
Abstract
Many tropical fisheries are data-poor and lack population demographic information needed for effective management and conservation. In this study we used mark-recapture of bonefish, Albula vulpes, an important species in catch-and-release recreational fisheries, to estimate capture probabilities. Moreover, for the first time we generated key demographic parameters including apparent survival, new entries and population size. We marked 9657 bonefish and recaptured 605 (6.3 % recapture rate) inside and outside protected areas in northern Belize and southern Mexico. We built 20 open population model types known as POPAN in program MARK. The model with a constant superpopulation and probability, and a time-dependent survival and capture probability was best supported by our data. A potentially stable superpopulation size of bonefish > 22 cm of approximately 197,350 individuals (SE = 16,010, lower bound = 168,382, upper bound = 231,302) inhabited a larger region beyond our sampled (40.8 km2 sample area). A combination of permanent and temporary immigration and emigration patterns resulted in seasonal variations in survival, capture probabilities, probability of entry of individuals and population size (or abundance). Approximately 188,000 adult bonefish migrate and congregate in near-shore pre-spawning aggregation sites of the Caribbean Sea near Belize and Mexico during the spawning season. Population stability is likely associated with bonefish protections enacted in 1977, protected areas, and conservation practices by fishing communities of Belize and Mexico. This highlights the importance of protected areas and interjurisdictional fisheries management and suggests the need for a paradigm shift in the Caribbean to include connectivity of habitats essential to all life stages for important fish species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Bonefish (Albula vulpes) is an ecologically, economically, and culturally important species targeted in the catch-and-release (CR) recreational fishery of the western North Atlantic and Caribbean Sea (Adams et al. 2008, 2019a). The species has an important niche as predator (Colton and Alevizon 1983; Danylchuk et al. 2008; Murchie et al. 2019) and prey (Danylchuk et al. 2007a, b; Torres-Chávez et al. 2018) in many coastal ecosystems. Its populations and habitats are important, as they sustain an entire industry that ranges from the manufacturers of fishing equipment, sales and retail, to guided fishing and accommodation services. In the Bahamas, the bonefish recreational fishery generates an annual economic impact of US $169 million (Fedler 2019) while multispecies flats fisheries in other locations generate $465 million in the Florida Keys (Fedler 2013), US $991 million in the Florida Everglades (Fedler 2009), and US $56 million in Belize (Fedler 2014), with an approximate total of US $1.68 billion. However, conservation and management of recreational fisheries continues to be a major challenge (FAO 2009, 2012; Perez-Cobb et al. 2014) due to the lack of data on biological, ecological (Adams 2017; Adams et al. 2019a; Pickett et al. 2020) and population dynamic characteristics (Ault et al. 2008; Larkin 2011; Filous et al. 2019; Perez et al. 2020) to inform decision making.
Biotic and abiotic density-dependent and density-independent factors affect survival, reproduction, distribution of organisms, population size and movement patterns in populations (Begon et al. 2006). Movement in particular is an almost universal behavioral characteristic that affects population dynamics and communities (Dingle 2014) and allows for habitat and ecosystem connectivity (Mumby 2005; Sheaves 2009; Perez et al. 2019b). These occur because movements are also affected by biotic and environmental factors (Begon et al. 2006; Binder et al. 2011; Dingle 2014; Acolas and Lambert 2016; Couto et al. 2016; Thurow 2016). Understanding changes in population size and population dynamics is important as these population characteristics are not often integrated into the present conservation and management systems (FAO 2009, 2012; Perez-Cobb et al. 2014). Assessing how these parameters are affected by behavioral dynamics is important to predict resiliency and achieve sustainability, but this can be difficult as they are often affected by human activities (Danylchuk et al. 2007a; Arlinghaus et al. 2013).
Bonefish was heavily exploited by artisanal subsistence and commercial (ASC) fisheries in the 1960s in Caribbean countries, such as Belize and Mexico, due to its predictable schooling behaviors associated with local movements and migration. Bonefish was first protected as a non-commercial species on December 31, 1977 in Belize (Government of Belize 2003), and further protected along with permit (Trachinotus falcatus) and Atlantic tarpon (Megalops atlanticus) as CR-only in 2009 (Government of Belize 2009a, b). These protections were used as tools in conservation and management to protect the economically valuable flats fishery. Protected areas using zoning schemes were also created to limit the use of unsustainable fishing gear such as nets (Government of Belize 2003). In Mexico, refuge zones in biosphere reserves and protected areas were created; outside of these zones bonefish remains unprotected, but local communities still practice CR to help ensure a sustainable fishery (Perez-Cobb et al. 2014). These measures, along with declines in commercially oriented fisheries in the Caribbean, prompted fishers to become involved in the CR fishery, making it a livelihood and more sustainable and economically valuable. In Belize, the fishery directly employs nearly 2100 individuals who benefit from nearly US $35 million in wages and salaries (Fedler 2014). Yet, bonefish is still Near Threatened due to ASC harvest, habitat loss and fragmentation caused by coastal development, urbanization and poor water quality (Adams et al. 2013).
The most immediate threat to bonefish, considering bonefish is CR, is habitat loss and degradation from coastal development (Steinberg 2015; Adams and Murchie 2015; Adams et al. 2019a, b; Brownscombe et al. 2019; Perez et al. 2019a; Sweetman et al. 2019), which results in fragmentation and habitat patchiness (Akçakaya 2000). Thus, ensuring bonefish ontogenetic habitat protection and connectivity (Perez et al. 2019b, 2020) of offshore pelagic larvae and benthic neritic juveniles and adults in seagrass, sandy and mangrove coastal habitats are important for a healthy and productive fishery (Murchie et al. 2019). Nonetheless, few studies in the area highlight anthropogenic impacts such as threats to biodiversity (Steinberg 2015) by the decrease in coastal vegetation cover (Sweetman et al. 2019), changes in composition and structure of macrobenthos by runoff of organic matter and pollutants from rivers (González et al. 2009), as well as altered water temperature, organic matter in sediment, oxygen concentrations, vegetation cover, structure of benthic community (Hernández-Arana and Ameneyro-Angeles 2011) and diversity, composition, and abundance of fish species (Schmitter-Soto and Herrera-Pavón 2019) from dredging and artificial canals.
Most biological and population studies on bonefish have been conducted outside the Caribbean and have provided important insights on behavioral dynamics associated with bonefish movements (Murchie et al. 2013; Boucek et al. 2019), pre-spawning and spawning activity (Danylchuk et al. 2011, 2019; Adams et al. 2019b) and on population size and dynamics (Ault et al. 2008; Larkin 2011). Unfortunately, few studies on movement, migration and shared resources have been conducted in Central America (Perez et al. 2019a, b; Perez et al. 2020) and none on population size and dynamics. This lack of biological information is especially disconcerting since local information is necessary to aid decision-making in coastal development and habitat protection, especially for bonefish which has a life history that requires a large habitat mosaic (Adams and Murchie 2015) and a new approach to conservation and management (Perez et al. 2020).
The ASC fisheries and coastal development has increased in the Yucatan Peninsula in the last two decades, leaving uncertainty on the status of bonefish as a shared resource after four decades of protection, conservation and management. To address this uncertainty, we used mark-recapture to evaluate for the first time the bonefish population size and dynamics in the Caribbean Sea and a tropical estuary shared by Belize and Mexico. We used the POPAN system for the analysis of mark-recapture data (Arnason and Schwarz 1999) in MARK to estimate capture probability and key demographic parameters: apparent survival, new entries and population size in open populations (White and Burnham 1999; Pine et al. 2003) to provide a foundational metric for evaluation of conservation effectiveness.
Materials and methods
Study area
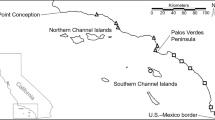
The study was conducted in the Western Caribbean of northern Belize and southern Mexico in the Yucatan Peninsula (Fig. 1). The study area includes a bay known as the Chetumal Bay in Mexico and Corozal Bay (hereafter Chetumal-Corozal Bay) and the adjacent Caribbean coast. These ecosystems encompass one reproductive population for bonefish because there is connectivity between the bay and the Caribbean mediated by spawning migration (Perez et al. 2019b) and thus in this study considered as a single catchment area.

Study in the Western Caribbean of northern Belize and southern Mexico in the Yucatan Peninsula. Map processed by J. Padilla
The region experiences three seasons: cold fronts (hereafter norths) from November to January, dry from February to May, and rainy from June to October [precipitation, wind, and other seasonal influences have been discussed previously (Perez et al. 2019a, b)]. Shallow flats with muddy, sandy, coral rubble and rocky bottoms, often with submerged aquatic vegetation and mangrove-lined creeks, wetlands, and lagoons predominate (Perez et al. 2019b). The western area consists of sources of freshwater during the wet season from sinkholes known as “ojos de agua” (Hernández-Arana and Ameneyro-Angeles 2011), the Hondo River separating Belize and Mexico, and the New River in Belize (Fig. 1). The eastern area is characterized by an artificial canal known as the Zaragoza Canal 50 m wide, 1300 m long and 2.5 m deep, which drastically modified the local hydrology from brackish to marine conditions, presence of hard corals (Hernández-Arana and Ameneyro-Angeles 2011) and salinity ranges of 18 to 40 psu (Perez et al. 2019b). The Caribbean coast is comprised of a backreef lagoon system (Adams et al. 2006), approximately 1 km wide from the shoreline to the reef crest (Perez et al. 2019b), typically between 2 and 3 m deep to a maximum of 6 m (Grimshaw and Paz 2004) and marine salinity ranges 34 to 36 psu (Perez et al. 2019b). The eastern area also has a natural channel known as Bacalar Chico of 30 m width that meanders among mangroves for approximately 3 km length (Hernández-Arana and Ameneyro-Angeles 2011) which forms the international border between Belize and Mexico, and a second natural channel known as Boca del Rio in San Pedro Town. Lastly, the southern region is largely characterized by a wide opening of the bay between San Pedro and the mainland.
The study area included portions of wildlife sanctuaries and marine protected areas (Fig. 1). In northern Belize, Bacalar Chico Marine Reserve and National Park (BCMRNP), Hol Chan Marine Reserve (HCMR) and Corozal Bay Wildlife Sanctuary (CBWS) are part of the Northern Belize Coastal Complex (Sarteneja Alliance for Conservation and Development 2015, 2020). In southern Mexico, the study area included the Manati Sanctuary State Reserve (Santuario de Manatí, MSSR) and Xcalak Reef National Park (Parque Nacional Arrecifes de Xcalak, XRNP), which borders Belize (Schmitter-Soto et al. 2018; Torres-Chávez et al. 2018; Schmitter-Soto and Herrera-Pavón 2019). Both CBWS and MSSR were further on the western side of the bay where sampling was not conducted. Part of HCMR is also within the bay, but the general use zone (Bajos Conservation Zone) consists of five sub-zones for the conservation of flats habitats (Government of Belize 2015) which were close to sampling areas. Most sampling occurred on the eastern side of MSSR in Mexico and Bacalar Chico National Park (BCNP) and general use zone of HCMR. Most of our sampling were inside the protected areas of BCMRNP, HCMR and PNAX (Fig. 1). These areas were designated to preserve and protect multiple species and habitats despite the lack of knowledge on the importance of key habitats to specific species such as bonefish (J. Azueta, pers. comm.).
Sampling
Sampling periods occurred in January, June, November and December 2016; every month of 2017, and February 2018 (Table 1). Sampling focused on sites that harbored bonefish based on local knowledge of guides. Not all sites were sampled during each sampling period. Sites were less than 1.2 m deep, with sturdy sand, rock, or seagrass bottoms. At each site, the search for bonefish occurred by boat propelled approximately 1–2 km/h by poling or motor and about 500 m from the coast, which allowed us to sight bonefish 500 m towards the shoreline and 500 m on the other side of the boat. Bonefish were sighted and then captured using two light-colored seine nets, each 45 m long, 1.2 m high, and with 2.5 cm mesh. The fish were encircled with the seines and then taken out with hand nets and kept in a nearly submerged floating cage (1 m x 0.5 m x 0.25 m). Each bonefish was measured (fork length, FL, to the nearest mm) and tagged with a dart tag (model PDS, Hallprint, Australia) in the left-side musculature, between the first dorsal pterygiophores (Boucek and Adams 2011). Following the tag manufacturers’ advice for the size and type of tag we used, to ensure survival only fish > 22 cm FL were tagged, the rest only measured and counted. At each site we recorded the date, time, latitude and longitude, strata (CC or CB), and tag number of any recaptured fish. Fish were handled for the shortest time possible, allowed to recover in another seine enclosure and then released en masse to reduce post-release mortality from predation (Adams et al. 2009) (for further details see Perez et al. 2019a, b). There are always unidentified and uncontrollable factors that influence animal detection (Mackenzie and Royle 2005; MacKenzie et al. 2006). Thus, our sampling effort was not controlled by the number of haul nets per day or month, but by time: eight consecutive days each month, four days in each country. In some instances, bonefish were so abundant that an entire day was spent marking-recapturing bonefish that had been captured in a few samples, meaning fewer sites were covered. In other instances more sites were sampled as bonefish were less abundant or had lower presence. As has been described in Perez et al. (2019a, b), weather conditions did not reduce sampling days but affected our mark- recapture through the seasons. We made our best effort to sample at least 30 days after each initial sampling period to reduce effects of tag loss (Boucek and Adams 2011). The largest differences in time intervals were between sample periods 1 and 2 (149 days) when mostly new and unmarked animals were captured and then marked.
Model structure and analysis
POPAN is a comprehensive statistical system used to manage and analyze data from mark-recapture experiments (Schwarz and Arnason 1996). There are several POPAN models which are open population model extensions (Arnason 1972, 1973; Schwarz and Arnason 1996; Arnason and Schwarz 1999) of the Jolly-Seber (JS) model (Jolly 1965; Seber 1965). The JS model of mark-recapture experiments has been used to estimate abundance of the population and number of new entries between samples. The general assumptions are that marked animals (i.e. recaptured) and unmarked animals (newly marked/tagged) have the same capture probability in the population (i.e. newly captured unmarked individuals are a random sample of all unmarked animals in the population). The JS model was parameterized in the POPAN formulation (known as POPAN-4) by Schwarz and Arnason (1996) to assume a superpopulation (N) consisting of all animals that would ever be born to the population and the probability that animals from this hypothetical superpopulation would enter the sampled population between sample periods. These two parameters and other parameters are represented below (summarized in Fig. 2):
-
pi the probability of capture at sample period i;
-
φi probability of an animal surviving and remaining in the population between sample periods i and i + 1, given it was alive and in the population at sample i.
-
bi probability of an animal from the superpopulation (\(\widehat{N}\)) would enter the population between periods i and i + 1 and survive to the next sample period i + 1.
-
Bi is the net number of new entrants between sample period i and i + 1 and ∑Bi is the total number of net new entrants.
-
\(\widehat{B}\)i is the gross number of new entrants (birth + immigration) between sample period i and i + 1 and ∑\(\widehat{B}\)i is the total number of gross new entrants.
-
M = ∑mi the total number of marked animals; mi the number of marked animals captured at sample period i.
-
U = ∑ui the total number of unmarked animals; ui the number of unmarked animals captured at sample period i.
-
ni = number of animals captured at sample period i, marked and unmarked (i.e. ni= mi + ui).
-
Ni = Bi + mi is the total number of animals present in the sample population between sample period i and i + 1; and,
-
N = (∑\(\widehat{B}\)I - ∑Bi) + M is the total population size estimate (i.e. superpopulation, which is the total number of animals, observed or unobserved individuals).

Other model assumptions: (1) there are losses on capture at every sample period; (2) fish retain the tags throughout the duration of the experiment, tags are read properly and sample is instantaneous; (3) the study area is constant in size; (4) apparent survival in the marked subset of animals provides information on the remaining unmarked animals in the population at large; (5) apparent survival probabilities are the same for all marked and unmarked fish between sample (homogeneous survival); (6) catchability for marked and unmarked fish is similar and estimated for each sample (i.e. homogeneous catchability); (7) animals leave the population by death or permanent emigration (i.e. apparent survival = 1 – (death + emigration)) or enter as new entries/births/unmarked fish from outside by natural births (fish grow into the catchable portion of the population) or immigration (for spawning); (8) the number of new animals, entrance probabilities and a superpopulation size are equivalent in the modelling process; and (9) in a fully-time dependent model (a) survival and catchability cannot be estimated for the final sample, only between sample periods and (b) entry between first and second period cannot be estimated, only between sample periods.
We labeled all marks and recaptures as 1 and non-encounters as 0. We then obtained an encounter history for each individual. For example, 1,000,000,000,110,000 means that a bonefish was marked in sample period 1, was not encountered from sample periods 2 to 10, recaptured in sample period 11, recaptured again in sample period 12 and not encountered afterwards (sample periods 13–16). Each encounter history was loaded in the program MARK. Time interval between sample periods was estimated by dividing the number of days between an end date (e.g. for sample period 1) and a start date (e.g., sample period 2) by 31 (days) (Table 1). We used Google Earth (www.googleearth.com) to draw polygons over the sampled areas. The polygon lines were approximately 1 km from and along the shoreline. The Google Earth data in kml format were then pasted in Earth Point (http://www.earthpoint.us/Shapes.aspx) to estimate the area and perimeter of the study area.
The POPAN models (Arnason and Schwarz 1999) were built using the sin link function and Run All function in MARK (White and Burnham 1999) to parameterize the Jolly-Seber model (Arnason 1972, 1973; Schwarz and Arnason 1996; Arnason and Schwarz 1999) in terms of a superpopulation: apparent survival (ϕ), capture probability given the animal is alive and on the study area, i.e., available for capture (p), probability of entry into the population (b) and superpopulation size (N). The top four models (Table 2) were then adjusted using the sin link function for ϕ and p, the Multinomial Logit link function or MLogit (1) in MARK for b, and log link function for N. Models were first assessed using Akaike’s Maximum Likelihood (Akaike 1973) and with a Least Regression Test (LRT) built in MARK. Finally, All figures were produced in RStudio Version 1.1.442 (RStudio-Team 2016).
Results
Our sample sites were distributed over approximately 71.6 km of shoreline along the coast of our study area and encompassed approximately 40.8 km2 of flats habitats. A total of 9657 bonefish was marked and 605 individuals were recovered (6.3 % recapture rate). Effective sample size (newly marked and recovered) was 10,272 bonefish (Table 3) meaning some marked bonefish were recaptured multiple times. Size of marked bonefish (average = 30.1 cm, min = 22.0 cm, max = 56.4 cm) recovered bonefish (average = 31.25 cm, min = 23.9 cm, max = 47.0 cm) were nearly similar. Because previous research (Perez et al. 2019a, b) demonstrated that bonefish in the bay and Caribbean were part of the same population, we used single modelling on our data (Arnason and Schwarz 1999; White and Burnham 1999; Pine et al. 2003). Hence, marked and recaptured bonefish in the bay and the Caribbean were not distinguished.
A total of 20 models were run using MARK. First, 16 models were run using the sin function for all parameters. From the top four models (Model parameters in Table 2), Model 1 [ϕ (t) p (t) b (.) N(.)] and Model 2 [ϕ (t) p (t) b (t) N (.)] had a constant N while models 3 and 4 had a time-varying N, higher AICc and lower model likelihood and AICc weight (Table 2). The differences between model 1 and model 2 were in the probability of entry (b) where Model 1 had a constant b and model 2 a time-varying b. The top four models were readjusted using the sin function for ϕ and p, mlogit (1) function for b, and log function for N (Table 2). Nonetheless, the constrained probability of entry to a fully-time dependent model provided 15 estimates for b (estimates in Table 2). An LRT was used on all 4 models and indicated that model 2 was the reduced model from the general model 3 (highest χ2 = -0.028, D.F. = 0, P = < 0.05) because the formulations were simply re-parameterizations. Thus, model 1 with the lowest AIC and a model likelihood of 1 with 47 estimated parameters (ϕ = 15, p = 16, b = 15 and N = 1) was better supported by our data and was also the most parsimonious model: superpopulation as constant and apparent survival, probability of entry and capture probability as fully-time dependent.
We obtained one estimate for the superpopulation (N) of bonefish > 22 cm. Using model 1 we estimated approximately 197,350 individuals (SE = 16,010, lower bound = 168,382, upper bound = 231,302) and using model 2 a similar estimate of 194,126 individuals (SE = 15,564, lower bound = 165,938, upper bound = 227,103). Based on the superpopulation estimate of model 1, during our study we marked 4.9 % and the remaining 95.1 % were unmarked. This estimated superpopulation is interpreted as the estimated total number of bonefish (summation of tagged and untagged) ever present in the spatial extent of the experiment and does not represent the number present at any particular point in time and beyond.
Our estimates reveal a larger sample population size (Ni) during the norths season than the dry season and both seasons were higher than the rainy season (Fig. 3; Table 3). The peak population size (> 40,000) started from sample period 2 (November - early norths season) of 2016 and was relatively high (> 188,000) in December 2016, January 2017 and February 2017 (Table 3; Fig. 3). Most of our sampling during the norths season was in CC as the bay was choppy and water visibility to sight bonefish was unfavorable. There were also peaks in sample period 13 (November 2017) and period 14 (December 2017) with a low estimate on the following period (January 2018) with 1256 bonefish. Both of these peak patterns were followed by steep drops after the norths season and into the dry season. In 2016 cold fronts started in early November and in 2017 and 2018 the cold fronts were in late November to December.

Population size estimates (Ni) or abundance with upper and lower bounds for bonefish > 22 cm in the Western Caribbean of northern Belize and southern Mexico in the Yucatan Peninsula
The probability estimate in the entry of migrant (bonefish > 22 cm) and/or new birth (fish reaching 22 cm FL) was time-dependent because of the mlogit link function specified in the modelling. Both the gross and net new entrant estimates were similar with peaks during the norths season (November and December) of 2016 and 2017 (Figs. 4 and 5) corresponding to a likely migration of ready to spawn adults or new births. There was a high entry estimate (75.9 %) of gross and net entrants of 148,616 bonefish during sample period 2 of November 2016. The major difference was in sample period 11 of the early rainy season of 2017 where gross entrants was 3640 and new entrants 2087. However, entry estimates of sample period 11 (August/September) was 1 % and of sample period 12 (October) 2 %. Both gross and net entrants were lower in sample period 11 than in sample period 12 (Figs. 4 and 5).

Gross entrant estimates of bonefish > 22 cm in the Western Caribbean of northern Belize and southern Mexico in the Yucatan Peninsula

Net entrant estimates of bonefish > 22 cm in the Western Caribbean of northern Belize and southern Mexico in the Yucatan Peninsula. Refer to the text for months and years that correspond to the seasons
The capture probabilities (Fig. 6) and apparent survival were also time-dependent (Fig. 7) and reflected an obvious result with a sampling design of uneven time intervals. Capture probabilities estimates were high during sample period 8 of the dry season (9 %), sample period 10 of the rainy season (10 %) and highest in sample period 15 of norths season (25 %), consistent with the high number of recaptures and the marking of new individuals (i.e. effective sample size in Table 3). The peak in apparent survival was followed by a decline in all seasons: north (2016), dry (2017) and norths (2018). In several sample periods of the norths season of 2016, apparent survival remained at 100 % but fell below 50 % in the dry and rainy season of 2017 with the lowest (11 %) during the dry season of 2017.

Capture probability (p) estimates of bonefish in 16 sample periods (16 estimates) of bonefish > 22 cm in the Western Caribbean of northern Belize and southern Mexico in the Yucatan Peninsula

Apparent survival (ϕ) estimates of bonefish in 16 sample periods (15 estimates) of bonefish > 22 cm in the Western Caribbean of northern Belize and southern Mexico in the Yucatan Peninsula
Discussion
We provide for the first time an estimate of the population size, survival and entry of migrants and new births associated with migration patterns of bonefish in Belize and Mexico. Our study also supports the need to consider the entire region of northern Belize and southern Mexico as a reproductive catchment area necessary to maintain a healthy and resilient bonefish population (Perez et al. 2020). The overall finding was a constant superpopulation size which suggests a stable and potentially resilient population. This is likely a result of good fisheries management and conservation measures adopted and implemented by local communities. Nonetheless, these are all threatened by the emerging unregulated expansion of tourism-related infrastructure and activities (Steinberg 2015).
Although the data on production and landings of bonefish is limited prior to 1977, local knowledge recognizes the bonefish population has recovered (A.P. unpubl. data). Thus, the observed population stability suggests the protection of bonefish in 1977, further protection with catch-and-release status in 2009, establishment of protected areas, and conservation actions by coastal communities likely had a positive effect. The establishment of marine reserves produces an increase in fish density and abundance following declines from fisheries harvest (Friedlander and Parrish 1998; Schmitter-Soto et al. 2018). This is particularly true as marine reserves are net exporters of adults (“spillover effect”) and propagules (“recruitment effect”) (Russ et al. 2004), and it is likely that the stable population observed in this study benefits from these protections. This stability can also be attributed to good conservation practices by the flats fishing communities, whereby guides and anglers have reduced stress by following best handling practices (Danylchuk et al. 2007a; Suski et al. 2007). The protection of sea turtle nesting habitats in BCMR in 1991 (Grimshaw and Paz 2004) serendipitously protected a major pre-spawning aggregation (PSA) site for adult bonefish which was only recently documented (Perez et al. 2019a, b) and is also likely supporting population stability. Reef fish (snappers and groupers) have been mostly documented to form offshore spawning aggregations (Sadovy de Mitcheson et al. 2008), but the formation of PSA aggregations by bonefish requires special considerations as they occur along the coast (subsequent spawning occurs in offshore waters; Danylchuk et al. 2011, Adams et al. 2019a, b). The PSAs are particularly vulnerable to harvest and habitat loss which could reduce the number of reproductive adults and eventually the population size.
Animals enter the study area through immigration or through births from within the study area and can exit the population through death or permanent emigration (Pine et al. 2003; Hightower and Pollock 2013). A combination of these with seasonal variations reflects an apparently stable bonefish population. First, our estimates of abundance or population size were very likely associated to spawning (i.e. bonefish migrating and in pre-spawning schools or ready to spawn). We assume this as most of our sampling during the norths season was in the Caribbean Sea in both countries. As has been largely reported, animals respond to environmental cues to migrate to spawn (Begon et al. 2006; Binder et al. 2011; Dingle 2014; Acolas and Lambert 2016). For bonefish, changes in abiotic conditions produced by the norths season coincide with the bonefish spawning migration (Perez et al. 2019a, b, 2020). Thus, spawning migration is a major factor that produced the observed peak in population size estimates during the first norths season. This seasonal population peak is similar to other species that undergo spawning migrations. For instance, snapper and groupers in the Caribbean have also been documented to have peak months during their spawning season (Heyman et al. 2005; Heyman and Kjerfve 2008). In our case, it seems the bonefish peak spawning month is November based on population size estimates. In contrast, the low population size estimate in the second norths season was likely due to weather reducing the amount of sampling, although fish migrating to a spawning event might also be reflected as emigration/death, thus reducing abundance estimates. The population peaks during the dry and rainy season were likely the result of new births of younger fish (bonefish < 22 cm FL) into the large size fish (> 22 cm) population. This suggests a size-based movement pattern and distribution of bonefish (Perez et al. 2019a, 2020) which deserves additional examination. The steep drops in population size (i.e. loss) after December and November of the norths season and into the dry season reflected the end of the spawning season and return of the seasonal spawning migrants to home-ranges.
The bidirectional seasonal spawning movement also explains a fully-time varying probability of entry, capture rate and apparent survival. The increase in numbers of individuals in the population resulted in a higher survival rate and capture rate of bonefish during the norths season and in some sample periods of the dry and rainy seasons. The lower peaks of survival and capture rate was likely the result of deaths/emigration and fewer fish entering the population. Both are also supported by the population size estimates as discussed above. Migration is important as it determines fecundity and fitness (Begon et al. 2006; Dingle 2014) but also reduces survival through predation (Shaw and Levin 2011), poaching (Schmitter-Soto and Herrera-Pavón 2019), ASC and recreational harvest (McGarvey and Feenstra 2002; McGarvey 2009), injury and predation from angling (Danylchuk et al. 2007c), dispersal after spawning (Begon et al. 2006), loss of site attachment (Murchie et al. 2013) and mortality by habitat loss.
The spatial extent of the superpopulation was likely larger than the sampled area. This means that although sampling was conducted in 40.8 km2 of flats habitats, the bonefish habitat mosaic that supports a healthy population is a larger catchment area that provides adults to the pre-spawning aggregation site and receives larvae from spawning at that and other sites. Thus, incorporating catchment on a spatial scale of habitats for adult, juvenile, and larval phases into fisheries and protected areas management is important for species that aggregate to spawn (Sadovy de Mitcheson et al. 2008) such as bonefish. Approximately 188,000 adult bonefish from the regional catchment area aggregated to spawn at some point during the extended spawning season (October through April, Danylchuck et al. 2011), which based on our estimates peaks in November-December-February. This highlights spatio-temporal vulnerability of adult bonefish during this season due to the massive numbers and behavioral schooling pattern in PSA sites. Additionally, it is possible there is a superpopulation size of bonefish < 22 cm similar to our N estimate (c.a 197,350) for bonefish > 22 cm. We deduce this from previous studies by Perez et al. (2019a) which show nearly half of the captured bonefish in sampling events were < 22 cm, all of which were released unmarked (50.1 % of the mean abundance). Lastly, because our sampling was in habitats of large-sized bonefish, we presume non-sampled habitats (i.e. non shallow and sturdy habitats) likely form part of home-ranges or recruitment habitats of younger bonefish, which further supports the need for a large catchment area to sustain a viable bonefish population.
Population size and stability depends on healthy ecosystems and sustainable use of resources. This suggests that additional stresses and vulnerability of bonefish from ASC fisheries, habitat degradation and the flats fishery should be considered in present and future conservation and management systems. As has been shown worldwide, unregulated harvest of pre-spawning aggregations of bonefish (Beets 2001; Filous et al. 2019) and spawning aggregations of reef fish have reduced populations rapidly (Sala et al. 2001), thus decreasing the viability of a population (Begon et al. 2006). Furthermore, coastal development causes detrimental effects to habitats through reduced water quality and altered physicochemical and biological processes from urban and agriculture runoff (Ortiz-Hernández and Sáenz-Morales 1997; Vidal-Martínez et al. 2003) and degraded and modified structure, diversity and abundances of organisms and ecosystems (Hernández-Arana and Ameneyro-Angeles 2011; Medina-Quej et al. 2009; Schmitter-Soto et al. 2018; Schmitter-Soto and Herrera-Pavón 2019). Finally, physical stresses by angling disrupts natural physiological processes and behaviors that ultimately affects survival (Danylchuk et al. 2007a). This is important because although CR is a valuable conservation tool, additional regulatory measures should be considered, including prohibiting angling in pre-spawning aggregations during the reproductive season, a continuous advocacy for the use of good handling practices, and fishery capacity (Perez et al. 2020), where fishery capacity is defined as: the amount of fishing effort the fishery can support while maintaining a high quality fishery (high catch rates, large fish size, intact habitats) (Adams 2017).
This study provides important population characteristics for protected areas and fisheries management with interjurisdictional implications for Belize and Mexico. Our recommendations for using the data from this and related studies that highlight shared resources include: protection of the PSA site to reduce vulnerability of adult bonefish to habitat loss and degradation, predation, angling, poaching and harvest; creating and implementing a fisheries sustainability plan (Medina-Quej et al. 2009) that incorporates the spatiotemporal factors identified here (e.g., seasonal migration, spawning migration); research on the interaction of the ASC fisheries, such as beach traps along the coastline, to enable formulation of conservation plans to reduce or mitigate impacts; establishing a catch-and-release sustainability plan; establish an evaluation system for coastal development, urban and agricultural activities, and enforcement of associated regulations, that incorporates information on fisheries and associated fish species and habitats; and a historical reconstruction of the bonefish population to enable evaluation of management success or failure as a function of bonefish population size.
References
Acolas M-L, Lambert P (2016) Life Histories of Anadromous Fishes. In: Morais P, Daverat F (eds) An introduction to fish migration. CRC Press, Boca Raton, pp 55–77
Adams AJ (2017) Guidelines for evaluating the suitability of catch and release fisheries: Lessons learned from Caribbean flats fisheries. Fish Res 186:672–680. https://doi.org/10.1016/j.fishres.2016.09.027
Adams AJ, Murchie KJ (2015) Recreational fisheries as conservation tools for mangrove habitats. Am Fish Soc Symp 83:43–56
Adams AJ, Dahlgren CP, Kellison GT et al (2006) Nursery function of tropical back-reef systems. Mar Ecol Prog Ser 318:287–301
Adams A, Wolfe R, Tringali M et al (2008) Rethinking the status of Albula spp. biology in the Caribbean and western Atlantic. In: Ault JS (ed) The biology and management of the world tarpon and bonefish fisheries. CRC Press, Boca Raton, pp 203–214
Adams AJ, Horodysky AZ, Mcbride RS et al (2013) Global conservation status and research needs for tarpons (Megalopidae), ladyfishes (Elopidae) and bonefishes (Albulidae). Fish Fish 15:280–311
Adams AJ, Rehage JS, Cooke SJ (2019a) A multi-methods approach supports the effective management and conservation of coastal marine recreational flats fisheries. Environ Biol Fishes 102:105–115. https://doi.org/10.1007/s10641-018-0840-1
Adams AJ, Shenker JM, Jud Z et al (2019b) Identifying pre-spawning aggregation sites for the recreationally important bonefish (Albula vulpes) to inform conservation. Environ Biol Fishes 102:159–173
Akaike H (1973) Maximum likelihood identification of gaussian autoregressive moving average models. Biometrics 60:255–265
Akçakaya HR (2000) Viability analyses with habitat-based metapopulation models. Popul Ecol 42:45–53. https://doi.org/10.1007/s101440050008
Arlinghaus R, Cooke SJ, Potts W (2013) Towards resilient recreational fisheries on a global scale through improved understanding of fish and fisher behaviour. Fish Manag Ecol 20:91–98. https://doi.org/10.1111/fme.12027
Arnason AN (1972) Parameter estimates from mark-recapture experiments on two population subjects to migration and death. Res Popul Ecol (Kyoto) 13:97–113
Arnason AN (1973) The estimation of population size, migration rates and survival in a stratified population. Res Popul Ecol (Kyoto) 15:1–8. https://doi.org/10.1007/BF02510705
Arnason NA, Schwarz CJ (1999) Using popan-5 to analyse banding data. Bird Study 46:S157–S168. https://doi.org/10.1080/00063659909477242
Ault JS, Moret S, Luo J et al (2008) Florida Keys bonefish population census. In: Ault JS (ed) Biology and management of the world tarpon and bonefish fisheries. CRC Press, Boca Raton, pp 383–398
Beets J (2001) Declines in finfish resources in Tarawa Lagoon, Kiribati, emphasize the need for increased conservation effort. Atoll Res Bull 1–14. https://doi.org/10.5479/si.00775630.490
Begon M, Harper JL, Townsend CR (2006) Ecology from individuals to ecosystems, 3rd edn. Blackwell Publishing, USA
Binder TR, Cooke SJ, Hinch SG (2011) The biology of fish migration. In: Farrell AP (ed) Encyclopedia of fish physiology: from genome to environment. Academic, San Diego, pp 1921–1927
Boucek R, Adams AJ (2011) Comparison of retention success for multiple tag types in common snook. North Am J Fish Manag 31:693–699. https://doi.org/10.1080/02755947.2011.611044
Boucek RE, Lewis JP, Stewart BD et al (2019) Measuring site fidelity and homesite-to-pre-spawning site connectivity of bonefish (Albula vulpes): using mark-recapture to inform habitat conservation. Environ Biol Fishes 102:185–195
Brownscombe JW, Danylchuk AJ, Adams AJ et al (2019) Bonefish in South Florida: status, threats and research needs. Environ Biol Fishes 102:329–348. https://doi.org/10.1007/s10641-018-0820-5
Colton DE, Alevizon WS (1983) Feeding ecology of bonefish in Bahamian waters. Trans Am Fish Soc 112:178–184
Couto A, BaptistaM, Furtado M, Sousa L, Queiroz N (2016) Life histories of oceanodromous fishes. In: Morais P, Daverat F (eds) An introduction to fish migration. CRC Press, Boca Raton, pp 123–146
Danylchuk AJ, Danylchuk SE, Cooke SJ et al (2007a) Post-release mortality of bonefish, Albula vulpes, exposed to different handling practices during catch-and-release angling in Eleuthera, The Bahamas. Fish Manag Ecol 14:149–154. https://doi.org/10.1111/j.1365-2400.2007.00535.x
Danylchuk AJ, Danylchuk SE, Cooke SJ, Goldberg TL (2007b) Do bonefish (Albula vulpes) use mangroves for protection from predators following catch-and-release angling? In: 59th Gulf and Caribbean Fisheries Institute, pp 417–422
Danylchuk SE, Danylchuk AJ, Cooke SJ et al (2007c) Effects of recreational angling on the post-release behavior and predation of bonefish (Albula vulpes): the role of equilibrium status at the time of release. J Exp Mar Bio Ecol 346:127–133. https://doi.org/10.1016/j.jembe.2007.03.008
Danylchuk AJ, Danylchuk SE, Cooke SJ et al (2008) Ecology and management of bonefish (Albula spp.) in the Bahamian archipelago. In: Ault JS (ed) Biology and management of the world tarpon and bonefish fisheries. CRC, Boca Raton, pp 79–92
Danylchuk AJ, Cooke SJ, Goldberg TL et al (2011) Aggregations and offshore movements as indicators of spawning activity of bonefish (Albula vulpes) in The Bahamas. Mar Biol 158:1981–1999
Danylchuk AJ, Lewis J, Jud Z et al (2019) Behavioral observations of bonefish (Albula vulpes) during prespawning aggregations in the Bahamas: clues to identifying spawning sites that can drive broader conservation efforts. Environ Biol Fishes 102:175–184. https://doi.org/10.1007/s10641-018-0830-3
Dingle H (2014) Migration: the biology of life on the move, 2nd edn. Oxford University Press, Oxford
FAO (2009) Fisheries management. 2. The ecosystem approach to fisheries. 2.2 Human dimensions of ecosystem approach to fisheries. Technical guidelines for responsible fisheries. No. 4, Suppl. 2, Add. 2. Rome
FAO (2012) Recreational fisheries. FAO technical guidelines for responsible fisheries. No. 13. Rome
Fedler AJ (2009) The economic impact of recreational fishing in the Everglades Region. Palmetto Bay
Fedler A (2013) Economic impact of the Florida Keys flats fishery. Vero Beach, FL
Fedler AJ (2014) 2013 economic impact of flats fishing in Belize. Vero Beach, FL
Fedler AJ (2019) The economic impact of flats fishing in The Bahamas. Miami, FL
Filous A, Lennox RJ, Eveson P et al (2019) Population dynamics of roundjaw bonefish Albula glossodonta at a remote coralline atoll inform community-based management in an artisanal fishery. Fish Manag Ecol 27:200–214. https://doi.org/10.1111/fme.12399
Friedlander AM, Parrish JD (1998) Habitat characteristics affecting fish assemblages on a Hawaiian coral reef. J Exp Mar Bio Ecol 224:1–30. https://doi.org/10.1016/S0022-0981(97)00164-0
González NE, Carrera-Parra LF, Salazar-Silva P et al (2009) Macrobentos. In: Espinoza-Ávalos J, Islebe GA, Hernández-Arana HA (eds) El sistema ecológico de la bahía de Chetumal/Corozal: costa occidental del Mar Caribe. El Colegio de la Frontera Sur, Chetumal, pp 88–101
Government of Belize (2003) Fisheries Act (extension of application) Order, Fisheries Act Chap. 210, revised edition 2003. 226
Government of Belize (2009a) S.I. 114 of the Fisheries Act, Chap. 210. Belize Government Gazzette, Belize
Government of Belize (2009b) S.I. 115 of the Coastal Zone Management Authority Act, Chap. 329. Belize Government Gazzette, Belize
Government of Belize (2015) Fisheries (Hol Chan Marine Reserve) Regulations 2015. Belize Government Gazzette, Belize
Grimshaw T, Paz G (2004) The revised Bacalar Chico National Park & Marine Reserve management plan. Green Reef Environmental Insitute, San Pedro
Hernández-Arana H, Ameneyro-Angeles B (2011) Benthic biodiversity changes due to the opening of an artificial channel in a tropical coastal lagoon (Mexican Caribbean). J Mar Biol Assoc United Kingdom 91:969–978. https://doi.org/10.1017/S0025315410002043
Heyman WD, Kjerfve B (2008) Characterization of transient multi-species reef fish spawning aggregations at Gladden Spit, Belize. Bull Mar Sci 83:531–551
Heyman WD, Kjerfve B, Graham RT et al (2005) Spawning aggregations of Lutjanus cyanopterus (Cuvier) on the Belize Barrier Reef over a 6 year period. J Fish Biol 67:83–101. https://doi.org/10.1111/j.1095-8649.2005.00714.x
Hightower JE, Pollock KH (2013) Tagging methods for estimating population size and mortality rates of inland striped bass populations. Am Fish Soc Symp, pp 249–262
Jolly GM (1965) Explicit estimates from capture-recapture data with both death and immigration- stochastic model. Biometrics 52:225–247
Larkin MF (2011) Assessment of South Florida’s bonefish stock. University of Miami, FL
Mackenzie DI, Royle JA (2005) Designing occupancy studies: general advice and allocating survey effort. J Appl Ecol 42:1105–1114. https://doi.org/10.1111/j.1365-2664.2005.01098.x
MacKenzie DI, Nichols JD, Royle JA et al (2006) Occupancy estimation and modeling: inferring patterns and dynamics of dpecies occurrence. Academic, Burlington
McGarvey R (2009) Methods of estimating mortality and movement rates from single-tag recovery data that are unbiased by tag non-reporting. Rev Fish Biol Fish 17:291–304. https://doi.org/10.1080/10641260802664841
McGarvey R, Feenstra JE (2002) Estimating rates of fish movement from tag recoveries: conditioning by recapture. Can J Fish Aquat Sci 59:1054–1064. https://doi.org/10.1139/f02-080
Medina-Quej A, Arce-Ibarra AM, Herrera-Pavón RL et al (2009) Pesquerías: sector social, recurso base y manejo. In: Espinoza-Ávalos J, Islebe GA, Hernández-Arana HA (eds) El sistema ecológico de la bahía de Chetumal/Corozal: costa occidental del Mar Caribe. El Colegio de la Frontera Sur, Chetumal, Mexico, pp 184–195
Mumby PJ (2005) Connectivity of reef fish between mangroves and coral reefs: algorithms for the design of marine reserves at seascape scales. Biol Conserv 128:215–222
Murchie KJ, Cooke SJ, Danylchuk AJ et al (2013) Movement patterns of bonefish (Albula vulpes) in tidal creeks and coastal waters of Eleuthera, The Bahamas. Fish Res 147:404–412
Murchie KJ, Haak CR, Power M et al (2019) Ontogenetic patterns in resource use dynamics of bonefish (Albula vulpes) in the Bahamas. Environ Biol Fishes 102:117–127. https://doi.org/10.1007/s10641-018-0789-0
Ortiz-Hernández MC, Sáenz-Morales R (1997) Effects of organic material and distribution of fecal coliforms in Chetumal Bay, Quintana Roo, Mexico. Environ Monit Assess 55:423–434. https://doi.org/10.1023/A:1005939100154
Perez AU, Schmitter-Soto JJ, Adams AJ, Herrera-Pavón RL (2019a) Influence of environmental variables on abundance and movement of bonefish (Albula vulpes) in the Caribbean Sea and a tropical estuary of Belize and Mexico. Environ Biol Fishes 102:1421–1434. https://doi.org/10.1007/s10641-019-00916-0
Perez AU, Schmitter-Soto JJ, Adams AJ, Heyman WD (2019b) Connectivity mediated by seasonal bonefish (Albula vulpes) migration between the Caribbean Sea and a tropical estuary of Belize and Mexico. Environ Biol Fishes 102:197–207. https://doi.org/10.1007/s10641-018-0834-z
Perez AU, Schmitter-Soto JJ, Adams AJ (2020) Towards a new shift in conservation and management of a fishery system and protected areas using bonefish (Albula vulpes) as an umbrella species in Belize and Mexico. Environ Biol Fishes 103:1359–1370. https://doi.org/10.1007/s10641-020-01028-w
Perez-Cobb AU, Arce-Ibarra AM, García-Ortega M et al (2014) Artisanal recreational fisheries: using a combined approach to fishery assessment aimed at providing insights for fishery managers. Source Mar Resour Econ 29:89–109
Pickett BD, Wallace EM, Ridge PG, Kauwe JSK (2020) Lingering taxonomic challenges hinder conservation and management of global bonefishes. Fisheries 45:347–358. https://doi.org/10.1002/fsh.10438
Pine WE, Pollock KH, Hightower JE et al (2003) A review of tagging methods for estimating fish population size and somponents of mortality. Fisheries 28:10–23. https://doi.org/10.1577/1548-8446(2003)28
RStudio-Team (2016) RStudio: Integrated development for R. RStudio, Inc. http://rstudio.com/
Russ GR, Alcala AC, Mayapa AP et al (2004) Marine reserve benefits local fisheries. Ecol Appl 14:597–606
Sadovy de Mitcheson Y, Cornish A, Domeier M et al (2008) A global baseline for spawning aggregations of reef fishes. Conserv Biol 22:1233–1244. https://doi.org/10.1111/j.1523-1739.2008.01020.x
Sala E, Ballesteros E, Starr RM (2001) Rapid decline of Nassau grouper spawning aggregations in Belize: fishery management and conservation needs. Fisheries 26:23–30. https://doi.org/10.1577/1548-8446(2001)026<0023:RDONGS>2.0.CO;2
Sarteneja Alliance for Conservation and Development (2015) Northern Belize coastal complex management action planning outputs, Summary. Sarteneja Alliance for Conservation and Development, Sarteneja, Belize, 42 p
Sarteneja Alliance for Conservation and Development (2020) Corozal Bay Wildlife Sanctuary Management Plan 2020–2024. Sarteneja Alliance for Conservation and Development, Sarteneja, Belize, 256 p
Schmitter-Soto JJ, Herrera-Pavón RL (2019) Changes in the fish community of a western Caribbean estuary after the expansion of an artificial channel to the sea. Water 11:w11122582. https://doi.org/10.3390/w11122582
Schmitter-Soto JJ, Aguilar-Perera A, Cruz-Martínez A et al (2018) Interdecadal trends in composition, density, size, and mean trophic level of fish species and guilds before and after coastal development in the Mexican Caribbean. Biodivers Conserv 27:459–474. https://doi.org/10.1007/s10531-017-1446-1
Schwarz CJ, Arnason AN (1996) A general methodology for the analysis of capture-recapture experiments in open populations. Biometrics 52:860–873
Seber GAF (1965) A note on the multiple-recapture census. Biometrics 49:269–271
Shaw AK, Levin SA (2011) To breed or not to breed: a model of partial migration. Oikos 120:1871–1879. https://doi.org/10.1111/j.1600-0706.2011.19443.x
Sheaves M (2009) Consequences of ecological connectivity: the coastal ecosystem mosaic. Mar Ecol Prog Ser 391:107–115. https://doi.org/10.3354/meps08121
Steinberg MK (2015) A nationwide assessment of threats to bonefish, tarpon, and permit stocks and habitat in Belize. Environ Biol Fishes 98:2277–2285. https://doi.org/10.1007/s10641-015-0429-x
Suski CD, Cooke SJ, Danylchuk AJ et al (2007) Physiological disturbance and recovery dynamics of bonefish (Albula vulpes), a tropical marine fish, in response to variable exercise and exposure to air. Comp Biochem Physiol - A Mol Integr Physiol 148:664–673
Sweetman BM, Cissell JR, Rhine S, Steinberg MK (2019) Land cover changes on Ambergris Caye, Belize: a case study of unregulated tourism development. Prof Geogr 71:123–134. https://doi.org/10.1080/00330124.2018.1501710
Thurow RF (2016) Life histories of potamodromous fishes. In: Morais P, Daverat F (eds) An introduction to fish migration. CRC Press, Boca Raton, pp 29–54
Torres-Chávez P, Schmitter-Soto JJ, Mercado-Silva N, Valdez-Moreno ME (2018) Movimiento entre hábitats de la barracuda Sphyraena barracuda, determinado por aproximaciones tróficas en el Caribe. Rev Mex Biodivers 89:865–872
Vidal-Martínez VM, Aguirre-Macedo ML, Noreña-Barroso E et al (2003) Potential interactions between metazoan parasites of the Mayan catfish Ariopsis assimilis and chemical pollution in Chetumal Bay, Mexico. J Helminthol 77:173–184. https://doi.org/10.1079/joh2002158
White GC, Burnham KP (1999) Program MARK: survival estimation from populations of marked animals. Bird Study 46:S120–S139. https://doi.org/10.1080/00063659909477239
Acknowledgements
Funding sources were the Mexican Consejo Nacional de Ciencia y Tecnología, via project No. 242558, and Bonefish & Tarpon Trust. We are thankful to fishers, guides (Omar Arceo, Jose Polanco, Geovanni Ortega, Rob Mukai), volunteers (Jon-Pierre Windsor, Felipe Martínez, Antonio Aguilar, Fernando Aguilar, Julio Cárdenas, Jason Maize, Rudy Castellanos, David González, Yasmin González, Norman Mercado, Axel Schmitter, Martha Valdez), fishing lodges (Omar Arceo Freelance Fishing, El Pescador Lodge and Villas, Costa de Cocos, Acocote Inn and Flats Fly-Fishing Guides Services) and Sarteneja Alliance for Conservation and Development. Special thanks to Janneth Padilla for the study area map. Fishing permits: PPF/DGOPA-053/15 in Mexico and 000008–16 in Belize, with additional authorization from the corresponding protected areas.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical approval
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed as highlighted in acknowledgements and methodology.
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Perez, A.U., Schmitter-Soto, J.J. & Adams, A.J. The first estimate of interjurisdictional population dynamics for bonefish, Albula vulpes, a shared resource in the western Caribbean. Environ Biol Fish 104, 341–356 (2021). https://doi.org/10.1007/s10641-021-01081-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-021-01081-z