
Summary
DNA damage repair plays essential roles in drug resistance, especially resistance to Poly (ADP-ribose) polymerase (PARP) inhibitors in the clinic. A subset of DNA repair proteins such as Breast cancer gene 1 (BRCA1), BRCA2 and RecA homolog (RAD51) are client proteins of heat shock protein 90 (Hsp90). Clearance of these DNA repair proteins by inhibition of Hsp90 is a promising strategy for overcoming resistance to PARP inhibitors. Here we report the pharmacological analysis of the highly potent second-generation Hsp90 inhibitor, ganetespib. Methods Nuclear BRCA1, BRCA2, and RAD51 expression in breast cancer cells were detected by subcellular fractionation and western blot analysis. Formation of nuclear RAD51 and γ-H2AX foci was analyzed by immunofluorescent staining. The cytotoxicity of ganetespib and ABT-888 in breast cancer cells were evaluated by cell proliferation, colony survival, and apoptosis assay. To investigate the efficacy of this therapy in vivo, SCID mice bearing MCF7 xenografts were treated with ganetespib and ABT-888, both as single agents and in combination. Results Ganetespib significantly destabilized nuclear BRCA1, BRCA2, and RAD51, and efficiently disrupted homologous recombination-mediated DNA double-strand break repair in breast cancer cells. The synergistic antitumor effects of ganetespib and the PARP inhibitor, ABT-888 were observed, and concurrent treatment with both inhibitors synergistically inhibited xenograft tumor growth. Importantly, the combined treatment was well tolerated, without significant loss of body weight or major histological changes in the breast cancer xenograft model. Conclusion These data provide a novel strategy for the treatment of breast cancer with wild type BRCA1 using combination therapy targeting Hsp90 to overcome resistance to PARP inhibitors.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Poly(ADP-ribose) polymerase (PARP) are key molecules in the repair of DNA single-strand breaks (SSBs); inhibition of PARP enzymes results in the accumulation of SSBs which can lead to the formation of potentially lethal double-strand DNA breaks (DSBs) at replication forks during the S-phase [1]. PARP inhibitors have garnered significant attention as antitumor agents. Breast cancer gene 1 (BRCA1) and Breast cancer gene 2 (BRCA2) are important DNA repair proteins that are required for the effective repair of DNA DSBs by homologous recombination (HR). Tumor cells with HR deficiency or “BRCAness” are likely to be particularly sensitive to PARP inhibitors because they are unable to repair the collapsed forks and the resulting DSBs are cytotoxic [2–4]. Although a fraction of Breast cancer harbors HR defects that may sensitize the affected tumors to DNA-damaging agents, most breast cancer do not carry this defect, and as such, are insensitive to PARP inhibitors as monotherapies.
Hsp90 is an ATP-dependent molecular chaperone and is the most abundant intracellular protein in mammalian cells. It is essential for protein folding, assembly, and degradation processes. Hsp90 is expressed at higher levels in malignant cells compared to normal cells. It is required for the activation and stabilization of more than 200 client proteins involved in critical signaling pathways necessary for cellular proliferation including cell cycle progression, apoptosis, transcriptional regulation, chromatin remodeling, and DNA repair [5, 6].
Tanespimycin (17-N-allylamino-17-demethoxygeldanamycin, 17-AAG) is a first-generation heat shock protein 90 (Hsp90) inhibitor derived from the antibiotic geldanamycin. It exhibits indirect inhibitory effects on HR function by downregulating the protein levels of BRCA1, BRCA2, and RecA homolog (RAD51), suggesting that the combined use of Hsp90 inhibitors with PARP inhibitors or DNA damaging agents may be particularly effective [7, 8]. However, due to its relatively low efficacy and high liver toxicity, 17-AAG could not be fully developed as a therapeutic agent. Ganetespib is a novel, small molecule, second-generation Hsp90 inhibitor with better pharmacological properties and safety profiles than 17-AAG. In addition, it has demonstrated preclinical activity against Non Small Cell Lung Cancer (NSCLC) in animal models, including those driven by mutant EGFR, rearranged ALK, and/or mutant KRAS [9–12]. In clinical trials, ganetespib monotherapy showed a manageable side effect profile as well as clinical activity in heavily pretreated patients with advanced NSCLC, particularly in patients with tumors harboring ALK gene rearrangement [13].
In this study, we investigated the effects of ganetespib alone and in combination with PARP inhibitors both in vitro and in vivo in breast cancer models. We demonstrate that ganetespib was effective in downregulating the protein levels of BRCA1, BRCA2 and RAD51. In addition, ganetespib potentiated PARP inhibitor-induced cell death, in part by indirectly inhibiting HR function.
Materials and methods
Cell culture, chemical reagent, and irradiation treatments
MCF7 and MDA-MB-231were purchased from the Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China). Certification of cell line authenticity was provided. Cells were maintained in basic DMEM supplemented with 10% fetal bovine serum (FBS) (16000–044, GIBCO), 100 U penicillin, and 100 mg/mL streptomycin (Thermo Fisher Scientific). Cell lines were subjected to DNA profiling annually (Short Tandem Repeat Analysis) at the Center for Medicolegal Expertise of Sun Yat-Sen University (Guangdong, China) to confirm their authenticity. For irradiation of cells, the Siemens ONCOR Impression Plus linear accelerator (6 MV) X-ray (Munich, Germany) was used at a dose rate 2 Gy/min. Control cells were mock irradiated. Ganetespib (STA-9090) was provided by Synta Pharmaceuticals (Lexington), and Veliparib (ABT-888) were purchased from Selleck (Shanghai, China). Each of the inhibitors was dissolved in dimethyl sulfoxide (DMSO) at a final concentration of 10 mM, aliquoted, and stored at −80 °C.
Antibodies
The antibodies used were anti-BRCA1 (Ab-1) mouse mAb (MS110) (Cat No. OP92; Calbiochem), anti-BRCA2 (ab27976; Abcam), phospho-Histone H2AX (Ser139) (2577; Cell Signaling), HSP90 (4874; Cell Signaling), GAPDH, (sc-25778; Santa Cruz), Histone H1 (AE-4), (sc-8030; Santa Cruz), anti-PARP (ab110915; Abcam), and RAD51 (H-92) (sc-8349; Santa Cruz). The secondary antibodies used were goat anti-mouse IgG-HRP, (sc-2005; Santa Cruz), goat anti-rabbit IgG-HRP, (sc-2004; Santa Cruz), anti-rabbit IgG(H + L) ab2 Fragment (Alexa Flour 488 conjugated, 4412S; Cell Signaling)
Cell proliferation, colony survival, and apoptosis assay
Cells were digested and re-plated into 96 well plates with density of 2000 cells/well in 100 μl medium. 16 h later the cells were then treated with 6 point concentrations of ganetespib (with 2 fold dilutions starting from 50nM) or ABT-888 (with 2 fold dilutions starting from 10uM) or with a combination of ganetespib and ABT-888 (fixed concentration ratio of 1:200) for 72 h. Cell number were counted using Cell Counting Kit-8(CCK-8) (DOJINDO, Japan) according to instructions. Each experimental condition was performed in triplicate and repeated at least three times. All values were normalized with respect to the viability of untreated cells.
For Colony formation assays, single cells were obtained by Trypsin-EDTA digestion and cell numbers were counted. Then the cells were re-plated into 6 well plates with density of 100–5000 cells/well. After 16 h when cells are attached to the plate, cells were treated by drugs or vehicle control as mentioned in Fig. 3. 72 h after the combined treatment, the medium was replaced with fresh medium without ganetespib and ABT-888. 12 days later, the colony were fixed by 75% ethanol and stained by 1% crystal violet and were counted. Survival fraction was calculated as:
(Number of colonies / number of cells plated) / (number of colonies for corresponding control / number of control cells plated).
For analysis of apoptosis, MCF7 cells were treated with Vehicle, ganetespib or ABT-888 alone or in combination of Ganetespid with ABT-888. 48 h post treatment, cells were harvested, and apoptotic cells were stained using the Annexin V-FITC Apoptosis Detection kit (BD PharMingen, San Diego, CA). The percentage of apoptotic cells was measured by flow cytometry.
Immunofluorescent staining
Immunofluorescence staining was performed as described previously [14]. Briefly, cells were grown on glass cover slips into 24-well plates and left to attach for 16 h. Then the cells were treated with ganetespib, ABT-888 or irradiated (or mock irradiated) at indicated doses and times. Cells were washed with PBS twice and fixed with ice cold methanol for 20 min. Cells were permeabilized for 10 min with 0.5%Triton X-100 (TX)/PBS and blocked by 2%BSA/0.1%TX/PBS for 40 min at RT. Then cover slips were incubated with primary antibody diluted with 2%BSA/0.1%TX/PBS for overnight at 4 °C. Slides were washed and incubated for 1 h with secondary antibody (1:1000 goat anti-mouse Alexa 488-conjugated antibody), stained with DAPI for 1–2 min, mounted with mounting medium, and analyzed by fluorescence microscopy.
Subcellular fractionation and western blot analysis
For nuclear and cytoplasmic protein extraction, cells were subjected to subcellular fractionation, according the manual of manufactory of Nuclear Extract Kit (Active Motif, Cat No.:40010, Shanghai China). Protein expression in each component was detected by Western blot. As a control for the purity of fractionation, histone H1 (nuclear) and GAPDH (cytosol) expression levels were also analyzed.
For western blot analysis, cell lysates were prepared using radioimmunoprecipitation assay (RIPA) buffer (150 mM NaCl, 50 mM Tris, pH 8.0, 5 mM EDTA, 0.5% sodium deoxycholate, 0.1% SDS, 1.0% Nonidet P-40) with protease and phophatase inhibitor cocktails (Sigma-Aldrich, St. Louis, MO) and subjected to SDS-PAGE analysis. GAPDH was used as a loading control in western blots.
In vivo tumor growth inhibition
Female (7 to 8 weeks old) immunodeficient SCID mice were purchased from SLAC laboratory animal company (Shanghai, China). Mice were maintained in a pathogen-free environment, and all in vivo procedures were approved by Guangzhou Medical University Animal Care and Use Committee. MCF7 cells (1 × 106 cells) were subcutaneously implanted into the animals. Mice bearing established tumors (100–200 mm3) were randomized into four treatment groups (n = 7). Animals were treated with a 50 mg/kg weekly dose of ganetespib or a 20 mg/kg daily dose of ABT-888, either alone or in combination. Intravenous administration with either vehicle or ganetespib (formulated in 10/18 DRD: 10% DMSO, 18% Cremophor RH 40, 3.6% dextrose, 68.4% water) was via the tail vein [11]. ABT-888 was diluted in vehicle solution (2.1 g sorbitol, 0.1 g citric acid, monohydrate, purified water) to a final concentration of 10 mg/ml, and 20 mg/kg ABT-888 was administered daily by oral gavage. The body weight of the mice was monitored every day. Tumor sizes were measured every 3 days in three dimensions using an electronic caliper, and tumor volumes (V) were calculated by caliper measurement of width (W), length (L), and thickness (T) as follows: V = 0.5236 (LWT). Mice were euthanized and sacrificed 2 days after the fifth dose of ganetespib. The organs including heart, lung, stomach, intestine, bowl, liver, and kidney were fixed in formalin and hematoxylin and eosin (H&E)-stained slides were examined under the microscope.
In vivo protein degradation assay
For the in vivo protein degradation assay, when established tumors were palpable (~200 mm3), animals were treated with a single bolus injection of ganetespib at a dose of 50 mg/kg. Mice were sacrificed and tumors were harvested 24, 48, and 72 h after treatment. For comparison, animals were treated with a single injection of vehicle and tumors were collected after 24 h. Tumor tissues were homogenized in RIPA buffer, after which the lysates were subjected to Western blotting.
Statistical analysis
The data were analyzed via analysis of variance (ANOVA) followed by a Bonferroni post-hoc test using GraphPad Prism 6 for windows. Data are presented as the mean and standard error of mean.
Results
Ganetespib significantly destabilized nuclear BRCA1, BRCA2, and RAD51 in breast cancer cells
We inverstigated whether ganetespib downregulates the protein levels of BRCA1, BRCA2, RAD51 and PARP. As shown in Fig. 1a and b, ganetespib downregulated BRCA1, BRCA2, and RAD51 in a dose- and time-dependent manner. However, it did not induce significant downregulation of PARP. A concomitant increase in Hsp90 levels was observed, indicative of Hsp90 inhibition.

Effect of Hsp90 inhibition in whole cell lysates and subcellular expression of key DNA repair proteins in breast cancer cells. A, MCF7 cells were exposed to graded concentrations of ganetespib (12.5–200 nM) for 24 h and cell lysates were immunoblotted with the indicated antibodies. B, MCF7 cells were treated with ganetespib (50 nM) at the indicated time points between 12 and 48 h. C, Subcellular localization of BRCA1, BRCA2, RAD51 and PARP after treatment with vehicle or 50 nM ganetespib
The repair of DNA DSBs requires that BRCA1 and other DNA repair proteins are present in the nucleus [15]. Then we tested whether ganetespib induces degradation of nuclear BRCA1, BRCA2, RAD51, and PARP proteins. MCF7 cells were treated with vehicle or 50 nM ganetespib for 24 h, and protein expression in nuclear and cytoplasmic compartments was assessed by subcellular fractionation. As shown in Fig. 1c BRCA1, BRCA2, and RAD51 were localized in both the nucleus and cytoplasm, and ganetespib downregulated the protein levels of BRCA1,BRCA2, and RAD51 in both cell compartments; The PARP protein was mainly localized in the nucleus, and ganetespib had almost no effects on its nuclear expression. Together, these data suggest that Hsp90 inhibition by ganetespib efficiently decreases the expression of DNA repair proteins in the nucleus, which may significantly inhibit their function.
Pretreatment of ganetespib altered DSB repair capacity in breast cancer cells treated with ionizing radiation and PARP inhibition
Given that ganetespib can degrade several central components of DNA DSB repair proteins, we assessed whether ganetespib pretreatment altered DSB repair capacity in breast cancer cells treated with ionizing radiation (IR). First, we analyzed RAD51 foci, a well-established functional marker of HR repair activity. When a DNA DSB occurs, RAD51 foci form at the sites of single-stranded DNA in lesions that promote HR [16]. In this study, MCF7 cells were pretreated with vehicle or 25 nM ganetespib for 24 h, and cells were mock irradiated or irradiated with 4 Gy IR. RAD51 foci levels were analyzed by immunofluorescence 2, 8, and 24 h after IR treatment. As shown in Fig. 2a, at 2 h after 4 Gy IR, about 47% of cells was RAD51 foci-positive without ganetespib treatment, which significantly decreased to about 13% after ganetespib treatment. Similar results were found 8 h after 4 Gy IR. Representative data of three independent experiments are shown as the percent of cells with Rad51 foci, and a representative stain of cells exhibiting RAD51 foci is shown in Fig. 2a.

Pretreatment of ganetespib reduces DNA repair after DNA damage. MCF7 cells were pretreated with vehicle or 25 nM ganetespib for 24 h, after which cells were mock irradiated or irradiated with 4Gy IR. RAD51 (A) and γ-H2AX (B) levels were analyzed by immunofluorescence 2, 8, and 24 h after IR. Shown is the representative data of three independent experiments of the percent of cells (mean ± SEM) with RAD51 or γ-H2AX foci, Inset, a representative staining of cell exhibiting RAD51 foci or γ-H2AX with the nucleus stained. C, MCF7 cells were treated with vehicle, ganetespib, and ABT-888 alone, or in combination at the indicated dose, and γ-H2AX levels were observed 12, 24, and 48 h after treatment. (*, P < 0.05; **, P < 0.01)
Next, we determined if compromised DNA repair by ganetespib would result in increased levels of persistent DSBs. To this end we assessed the accumulation of γ-H2AX foci, a marker of unrepaired DSBs [17], in MCF7 cells treated with ganetespib and 4 Gy IR. As shown in Fig. 2b, at 8 h and 24 h after 4 Gy IR, the amount of unrepaired DSBs significantly decreased in a time-dependent manner in cells without ganetespib treatment, and only modestly decreased in cells with ganetespib treatment. These data indicate that ganetespib pretreatment decreases HR-mediated DSB repair, resulting in the increased accumulation of unrepaired DSBs in MCF7 cells. Representative data of three independent experiments are shown as the percent of cells with γ-H2AX foci. A representative stain of cells exhibiting γ-H2AX foci is shown in Fig. 2b.
As mentioned above, PARP plays a key role in the repair of SSBs, which in dividing cells, are ultimately converted to DSBs that are repaired by HR-mediated repair machinery. Since ganetespib degrades several key components of the HR-mediated repair of DSBs, we reasoned that the combination of ganetespib and ABT-888 would result in additional DNA DSB damage. To evaluate this, we performed a time course analysis of γ-H2AX foci with vehicle, ganetespib, and ABT-888 alone or in combination. As shown in Fig. 2c ABT-888 alone only modestly enhanced formation of γ-H2AX foci; however, there was marked increase of γ-H2AX foci in cells treated with a combination of ganetespib and ABT-888 in a time-dependent manner. These data suggest that ABT-888 significantly increases DNA DSB damage when combined with ganetspib.
Combination of ganetespib with ABT-888 enhances cytotoxicity in breast cancer cells
We evaluated the combined drug interaction according to the median-effect method of Chou and Talalay. The combination index (CI) was calculated using the CompuSyn Software (www.combosyn.com) [18]. A CI of 1 indicates an additive drug interaction, a CI of more than 1 indicates an antagonistic interaction, and a score lower than 1 suggests a synergistic interaction. To examine whether the schedule of administration modulated efficacy, three types of sequential combinations were evaluated. For the first sequential combination, cells were treated with ganetespib for 16 h after which ABT-888 was added. For the second sequential combination, cells were simultaneously treated with ganetespib and ABT-888. For the third sequential treatment, cells were treated with ABT-888 for 16 h after which ganetespib was added. Then 72 h after the combined treatment, the proliferation of cells was assayed. A constant concentration ratio (1:200) of ganetespib and ABT-888 was used. As shown in Table 1, the schedule of administration had substantial effects on cytotoxicity of MCF7 cells. The first sequential combination resulted in markedly synergistic effects for all doses used (lower values of CIs).
Since maximal efficacy was observed in cells pretreated with ganetespib for 16 h, this schedule of administration was used to further evaluate the CI value of combination treatment in other breast cancer cell line MDA-MB-231. As shown in Table 1, similar results were obtained in this cell line.
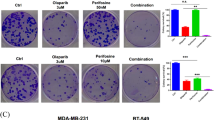
To further determine the synergistic effects of combined ganetespib and ABT-888 treatment on cell proliferation, we performed the colony formation assay. As shown in Fig. 3a and b, ABT-888 alone failed to have any cytotoxic effects on the Breast cancer cells MCF7 and MDA-MB-231, whereas those pretreated with ganetespib had increased dose-dependent cytotoxicity to ABT-888. In general, the synergistic effects were observed in the cell proliferation and colony survival assays, and the higher doses of ganetespib and ABT-888 resulted in more points of synergism for all of the cell lines, especially when ganetespib was administered before ABT-888.

Ganetespib enhanced cytotoxicity of the PARP inhibitor. Cells were pretreated with various doses of ganetespib for 16 h, after which different doses of ABT-888 were added. 72 h after the combined treatment, the media was replaced with fresh media lacking ganetespib and ABT-888. 12 days later, cell colonies were fixed and stained. Shown is the representative data of at least three independent experiments (mean ± SEM). A, MCF7, B, MDA-MB-231
Enhanced cytotoxicity of the PARP inhibitor by ganetespib is mediated by the apoptosis pathway
To investigate whether cellular apoptosis is involved in ganetespib-mediated cell cytotoxicity in Breast cancer cells, we evaluated the activation of apoptotic pathways in MCF7 cells treated with ganetespib and ABT-888 by assessing cleaved caspase 3, caspase 9, and annexin V. As shown in Fig. 4a, increased caspase 3 and caspase 9 cleavage was observed in cells treated with ganetespib alone or combined with ABT-888. No significant difference was observed in cells treated with ABT-888 alone. The reduction in ganetespib-induced apoptosis was verified with annexin V staining. As shown in Fig. 4b and c, activation of apoptosis was significantly greater in both MCF7 and MDA-MB-231 cells upon combined treatment of ganetespib and ABT-888 compared to treatment with either agent alone (P < 0.01). Together, these results suggest that apoptosis contributes to the combined effects of ganetespib and ABT-888 on inhibition of DNA damage repair.

Enhanced cytotoxicity of PARP inhibitor by ganetespib is driven by apoptosis. A, Combined inhibition of Hsp90 and PARP increased levels of cleaved caspase 3 and caspase 9 in MCF7 cells. MCF7 cells were treated with ganetespib alone or a combination of ganetespib and ABT-888 for 48 h after which cell lysates were immunoblotted with the indicated antibodies. B and C, Combined inhibition of Hsp90 and PARP increased the number of apoptotic cells in MCF7, and MDA-MB-231 cells. B, a representative flow cytometry analysis of apoptosis after annexin V staining. C, Shown is the representative data of at least three independent experiments (mean ± SEM). (**, P < 0.01)
Ganetespib degrades DNA repair proteins and inhibits tumor growth in vivo
Given that ganetespib can down regulate several important DNA repair proteins such as BRCA1, BRCA2, and RAD51 in vitro, we tested whether it could degrade these proteins in MCF7 xenograft tumors in SCID mice. As shown in Fig. 5a, the protein expression of BRCA1, BRCA2, and RAD51 was reduced within 24 h of ganetespib treatment. Importantly, these effects were sustained over time, as recovery did not occur until 72 h.

Hsp90 inhibition augments PARP inhibitor efficacy in tumor growth in vivo, without significant effects on body weight or significant pathology changes. A. Ganetespib inhibited BRCA1, BRCA2 and RAD51 expression in a xenograft model of breast cancer. SCID mice bearing MCF7 xenografts received a single dose of vehicle or ganetespib (50 mg/kg). At 24, 48, and 72 h, lysates from harvested xenografts (n = 3 per group) were subjected to Western blotting with the indicated antibodies. B, Ganetespib combined with the PARP inhibitor ABT-888 inhibited tumor growth. SCID mice bearing MCF7 xenografts were randomized into four treatment groups (n = 7): control, i.v. dosed with ganetespib (50 mg/kg) once weekly for consecutive 5 weeks, oral dosed with ABT-888 (20 mg/kg) daily, ganetespib + ABT-888. Tumor size was measured every 3 days. Shown is the tumor volume (mean ± SEM) over time. The combination treatment of ganetespib and ABT-888 induced a significant delay in tumor growth compared to either agent alone. (**, P < 0.01). C, For mice treated in B, the average change in body weight was followed over the treatment time course, without significant loss of body weight was observed. D, Shown is the representative picture of eosinophilic changes at scattered areas of the liver for mice treated in B
To evaluate whether the combined benefits on cell viability detected in vitro translated to improved efficacy in vivo, mice bearing MCF7 xenografts were treated with 50 mg/kg ganetespib and ABT-888, both as single agents and in combination. It was previously reported that the highest non-toxic dose of ganetespib that can be given weekly is 150 mg/kg [19]. We selected a low ganetespib dose (one-third of this dose) for these experiments in order to readily permit evaluation of potential combinatorial improvements in efficacy between the two drugs. As shown in Fig. 5b, As a single agent, weekly administration of a suboptimal dose of ganetespib (50 mg/kg) or daily dosing with ABT-888 (20 mg/kg) resulted in only modest antitumor activity, however, concurrent treatment with both agents resulted in a significant enhancement of antitumor activity, which is consistent with our in vitro findings. These results indicate that the combination of ganetespib with ABT-888 results in a superior therapeutic response compared to the single-agent activity of each compound alone.
No significant loss of body weight or histology changes were induced by ganetespib
To confirm whether any severe side effects were caused by the concurrent treatment of ganetespib and ABT-888, the body weight of mice was monitored daily. Mice treated with ganetespib alone or with both ganetespib and ABT-888 suffered from diarrhea within 2 days of dosing of ganetespib, and about 5% body weight loss accompanied those symptoms. On the third day, the symptoms subsided and the body weight gradually increased. No significant difference in average body weight was observed between the four experimental groups after treatments with five doses of ganetespib (Fig. 5c). After the mice were sacrificed at the end of the experiments, organs including the heart, lung, stomach, intestine, bowl, liver, and kidney were fixed in formalin, and H&E-stained slides were examined under a microscope. Scattered eosinophils in the liver were observed in one of seven mice treated with ganetespib alone or with both ganetespib and ABT-888. A representative picture is shown in Fig. 5d. Histological changes were not observed in the vehicle control and ABT-888 treatment groups. In addition, no specific changes were observed in the tissues of other organs. In summary, the combination treatment of ganetespib and ABT-888 was well tolerated and no significant loss of body weight or significant histological changes were observed.
Discussion
PARP inhibitors as potential anti-cancer drugs have been tested in clinical trials [20]. Tumor cells with an HR deficiency or “BRCAness” are likely to be particularly sensitive to PARP inhibitors [2, 3]. As specific HR inhibitors are not available, we demonstrated that the second-generation Hsp90 inhibitor, ganetespib, downregulates BRCA1, BRCA2, and RAD51, key regulators of HR, and enhances the efficacy of the PARP inhibitor, ABT-888, by blocking DNA repair. Ganetespib and ABT-888 work synergistically in vitro and in mouse xenograft models.
Ganetespib is a novel small molecule, second-generation Hsp90 inhibitor with improved pharmacological properties and safety profiles compared to first-generation Hsp90 inhibitors. Our data shows that ganetespib effectivly destabilizes key DNA repair proteins. We found that BRCA1, BRCA2, and RAD51, were localized in both the nucleus and cytoplasm, and ganetespib downregulated the protein expression of BRCA1, BRCA2 and RAD51 in both compartments. However, PARP is mainly localieze in nucleus, and ganetespib had almost no effects on the nuclear proteins PARP. These data suggest that for some Hsp90 client proteins, both cytoplasmic and nuclear proteins are sensitive to ganetespib, which is not the case for all protein, although the underlying basis of resistance is not known.
Consistent with the fact that ganetespib can degrade several central components of the DNA DSB repair proteins, we confirmed that ganetespib pretreatment altered the DSB repair capacity of MCF7 cells by analyzing RAD51 and γ-H2AX foci formation after IR treatment. Studies from our group and others have shown that upstream proteins, which promote assembly of subnuclear RAD51 foci, are sensitive to ganetespib [14, 21], and depletion of RAD51 and upstream factors may account for the decreased RAD51 foci and subsequent impaired DSB repair [22].
PARP plays a key role in the repair of SSBs. In dividing cells, inhibition of PARP leads to the accumulation of unrepaired SSBs which are converted into DSBs at replication forks, allowing subsequent repair by unaffected HR mechanisms [23]. Cells treated with ABT-888 alone did not have significantly enhanced DSB formation. However HR deficiency caused by pretreatment of ganetespib inhibited the repair of DSBs, which manifested as persistent γ-H2AX foci. Since ganetespib only had modest effects on PARP protein expression, sensitization of cells to cytoxicity of the PARP inhibitor is not through downregulation of PARP protein, but rather, is through downregulation of other DSB repair proteins.
The combination of ganetespib with ABT-888 enhanced cytotoxicity in MCF7 and MDA-MB-231 cells, and this effect was affected by sequential treatment. Specifically, when ganetespib was administered 16 h earlier than ABT-888, the best synergistic effects occurred, suggesting that the combination treatment will be most effective when ganetespib is added first to induce HR deficiency. These data are consistent with a report that exposure to 17-DMAG for 16 h enhances the radiosensitivity of MiaPaca human pancreatic cancer cells [24]. Thus it is clear that the timing and sequence of treatment are important in order to achieve ganetespib-induced tumor sensitization to PARP inhibition.
The combined benefits on cell viability observed in vitro also translated to improved efficacy in vivo. When ganetespib or ABT-888 are administered as monotherapies, a modest degree of tumor growth inhibition occurred. However, concurrent treatment with both drugs resulted in a significantly improved antitumor response. Most importantly, the combination treatment was well tolerated, with the exception of some minor changes to the liver such as a eosinophilic changes in scattered areas of the liver. These findings are consistent with the clinical safety profile of ganetespib, which only showed a minor incidence of visual disorders in more than 800 patients [25, 26]. We confirmed that the in vivo toxicity of ganetespib was acceptable, and the addition of ABT-888 did not increase toxicity.
Conclusion
Our preclinical data suggest the ganetespib can be combined with PARP inhibitors to inhibit breast cancer tumor growth by targeting DNA repair proteins. No additional systemic toxicity was observed in the in vivo mouse model. The data presented here provide strong preclinical support for the exploration of this combination treatment as a novel therapeutic approach in patients suffering from breast cancer with wild type BRCA1.
References
de Murcia JM, Niedergang C, Trucco C, Ricoul M, Dutrillaux B, Mark M et al (1997) Requirement of poly(ADP-ribose) polymerase in recovery from DNA damage in mice and in cells. Proc Natl Acad Sci U S A 94(14):7303–7307
Bryant HE, Schultz N, Thomas HD, Parker KM, Flower D, Lopez E et al (2005) Specific killing of BRCA2-deficient tumours with inhibitors of poly(ADP-ribose) polymerase. Nature 434(7035):913–917
Rottenberg S, Jaspers JE, Kersbergen A, van der Burg E, Nygren AO, Zander SA et al (2008) High sensitivity of BRCA1-deficient mammary tumors to the PARP inhibitor AZD2281 alone and in combination with platinum drugs. Proc Natl Acad Sci U S A 105(44):17079–17084
Gudmundsdottir K, Ashworth A (2006) The roles of BRCA1 and BRCA2 and associated proteins in the maintenance of genomic stability. Oncogene 25(43):5864–5874
Li J, Buchner J (2013) Structure, function and regulation of the hsp90 machinery. Biomed J 36(3):106–117
Banerji U (2009) Heat shock protein 90 as a drug target: some like it hot. Clin Cancer Res 15(1):9–14
Stecklein SR, Kumaraswamy E, Behbod F, Wang W, Chaguturu V, Harlan-Williams LM et al (2012) BRCA1 and HSP90 cooperate in homologous and non-homologous DNA double-strand-break repair and G2/M checkpoint activation. Proc Natl Acad Sci U S A 109(34):13650–13655
Dungey FA, Caldecott KW, Chalmers AJ (2009) Enhanced radiosensitization of human glioma cells by combining inhibition of poly(ADP-ribose) polymerase with inhibition of heat shock protein 90. Mol Cancer Ther 8(8):2243–2254
Acquaviva J, Smith DL, Sang J, Friedland JC, He S, Sequeira M et al (2012) Targeting KRAS-mutant non-small cell lung cancer with the Hsp90 inhibitor ganetespib. Mol Cancer Ther 11(12):2633–2643
Shimamura T, Perera SA, Foley KP, Sang J, Rodig SJ, Inoue T et al (2012) Ganetespib (STA-9090), a nongeldanamycin HSP90 inhibitor, has potent antitumor activity in in vitro and in vivo models of non-small cell lung cancer. Clin Cancer Res 18(18):4973–4985
Sang J, Acquaviva J, Friedland JC, Smith DL, Sequeira M, Zhang C et al (2013) Targeted inhibition of the molecular chaperone Hsp90 overcomes ALK inhibitor resistance in non-small cell lung cancer. Cancer Discov 3(4):430–443
Proia DA, Sang J, He S, Smith DL, Sequeira M, Zhang C et al (2012) Synergistic activity of the Hsp90 inhibitor ganetespib with taxanes in non-small cell lung cancer models. Investig New Drugs 30(6):2201–2209
Socinski MA, Goldman J, El-Hariry I, Koczywas M, Vukovic V, Horn L et al (2013) A multicenter phase II study of ganetespib monotherapy in patients with genotypically defined advanced non-small cell lung cancer. Clin Cancer Res 19(11):3068–3077
Bhattacharyya A, Ear US, Koller BH, Weichselbaum RR, Bishop DK (2000) The breast cancer susceptibility gene BRCA1 is required for subnuclear assembly of Rad51 and survival following treatment with the DNA cross-linking agent cisplatin. J Biol Chem 275(31):23899–23903
Jiang J, Yang ES, Jiang G, Nowsheen S, Wang H, Wang T et al (2011) p53-dependent BRCA1 nuclear export controls cellular susceptibility to DNA damage. Cancer Res 71(16):5546–5557
Yang ES, Nowsheen S, Rahman MA, Cook RS, Xia F (2012) Targeting BRCA1 localization to augment breast tumor sensitivity to poly(ADP-ribose) polymerase inhibition. Cancer Res 72(21):5547–5555
Sedelnikova OA, Rogakou EP, Panyutin IG, Bonner WM (2002) Quantitative detection of (125)IdU-induced DNA double-strand breaks with gamma-H2AX antibody. Radiat Res 158(4):486–492
Ashton JC (2015) Drug combination studies and their synergy quantification using the Chou-Talalay method--letter. Cancer Res 75(11):2400
Ying W, Du Z, Sun L, Foley KP, Proia DA, Blackman RK et al (2012) Ganetespib, a unique triazolone-containing Hsp90 inhibitor, exhibits potent antitumor activity and a superior safety profile for cancer therapy. Mol Cancer Ther 11(2):475–484
Tutt A, Robson M, Garber JE, Domchek SM, Audeh MW, Weitzel JN et al (2010) Oral poly(ADP-ribose) polymerase inhibitor olaparib in patients with BRCA1 or BRCA2 mutations and advanced breast cancer: a proof-of-concept trial. Lancet 376(9737):235–244
Suhane T, Laskar S, Advani S, Roy N, Varunan S, Bhattacharyya D et al (2015) Both the charged linker region and ATPase domain of Hsp90 are essential for Rad51-dependent DNA repair. Eukaryot Cell 14(1):64–77
Ko JC, Chen HJ, Huang YC, Tseng SC, Weng SH, Wo TY et al (2012) HSP90 inhibition induces cytotoxicity via down-regulation of Rad51 expression and DNA repair capacity in non-small cell lung cancer cells. Regul Toxicol Pharmacol 64(3):415–424
Davar D, Beumer JH, Hamieh L, Tawbi H (2012) Role of PARP inhibitors in cancer biology and therapy. Curr Med Chem 19(23):3907–3921
Dote H, Burgan WE, Camphausen K, Tofilon PJ (2006) Inhibition of hsp90 compromises the DNA damage response to radiation. Cancer Res 66(18):9211–9220
Proia DA, Bates RC (2014) Ganetespib and HSP90: translating preclinical hypotheses into clinical promise. Cancer Res 74(5):1294–1300
Ramalingam S, Goss G, Rosell R, Schmid-Bindert G, Zaric B, Andric Z et al (2015) A randomized phase II study of ganetespib, a heat shock protein 90 inhibitor, in combination with docetaxel in second-line therapy of advanced non-small cell lung cancer (GALAXY-1). Ann Oncol 26(8):1741–1748
Acknowledgements
We thanks Synta Pharmaceuticals (Lexington) for providing Ganetespib (STA-9090).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interests.
Funding
This work was supported by the National Natural Science Foundation of China (No. 81272901 to Juhong Jiang and No. 81372298 to Yuanzhi Lu).
Ethics approval
We declare that all experiments were performed in compliance with all laws of China.
Informed consent
For this type of study, formal consent is not required.
Additional information
Juhong Jiang and Yuanzhi Lu contributed equally
Rights and permissions
About this article

Cite this article
Jiang, J., Lu, Y., Li, Z. et al. Ganetespib overcomes resistance to PARP inhibitors in breast cancer by targeting core proteins in the DNA repair machinery. Invest New Drugs 35, 251–259 (2017). https://doi.org/10.1007/s10637-016-0424-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10637-016-0424-x