
Abstract
Climate change and its associated symptoms, such as warming air temperatures, glacier retreat, reduced snowpack, and increased variability in precipitation, are warming rivers and lakes. Such warming water temperatures threaten cold-water fishes in some regions. For instance, warm water temperatures can kill or harm anadromous Pacific salmon as they migrate upstream to spawning grounds. In this study, we assessed how tributaries, and their relative watershed properties, shape temporal and spatial dynamics of temperatures during the summer months in a salmon migratory river. We focused on the mainstem of the Babine River of British Columbia, an important migratory corridor for steelhead and the five eastern Pacific salmon species, but particularly for sockeye salmon that spawn in stream reaches above the Babine Lake, at the river’s headwaters. We discovered that large glacier- and snowpack-fed tributaries cooled the Babine River by approximately 2 °C over its 96 km length. Different tributaries played different temperature functions. Cooler and more glacierized rivers were associated with a bigger change in temperature between upstream and downstream sites. Understanding how water temperatures change during adult salmon migration, especially within riverscapes on the edge of potentially harmful temperatures, can help inform proactive management options in a warming world.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Climate change is warming rivers and lakes and threatening cold-water freshwater fishes (Schindler 2001; Strayer and Dudgeon 2010). For example, the mean summer water temperature of the Fraser River in British Columbia, Canada, has increased by ~ 1.5 °C since the 1950s (Patterson et al. 2007). This warming has caused mass mortality to some populations of adult migrating Pacific salmon (Oncorhynchus spp.) in particularly hot years, and is predicted to drive a 9–16% decrease in Pacific salmon survival by the end of the century (Martins et al. 2011). More broadly, increasing air temperatures due to climate change (IPCC 2013), associated decreases in glaciers (Zemp et al. 2019), decline in annual mountain snowpack (Mote et al. 2005), earlier spring snowmelt (Rauscher et al. 2008; Islam and Déry 2017), and changes in annual distribution of precipitation are all affecting stream flow and temperature (Hartmann et al. 2013). Understanding the landscape processes driving temperature regimes in rivers (Isaak et al. 2010; Mantua et al. 2010; Martins et al. 2011) can inform exposure, and thus risk, of climate warming to freshwater biodiversity such as Pacific salmon (Crozier et al. 2021; FitzGerald et al. 2021).
The impacts on fishes of climate warming in river systems will be influenced by the spatial configuration of aquatic habitats (Fullerton et al. 2017; Steel et al. 2017). River systems, with their branching network structure and potential high connectivity, are particularly interesting thermal landscapes (Isaak et al. 2010; Peterson et al. 2013; Fullerton et al. 2015). While the diverse climate portfolios of each tributary watershed can buffer downstream flow patterns (Moore et al. 2015; Chezik et al. 2017), climatic changes may lead to stream flow and temperatures changes within tributary catchments that may alter the temperature of downstream rivers (Fullerton et al. 2015; Steel et al. 2017). In mountainous watersheds, like many in British Columbia, tributary rivers are commonly fed from glacierized and snow-covered mountains that cool downstream rivers, many of which are major migratory corridors for Pacific salmon (Islam and Déry 2017; Milner et al. 2017). However, the amount these tributaries cool the mainstem may be increasingly compromised by rapid glacier retreat (Clarke et al. 2015) and diminishing persistent spring snowpack (Islam and Déry 2017). The spatial distribution of temperatures in river networks influences the management options for effective conservation of cold-water fishes in a warming world (Ebersole et al. 2020).
Stream temperature is a product of local climate, landscape features, and human land-use (Lisi et al. 2013; Steel et al. 2017). Warmer water temperatures generally rise with increasing air temperatures or watershed size, likely due to the interaction between the size of the watershed and elevation (Caissie 2006). However, these relationships may be modulated by multiple complex processes that operate at different scales to ultimately control stream temperature (Webb et al. 2008; Garner et al. 2014; Lisi et al. 2015). For example, landscape features and characteristics such as glacier cover, snowpack, watershed elevation, and presence of lakes can all influence temperature dynamics. For example, during periods of warm weather, glaciers and persistent snowpack experience a significant amount of melt, contributing cold water to downstream water temperatures (Jansson et al. 2003), which can decouple the air to water temperature relationship. Over longer timescales, persistent declining spring snowpack and retreating glaciers will lead to a decline of glacier meltwater contributions to stream runoff (Bliss et al. 2014; Huss and Hock 2018), a processes that would inevitably increase water temperatures. Additionally, large lakes can stabilize and in some instances warm downstream water temperatures depending on the lake-feeding water source (Dorava and Milner 2000; Schoen et al. 2017). Also, depending on the extent and proximity, human land use, such as forest harvest (Pollock et al. 2009), irrigation from agriculture (Haddeland et al. 2006), and major dams (Angilletta et al. 2008), can also increase stream temperatures and alter flow patterns. Thus, local climate and landscape characteristics of watershed sub-catchments can influence tributary temperatures, and thus control whether tributaries act to cool, or warm, downstream rivers (Bellmore and Baxter 2014; Fullerton et al. 2015).
There are many approaches to quantifying thermal regimes of rivers, ranging from empirical studies that quantify spatio-temporal variability in water temperatures with field measurements from networks of deployed sensors (Isaak et al. 2010), to process-based models that rely on physics and first principles (Paniconi and Putti 2015; Fatichi et al. 2016). At the most basic level, the influence of tributaries on mainstem water temperatures should be a simple mass-balance mixing model, where downstream temperature is the discharge-weighted sum of the upstream temperatures. Yet, predicting the tributary’s discharge and temperature can be challenging in locations with high spatial variation in precipitation, elevation, glaciers, snowpack, and other factors. Furthermore, the daily or seasonal temporal dynamics of water temperatures in different tributaries could be asynchronous with each other and the mainstem, further complicating simple predictions. Thus, studies often deploy temperature loggers to directly quantify the spatial and temporal dynamics of water temperatures in complex and data-limited river systems (Lisi et al. 2013; Fellman et al. 2014), which include important salmon migratory corridors such as the Babine River.
Pacific salmon populations are increasingly adversely impacted by warming water temperatures in rivers throughout North America (Eliason et al. 2011; Martins et al. 2011; Barnett et al. 2020; Crozier et al. 2021) as well as many other stressors (Xanthippe 2005; Walsh et al. 2020). High water temperature can alter the rate of physiological processes of adult migrating Pacific salmon, which depletes their energy reserves increasing mortality rates (Rand et al. 2006; Martins et al. 2011; Zillig et al. 2021). For example, sockeye salmon (O. nerka) start to show signs of reduction in aerobic scope or mortality when water temperatures exceed ~ 18 °C (Naughton et al. 2005; Keefer et al. 2008; Martins et al. 2011; Atlas et al. 2021); however, this value may vary based on population and their local adaptions (Eliason et al. 2011; Martins et al. 2011). While salmon have the capacity for plastic and adaptive responses to environmental change (Crozier and Hutchings 2014), such as by migrating at different times to avoid excessively warm waters (Reed et al. 2011), obtaining a better understanding of the thermal profile of important migratory corridors is a key step in understanding the exposure of salmon to climate warming (Crozier et al. 2021; FitzGerald et al. 2021).
The Babine River, British Columbia, Canada, is an important migratory corridor for steelhead (O. mykiss) and the five eastern Pacific salmon species, but particularly for sockeye salmon that spawn in stream reaches above the Babine Lake, at the river’s headwaters (Gottesfeld and Rabnett 2008). The Babine River is the origin of most sockeye salmon (80–94%) in the Skeena River Watershed and supports commercial, recreational, and cultural fisheries (Gottesfeld and Rabnett 2008). In some years with warmer temperatures and higher flows, it appears that migration by sockeye salmon may be challenged or delayed through this steep river (Stiff et al. 2015). Water temperature at the outlet of Babine Lake draining into Babine River has increased by 1.5 °C since the 1900s (Stiff et al. 2015). Unlike other major salmon migratory rivers such as the Fraser River (Patterson et al. 2007; Martins et al. 2011), there is little understanding of the seasonal and spatial patterns of the thermal regime for the Babine River and its tributaries.
Here we investigated the role of tributaries in controlling water temperatures of an important sockeye salmon migratory river during peak salmon migration period of a climatically “normal” year. More specifically, we quantified monthly and daily temperature profiles along the Babine River mainstem and its major tributaries in 2016 and 2017, determined if tributary rivers entering the Babine River provided cool water inputs, and assessed potential relationships between climate and landscape characteristics of the tributary rivers. We hypothesized that size of tributary watershed would correlate with the change in water temperature between tributary and downstream water temperature. We also hypothesized that the tributaries and Babine River mainstem seasonal temperature patterns would vary, driving asynchronous contributions to downstream temperatures. These analyses provide specific insight into the thermal profile of the mainstem Babine River at the basin and sub-basin level (Torgersen et al. 2012) to help understand climate exposure of culturally and economically important fishes. More generally, this study showcases the diverse impacts that tributaries with different landscape features can have on river temperatures.
Methods
Babine River watershed
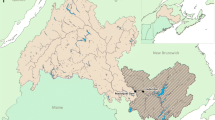
This study was conducted in the Babine River watershed, which drains 10,477 km2 from the Nechako Plateau through the southern Skeena Mountains (Fig. 1). The Babine River watershed is broken into an upper and lower portion, with the upper watershed being situated in the low-relief Nechako Plateau composed mainly of Babine Lake, the longest natural lake in British Columbia, and the tributaries that flow into the lake. The lower watershed, draining from the Babine Lake, consists of the 96-km-long Babine River and other major tributaries, including the Nichyeskwa, Nilkitkwa, Shelagyote, and Shedin drainages (Fig. 1). The elevation at the Babine Lake outlet is ~ 700 m, and ~ 320 m at the Skeena River confluence with a ~ 380 m loss in elevation along the river’s profile. The Babine River is predominantly in a canyon with steep gradients containing falls and rapids rated up to class IV. Major tributaries, some of which originate from high-elevation snow-capped mountains and glaciers, are steep with widely fluctuating water temperatures, flows, and sediment supply (Gottesfeld and Rabnett 2008). There is presently, and has historically been, extensive forestry within the watershed with some areas such as the Nilkitkwa River drainage being more heavily impacted than others (de Groot 2014). In 1999, the Babine River and its 1-km riparian buffer were protected from future development activities by the creation of the Babine River Corridor Provincial Park. However, this protection does not include much of the Babine River watershed area nor its tributaries. The upper part of the Babine River watershed is within Lake Babine Nation territory and the lower portion is within Gitxsan territory.

Babine River watershed and its major tributary watersheds overlaid on a digital elevation model in British Columbia, Canada. Red points represent location of temperature logger sites. Inset map shows temperature logger placement at upstream, within stream and downstream tributary sites. Tributary watersheds identified in bold are those used in our analyses. The Babine River flows northwest from Babine and Nilkitkwa Lake downstream to the Skeena River
The Babine River watershed is a major producer of all five species of eastern Pacific salmon, including Chinook salmon (O. tshawytscha), chum salmon (O. keta), coho salmon (O. kisutch), pink salmon (O. gorbuscha), and sockeye salmon fished by Indigenous, commercial, and recreational fisheries. The Babine River system produces approximately 90% of the sockeye salmon within the Skeena River watershed, with enhancement channels above Babine Lake (Gottesfeld and Rabnett 2008; Cox-Rogers and Spilsted 2012). Sockeye salmon typically migrate up the Babine River mainstem between early July and the end of August (Gottesfeld and Rabnett 2008). The Skeena River downstream of the Babine River is generally cool and has a lower gradient than the Babine River (Stiff et al. 2015); thus, the Babine River represents the portion of the Babine salmon migration that potentially contains the warmest water temperatures and steepest gradient to swim against.
Stream temperature data
We deployed temperature loggers in the Babine River watershed with the goal of understanding how major tributaries influence the Babine River mainstem temperature. Loggers were placed upstream, within, and downstream of major tributaries to capture the relative contribution of each stream to the subsequent downstream temperatures (Marsha et al. 2018). In August/September 2015, we deployed 32 temperature loggers (HOBO Pendants®) upstream, within, and downstream (~ 1 km) of the Nichyeskwa, Nilkitkwa, Hanawald, Shelagyote, Gail, Thomlinson, Shedin, and Shegisic rivers (major tributaries) and along the mainstem of the Babine River (Fig. 1). In 2016, we obtained, downloaded, and redeployed temperature loggers, and added 15 additional loggers to the Babine River mainstem and at Boucher, Cataline, Shanagha, Le Claire, and Sam Green tributaries (Fig. 1). We retrieved all temperature loggers from the Babine River in October 2017. The Babine River is remote and has Class IV whitewater rapids. Thus, we accessed and navigated the length of the Babine River by raft, entering the river at the Babine Lake outlet and exiting at the Skeena River confluence. This project was developed with full recognition of Indigenous Rights, and all research was conducted with permission to access the territories of Lake Babine and Gitxsan Nations.
Stream temperatures were monitored at two-hour intervals from August/September 2015 to mid-October 2017. At each site, we anchored the loggers by attaching cable to large healthy trees on the riverbank beyond predicted extreme river flows levels. The loggers were cased with white PVC shields and attached to the cable protecting them from debris and solar radiation that may bias temperature readings (Isaak and Horan 2011). To protect the loggers from de-watering events or being torn out from debris, we attached weights to the loggers and sunk them in deep pools outside of the rivers thalweg. Upon retrieval and downloading of the data, we manually inspected the data and removed data believed to be associated with de-watering events (e.g., Sowder & Steel 2012), and averaged remaining raw temperature readings to the daily level. All 2017 data were eliminated from our analysis due to a large flood in early May that destroyed 44% of the data loggers via de-watering events, wedging beneath boulders, or being ripped off anchored trees. In addition, Gail River temperature logger data were eliminated from 2016 due to a de-watering event. Our analysis of stream temperature focuses on July 1–August 31, 2016, which is typically the period of warmest annual water temperatures and when most Babine River sockeye salmon populations migrate upstream (Stiff et al. 2015).
We assessed thermal dynamics of the Babine River mainstem and tributaries throughout July and August 2016 (hereafter “summer”) to understand how temperatures changed during the warmest annual period and when adult sockeye salmon typically migrate within the Babine River watershed (Gottesfeld and Rabnett 2008; Stiff et al. 2015). First, we averaged July/August stream temperatures from the 15 sites along the Babine River, and six tributary sites to determine if tributaries that had various watershed sizes and characteristics played a role in cooling the Babine River during peak summer temperature months. Second, we assessed mean daily water temperatures from July 1–August 31, 2016, for each tributary upstream, within stream, and downstream sites to understand how the varying seasonal temperature patterns of the tributaries would affect Babine River mainstem temperatures. Last, we calculated the difference in summer water temperature between upstream and downstream sites, to determine how much each tributary either cooled, warmed, or had little effect on the mainstem Babine River water temperature.
Climate and landscape variables associated with tributary water temperature
Many climate and landscape (hereafter “watershed”) variables are known to potentially contribute to stream temperatures (e.g., Isaak et al. 2010). Using Geographic Information Systems (GIS), we obtained watershed data from several sources. Climate data were calculated using the open-source tool ClimateBC (Wang et al. 2016), which extracts and downscales PRISM climate normal data and extrapolates to any location within British Columbia (Daly et al. 2008). We used climate data averages for each of the six tributary watersheds by sampling at a 1-km resolution across the Babine River watershed, then used ClimateBC to estimate mean summer air temperature and precipitation in 2016, and used previous winter precipitation as snow (Table 1). Landscape variables included glacier coverage obtained from the Randolph Glacier Inventory (v6.0) (RGI Consortium 2017), watershed and lake areas calculated from polygons within the Freshwater Atlas of British Columbia, and elevation values extracted from a 25-m digital elevation model (DEM). We extracted forest harvest data from the National Forest Information System, derived from Landsat images, identifying yearly forest harvest activity at a 30-m resolution. We calculated the total area of forest harvest within each tributary watershed over a 30-year (1985–2015) period. To characterize the contribution of glaciers and forest harvest relative to the size of each tributary watershed, we divided these variables by their watershed area to calculate their proportional coverage.
We used a Pearson’s correlation analysis to identify relationships between watershed characteristics of each tributary, with emphasis on how watershed characteristics were associated with summer stream temperature or the proportional change of water temperature from each tributary to the Babine River mainstem. We were limited in the number of tributaries (n = 6) and therefore did not conduct further statistics. The watersheds and their characteristics are included in our correlation analyses listed in Table 1.
Results
River temperature patterns
Mean summer stream temperature of the Babine River decreased by ~ 2.0 °C from Babine Lake outlet site (15.79 °C ± 0.80 °C) to the Skeena River confluence site (13.76 °C ± 1.13 °C) (Fig. 1). The large and glacierized tributary rivers—Nilkitkwa, Shelagyote, and Shedin—contributed cold water (mean summer temperatures of 12.4 °C, 10.5 °C, and 9.8 °C, respectively), which appeared to reduce summer stream temperature of the Babine River mainstem (Table 1; Fig. 2). The smaller tributary watersheds, Thomlinson and Shegisic, with cold mean summer temperatures of 10.3 °C and 8.2 °C (Table 1), and the warmer Hanawald River (15.0 °C mean summer temperature) appeared to have minimal effect in changing the Babine River mainstem temperatures (Fig. 2). The Babine River mainstem water temperature warmed between the major glacierized watersheds, with summer stream temperature increasing by 1.2 °C between the downstream Nilkitkwa tributary and upstream Shelagyote tributary, and 0.2 °C between the downstream Shelagyote tributary and upstream Shedin tributary (Fig. 2).

Mean summer temperature (°C) ± one standard deviation (shown in gray shading) from temperature sites located along the Babine River, from Babine Lake (distance = 0) to the Skeena River (distance = 96). Blue points represent mean summer temperature (°C) for each tributary that feed into the Babine River, scaled to watershed size. Solid gray line represents the interpolation between point estimates obtained from temperature loggers. Note: Hanawald Creek drains from a small watershed with no glaciers, and Shedin and Shegisic are in close proximity and therefore share the same upstream and downstream temperature loggers
Seasonal dynamics of temperatures in the different tributaries and mainstem were surprisingly similar. Apart from Hanawald, all tributary sites had mean daily stream temperatures colder than the Babine River mainstem (upstream and downstream) sites between July 1 and August 31, in 2016 (Fig. 3). Throughout the summer months, mean daily temperatures of the Nilkitkwa, Shelagyote, and Shedin within-stream temperature sites appeared to reduce Babine River downstream mean daily temperatures (Fig. 3a, c, and e). Meanwhile the Hanawald, Thomlinson, and Shegisic within-stream temperature sites had little to no effect on the Babine River downstream sites. All sites had the warmest mean daily temperature on July 19, 2016, with the peak temperature varying by site. For each within-stream temperature site, the warmest mean daily temperature was 17.7 °C for Nilkitkwa, 18.9 °C for Hanawald, 17.6 °C for Shelagyote, 16.7 °C for Thomlinson, 17.0 °C for Shedin, and 16.2 °C for Shegisic. The warmest mean daily water temperature within the Babine River mainstem was 17.7 °C on July 20, 2016. Thus, counter to predictions, seasonal patterns of water temperatures were generally coherent among tributaries and the mainstem.

Mean daily temperatures (°C) ± one standard deviation (shown in gray shading) for upstream, within stream, and downstream tributary sites from July 1–August 31, 2016. (a) Nilkitkwa (b) Hanawald (c) Shelagyote (d) Thomlinson (e) Shedin (f) Shegisic
The contribution of stream temperature from each tributary site to the Babine River was different, with cooler and more glacierized rivers causing bigger differences in water temperatures upstream and downstream. The temperature difference between upstream and downstream at the Nilkitkwa, Shelagyote, and Shedin tributary sites was − 1.5 °C, − 1.0 °C, and − 0.7 °C, respectively (Fig. 4). The temperature difference at Hanawald was positive (0.6 °C), with the difference at Thomlinson (− 0.1 °C) and Shegisic (− 0.2 °C) tributaries being trivial. Thus, stream temperatures from large glacierized watersheds appear to be the major contributors to temperature changes within the Babine River mainstem.

Map showing the temperature (°C) change between upstream and downstream sites at each tributary site. Blue shading represents a cooling of the Babine River mainstem temperature, red represents a warming of the Babine River mainstem, and yellow represents a minimal change. Red points represent location of temperature logger sites
Watershed characteristics associated with tributary stream temperature
There were strong relationships between some of the six watershed characteristics, tributary temperatures, and temperature difference upstream to downstream (Fig. 5). Mean summer water temperatures were inversely related to previous winter precipitation as snow (r = − 0.93; p = 0.01) and percent glacier cover (r = − 0.87; p = 0.02). Not surprisingly, these two variables were strongly correlated with each other (r = 0.9, p = 0.02). Mean summer water temperature had a positive association with mean elevation (r = 0.76), mean summer air temperature (r = 0.74), and lake area (r = 0.67), although insignificant (p > 0.05; Figs. 5 and 6). There was no relationship between mean summer water temperature and summer precipitation (r = − 0.15), watershed area (r = 0.13), and percent harvest (30 years) (r = 0.24). Water temperature difference upstream to downstream had a strong positive association with watershed area (r = − 0.89; p = 0.02). Thus, watershed variables vulnerable to the effects of climate change that have the strongest effect on mean summer stream temperature are previous winter precipitation as snow or percent glacier and mean summer air temperature. Water temperature difference between upstream and downstream sites is greatest in large watersheds.

Correlations between watershed landscape characteristics. Black box represents correlations between either mean summer water temperature or temperature difference upstream to downstream and watershed characteristics. Values represent Pearson’s correlation coefficients, with higher values shown by increasingly intense colors; blue indicates negative correlations and orange, positive. Asterisks represent statistically significant correlations. N = 6 throughout

Relationship between watershed characteristics and summer stream temperature for the six tributaries. Tributary sites are listed and colored by percent glacier, with values indicated in parenthesis. The x-scales vary depending on the watershed characteristic. Regression lines (in gray) are for visual purposes only
Discussion
We assessed the diverse role of tributaries in cooling a major salmon migration corridor. While water temperatures often increase as watershed size increases due to cumulative thermal loading (Caissie 2006; Lisi et al. 2013), here we showcase how large tributaries that drain regions with glaciers and snow can decrease downstream mainstem river temperatures. Particularly, tributaries with more snowpack throughout the previous winter and glaciers in their catchment had cooler water temperatures. These glacier- and snowpack-fed tributaries cooled the Babine River by approximately 2 °C over its 96 km length. Specifically, the large and cold Nilkitkwa, Shelagyote, and Shedin rivers, with mean summer water temperatures of 12.4 °C, 10.5 °C, and 9.8 °C, respectively, provided the most cooling to the Babine River mainstem, and reduced the Babine River temperature by − 1.5 °C, − 1.0 °C, and − 0.71 °C, respectively. Other studies have also found that glaciers have a cooling effect on summer stream temperature, where water temperatures are reported to decrease by ~ 1 °C for every 10% increase in glacier cover (Moore 2006; Fellman et al. 2014). Meanwhile, the Thomlinson and Shegisic rivers, which are also cold and glacierized, contributed very little to cooling the Babine River mainstem temperatures. This lack of cooling is presumably due to their smaller watershed size, and thus smaller flow output. Indeed, as expected, there was a strong positive correlation between watershed area and the temperature difference upstream to downstream. Surprisingly, seasonal temperature dynamics across sites, both mainstem and tributaries, were synchronous. Regardless, cold meltwater from mountainous tributaries is currently decreasing thermal stress to migrating adult Pacific salmon, which agrees with previous work on thermal riverscapes in mountainous regions (Fellman et al. 2014).
The Babine River is currently on the edge of posing thermal challenges to adult migrating sockeye salmon. The Babine River summer water temperatures are approaching those that could potentially harm sockeye salmon, depending on the thermal tolerance of the Babine River sockeye salmon population. For example, sockeye salmon survival declined when water temperatures exceeded 15 °C in the Koeye River, a small watershed on the central coastal of British Columbia (Atlas et al. 2021), whereas some sockeye salmon populations in the Fraser River appear to be adapted to tolerate warmer water temperatures (Eliason et al. 2011). While there are no population-specific studies of Babine River sockeye thermal tolerance, there is some evidence suggesting that when water temperatures near the Babine Lake outlet exceed ~ 18 °C, sockeye salmon delay migration and stay in cooler waters downstream (Stiff et al. 2015). Thus, while the Babine River may be on the edge of suitable water temperatures, it appears that cold water inputs from major tributaries currently play a key role in maintaining suitable water temperatures for migratory salmon.
The longitudinal thermal profile of rivers can be complex (Fullerton et al. 2015), but all migratory Pacific salmon accessing headwater streams and lakes for spawning must transit through the entire thermal profile of the Babine River at some point in time. In the Babine River watershed, we discovered that the upstream portion is warmer than the downstream portion. This downstream portion is also the location of waterfalls, canyons, and steep sections that could present salmon migratory challenges (Gottesfeld and Rabnett 2008). Maximum aerobic and anaerobic metabolism are required to ascend through difficult portions of upstream migration for populations of sockeye salmon (Eliason and Farrell 2016), and sockeye salmon have been observed recovering in pools or back eddies, sometimes fed from cold upwelling groundwater (Ebersole et al. 2001), while migrating through challenging river sections (Brett 1995). In addition, lower sections of the Babine River have been reported to act as a temperature refugium where migrating sockeye salmon delay migration until waters cool down before continuing their migration (Stiff et al. 2015). Thus, the major tributaries of the Babine River appear essential in cooling the more challenging lower sections, providing more suitable conditions for migrating sockeye salmon. However, the cold tributaries do not cool the entire river, such as the upper portion, which is still required for transit to spawning grounds.
Like rivers across western North American and beyond (Isaak et al. 2012; Luce et al. 2014), Babine River water temperatures will likely continue to warm. Glacier- and snow-fed rivers maintain cool water temperatures in the Babine River, and projections show a 70% decline of glaciers in British Columbia over the next century (Clarke et al. 2015), and 20% decline in winter snowpack levels by 2050 (Islam and Déry 2017), thus impacting the cold water runoff from snow and ice. In addition, the Babine Lake surface water temperatures has increased in recent years (Stiff et al. 2015), and forest harvest has continued to rise (de Groot 2014). Thus, warming temperatures are a potentially pending problem for Babine River and its migratory sockeye salmon. However, other factors can also influence salmon migration and juvenile rearing success, which typically interact with water temperature, but include stream runoff, physical habitat, physiological adaptions, biotic interactions, and food quality and quantity (Martins et al. 2012).
Understanding the spatial and temporal dynamics of water temperatures in riverscapes can help inform management options in a warming world (Ebersole et al. 2020). While climate change requires a global effort in reducing greenhouse gas emissions (IPCC 2013), there are options for localized management actions to try to mitigate climate impacts (Johnson and Jones 2000; Pollock et al. 2009; Ebersole et al. 2020). While we acknowledge that we were limited in our scope to assess the Babine River over multiple years, and that we were limited in sample size representing different tributary rivers (N = 6), here we showcase that on the Babine River, tributaries do measurably cool a major salmon migratory corridor. Land use in the major tributary catchments will influence their ongoing role in cooling the downstream river. For example, forest harvest increases stream temperatures (Mellina et al. 2002; Pollock et al. 2009), and some of these catchments have already been substantially harvested (e.g., Nilkitkwa River watershed, 5.9% forest harvest). While a 1-km buffer surrounding the Babine River mainstem is protected from future development, this does not protect these key tributaries and the cooling function they provide. Additionally, we do note that our findings did not indicate a correlative relationship between forest harvest and mean summer water temperature, not surprising given the sample size of our analysis. We suggest further research on the impacts forest harvest is having on major important tributary water temperatures, such as the Nilkitkwa River. Further restriction of forest harvest within important tributaries that cool downstream Babine River sections would likely help maintain the cold water during upstream migration. Spatial heterogeneity in river temperatures, whether it is small-scale groundwater seepages or tributaries that act as cold-water refuges or the cooling inputs of major tributaries, plays a key role in the persistence and vulnerability of cold-water fishes (Monk et al. 2013; Ebersole et al. 2020).
Data availability
Open datasets used are referenced throughout the manuscript. All other data, material, and code are available upon request.
References
Angilletta MJ, Steel EA, Bartz KK, Kingsolver JG, Scheuerell MD, Beckman BR, Crozier LG (2008) Big dams and salmon evolution: changes in thermal regimes and their potential evolutionary consequences. Evol Appl 1:286–299. https://doi.org/10.1111/j.1752-4571.2008.00032.x
Atlas WI, Seitz KA, Jorgenson JWN, Millard-Martin B, Housty WG, Ramos-Espinoza D, Burnett NJ, Reid M, Moore JW (2021) Thermal sensitivity and flow-mediated migratory delays drive climate risk for coastal sockeye salmon. Facets 71–89. https://doi.org/10.1139/facets-2020-0027
Barnett HK, Quinn TP, Bhuthimethee M, Winton JR (2020) Increased prespawning mortality threatens an integrated natural- and hatchery-origin sockeye salmon population in the Lake Washington Basin. Fish Res 227:105527. https://doi.org/10.1016/j.fishres.2020.105527
Bellmore JR, Baxter CV (2014) Effects of geomorphic process domains on river ecosystems: a comparison of floodplain and confined valley segments. River Res Appl 30:617–630. https://doi.org/10.1002/rra.2672
Bliss A, Hock R, Radić V (2014) Global response of glacier runoff to twenty-first century climate change. J Geophys Res Earth Surf 119:717–730. https://doi.org/10.1002/2013JF002931
Brett JR (1995) Energetics. In: Margolis GC, Clarke WC (eds) Physiological Ecology of Pacific Salmon. UBC Press, Vancouver, BC, pp 1–68
Caissie D (2006) The thermal regime of rivers: a review. Freshw Biol 51:1389–1406. https://doi.org/10.1111/j.1365-2427.2006.01597.x
Chezik KA, Anderson SC, Moore JW (2017) River networks dampen long-term hydrological signals of climate change. Geophys Res Lett 44:7256–7264. https://doi.org/10.1002/2017GL074376
Clarke GKC, Jarosch AH, Anslow FS, Radić V, Menounos B (2015) Projected deglaciation of western Canada in the twenty-first century. Nat Geosci 8:372–377. https://doi.org/10.1038/ngeo2407
Cox-Rogers S, Spilsted B (2012) Update Assessment of Sockeye Salmon Production from Babine Lake, British Columbia. Can Tech Rep Fish Aquat Sci 2956
Crozier LG, Burke BJ, Chasco BE, Widener DL, Zabel RW (2021) Climate change threatens Chinook salmon throughout their life cycle. Commun Biol 4:222. https://doi.org/10.1038/s42003-021-01734-w
Crozier LG, Hutchings JA (2014) Plastic and evolutionary responses to climate change in fish. Evol Appl 7:68–87. https://doi.org/10.1111/eva.12135
Daly C, Halbleib M, Smith JI, Gibson WP, Doggett MK, Taylor GH, Curtis J, Pasteris PP (2008) Physiographically sensitive mapping of climatological temperature and precipitation across the conterminous United States. Int J Climatol 28:2031–2064. https://doi.org/10.1002/joc.1688
de Groot A (2014) Babine Watershed: indicator data summary report. Smithers, BC, Babine Watershed Monitoring Trust
Dorava JM, Milner AM (2000) Role of lake regulation on glacier-fed rivers in enhancing salmon productivity: the Cook Inlet watershed, south-central Alaska, USA. Hydrol Process 14:3149–3159
Ebersole JL, Liss WJ, Frissell CA (2001) Relationship between stream temperature, thermal refugia and rainbow trout (Oncorhynchus mykiss) abundance in arid-land streams in the northwestern United States. Ecol Freshw Fish 10:1–10. https://doi.org/10.1034/j.1600-0633.2001.100101.x
Ebersole JL, Quiñones RM, Clements S, Letcher BH (2020) Managing climate refugia for freshwater fishes under an expanding human footprint. Front Ecol Environ 18:271–280. https://doi.org/10.1002/fee.2206
Eliason EJ, Clark TD, Hague MJ, Hanson LM, Gallagher ZS, Jeffries KM, Gale MK, Patterson DA, Hinch SG, Farrell AP (2011) Differences in thermal tolerance among sockeye salmon populations. Science 332:109–112. https://doi.org/10.1126/science.1199158
Eliason EJ, Farrell AP (2016) Oxygen uptake in Pacific salmon Oncorhynchus spp.: when ecology and physiology meet. J Fish Biol 88:359–388. https://doi.org/10.1111/jfb.12790
Fatichi S, Vivoni ER, Ogden FL, Ivanov VY, Mirus B, Gochis D, Downer CW, Camporese M, Davison JH, Ebel B, Jones N, Kim J, Mascaro G, Niswonger R, Restrepo P, Rigon R, Shen C, Sulis M, Tarboton D (2016) An overview of current applications, challenges, and future trends in distributed process-based models in hydrology. J Hydrol 537:45–60. https://doi.org/10.1016/j.jhydrol.2016.03.026
Fellman JB, Nagorski S, Pyare S, Vermilyea AW, Scott D, Hood E (2014) Stream temperature response to variable glacier coverage in coastal watersheds of southeast Alaska. Hydrol Process 28:2062–2073. https://doi.org/10.1002/hyp.9742
FitzGerald AM, John SN, Apgar TM, Mantua NJ, Martin BT (2021) Quantifying thermal exposure for migratory riverine species: phenology of Chinook salmon populations predicts thermal stress. Glob Change Biol 27:536–549. https://doi.org/10.1111/gcb.15450
Fullerton AH, Burke BJ, Lawler JJ, Torgersen CE, Ebersole JL, Leibowitz SG (2017) Simulated juvenile salmon growth and phenology respond to altered thermal regimes and stream network shape. Ecosphere 8:e02052. https://doi.org/10.1002/ecs2.2052
Fullerton AH, Torgersen CE, Lawler JJ, Faux RN, Steel EA, Beechie TJ, Ebersole JL, Leibowitz SG (2015) Rethinking the longitudinal stream temperature paradigm: region-wide comparison of thermal infrared imagery reveals unexpected complexity of river temperatures. Hydrol Process 29:4719–4737. https://doi.org/10.1002/hyp.10506
Garner G, Hannah DM, Sadler JP, Orr HG (2014) River temperature regimes of England and Wales: spatial patterns, inter-annual variability and climatic sensitivity. Hydrol Process 28:5583–5598. https://doi.org/10.1002/hyp.9992
Gottesfeld AS, Rabnett KA (2008) Skeena River: fish and their habitat. Portland, USA, Ecotrust
Haddeland I, Lettenmaier DP, Skaugen T (2006) Effects of irrigation on the water and energy balances of the Colorado and Mekong river basins. J Hydrol 324:210–223. https://doi.org/10.1016/j.jhydrol.2005.09.028
Hartmann DL, Kelin Tank AMG, Rusticucci M, Alexander LV, Brömmimann S, Charabi Y, Dentener F, Dlugokencky EJ, Easterling DR, Kaplan A, Soden BJ, Thorne PW, Wild M, Zhai PM (2013) Observations: atmosphere and surface. In: Stocker TF, Qin D, Plattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM (eds) Climate Change 2013: The Physical Science Basis. Contribution of Working Group 1 to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, United Kingdom and New York, NY, USA, pp 159–254
Huss M, Hock R (2018) Global-scale hydrological response to future glacier mass loss. Nat Clim Change 8:135–140. https://doi.org/10.1038/s41558-017-0049-x
IPCC (2013) Climate Change 2013: the physical science basis. Contribution of working group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA
Isaak DJ, Horan DL (2011) An evaluation of underwater epoxies to permanently install temperature sensors in mountain streams. North Am J Fish Manag 31:134–137. https://doi.org/10.1080/02755947.2011.561172
Isaak DJ, Luce CH, Rieman BE, Nagel DE, Peterson EE, Horan DL, Parkes S, Chandler GL (2010) Effects of climate change and wildfire on stream temperatures and salmonid thermal habitat in a mountain river network. Ecol Appl 20:1350–1371
Isaak DJ, Wollrab S, Horan D, Chandler G (2012) Climate change effects on stream and river temperatures across the northwest U.S. from 1980–2009 and implications for salmonid fishes. Clim Change 113:499–524. https://doi.org/10.1007/s10584-011-0326-z
Islam SU, Déry SJ (2017) Future climate change impacts on snow and water resources of the Fraser River basin, British Columbia. J Hydrometeorol 18:473–496. https://doi.org/10.1175/JHM-D-16-0012.1
Jansson P, Hock R, Schneider T (2003) The concept of glacier storage: a review. J Hydrol 282:116–129. https://doi.org/10.1016/S0022-1694(03)00258-0
Johnson SL, Jones JA (2000) Stream temperature responses to forest harvest and debris flows in western Cascades, Oregon. Can J Fish Aquat Sci 57:30–39
Keefer ML, Peery CA, Heinrich MJ (2008) Temperature-mediated en route migration mortality and travel rates of endangered Snake River sockeye salmon. Ecol Freshw Fish 17:136–145. https://doi.org/10.1111/j.1600-0633.2007.00267.x
Lisi PJ, Schindler DE, Bentley KT, Pess GR (2013) Association between geomorphic attributes of watersheds, water temperature, and salmon spawn timing in Alaskan streams. Geomorphology 185:78–86. https://doi.org/10.1016/j.geomorph.2012.12.013
Lisi PJ, Schindler DE, Cline TJ, Scheuerell MD, Walsh PB (2015) Watershed geomorphology and snowmelt control stream thermal sensitivity to air temperature. Geophys Res Lett 42:3380–3388. https://doi.org/10.1002/2015GL064083
Luce C, Staab B, Kramer M, Wenger S, Isaak D, McConnell C (2014) Sensitivity of summer stream temperatures to climate variability in the Pacific Northwest. Water Resour Res 50:3428–3443. https://doi.org/10.1002/2013WR014329
Mantua NJ, Tohver I, Hamlet A (2010) Climate change impacts on streamflow extremes and summertime stream temperature and their possible consequences for freshwater salmon habitat in Washington State. Clim Chang 102:187–223. https://doi.org/10.1007/s10584-010-9845-2
Marsha A, Steel EA, Fullerton AH, Sowder C (2018) Monitoring riverine thermal regimes on stream networks: Insights into spatial sampling designs from the Snoqualmie River, WA. Ecol Ind 84:11–26. https://doi.org/10.1016/j.ecolind.2017.08.028
Martins EG, Hinch SG, Cooke SJ, Patterson DA (2012) Climate effects on growth, phenology, and survival of sockeye salmon (Oncorhynchus nerka): a synthesis of the current state of knowledge and future research directions. Rev Fish Biol Fisheries 22:887–914. https://doi.org/10.1007/s11160-012-9271-9
Martins EG, Hinch SG, Patterson DA, Hague MJ, Cooke SJ, Miller KM, Lapointe MF, English KK, Farrell AP (2011) Effects of river temperature and climate warming on stock-specific survival of adult migrating Fraser River sockeye salmon (Oncorhynchus nerka). Glob Change Biol 17:99–114. https://doi.org/10.1111/j.1365-2486.2010.02241.x
Mellina E, Moore RD, Hinch SG, Macdonald JS, Pearson G (2002) Stream temperature responses to clearcut logging in British Columbia: the moderating influences of groundwater and headwater lakes. Can J Fish Aquat Sci 59:1886–1900. https://doi.org/10.1139/f02-158
Milner AM, Khamis K, Battin TJ, Brittain JE, Barrand NE, Füreder L, Cauvy-Fraunie S, Gíslason GM, Jacobsen D, Hannah DM, Hodson AJ, Hood E, Lencioni V, Ólafsson JS, Robinson CT, Tranter M, Brown LE (2017) Glacier shrinkage driving global changes in downstream systems. Proc Natl Acad Sci 114:9770–9778
Monk WA, Wilbur NM, Allen Curry R, Gagnon R, Faux RN (2013) Linking landscape variables to cold water refugia in rivers. J Environ Manage 118:170–176. https://doi.org/10.1016/j.jenvman.2012.12.024
Moore JW, Beakes MP, Nesbitt HK, Yeakel JD, Patterson DA, Thompson LA, Phillis CC, Braun DC, Favaro C, Scott D, Carr-Harris C, Atlas WI (2015) Emergent stability in a large, free-flowing watershed. Ecology 96:340–347
Moore RD (2006) Stream temperature patterns in British Columbia, Canada, based on routine spot measurements. Can Water Resour J 31:41–56
Mote PW, Hamlet AF, Clark MP, Lettenmaier DP (2005) Declining mountain snowpack in western North America. Bull Am Meteor Soc 86:39–50
Naughton GP, Caudill CC, Keefer ML, Bjornn TC, Stuehrenberg LC, Peery CA (2005) Late-season mortality during migration of radio-tagged adult sockeye salmon (Oncorhynchus nerka) in the Columbia River. Can J Fish Aquat Sci 62:18
Paniconi C, Putti M (2015) Physically based modeling in catchment hydrology at 50: survey and outlook. Water Resour Res 51:7090–7129. https://doi.org/10.1002/2015WR017780
Patterson DA, Macdonald JS, Skibo KM, Barnes D, Guthrie I, Hills J (2007) Reconstructing the thermal history for the lower Fraser River, 1941 to 2006, and implications for adult sockeye salmon (Oncorhynchus nerka) spawning migration. Can Tech Rep Fish Aquat Sci
Peterson EE, Ver Hoef JM, Isaak DJ, Falke JA, Fortin M-J, Jordan CE, McNyset K, Monestiez P, Ruesch AS, Sengupta A, Som N, Steel EA, Theobald DM, Torgersen CE, Wenger SJ (2013) Modelling dendritic ecological networks in space: an integrated network perspective. Ecol Lett 16:707–719. https://doi.org/10.1111/ele.12084
Pollock MM, Beechie TJ, Liermann M, Bigley RE (2009) Stream temperature relationships to forest harvest in western Washington. J Am Water Resour Assoc 45:141–156
Rand PS, Hinch SG, Morrison J, Foreman MGG, MacNutt MJ, Macdonald JS, Healey MC, Farrell AP, Higgs DA (2006) Effects of river discharge, temperature, and future climates on energetics and mortality of adult migrating Fraser River sockeye salmon. Trans Am Fish Soc 135:655–667. https://doi.org/10.1577/T05-023.1
Rauscher SA, Pal JS, Diffenbaugh NS, Benedetti MM (2008) Future changes in snowmelt-driven runoff timing over the western US. Geophys Res Lett 35:L16703. https://doi.org/10.1029/2008GL034424
Reed TE, Schindler DE, Hague MJ, Patterson DA, Meir E, Waples RS, Hinch SG (2011) Time to Evolve? Potential evolutionary responses of Fraser River sockeye salmon to climate change and effects on persistence. PLoS ONE 6:e20380. https://doi.org/10.1371/journal.pone.0020380
RGI Consortium (2017) Randolph Glacier Inventory - a dataset of global glacier outlines: version 6.0. Colorado, USA
Schindler DW (2001) The cumulative effects of climate warming and other human stresses on Canadian freshwaters in the new millennium. Can J Fish Aquat Sci 58:18–29
Schoen ER, Wipfli MS, Trammell EJ, Rinella DJ, Floyd AL, Grunblatt J, McCarthy MD, Meyer BE, Morton JM, Powell JE, Prakash A, Reimer MN, Stuefer SL, Toniolo H, Wells BM, Witmer FDW (2017) Future of Pacific salmon in the face of environmental change: lessons from one of the world’s remaining productive salmon regions. Fisheries 42:538–553. https://doi.org/10.1080/03632415.2017.1374251
Sowder C, Steel EA (2012) A note on the collection and cleaning of water temperature data. Water 4:597–606. https://doi.org/10.3390/w4030597
Steel EA, Beechie TJ, Torgersen CE, Fullerton AH (2017) Envisioning, quantifying, and managing thermal regimes on river networks. Bioscience 67:506–522. https://doi.org/10.1093/biosci/bix047
Stiff HW, Hyatt KD, Hall P, Finnegan B, Macintyre D (2015) Water temperature, river discharge, and adult sockeye salmon migration observations in the Babine Watershed, 1946–2014. Can Man Rep Fish Aquat Sci 3053
Strayer DL, Dudgeon D (2010) Freshwater biodiversity conservation: recent progress and future challenges. J N Am Benthol Soc 29:344–358. https://doi.org/10.1899/08-171.1
Torgersen CE, Ebersole JL, Keenan DM (2012) Primer for Identifying Cold-Water Refuges to Protect and Restore Thermal Diversity in Riverine Landscapes. Seattle, WA
Walsh JC, Connors K, Hertz E, Kehoe L, Martin TG, Connors B, Bradford MJ, Freshwater C, Frid A, Halverson J, Moore JW, Price MHH, Reynolds JD (2020) Prioritizing conservation actions for Pacific salmon in Canada. J Appl Ecol 57:1688–1699. https://doi.org/10.1111/1365-2664.13646
Wang T, Hamann A, Spittlehouse D, Carroll C (2016) Locally downscaled and spatially customizable climate data for historical and future periods for North America. PLoS ONE 11:e0156720. https://doi.org/10.1371/journal.pone.0156720
Webb BW, Hannah DM, Moore RD, Brown LE, Nobilis F (2008) Recent advances in stream and river temperature research. Hydrol Process 22:902–918. https://doi.org/10.1002/hyp.6994
Xanthippe A (2005) Atlas of Pacific Salmon: the first map-based status assessment of salmon in the North Pacific
Zemp M, Huss M, Thibert E, Eckert N, McNabb R, Huber J, Barandun M, Machguth H, Nussbaumer SU, Gärtner-Roer I, Thomson L, Paul F, Maussion F, Kutuzov S, Cogley JG (2019) Global glacier mass changes and their contributions to sea-level rise from 1961 to 2016. Nature 568:382–386. https://doi.org/10.1038/s41586-019-1071-0
Zillig KW, Lusardi RA, Moyle PB, Fangue NA (2021) One size does not fit all: variation in thermal eco-physiology among Pacific salmonids. Rev Fish Biol Fisheries 31:95–114. https://doi.org/10.1007/s11160-020-09632-w
Acknowledgements
We gratefully acknowledge funding from the Liber Ero Foundation, which provided the opportunity to conduct research on the remote Babine River, British Columbia. We are grateful for Skeena Watershed Conservation Coalition, Alfred Schaefer, Morag Gibb, Todd James, Ben Pittman, Ciara Sharpe, and Ali Howard for safely navigating us down the river by raft or kayak. We are grateful for Babine Norlakes Steelhead camp and Silver Hilton Steelhead lodges for assisting in logger placement and retrieval. Thank you to Kenny Rabnett for providing endless knowledge of the Skeena River watershed. Kara Pitman was supported by National Science and Engineering Research Council and Association of Canadian Universities for Northern Studies. We are extremely grateful for Gitxsan and Lake Babine Nations for permitting us to conduct research within their traditional territories.
Funding
This work was financially supported by the Liber Ero Foundation funding agency. Kara J. Pitman was supported by National Science and Engineering Research Council and Association of Canadian Universities for Northern Studies.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection, and analysis were performed by Kara J. Pitman. The first draft of the manuscript was written by Kara J. Pitman, and Jonathan W. Moore commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval
This research does not involve human participants and/or animals; therefore, no ethics approval was needed.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Pitman, K.J., Moore, J.W. The role of large, glaciated tributaries in cooling an important Pacific salmon migration corridor: a study of the Babine River. Environ Biol Fish 104, 1263–1277 (2021). https://doi.org/10.1007/s10641-021-01152-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-021-01152-1