounds and their purification from Penicillium citrinum. Applied Biochemistry and Biotechnology, 165, 639. https://doi.org/10.1007/s12010-011-9282-3
Bajpai, B., & Patil, S. (2008). A new approach to microbial production of gallic acid. Brazilian Journal of Microbiology, 39, 708–711. https://doi.org/10.1590/S1517-83822008000400021
Bakina, L. G., Chugunova, M. V., Polyak, Y. M., Mayachkina, N. V., & Gerasimov, A. O. (2021). Bioaugmentation: Possible scenarios due to application of bacterial preparations for remediation of oil contaminated soil. Environmental Geochemistry and Health, 43(6), 2347–2356. https://doi.org/10.1007/s10653-020-00755-4
Barazani, O., & Friedman, J. (2001). Allelopathic bacteria and their impact on higher plants. Critical Reviews in Microbiology, 27(1), 41–55. https://doi.org/10.1080/20014091096693
Battilani, P., Stroka, J., & Magan, N. (2016). Foreword: Mycotoxins in a changing world. World Mycotoxin Journal, 9, 647–651. https://doi.org/10.3920/WMJ2016.x004
Boer, M., Ellers, J., Gestel, C. M., Dunnen, J., Straalen, N., & Roelofs, D. (2013). Transcriptional responses indicate attenuated oxidative stress in the springtail Folsomia candida exposed to mixtures of cadmium and phenanthrene. Ecotoxicology, 22, 619–631. https://doi.org/10.1007/s10646-013-1053-1
Borowik, A., Wyszkowska, J., & Oszust, K. (2017). Functional diversity of fungal communities in soil contaminated with diesel oil. Frontiers in Microbiology, 8, 1862. https://doi.org/10.3389/fmicb.2017.01862
Chesire, M. V. (1977). Origins and stability of soil polysaccharide. European Journal of Soil Science, 98, 371–376.
Chugunova, M. V., Bakina, L. G., Mayachkina, N. V., Polyak, Y. M., & Gerasimov, A. O. (2022). Features of the processes of detoxification and self-restoration of oil-contaminated soils—a field study. Journal of Soils and Sediments, 22, 3087–3105. https://doi.org/10.1007/s11368-022-03272-2
Coelho, R. R. R., Linhares, L. F., & Martin, J. P. (1988). Sugars in hydrolysates of fungal melanins and soil humic acids. Plant and Soil, 106(1), 127–133.
Cosme, P., Rodríguez, A. B., Espino, J., & Garrido, M. (2020). Plant phenolics: Bioavailability as a key determinant of their potential health-promoting applications. Antioxidants (basel), 9(12), 1263. https://doi.org/10.3390/antiox9121263
Danilova, A. A. (2010). Phytotoxicity of fungal complex in leached chernozem of Priob’e under different methods of main soil cultivation. Agricultural Biology, 3, 108–111. In Russian.
Dindar, E., Şağban, F. O. T., & Başkaya, H. S. (2015). Variations of soil enzyme activities in petroleum-hydrocarbon contaminated soil. International Biodeterioration and Biodegradation, 105, 268–275. https://doi.org/10.1016/j.ibiod.2015.09.011
Dospehov, B. A. (1985). Methods of field experience. Agroprodpublishing. In Russian.
El-Hanafy, A. A., Anwar, Y., Sabir, J. S. M., Mohamed, S. A., Al-Garni, S. M. S., Zinadah, O. S. H., et al. (2017). Characterization of native fungi responsible for degrading crude oil from the coastal area of Yanbu, Saudi Arabia. Biotechnology and Biotechnological Equipment, 31, 105–111. https://doi.org/10.1080/13102818.2016.1249407
Evtyugin, D. D., Magina, S., & Evtuguin, D. V. (2020). Recent advances in the production and applications of ellagic acid and its derivatives. A review. Molecules, 25(12), 2745. https://doi.org/10.3390/molecules25122745
Geng, S., Xu, G., You, Y., Xia, M., Zhu, Y., Ding, A., Fan, F., & Dou, J. (2022). Occurrence of polycyclic aromatic compounds and interdomain microbial communities in oilfield soils. Environmental Research, 212(Part A), 113191. https://doi.org/10.1016/j.envres.2022.113191
Goldberg, I., Rokem, J. S., & Pines, O. (2006). Organic acids old metabolites new themes. Journal of Chemical Technology and Biotechnology, 81, 1601–1611. https://doi.org/10.1002/jctb.1590
Griffin, J. L. (2014). Metabolomic analysis of soil communities can be used for pollution assessment. Environmental Toxicology and Chemistry, 33, 61–64. https://doi.org/10.1002/etc.2418
Gunina, A., & Kuzyakov, Y. (2015). Sugars in soil and sweets for microorganisms: Review of origin, content, composition and fate. Soil Biology and Biochemistry. https://doi.org/10.1016/j.soilbio.2015.07.021
Haziev, F. H. (2005). Methods of soil enzymology. Nauka. In Russian.
Hernandez-Soriano, M. C., Jimenez-Lopez, J. C. (2014). Metabolomics for soil contamination assessment. In Environmental risk assessment of soil contamination (pp. 459–481). Rijeka. https://doi.org/10.5772/58294
Hewelke, E., Szatyłowicz, J., Hewelke, P., Gnatowski, T., & Aghalarov, R. (2018). The impact of diesel oil pollution on the hydrophobicity and CO2 efflux of forest soils. Water Air Soil Pollution, 229(2), 51. https://doi.org/10.1007/s11270-018-3720-6
Hu, S., Coleman, D. C., Beare, M. H., & Hendrix, P. F. (1995). Soil carbohydrate in aggrading and degrading ecosystem: Influences of fungi and aggregates. Agriculture, Ecosystems and Environment, 54, 77–88. https://doi.org/10.1016/0167-8809(95)00588-J
Ismaiel, A. A., & Papenbrock, J. (2015). Mycotoxins: Producing fungi and mechanisms of phytotoxicity. Agriculture, 5, 492–537. https://doi.org/10.3390/agriculture5030492
IUSS Working Group WRB (2006). world reference base for soil resources 2006. World Soil Resources Report No. 103. FAO, Rome.
Jiang, S., Wang, W., Xue, X., Cao, C., & Zhang, Y. (2016). Fungal diversity in major oil-shale mines in China. Journal of Environmental Sciences, 41, 81–89. https://doi.org/10.1016/j.jes.2015.04.032
Jiménez-Quero, A., Pollet, E., Zhao, M., Marchioni, E., Averous, L., & Phalip, V. (2017). Fungal fermentation of lignocellulosic biomass for itaconic and fumaric acid production. Journal of Microbiology and Biotechnology, 27, 1–8. https://doi.org/10.4014/jmb.1607.07057
Johnsen, L. G., Skou, P. B., Khakimov, B., & Bro, R. (2017). Gas chromatography—Mass spectrometry data processing made easy. Journal of Chromatography A, 1503, 57–64. https://doi.org/10.1016/j.chroma.2017.04.052
Johnson, J. L., & Temple, K. L. (1964). Some variables affecting measurement of catalase activity in soil. Soil Science Society of America Journal, 28, 207–216.
Kapel’kina L. P., Bardina T. V., Bakina L. G., Chugunova M.V., Gerasimov A. O., Mayachkina N. V., & Galdiyants A. A. (2009). Measurement of seed germination rate and root length of higher plant seedlings for determining toxicity of technogenically contaminated soils: MP-2006. FR.1.39.2006.02264. St. Petersburg. (in Russian).
Karaffa, L., Fekete, E., & Kubicek, C. P. (2021). The role of metal Ions in fungal organic acid accumulation. Microorganisms, 9(6), 1267. https://doi.org/10.3390/microorganisms9061267
Latif, S., Chiapusio, G., & Weston, L. A. (2017). Chapter Two-allelopathy and the role of allelochemicals in plant defense. Advances in Botanical Research, 82, 19–54. https://doi.org/10.1016/bs.abr.2016.12.001
Ławniczak, Ł, Woźniak-Karczewska, M., Loibner, A. P., Heipieper, H. J., & Chrzanowski, Ł. (2020). Microbial degradation of hydrocarbons-basic principles for bioremediation: A review. Molecules, 25(4), 856. https://doi.org/10.3390/molecules25040856
Lee, S.-H., Kim, M.-S., Kim, J.-G., & Kim, S.-O. (2020). Use of soil enzymes as indicators for contaminated soil monitoring and sustainable management. Sustainability, 12, 8209. https://doi.org/10.3390/su12198209
Lesage-Meessen, L., Delattre, M., Haon, M., Thibault, J. F., Ceccaldi, B. C., Brunerie, P., et al. (1996). A two-step bioconversion process for vanillin production from ferulic acid combining Aspergillus niger and Pycnoporus cinnabarinus. Journal of Biotechnology, 50, 107–113. https://doi.org/10.1016/0168-1656(96)01552-0
Li, X., Qu, C., Bian, Y., Gu, C., Jiang, X., & Song, Y. (2019). New insights into the responses of soil microorganisms to polycyclic aromatic hydrocarbon stress by combining enzyme activity and sequencing analysis with metabolomics. Environmental Pollution, 255(Part A), 113312. https://doi.org/10.1016/j.envpol.2019.113312
Mäkelä, M. R., Marinović, M., Nousiainen, P., Liwanag, A. J., Benoit, I., Sipilä, J., Hatakka, A., de Vries, R. P., & Hildén, K. S. (2015). Aromatic metabolism of filamentous fungi in relation to the presence of aromatic compounds in plant biomass. Advances in Applied Microbiology, 91, 63–137. https://doi.org/10.1016/bs.aambs.2014.12.001
Marchand, C., St-Arnaud, M., Hogland, W., Bell, T. H., & Hijri, M. (2017). Petroleum biodegradation capacity of bacteria and fungi isolated from petroleum-contaminated soil. International Biodeterioration and Biodegradation, 116, 48–57. https://doi.org/10.1016/j.ibiod.2016.09.030
Mirkin, B. M., Naumova, L. G., & Solomeshch, A. I. (2002). Modern science of vegetation. Logos. in Russian.
Okoh, A. I. (2006). Biodegradation alternative in the cleanup of petroleum hydrocarbon pollutants. Biotechnology and Molecular Biology Reviews, 1(2), 38–50.
Onwosi, C. O., Odimba, J. N., Igbokwe, V. C., Nduka, F. O., Nwagu, T. N., Aneke, C. J., & Eke, I. E. (2020). Principal component analysis reveals microbial biomass carbon as an effective bioindicator of health status of petroleum-polluted agricultural soil. Environmental Technology, 41(24), 3178–3190. https://doi.org/10.1080/09593330.2019.1603252
Pagano, L., Serving, A. D., De La Torre-Roche, R., Mukherjee, A., Majumdar, S., Hawthorne, J., Marmiroli, M., Maestri, E., Marra, R. E., Isch, S. M., Dhankher, O. P., White, J. C., & Marmiroli, N. (2016). Molecular response of crop plants to engineered nanomaterials. Environmental Science and Technology, 50, 7198–7207. https://doi.org/10.1021/acs.est.6b01816
Patent WO2009107328. (2009). Process for production of quinic acid and/or caffeic acid.
Patil, C., Calvayrac, C., Zhou, Y., Romdhane, S., Salvia, M. V., Cooper, J. F., Dayan, F. E., & Bertrand, C. (2016). Environmental metabolic footprinting: A novel application to study the impact of a natural and a synthetic β-triketone herbicide in soil. Science of the Total Environment, 566–567, 552–558. https://doi.org/10.1016/j.scitotenv.2016.05.071
PND F 16.1:2.21-98. (2012). Quantitative chemical analyses of soils. Moscow (in Russian).
Polyak, Y. M., Bakina, L. G., Chugunova, M. V., Mayachkina, N. V., Gerasimov, A. O., & Bure, V. M. (2018). Effect of remediation strategies on biological activity of oil-contaminated soil—A field study. International Biodeterioration and Biodegradation, 126, 57–68. https://doi.org/10.1016/j.ibiod.2017.10.004
Polyak, Y., Bakina, L., Mayachkina, N., & Polyak, M. (2020). The possible role of toxigenic fungi in ecotoxicity of two contrasting oil-contaminated soils—A field study. Ecotoxicology and Environmental Safety, 202, 110959. https://doi.org/10.1016/j.ecoenv.2020.110959
Polyak, Y. M., & Sukcharevich, V. I. (2019). Allelopathic interactions between plants and microorganisms in soil ecosystems. Biology Bulletin Reviews, 9, 562–574. https://doi.org/10.1134/S2079086419060033
Pratyusha, S. (2022). Phenolic compounds in the plant development and defense: An overview. In M. Hasanuzzaman & K. Nahar (Eds.), Plant stress physiology—Perspectives in agriculture. IntechOpen. https://doi.org/10.5772/intechopen.102873
Robledo, A., Aguilera-Carbó, A., Rodriguez, R., Martinez, J. L., Garza, Y., & Aguilar, C. N. (2008). Ellagic acid production by Aspergillus niger in solid state fermentation of pomegranate residues. Journal of Industrial Microbiology and Biotechnology, 35(6), 507–513. https://doi.org/10.1007/s10295-008-0309-x
Rochfort, S., Ezernieks, V., Mele, P., & Kitching, M. (2015). NMR metabolomics for soil analysis provide complementary, orthogonal data to MIR and traditional soil chemistry approaches—A land use study. Magnetic Resonance in Chemistry, 53, 719–725. https://doi.org/10.1002/mrc.4187
Salanitro, J. P., Dorn, P. B., Huesemann, M. H., Moore, K. O., Rhodes, I. A., Rice Jackson, L. M., Vipond, T. E., Western, M. M., & Wisniewski, H. L. (1997). Crude oil hydrocarbon bioremediation and soil ecotoxicity assessment. Environmental Science and Technology, 31, 1769–1776. https://doi.org/10.1021/es960793i
Selbmann, L., Egidi, E., Isola, D., Onofri, S., Zucconi, L., De Hoog, G. S., Chinaglia, S., Testa, L., Tosi, S., Balestrazzi, A., Lantieri, A., Compagno, R., Tigini, V., & Varese, G. C. (2013). Biodiversity, evolution and adaptation of fungi in extreme environments. Plant Biosystems—an International Journal Dealing with All Aspects of Plant Biology, 147(1), 237–246. https://doi.org/10.1080/11263504.2012.753134
Shahi, A., Aydin, S., Ince, B., & Ince, O. (2016). Evaluation of microbial population and functional genes during the bioremediation of petroleum-contaminated soil as an effective monitoring approach. Ecotoxicology and Environmental Safety, 125, 153–160. https://doi.org/10.1016/j.ecoenv.2015.11.029
Sharma, P., Singh, S. P., Iqbal, H. M. N., & Tong, Y. W. (2022). Omics approaches in bioremediation of environmental contaminants: An integrated approach for environmental safety and sustainability. Environmental Research, 211, 113102. https://doi.org/10.1016/j.envres.2022.113102
Silva, D. P., Cavalcanti, D. D., de Melo, E. V., dos Santos, P. N. R., da Luz, E. L. P., de Gusmao, N. B., et al. (2015). Bio-removal of diesel oil through amicrobial consortium isolated from a polluted environment. International Biodeterioration and Biodegradation, 97, 85–89. https://doi.org/10.1016/j.ibiod.2014.09.021
Silva-Castro, G. A., Uad, I., Rodríguez-Calvo, A., González-López, J., & Calvo, C. (2015). Response of autochthonous microbiota of diesel polluted soils to landfarming treatments. Environmental Research, 137, 49–58. https://doi.org/10.1016/j.envres.2014.11.009
Souza, E. C., Vessoni-Penna, T. C., & de Souza Oliveira, R. P. (2014). Biosurfactant-enhanced hydrocarbon bioremediation: An overview. International Biodeterioration and Biodegradation, 89, 88–94. https://doi.org/10.1016/j.ibiod.2014.01.007
Stroud, J. L., Paton, G. I., & Semple, K. T. (2007). Microbe-aliphatic hydrocarbon interactions in soil: Implications for biodegradation and bioremediation. Journal of Applied Microbiology, 102(5), 1239–1253. https://doi.org/10.1111/j.1365-2672.2007.03401.x
Sumner, L. W., Amberg, A., Barrett, D., Beale, M. H., Beger, R., Daykin, C. A., Fan, T. W., Fiehn, O., et al. (2007). Proposed minimum reporting standards for chemical analysis. Metabolomics, 3, 211–221. https://doi.org/10.1007/s11306-007-0082-2
Sun, L., Wang, Y., Ma, D., Wang, L., Zhang, X., Ding, Y., Fan, K., Xu, Z., Yuan, C., Jia, H., et al. (2022). Differential responses of the rhizosphere microbiome structure and soil metabolites in tea (Camellia sinensis) upon application of cow manure. BMC Microbiology, 22, 55. https://doi.org/10.1186/s12866-022-02470-9
Swenson, T. L., & Northen, T. R. (2019). Untargeted soil metabolomics using liquid chromatography–mass spectrometry and gas chromatography–mass spectrometry. In E. Baidoo (Ed.), Microbial metabolomics. Methods in molecular biology. (Vol. 1859). Humana Press. https://doi.org/10.1007/978-1-4939-8757-3_4
Tan, X., Wang, Z., Lu, G., He, W., Wei, G., Huang, F., Xu, X., & Shen, W. (2017). Kinetics of soil dehydrogenase in response to exogenous Cd toxicity. Journal of Hazardous Materials, 329, 299–309. https://doi.org/10.1016/j.jhazmat.2017.01.055
Tate, R. L. I. I. I. (2002). Microbiology and enzymology of carbon and nitrogen cycling. In R. G. Burns & R. P. Dick (Eds.), Enzymes in the environment: Activity, ecology and applications (pp. 227–248). Marcel Dekker Inc.
Tavares, D. G., Barbosa, B. V. L., Ferreira, R. L., Duarte, W. F., & Cardoso, P. G. (2018). Antioxidant activity and phenolic compounds of the extract from pigment-producing fungi isolated from Brazilian caves. Biocatalysis and Agricultural Biotechnology, 16, 148–154. https://doi.org/10.1016/j.bcab.2018.07.031
Tillett, R., & Walker, J. R. L. (1990). Metabolism of ferulic acid by a Penicillium sp. Archives of Microbiology, 154, 206–208. https://doi.org/10.1007/BF00423334
Tremaroli, V., Workentine, M. L., Weljie, A. M., Vogel, H. J., Ceri, H., Viti, C., Tatti, E., Zhang, P., Hynes, A. P., Turner, R. J., & Zannoni, D. (2009). Metabolomic investigation of the bacterial response to a metal challenge. Applied and Environmental Microbiology, 75, 719–728. https://doi.org/10.1128/AEM.01771-08
Uzoho, B. U., & Igbojionu, G. U. (2014). Carbohydrate distribution of particle size fractions of soils in relation to land-use types in Mbaise, Southeastern Nigeria. Journal of Biology, Agriculture and Healthcare, 4(2), 27–36.
Valanciene, E., Jonuskiene, I., Syrpas, M., Augustiniene, E., Matulis, P., Simonavicius, A., & Malys, N. (2020). Advances and prospects of phenolic acids production, biorefinery and analysis. Biomolecules, 10(6), 874. https://doi.org/10.3390/biom10060874
Varjani, S. J., & Upasani, V. N. (2017). A new look on factors affecting microbial degradation of petroleum hydrocarbon pollutants. International Biodeterioration and Biodegradation, 120, 71–83. https://doi.org/10.1016/j.ibiod.2017.02.006
Vinayavekhin, N., & Saghatelian, A. (2010). Untargeted metabolomics. Current Protocols in Molecular Biology Chapter 30, Unit, 30(1), 1–24. https://doi.org/10.1002/0471142727.mb3001s90
Vuolo, M. M., Lima, V. S., & Maróstica-Junior, M. R. (2018). Phenolic compounds: Structure, classification, and antioxidant power. In M. R. Segura-Campos (Ed.), Bioactive compounds: Health benefits and potential applications (pp. 33–50). Elsevier Inc.
West, T. P. (2015). Fungal biotransformation of crude glycerol into malic acid. Zeitschrift Für Naturforschung C, 70(5–6), 165–167. https://doi.org/10.1515/znc-2015-0115
Withers, E., Hill, P. W., Chadwick, D. R., & Jones, D. L. (2020). Use of untargeted metabolomics for assessing soil quality and microbial function. Soil Biology and Biochemistry, 143, 107758. https://doi.org/10.1016/j.soilbio.2020.107758
Wolińska, A., & Stępniewska, Z. (2012). Dehydrogenase activity in the soil environment. In R. A. Canuto (Ed.), Dehydrogenases. InTech. https://doi.org/10.5772/48294
Wright, J. D. (1993). Fungal degradation of benzoic acid and related compounds. World Journal of Microbiology and Biotechnology, 9, 9–16. https://doi.org/10.1007/BF00656508
Wyszkowska, J., Borowik, A., & Kucharski, J. (2015). Response of Avena sativa, microorganisms and enzymes to contamination of soil with diesel oil. Plant, Soil and Environment, 61, 483–488. https://doi.org/10.17221/463/2015-PSE
Xu, X., Liu, W., Tian, S., Wang, W., Qi, Q., Jiang, P., Gao, X., Li, F., Li, H., & Yu, H. (2018). Petroleum hydrocarbon-degrading bacteria for the remediation of oil pollution under aerobic conditions: A perspective analysis. Frontiers in Microbiology, 9, 2885. https://doi.org/10.3389/fmicb.2018.02885
Yang, L., Lübeck, M., & Lübeck, P. S. (2017). Aspergillus as a versatile cell factory for organic acid production. Fungal Biology Reviews, 31, 33–49. https://doi.org/10.1016/j.fbr.2016.11.001
Zvyagintsev, D. G. (1991). Methods of soil microbiology and biochemistry. MSU. In Russian.
Acknowledgements
The authors acknowledge the financial support from the RSF. We are grateful to the staffs of Laboratory of Methods for Remediation of Technogenic Landscapes for their assistance in the annual field sample collection.
Funding
This work was supported by the Russian Science Foundation (RSF) (Grant No. 22-24-00580).
Author information
Authors and Affiliations
Contributions
YP and LB contributed to the conceptualization of the study and analysis of results; YP, LB, NM, MC, KY, and AS contributed to the laboratory measurements; YP, LB, KY, NB, and AS contributed to methodology of the study; YP prepared the manuscript which was reviewed by all authors.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Polyak, Y.M., Bakina, L.G., Mayachkina, N.V. et al. Long-term effects of oil contamination on soil quality and metabolic function. Environ Geochem Health 46, 13 (2024). https://doi.org/10.1007/s10653-023-01779-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10653-023-01779-2