
Abstract
Characterizing the potential for, and mechanisms of, organismal response to environmental stress, particularly in the context of temperature, remains important for predicting effects of global change on natural populations. A number of studies have looked at changes in behavioral variation, focusing primarily on activity, boldness and aggression, while many behavioral traits have gone unexplored. In this study, I examine responses of a widespread neotropical livebearer, the barred killifish, to variation in water temperature. In addition to activity and boldness, I assayed variation in anxiety (measured as thigmotaxis – preference for edges) following a short-term exposure to thermal stress. Similar to other studies, elevated temperatures were associated with significant increases in activity and propensity for risk-taking behavior. There was no effect of thermal stress on anxiety, but males did have overall lower anxiety than females. An increase in activity and boldness may be a general response to thermal stress, even if competing hypotheses of the adaptive benefit remain unresolved. The insensitivity of thigmotaxis to thermal stress suggests that this measure of anxiety may be independent of variation along the shy-bold axis and therefore represent a useful measure for future studies of behavioral variation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Characterizing context-dependent behavior is important for a variety of reasons. For example, on one hand, plasticity has important implications for predicting resilience and evolutionary response during periods of environmental change. On the other hand, the extent and time-course of plastic responses (or acclimation effects) has consequences for experimental design and interpretation of results (Allen et al. 2012). This is particularly true given that environments can change over relatively short time-scales (e.g., diel variation; Biro et al. 2009). Of the many biotic and abiotic environmental variables that may impact behavior, temperature is among the most important as it has far-reaching effects on organismal biology (Somero et al. 2017).
Temperature-dependence of behavior is widely documented across animal taxa. At its most basic, behavioral thermoregulation can help an organism proactively change its thermal environment (Stevenson 1985), thereby mitigating potential costs of sub-optimal temperatures. Many behavioral responses to temperature, however, are considered to be the result of shifting energetic demands (Houston 2010; Briffa et al. 2013). As temperature increases, so too do metabolic costs (Gillooly et al. 2001; Clarke and Fraser 2004). It has been suggested that changes in behavioral traits such as activity and boldness (risk-taking behavior) reflect a fundamental underlying link between metabolism and behavior (Careau et al. 2008). Two competing models have been suggested: (1) a performance model in which metabolic rate should be positively associated with activity and related traits because sustaining high activity requires increased maintenance costs and (2) an allocation model which posits that many organisms have finite food resources, resulting in a trade-off between energy allocated to behavior compared to energy used to sustain metabolic rate (Careau et al. 2008). The performance model would suggest increased activity and related traits under thermals stress, while the allocation model would predict reductions in such behaviors. Alternatively, behavioral variation and metabolic rate may be uncorrelated (Polverino et al. 2018), as variation in basal energetic demands may not increase or decrease energy available for other organismal requirements (Mathot and Dingemanse 2015). Existing evidence remains mixed (Careau et al. 2008) with evidence for individual variation within populations (Biro et al. 2009; McDonnell et al. 2019). What remains largely unexplored is how traits reflecting other dimensions of personality are impacted by thermal stress.
In the present study, multiple behaviors of a livebearing fish following short-term exposure to two different thermal regimes were evaluated. Aside from activity and boldness, anxiety was also measured, which is rarely evaluated in studies of thermal stress. It was predicted that thermal stress would either increase or decrease activity and boldness (fitting one of the proposed models described above), but there was no a priori expectation of which direction those behaviors would shift. Similarly, it was predicted that anxiety may also shift in response to thermal stress, especially if it reflects variation along the typical shy-bold axis of behavior. If anxiety reflects variation along the shy-bold axis of behavior, then the change in anxiety under thermal stress should match that of boldness.
Methods
Study species and collection
Pseudoxiphophorus jonesii are a small, livebearing fish native to Northeastern Mexico north of the Trans-Mexican Volcanic Belt (Agorreta et al. 2013). The species occurs across a broad range of elevations from the lowlands near sea-level to at least ~1500 m in streams along the slopes of the Sierra Madre Oriental (Bautista-Hernández et al. 2015; ZWC personal observation). Adult male (N = 13) and female P. jonesii (N = 11) were collected from the Rio Atlapexco (21.015, −98.339) in the state of Hidalgo, Mexico in June 2019. The water temperature at the time of collection was 27 °C. Fish were transported to the Centro de Investigaciones de las Huastecas Aguazarca (CICHAZ) field station in an insulated container with water from their collection site.
In the lab, individuals were randomly assigned to one of four, 40 L aquaria, where they were acclimated overnight. The following morning, two tanks were designated as the ‘cool’ treatment and two as the ‘warm’ treatment. Submersible heaters were used with internal thermostats to maintain the cool tanks between 27 and 28 °C (mean ± SD: 27.7 ± 0.66; range: 26.2–28.7 °C) and to raise the temperature of the warm tanks to 33 °C (mean ± SD: 33.2 ± 0.41; range: 32.2–34.3 °C). Temperatures were logged at 5-min intervals over the course of the experiment with a submersible temperature logger (Onset Computer Corp., Bourne, MA, USA). To avoid any acute stress, temperature in the warm treatment was increased slowly, averaging 0.34 °C/h (range 0.29–0.5 °C/h) or 0.006 °C/min. As a point of reference, the rate of increase typically used to estimate thermal limits is 50x greater (0.3 C/min; Lutterschmidt and Hutchison 1997). Once the temperature in the warm tanks had reached 33 °C, fish were maintained under their respective thermal conditions for approximately 3 days (range: 68–70.5 h). The effect of trial time which would reflect any time of day or order effects on performance in the behavioral assay was tested, but none were found. Fish were fed ad libitum with premium flake food twice per day and maintained on an ambient light cycle (~14 L:10 D).
Behavioral assay
On the final day of the experiment, each individual was separately tested in an open field trial (OFT). The OFT is a standard approach for quantifying a variety of non-social behaviors. A single individual was transferred to an opaque plastic tub (57 cm top diameter × 47 cm bottom diameter × 25 cm depth) filled to a depth of 15 cm with aged tap water. Water used in the tubs for OFT was preheated to match the water temperature in the respective cool and warm treatments. The tub was empty with no structure for the focal fish to use as cover. A digital camera was mounted 1 m above the testing arena to record each trial. As in other studies of personality in livebearing fish (White et al. 2019), the OFT was run for 5 min and the first 30 s of each recording were discarded as acclimation time. At the end of the OFT, the individual was removed, measured for standard length and transferred to a holding tank.
From the OFT recordings, I measured multiple non-social behaviors. I measured activity as the total time the focal fish spent actively swimming in the arena. Boldness was measured as the amount of time and the number of times an individual visited an exposed, putatively risky zone in the middle of the arena. This zone was delineated by an 11 cm diameter circle drawn on the bottom of the tub. This zone was placed exactly in the center of the arena and corresponded to ~5% of the total area of the bottom of the arena. Anxiety was estimated based on thigmotaxis (preference for tank edge) and measured as the distance of the focal individual from the edge of the OFT arena. This distance was averaged from measurements made in imageJ (Schneider et al. 2012) from 19 still images, captured at 15 s intervals over the course of each trial. Due to a strong correlation between time spent in the center zone and the measure of anxiety, subsequent analyses of boldness focused only on the propensity of individuals to engage in risky behavior (i.e., whether or not an individual entered the center).
Statistical analyses
I conducted separate general linear models (glm) for activity and anxiety using treatment, sex, and standard length as fixed effects. The interaction between treatment and sex was non-significant for both behaviors and was subsequently removed from final models, since there was no a priori expectation that the sexes would differ in response between treatments, and to avoid model over-fitting due to the limited sample size. The residuals met assumptions for glms. I evaluated the difference in propensity to engage in risky behavior between treatments using a Fisher’s exact test based on whether or not each individual visited the center zone at least once or never at all. Analyses were conducted in SPSS v25 (IBM Inc.) and all tests were two-tailed.
Results
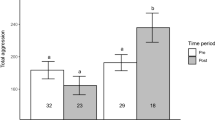
There was a significant difference in activity levels between treatments (F1,19 = 4.84, P = 0.04; Table 1), but not between sexes (Fig. 1a, b). Individuals in the warm treatment were 40% more active. Body size also had no effects on overall activity. There was no effect of temperature treatment (Fig. 1c) or body size on anxiety. The only significant difference observed was between the sexes (F1,19 = 9.78, P = 0.006; Table 1; Fig. 1d). Males exhibited lower anxiety than females across both treatments. There was a difference in the propensity for individuals to visit the center zone between treatments. Individuals in the hot treatment were more likely to engage in risky behavior than those in the cool treatment (Fisher’s exact test: P = 0.012). In the hot treatment, 73% of individuals visited the center zone of the arena compared to only 17% of individuals in the cool treatment.

Estimated marginal means (±SE) for activity (a, b) and anxiety (c, d). Activity was significantly different between treatments, but not sexes. Anxiety was significantly different between sexes but not treatments (Table 1). Anxiety values are presented as 1/thigmotaxis scores so that larger values reflect greater anxiety
Discussion
Behavioral variation in response to environmental stress has important implications both for the capacity and mechanisms by which natural populations respond to environmental challenges but also for experimental design. Short-term thermal stress increased locomotor activity and risk-taking behavior in Pseudoxiphophorus jonesii. Males showed reduced anxiety compared to females but there was no effect of temperature on anxiety for either sex. These results suggest that livebearing fish may exhibit similar patterns to other species in that they may compensate for increased metabolic demands by increasing activity and risk-taking. The results also demonstrate that the measure of anxiety used here was independent of activity and boldness and was insensitive to physiological stress.
It is generally assumed that increased activity and boldness under thermal stress reflects a general response related to increased metabolic demands (Careau et al. 2008; Biro et al. 2009). For example, to meet energetic needs, individuals may become more active or take more risks in their search for food. Nonetheless, some studies find considerable variation among individuals or mean reductions in locomotor activity (Martinez et al. 2007; Biro et al. 2009; McDonnell et al. 2019). In this study, fish were fed ad libitum twice per day in order to potentially alleviate energetic costs of increased temperature, but I cannot rule out that energetic costs were a driving factor in behavioral differences between temperature treatments. One prior study suggested that metabolic demands alone could not entirely explain observed behavioral shifts of a marine fish under thermal stress (Biro et al. 2009). Indeed, at least two alternative explanations may influence behavioral changes observed in this and other studies. One alternative is that increased activity and engagement in risky behavior reflects a strategy simply to avoid (or escape) a stressful condition similar to behavioral thermoregulation in other ectotherms (Seymour 1972; Huey 1974). In other words, if your environment becomes too warm, then move until you find a more favorable environment. A second alternative is that thermal stress may compromise cognition and decision-making abilities, resulting in suboptimal behavior that may lead to increased exposure to predators. These competing hypotheses are likely to remain difficult to disentangle because they are not mutually exclusive. Regardless of the underlying mechanism, variation in behavior or correlations among behaviors in response to even short-term physiological stress could have important implications for selection and evolution in natural populations.
In P. jonesii, activity and boldness were significantly affected by temperature but anxiety was not, suggesting that this measure of anxiety reflects variation along a distinct behavioral axis (i.e., not just another metric along the shy-bold continuum). One largely unexplored question is how far-reaching are the effects of physiological stress on behavior and personality. Anxiety is not commonly tested in response to thermal stress. At least one study with rats found similar results in that anxiety (measured as entries into open arms in a y-maze) was insensitive to thermal stress (Martinez et al. 2007). The authors concluded that while general motor activities varied across temperature, traits related to emotional state (such as anxiety) were not affected by thermal stress. The present study with P. jonesii provides additional support for limited or no effects of thermal stress on emotional state. However, it cannot be ruled out that a larger sample size may have helped detect a small, but significant effect of treatment on anxiety that was not detectable in the present study. In any case, additional studies in other taxa and that evaluate a wider range of behaviors are needed in order to understand the generality of these patterns.
Global change is impacting natural populations, with particular effects on thermal regimes. Examining species responses to thermal stress is critical not just to understanding how and whether species can withstand environmental changes, but for predicting evolutionary responses to changes in selection. There appear to be consistent patterns in some traits such as activity and boldness, but many behavioral traits remain underexplored. Furthermore, whether sex-specific variation in emotional traits such as anxiety reflect selection related to sexual conflict, exposure to increased predation in males, or other factors, remains unclear. Ultimately, studies that examine a wider variety of traits and test responses comparatively across species with variation in reproductive biology should provide critical insights into these questions.
References
Agorreta A, Domínguez-Domínguez O, Reina RG, Miranda R, Bermingham E, Doadrio I (2013) Phylogenetic relationships and biogeography of Pseudoxiphophorus (Teleostei: Poeciliidae) based on mitochondrial and nuclear genes. Mol Phylogenet Evol 66:80–90
Allen JL, Clusella-Trullas S, Chown SL (2012) The effects of acclimation and rates of temperature change on critical thermal limits in Tenebrio molitor (Tenebrionidae) and Cyrtobagous salviniae (Curculionidae). J Insect Physiol 58:669–678
Bautista-Hernández CE, Monks S, Pulido-Flores G, Miranda R (2015) A new species of Paracreptotrema (Digenea, Plagiorchiformes, Allocreadiidae) infecting two species of poeciliids in Rio Malila of the Rio Pánuco basin, Hidalgo, México, with a key to the species of the genus. Zookeys 55
Biro PA, Beckmann C, Stamps JA (2009) Small within-day increases in temperature affects boldness and alters personality in coral reef fish. Proc R Soc B Biol Sci 277:71–77
Briffa M, Bridger D, Biro PA (2013) How does temperature affect behaviour? Multilevel analysis of plasticity, personality and predictability in hermit crabs. Anim Behav 86:47–54
Careau V, Thomas D, Humphries MM, Réale D (2008) Energy metabolism and animal personality. Oikos 117:641–653
Clarke A, Fraser KPP (2004) Why does metabolism scale with temperature? Funct Ecol 18:243–251
Gillooly JF, Brown JH, West GB et al (2001) Effects of size and temperature on metabolic rates. Science (80- ) 239:2248–2251
Houston AI (2010) Evolutionary models of metabolism, behaviour and personality. Philos Trans R Soc B Biol Sci 365:3969–3975
Huey RB (1974) Behavioral thermoregulation in lizards: importance of associated costs. Science (80- ) 184:1001–1003
Lutterschmidt WI, Hutchison VH (1997) The critical thermal maximum: history and critique. Can J Zool Can Zool 75:1561–1574
Martinez RCR, Garcia AMB, Lamprea MR, Morato S (2007) Thermal stress decreases general motor activity of rats in the elevated plus-maze but does not alter aversion to the open arms. Behav Brain Res 182:135–139
Mathot KJ, Dingemanse NJ (2015) Energetics and behavior: unrequited needs and new directions. Trends Ecol Evol 30:199–206
McDonnell LH, Reemeyer JE, Chapman LJ (2019) Independent and interactive effects of long-term exposure to hypoxia and elevated water temperature on behavior and thermal tolerance of an equatorial cichlid. Physiol Biochem Zool 92:253–265
Polverino G, Santostefano F, Díaz-Gil C, Mehner T (2018) Ecological conditions drive pace-of-life syndromes by shaping relationships between life history, physiology and behaviour in two populations of eastern mosquitofish. Sci Rep 8:14673
Schneider CA, Rasband WS, Eliceiri KW (2012) NIH image to ImageJ: 25 years of image analysis. Nat Methods 9:671–675
Seymour RS (1972) Behavioral thermoregulation by juvenile green toads, Bufo debilis. Copeia 1972:572–575
Somero GN, Lockwood BL, Tomanek L (2017) Biochemical adaptation: response to environmental challenges, from life’s origins to the Anthropocene. Sinauer Associates, Incorporated Publishers
Stevenson RD (1985) The relative importance of behavioral and physiological adjustments controlling body temperature in terrestrial ectotherms. Am Nat 126:362–386
White SJ, Houslay TM, Wilson AJ (2019) Evolutionary genetics of personality in the Trinidadian guppy II: sexual dimorphism and genotype-by-sex interactions. Heredity (Edinb) 122:15
Acknowledgements
I thank the American Livebearer Association Langhammer Conservation Fund for financial support. Research followed approved IACUC procedures and conformed to standards for animal care and use where the research was carried out.
Funding
This study was funded by an American Livebearer Association Langhammer Conservation Fund grant to ZWC.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The author declares that there are no conflicts of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Culumber, Z.W. Thermal stress increases activity and risk-taking behavior but not anxiety in a livebearing fish. Environ Biol Fish 103, 313–317 (2020). https://doi.org/10.1007/s10641-020-00966-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-020-00966-9