
Abstract
In territorial stream salmonids, asymmetric competition can perpetuate individual size differences over time, but the extent to which this is manifested can be environmentally mediated. Here we study the variation in juvenile steelhead (Oncorhynchus mykiss) growth rates to identify the conditions (population density and water temperature) under which an individual’s size relative to its conspecifics conferred an advantage. Among steelhead rearing in the same stream section we found that relatively larger individuals on average grew faster than smaller conspecifics. However, comparing across stream sections there was a negative interaction between relative size and water temperature. The effect of an individual’s relative size on its growth rate decreased as temperatures were increasing, indicating that the advantages of being large diminished during periods of high temperatures or in locations with relatively higher temperatures. Compared to temperature, the effects of population density on the growth rate were less substantial. The results suggest that larger individuals on average acquire more resources than smaller individuals, and demonstrate that water temperature exerts an important, modulating control over growth performance in heterogeneous environments.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Across taxa, body size is a key characteristic which determines individual life history decisions and lifetime fitness (Quinn and Peterson 1996; Fujiwara et al. 2004; Begon et al. 2006; Satterthwaite et al. 2010). A large body size relative to conspecifics can confer advantages in the competition for food and space, aid in the endurance of adverse conditions and periodic food shortages, and help attract and safeguard mates (Abbott and Dill 1989; Nakano 1995; Rose et al. 2001; Fujiwara et al. 2004). In the face of limited resources, size disparities within an age class typically arise when individual demands increase and the competition intensifies (Begon et al. 2006). While variations in size distributions are fairly well documented (Rubenstein 1981; DeAngelis et al. 1993; Crozier et al. 2010; Parra et al. 2012; Ohlberger et al. 2013), it is less well understood how size distributions are created and maintained in wild populations (Lomnicki 1999; Pfister and Stevens 2002; Peacor and Pfister 2006; Peacor et al. 2007a).
When the timing of birth is fairly synchronized, and where individuals experience similar conditions as their conspecifics, size variation within a cohort is ultimately generated by individual differences in their ability to acquire resources and grow under the prevailing conditions (Ricker 1958; Uchmański 1999; Pfister and Stevens 2002; Pfister 2003). The origin of this size variation arise from the advantages (or disadvantages) conferred by a certain size at a given point in time (Ricker 1958), or from persistent individual traits that have a genetic or learned basis such as physiology and foraging behavior (Pfister and Stevens 2002; Pfister 2003; Fujiwara et al. 2004; Peacor et al. 2007a, b). The two categories are not mutually exclusive, and may be difficult to disentangle in wild populations (Pfister and Stevens 2002). Ecologically, both generate asymmetric competition, which result in a range of individual sizes within a population in response to the dynamic selection pressures (Ebenman and Persson 1988; Pfister and Stevens 2002; Fujiwara et al. 2004; Peacor et al. 2007a).
In variable environments and over varying degrees of competition for food and space, it is thus likely that size disparities among individuals manifest unequally between local populations (Rubenstein 1981; Parra et al. 2012; Ohlberger et al. 2013). For example, do initially larger individuals always grow faster, hence perpetuating the size inequality in a population, or do conditions exist where a relatively smaller body size is more beneficial in terms of growth? Individual differences can be particularly evident in fishes because they exhibit relatively flexible and indeterminate growth within a life stage (Rose et al. 2001; Pfister and Stevens 2002) and because phenotypic plasticity can generate multiple life history expressions (Satterthwaite et al. 2010; Benjamin et al. 2013; Phillis et al. 2016). Further, because they are poikilothermic, with a sensitive relationship between temperature and energy use, fishes represent a tractable system with which to study the interacting effects of demographic density dependence and abiotic factors on individual size and growth (Diana 2003; Myrvold and Kennedy 2015a).
Due to their plastic potential and sensitivity to abiotic conditions, a number of studies have investigated the effects of water temperature and population density on the size-at-age and growth rates of stream salmonids. A vast range of results have been reported, which is not surprising given the wide variety of environments that salmonids inhabit and the fact that limiting factors rarely operate in isolation (Crozier et al. 2010; Sloat and Reeves 2014; Myrvold and Kennedy 2015a; Thompson and Beauchamp 2016). Environmental conditions, population density, food availability, and interspecific interactions can greatly influence average demographic responses in local populations, but there is increasing interest in the characteristics of the individuals that make up those populations (Nakano et al. 1991; Nakano 1995; Satterthwaite et al. 2010; Bærum et al. 2013). Individuals in the same stream reach compete for access to food and space, which makes an individual’s ability to compete not only dependent on itself but also on the abilities of its competitors (Nakano et al. 1991; Nakano 1995). In stream salmonids, an individual’s rank in the dominance hierarchy (as evidenced by aggressive displays, quality of feeding territory, and growth performance) has often been attributed to its size (Abbott and Dill 1989; Nicieza and Metcalfe 1999), but the relationships between size and growth are complex and depend on environmental context and individual traits (Höjesjö et al. 2004; Reid et al. 2011). An interesting question, then, is to identify the conditions which may govern size-dependent growth performance.
In the present study we were interested in understanding the circumstances under which an individual’s size relative to conspecifics within a stream reach conferred a growth advantage (Fig. 1). The factors we tested for were population density and water temperature, as both have been shown to influence individual growth rates in stream salmonids (e.g., Crozier et al. 2010; Parra et al. 2012; Bærum et al. 2013; Myrvold and Kennedy 2015a). We use detailed data on individual growth from discrete study sites, which cover a range of densities and temperatures. While we expect that larger individuals are generally better competitors (i.e. when there is sufficient food relative to the water temperature), we hypothesize that high water temperatures can slow down this advantage due to the relationship between metabolic demands and temperature (Myrvold and Kennedy 2015b). We therefore expect a negative effect of the interaction between temperature and relative body mass on individual growth rates. Finally, because body size may confer a dominance advantage, we do not expect density to substantially impede growth in relatively larger individuals.

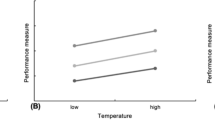
Schematic showing three processes of size-dependent growth. In the top panel the initially larger individual grows faster than the smaller individuals, whereas the bottom panel shows a situation where the initially smaller individual grows faster than the larger individuals. This study aimed at identifying the circumstances when this was occurring
Methods
Study area
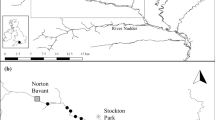
This study took place in the Lapwai Creek watershed in Idaho, United States. The 694 km2 watershed is located at the transition between the Columbia River Plateau and the Northern Rockies ecoregions (McGrath et al. 2002), and spans a gradient of land cover and land use from its headwaters on Craig Mountain (elevation 1530 m) to its confluence with the Clearwater River (elevation 237 m). Lapwai Creek is designated as critical habitat for a threatened population of wild Snake River steelhead (NMFS 2010). Summer water temperatures in lower reaches approach the tolerance level for steelhead (Myrvold and Kennedy 2015b), whereas winter air temperatures are at or below freezing from December through February. The steelhead population in this watershed has been reported on previously (Myrvold and Kennedy 2015a, b, c, d). As part of this work, we established an array of study sites representative of the physiographic variation in the watershed (Fig. 2). The study sites were approximately 100 m long, and key characteristics have been provided in Myrvold and Kennedy (2015c).

Study sites in the Lapwai Creek watershed, Idaho, United States (inset)
Juvenile steelhead data
We captured fish via three-pass depletion electrofishing using a Smith-Root LR-24 backpack electrofisher (Smith-Root Inc., Vancouver, Washington, USA). We visited each study site monthly from June to October in 2010–2012. Prior to any handling we anesthetized the fish with tricaine methanesulfonate (MS-222), and measured fork length in millimeters and mass to the nearest decigram. Individuals were classified as subyearling (hatched in May the same year) or overyearling (hatched in a previous year) based on size frequency histograms at the sampling visit. We tagged individuals 65 mm and longer with Passive Integrated Transponder (PIT) tags (Biomark HPT 12 mm full duplex tags, Biomark Inc., Boise, Idaho, USA). Following handling, fish were first allowed to recover in buckets with aerated water, and then released back into the stream. We estimated the size of each age class using Carle and Strub’s (1978) maximum weighted likelihood estimator for removal data, and calculated density by dividing the population estimate by the area sampled. The density estimate was expressed as the number of juvenile steelhead per 100 m2 and natural log-transformed.
We were interested in measuring the growth of subyearling steelhead compared to the other subyearlings in the respective stream reaches. We therefore considered periods in each site (from visit t to t + 1) for which we obtained growth measurements from four or more individuals. We calculated specific individual growth rate as the percent change in body mass per day between capture and recapture. Relative size was calculated as the difference between the mass of a fish and the average mass of fish in the same site at the sampling visit, and expressed in grams.
Temperature data
Streamwater temperatures were recorded every 30 min in each study site for the duration of the study using HOBO TidbiT v2 temperature loggers (Onset Computer Corporation, Pocasset, Massachusetts, USA). In the modeling we used the average temperature (°C) during the 4 week period between capture and recapture. The distribution of water temperatures among the study sites are given in Appendix 1.
Statistical methods
In order to quantify how much of the variation in the juvenile steelhead growth rates that could be attributed to the different levels in the data (individual and sampling visit to a study site) we performed a one-way analysis of variance (Raudenbush and Bryk 2002). The model for the variance components for the individual- and visit levels (also known as the empty or unconditional model) can be written as
Here, γ 00 is the grand mean growth rate of all individuals across all sampling visits; u 0j is the random visit effect, i.e. the deviation of visit j from the grand mean; and r ij is the residual individual effect, i.e. the deviation of individual ij from the sampling visit mean. Because individual steelhead and sampling visits to a study site were randomly sampled from a larger statistical population of potential fish and visits we can assume that u 0j ~ N(0, τ 00) and r ij ~ N(0,σ 2) (Raudenbush and Bryk 2002). We found substantial clustering by sampling visit, which means that observations made within a sampling visit were more similar than when compared with observations from all sampling visits. We hence modeled the growth rate variation under a hierarchical framework, that is, with both individual- and visit-level effects (Raudenbush and Bryk 2002).
We investigated the effects of individual mass, mass relative to conspecifics (relative mass), population density, and temperature on individual growth rate (Table 1). The complete derivation of the models are given in Appendix 2. Individual mass and relative mass are at the same hierarchical level as the response variable growth. However, to introduce population density and temperature we had to account for the multilevel structure in the data as these variables are not independent among the individuals in a site (Raudenbush and Bryk 2002). This means that the effects of individual mass and relative mass on its growth rate also depend on characteristics of the sampling visit to a study site. A hierarchical linear model with cross-level interactions between characteristics at the sampling visit level (here: temperature) and the individual level can be expressed as
This model tests the effects of individual mass, its mass relative to PIT-tagged conspecifics, and the effects of temperature alone and in interaction with individual specific mass, on an individual’s growth rate. We performed the same analysis with population density as the visit-level predictor variable (see Table 1 for a list of the models considered).
We used SAS 9.2 Proc MIXED (SAS Institute, Cary, North Carolina, USA) specified with the Kenward and Roger (1997) approximation of denominator degrees of freedom, and maximum likelihood as the estimator in all the analyses. Models were ranked using Akaike’s Information Criterion (Akaike 1973); the model with the lowest AIC value is the best approximating model of the data (Burnham and Anderson 2002). Variance inflation factors were less than 2 among the independent variables, and there were hence no problems with multicollinearity. Finally, we visually inspected the residual plots using the ods graphics option in SAS to ensure that statistical assumptions were met (i.e. that the variance components had means of zero and constant variances).
Results
Subyearling steelhead growth rates in the Lapwai Creek watershed averaged 0.91% body mass per day (SE = 0.046) among the 1228 individuals in this study. The variance could be attributed almost equally to individual-level (48% of the variance) and visit-level factors (52%), which suggest that both levels of effects contributed substantially towards the overall variation in subyearling steelhead growth rates.
Model 6 was the best approximating model of growth rate variation, receiving 61% of the support, and model 2 received 22% of the support among the models (Table 1). Because model 6 received more support than model 2, and because model 2 is a subset of model 6, we focus our treatment primarily on model 6. In addition to controlling for mass and relative mass (as with all other candidate models), model 6 included interaction terms with temperature (Table 2). Importantly, the cross-level interaction between relative size and temperature was negative (estimate = −0.0236, SE = 0.0097, t = −2.45, P(|t|) = 0.017). Cross-level interactions show how much the effect of the individual-level variable changes depending on the value of a visit-level variable. This shows that, controlling for an individual’s absolute and relative size, the effect of relative size on its growth rate decreased as temperatures were increasing (Fig. 3). The interaction between absolute mass and temperature was positive. This means that the effect of an individual’s mass on its growth rate increased with increasing temperatures. This owes to the larger average body sizes of steelhead rearing in sites with relatively warmer temperatures and lower densities.

Predicted growth rates (from the best approximating model) for three sizes of steelhead (6 g, 10 g, and 14 g) and three relative sizes (average, 4 g smaller than average, and 4 g larger than average)
Overall, the best model explained 11% of the variation in the growth data (Table 2). The covariates mass and relative mass explained 4% of the variation on the individual level, and inclusion of temperature in interaction with mass and relative mass explained 19% of the visit-level variation. Because the visit-level covariate temperature explained more of the variation in our data, there is reason to believe that the environmental context (i.e. variation in temperature between sites) largely affected the growth dynamics within our study reaches.
Compared to the effects of temperature, we found no substantial effects of density on individual growth rates (Table 1). This suggests that the impacts of temperature were stronger than those of competition when considering an individual’s size relative to its conspecifics; however, this does not imply that density is unimportant.
Discussion
Using data on individual steelhead growth in relation to other known individuals we found that relatively larger fish grew faster, but that their growth advantage depended upon the context. Specifically, there was a negative interaction between an individual’s relative mass and the water temperature. This shows that, controlling for an individual’s absolute size, the positive effects of an individual’s relative mass on its growth rate decreased as temperatures were increasing.
Body size influences most aspects of the relations between an organism and its environment (Ebenman and Persson 1988; Lomnicki 1999; DeRoos et al. 2003; Peacor et al. 2007a). At the level of species, size can determine trophic interactions, and at the level of individuals, larger size often confers advantages in the competition over limited resources and survival (Werner and Gilliam 1984; Ebenman and Persson 1988; Pfister and Stevens 2002). Because body size is closely tied to fitness, individual size variation can affect population density and dynamics (DeAngelis et al. 1993; DeAngelis and Mooij 2005). Some of the variation in size might reflect heritable traits, which underlie natural selection (Gårdmark et al. 2003). As Peacor et al. (2007a) noted the factors that affect size variation within a cohort are hence implicated in exposing different traits to natural selection or masking this variation from selection by preventing expression of trait characters. However, we still have limited understanding of the processes that ultimately generate size variation within a cohort, and the circumstances under which size disparities persist in a dynamic environment (Pfister and Stevens 2002; Peacor and Pfister 2006; Peacor et al. 2007a). Studying individual growth performance within a cohort over relatively short time intervals can help unravel the relationships between individual size, growth, and the environment.
In stream salmonids, growth performance has been linked to an individual’s rank in a local dominance hierarchy. Dominant fish typically acquire better microhabitats and food resources, and as a result, they often grow faster than subordinate conspecifics (Abbott and Dill 1989; Nicieza and Metcalfe 1999; Keeley 2001). Nakano et al. (1991) found that within pools in a Japanese mountain stream, relatively larger red spotted masu salmon (O. masou) grew faster than smaller individuals inhabiting the same pool, despite there being no relationship between initial size and growth when considering the stream as a whole. Subsequently, Nakano (1995) investigated the role of dominance status in explaining this variation in growth rates. Dominant individuals occupied better feeding stations, chose larger prey, and consequently grew faster. Further, dominant individuals were also larger, which is common in stream salmonids when food is abundant. For example, Abbott and Dill (1989) found that dominant juvenile steelhead grew faster than their subordinate counterparts because the appetite and conversion efficiency in subordinates were greatly reduced in the presence of dominant individuals. In experimental channels, Keeley (2001) found that large, aggressive juvenile steelhead consumed a disproportionate amount of the food. Consequently, size inequality within the juvenile stage increased with stocking density and decreased with increases in food availability (Keeley 2001).
There are, however, conditions under which dominance and/or large body size are not beneficial in the sense that they do not confer a growth advantage. Martin-Smith and Armstrong (2002) studied juvenile Atlantic salmon (Salmo salar) in a flow-regulated channel with fluctuating food supply. They found support for the hypothesis that individual growth correlates with movement but not with the ability to dominate good feeding locations. Further, aggressiveness (which can confer social status) has also been linked to physiology and behavior. Reid et al. (2011) found that standard metabolic rate was a strong predictor of social rank, which in turn was related to resource acquisition. However, standard metabolic rate did not predict growth performance, presumably due to the higher associated metabolic costs (Reid et al. 2011). In another study, Vøllestad and Quinn (2003) found that aggressive coho salmon (O. kisutch) grew slower than individuals exhibiting lesser levels of aggression, likely due to the same reason.
The influence of behavior and genotype on growth performance may therefore depend on the environmental context. Höjesjö et al. (2002) studied brown trout (S. trutta) in Sweden and found that although dominant individuals outgrew subordinates, nonaggressive individuals (those not partaking in territorial disputes) grew as fast as the aggressive, dominant individuals. Reid et al. (2012) found that resting metabolic rate in juvenile Atlantic salmon was related to dominance, but that when the food supply was less predictable and the habitat more visually complex, this social dominance was not advantageous in terms of growth. Höjesjö et al. (2004) found a similar pattern in brown trout, where the growth advantage of aggressive and dominant individuals diminished with increasing habitat complexity. In a study of juvenile steelhead in experimental streams, Sloat and Reeves (2014) found a positive directional selection on standard metabolic rate when the distribution of food was predictable. However, when the delivery of food resources was spatially unpredictable there was a negative selection for this trait (Sloat and Reeves 2014). This shows that the local environment may exert an important selection pressure because of physical heterogeneity and dynamic food supply.
Which biotic or environmental factors are limiting to growth depend on the conditions relative to the needs of the organism (Diana 2003; Begon et al. 2006). In previous studies in the Lapwai Creek watershed we have documented the various influences of water temperature on the density variation, cohort regulation, and individual energetics in juvenile steelhead (Myrvold and Kennedy 2015a, b, d). The Lapwai Creek population is located within the range of steelhead and redband rainbow trout, but at a relatively low elevation (250–700 m). The area has a dry summer (June–October), and in lower reaches the water temperatures can approach and temporarily exceed the thermal tolerance of rearing steelhead (Myrvold and Kennedy 2015b). Water temperature thus exerts an important control over individual performance and population dynamics in this system, and may hence be an important driver of growth compensation (i.e. a reduction in the size variability over time).
The discussion has focused on the role of physiological and behavioral underpinnings that may generate size differences in cohorts at different densities and temperatures. Our analysis concerned the subyearling cohort (i.e. individuals hatched in the same year as the sampling took place) to avoid the confounding factor of ontogenetic changes such as those associated with smoltification and migration (Diana 2003). Steelhead in the Lapwai Creek watershed outmigrate primarily at age two, and as yearlings at the earliest (Hartson and Kennedy 2015). However, can we completely discount the role of ontogenetic changes on the size and growth of individuals in such a diverse and plastic species? For the subyearling cohorts studied here we most likely could. Physiological changes occur after the decision to migrate has been made, which is usually months prior to the initiation of movement (Sloat et al. 2014). These physiological changes affect the partitioning of energy to various functions in the body, including somatic growth (Metcalfe 1998; Nicieza and Metcalfe 1999; Sloat et al. 2014; Kendall et al. 2015). However, due to the timing of our sampling (June to October) being 5–6 months prior to the primary outmigration period in this watershed (March–April) we expect the potential for such changes to be very limited in this study.
Ecologists are increasingly viewing a population as a set of individuals rather than as an entity with a mean and some deviation (Rubenstein 1981; Nakano 1995; Lomnicki 1999; DeAngelis and Mooij 2005). Body size influences most aspects of how an organism interacts with its environment directly, its fitness, and how it contributes to the dynamics of its population (Ebenman and Persson 1988; DeAngelis et al. 1993; Lomnicki 1999; Uchmański 1999). Understanding the intrinsic processes that generate size variation in a population (Pfister and Stevens 2002; Peacor et al. 2007a) in concert with abiotic constraints (Reid et al. 2012) can therefore provide great insights into the regulation of populations of mobile organisms and the selection on individual traits (Peacor et al. 2007a). Although natural settings provide the most realistic context to study these processes, they also prove challenging in disentangling the mechanisms generating size variation. More empirical studies from a variety of taxa and ecosystems are thus needed to address the causes and consequences of body size variation in natural populations.
References
Abbott JC, Dill LM (1989) The relative growth of dominant and subordinate juvenile steelhead trout (Salmo Gairdneri) fed equal rations. Behaviour 108:104–113
Akaike H (1973) Information theory and an extension of the maximum likelihood principle. In: Petrov BN, Csaki F (eds) Second international symposium on information theory. Akademiai Kiado, Budapest, Hungary, pp 267–281
Bærum KM, Haugen TO, Kiffney P et al (2013) Interacting effects of temperature and density on individual growth performance in a wild population of brown trout. Freshw Biol 58:1329–1339. doi:10.1111/fwb.12130
Begon M, Townsend CR, Harper JL (2006) Ecology: from individuals to ecosystems, 4th edn. Blackwell Publishing, Malden, Massachussets
Benjamin JR, Connolly PJ, Romine JG, Perry RW (2013) Potential Effects of Changes in Temperature and Food Resources on Life History Trajectories of Juvenile. Trans Am Fish Soc 142(1):208–220
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer-Verlag, New York
Carle FL, Strub MR (1978) A new method for estimating population size from removal data. Biometrics 34:621–630
Crozier LG, Zabel RW, Hockersmith EE, Achord S (2010) Interacting effects of density and temperature on body size in multiple populations of Chinook salmon. J Anim Ecol 79:342–349
DeAngelis DL, Mooij WM (2005) Individual-based modeling of ecological and evolutionary processes. Annu Rev Ecol Evol Syst 36:147–168
DeAngelis DL, Rose KA, Crowder LB et al (1993) Fish cohort dynamics: application of complementary modeling approaches. Am Nat 142:604–622
DeRoos AM, Persson L, McCauley E (2003) The influence of size-dependent life-history traits on the structure and dynamics of populations and communities. Ecol Lett 6:473–487
Diana JS (2003) Biology and ecology of fishes, 2nd edn. Cooper Publishing Group, Traverse City, Michigan
Ebenman B, Persson L (1988) Size-structured populations: ecology and evolution. Springer-Verlag, Berlin, Germany
Fujiwara M, Kendall BE, Nisbet RM (2004) Growth autocorrelation and animal size variation. Ecol Lett 7:106–113. doi:10.1046/j.1461-0248.2003.00556.x
Gårdmark A, Dieckmann U, Lundberg P (2003) Life-history evolution in harvested populations: the role of natural predation. Evol Ecol Res 5:239–257
Hartson RB, Kennedy BP (2015) Competitive release modifies the impacts of hydrologic alteration for a partially migratory stream predator. Ecol Freshw Fish 24(2):276–292
Höjesjö J, Johnsson J, Bohlin T (2002) Can laboratory studies on dominance predict fitness of young brown trout in the wild? Behav Ecol Sociobiol 52:102–108
Höjesjö J, Johnsson J, Bohlin T (2004) Habitat complexity reduces the growth of aggressive and dominant brown trout (Salmo trutta) relative to subordinates. Behav Ecol Sociobiol 56:286–289
Keeley ER (2001) Demographic responses to food and space competition by juvenile steelhead trout. Ecology 82:1247–1259
Kendall NW, McMillan JR, Sloat MR, Buehrens TW, Quinn TP, Pess GR, Kuzishchin KV, McClure MM, Zabel RW, Bradford M (2015) Anadromy and residency in steelhead and rainbow trout (): a review of the processes and patterns. Can J Fish Aquat Sci 72(3):319–342
Kenward MG, Roger JH (1997) Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 53:983–997
Lomnicki A (1999) Individual-based models and the individual-based approach to population ecology. Ecol Model 115:191–198
Martin-Smith KM, Armstrong JD (2002) Growth rates of wild stream-dwelling Atlantic salmon correlate with activity and sex but not dominance. J Anim Ecol 71:413–423
McGrath CL, Woods JA, Omernik JM, et al (2002) Ecoregions of Idaho (map scale 1:1,350,000)
Metcalfe NB (1998) The interaction between behavior and physiology in determining life history patterns in Atlantic salmon (Salmo Salar). Can J Fish Aquat Sci 55:93–103
Myrvold KM, Kennedy BP (2015a) Density dependence and its impact on individual growth rates in an age-structured stream salmonid population. Ecosphere 6:281
Myrvold KM, Kennedy BP (2015b) Local habitat conditions explain the variation in the strength of self-thinning in a stream salmonid. Ecol Evol 5:3231–3242. doi:10.1002/ece3.1591
Myrvold KM, Kennedy BP (2015c) Variation in juvenile steelhead density in relation to intream habitat and watershed characteristics. Trans Am Fish Soc 144:577–590
Myrvold KM, Kennedy BP (2015d) Interactions between body mass and water temperature cause energetic bottlenecks in juvenile steelhead. Ecol Freshw Fish 24:373–383
Nakano S (1995) Individual differences in resource use, growth and emigration under the influence of a dominance hierarchy in fluvial red-spotted masu salmon in a natural habitat. J Anim Ecol 64:75–84. doi:10.2307/5828
Nakano S, Kachi T, Nagoshi M (1991) Individual growth variation of red-spotted masu salmon, Oncorhynchus masou rhodurus, in a mountain stream. Japanese J Ichthyol 38:263–270
Nicieza AG, Metcalfe NB (1999) Costs of rapid growth: the risk of aggression is higher for fast-growing salmon. Funct Ecol 13:793–800
NMFS (2010) Endangered species act - section 7 formal consultation biological opinion and Magnuson-Stevens fishery conservation act essential fish habitat consultation for the operation and maintenance of the Lewiston orchards project. National Marine Fisheries Service, Seattle, Washington
Ohlberger J, Otero J, Edeline E et al (2013) Biotic and abiotic effects on cohort size distributions in fish. Oikos 122:835–844. doi:10.1111/j.1600-0706.2012.19858.x
Parra I, Almodovar A, Ayllon D et al (2012) Unravelling the effects of water temperature and density dependence on the spatial variation of brown trout (Salmo trutta) body size. Can J Fish Aquat Sci 69:821–832
Peacor SD, Pfister CA (2006) Experimental and model analyses of the effects of competition on individual size variation in wood frog (Rana sylvatica) tadpoles. J Anim Ecol 75:990–999. doi:10.1111/j.1365-2656.2006.01119.x
Peacor SD, Bence JR, Pfister CA (2007a) The effect of size-dependent growth and environmental factors on animal size variability. Theor Popul Biol 71:80–94. doi:10.1016/j.tpb.2006.08.005
Peacor SD, Schiesari L, Werner EE (2007b) Mechanisms of nonlethal predator effect on cohort size variation: ecological and evolutionary implications. Ecology 88:1536–1547
Pfister, CA (2003) Some consequences of size variability in juvenile prickly sculpin, Cottus asper. Env Bio Fishes 66:383–390
Pfister CA, Stevens FR (2002) The genesis of size variability in plants and animals. Ecology 83:59–72. doi:10.1890/0012-9658(2002)083[0059:TGOSVI]2.0.CO;2
Phillis CC, Moore JW, Buoro M, Hayes SA, Garza JC, Pearse DE (2016) Shifting Thresholds: Rapid Evolution of Migratory Life Histories in Steelhead/Rainbow Trout. J Hered 107(1):51–60
Quinn TP, Peterson NP (1996) The influence of habitat complexity and fish size on over-winter survival and growth of individually marked juvenile coho salmon (Oncorhynchus kisutch) in big beef creek, Washington. Can J Fish Aquat Sci 53:1555–1564
Raudenbush SW, Bryk AS (2002) Hierarchical linear models, 2nd edn. Sage Publications, Thousand Oaks, California
Reid D, Armstrong JD, Metcalfe NB (2011) Estimated standard metabolic rate interacts with territory quality and density to determine the growth rates of juvenile Atlantic salmon. Funct Ecol 25:1360–1367
Reid D, Armstrong JD, Metcalfe NB (2012) The performance advantage of a high resting metabolic rate in juvenile salmon is habitat dependent. J Anim Ecol 81:868–875
Ricker WE (1958) Handbook of computations for biological statistics of fish populations. Fish Res Board Canada Bull 119
Rose KA, Cowan JH, Winemiller KO, Myers RA, Hilborn R (2001) Compensatory density dependence in fish populations: importance, controversy, understanding and prognosis. Fish Fish 2(4):293–327
Rubenstein DI (1981) Individual variation and competition in the Everglades pygmy sunfish. J Anim Ecol 50:337–350
Satterthwaite WH, Beakes MP, Collins EM et al (2010) State-dependent life history models in a changing (and regulated) environment: steelhead in the California Central Valley. Evol Appl 3:221–243
Sloat MR, Reeves GH (2014) Individual condition, standard metabolic rate, and rearing temperature influence steelhead and rainbow trout (Oncorhynchus mykiss) life histories. Can J Fish Aquat Sci 71:491–501. doi:10.1139/cjfas-2013-0366
Sloat MR, Fraser DJ, Dunham JB et al (2014) Ecological and evolutionary patterns of freshwater maturation in Pacific and Atlantic salmonines. Rev Fish Biol Fish 24:689–707. doi:10.1007/s11160-014-9344-z
Thompson JN, Beauchamp DA (2016) Growth of juvenile steelhead Oncorhynchus mykiss under size-selective pressure limited by seasonal bioenergetic and environmental constraints. J Fish Biol 89:1720–1739. doi:10.1111/jfb.13078
Uchmański J (1999) What promotes persistence of a single population: an individual-based model. Ecol Model 115:227–241. doi:10.1016/S0304-3800(98)00179-3
Vøllestad LA, Quinn TP (2003) Trade-off between growth rate and aggression in juvenile coho salmon, Oncorhynchus kisutch. Anim Behav 66(3):561–568
Werner EE, Gilliam JF (1984) The ontogenetic niche and species interactions in size-structured populations. Annu Rev Ecol Evol Syst 15:393–425
Acknowledgements
We are grateful for the financial support from the United States Bureau of Reclamation, the University of Idaho Waters of the West Program, and United States Geological Survey. Thanks go to technicians E. Benson, N. Chuang, R. Hartson, and K. Pilcher and NSF-REU students J. Caisman, T. Kuzan, A. Merchant, J. Nifosi and T. Taylor for tremendous help obtaining the field data, and the Lewiston Orchards Irrigation District, Nez Perce Tribe, and private landowners for granting us access to their properties.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical approval
All sampling and handling procedures were permitted as part of the Section 7 consultation for the Lewiston Orchards Biological Opinion (NMFS 2010), and reviewed by the Idaho Department of Fish and Game and the University of Idaho Institutional Animal Care and Use Committee.
Rights and permissions
About this article

Cite this article
Myrvold, K.M., Kennedy, B.P. Size and growth relationships in juvenile steelhead: The advantage of large relative size diminishes with increasing water temperatures. Environ Biol Fish 100, 1373–1382 (2017). https://doi.org/10.1007/s10641-017-0649-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-017-0649-3