
Abstract
Background and Aims
Bioartificial livers (BALs) have attracted much attention as potential supportive therapies for liver diseases. A serum-free microcarrier culture strategy for the in vitro high-density expansion of human-induced hepatocyte-like cells (hiHeps) suitable for BALs was studied in this article.
Methods
hiHeps were transdifferentiated from human fibroblasts by the lentiviral overexpression of FOXA3, HNF1A, and HNF4A. Cells were cultured on microcarriers, their proliferation was evaluated by cell count and CCK-8 assays, and their function was evaluated by detecting liver function parameters in the supernatant, including urea secretion, albumin synthesis, and lactate dehydrogenase levels. The expressions of hepatocyte function-associated genes of hiHeps were measured by qRT-PCR in 2D and 3D conditions. The expression of related proteins during fibronectin promotes cell adhesion, and proliferation on microcarrier was detected by western blotting.
Results
During microcarrier culture, the optimal culture conditions during the adherence period were the use of half-volume high-density inoculation, Cytodex 3 at a concentration of 3 mg/mL, a cell seeding density of 2.0 × 105 cells/mL, and a stirring speed of 45 rpm. The final cell density in self-developed, chemically defined serum-free medium (SFM) reached 2.53 × 106 cells/mL, and the maximum increase in expansion was 12.61-fold. In addition, we found that fibronectin (FN) can promote hiHep attachment and proliferation on Cytodex 3 microcarriers and that this pro-proliferative effect was mediated by the integrin-β1/FAK/ERK/CyclinD1 signaling pathway. Finally, the growth and function of hiHeps on Cytodex 3 in SFM were close to those of hiHeps on Cytodex 3 in hepatocyte maintenance medium (HMM), and cells maintained their morphology and function after harvest on microcarriers.
Conclusions
Serum-free microcarrier culture has important implications for the expansion of a sufficient number of hiHeps prior to the clinical application of BALs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The liver, the largest and most substantial organ in the body, has a very complex structure and carries out important physiological functions [1]. Currently, various liver diseases caused by liver injury are increasingly threatening human health and survival [2,3,4]. Over recent decades, orthotopic liver transplantation (OLT) has been the most effective way to treat various end-stage liver diseases [5]. However, due to a severe shortage of available donor organs, bioartificial livers (BALs) have attracted much attention and serve as potential supportive therapies for the treatment of patients awaiting tissue regeneration [6, 7].
BALs system is composed of hepatocytes, bioreactors, and extracorporeal circulation devices that play an essential role in detoxification. The core of the BAL system is functional hepatocytes cultured in vitro at a high density with high activity [8]. At present, primary human hepatocytes, cryopreserved porcine hepatocytes, and an immortalized human hepatocyte line are the most frequently used hepatocytes in the BAL system. However, the poor proliferative capacity of human primary hepatocytes in vitro, risk of infectious zoonotic diseases from porcine hepatocytes, and limited liver function of the immortalized human hepatocyte line limit their widespread application in BALs [9, 10]. Some recent studies have established human-induced hepatocyte-like cells through transdifferentiation technology, which is expected to solve the problems with hepatocyte sources in BALs [11].
Human-induced hepatocyte-like cells (hiHeps) are transdifferentiated from human skin fibroblasts by the lentiviral expression of FOXA3, HNF1A, and HNF4A with SV40 large T. The resultant hiHeps share numerous functions, such as biliary drug clearance, cytochrome P450 enzymatic activity, and the synthesis of urea and albumin, with mature hepatocytes [12]. In addition, hiHeps display a typical epithelial phenotype and are capable of expansion in vitro, thereby serving as an important cell source for BAL treatment [12]. Typically, at least 1010 functional hepatocytes are required for BAL-based ALF clinical treatment, and the expansion of these cells must be performed in serum-free medium (SFM) in vitro prior to their clinical application [13, 14]. In a previous two-dimensional (2D) plate culture process, we achieved hiHep proliferation and the stable expression of specific genes during the long-term passage of hiHeps in a self-developed, chemically defined SFM [15]. Although it is theoretically possible to generate 1010 functional hiHeps in vitro in SFM, this method not only does not utilize space effectively but also has a high cost, and it would be difficult to obtain enough hiHeps for BALs in a short time. Therefore, it has become necessary to consider alternative systems that allow a larger specific surface area and exhibit higher productivity, and microcarriers have been used to culture and expand adherent cells.
The microcarrier culture technique is a common high-density cell culture procedure with advantages including a large specific surface area, the use of microcarrier stirred-suspension culture, and easy scale-up [16]. Over the past few decades, a variety of commercial and experimental microcarriers have emerged and been used. For example, the Arifin group achieved a maximum Vero cell concentration of approximately 7.95 × 105 cells/mL with the use of Cytodex 1 [17]. Fernandes et al. reported the culture of mouse embryonic stem cells (ESCs) with either Cytodex 3 or CultiSpher-S microcarriers; after 8d, the maximal cell densities achieved were (1.9 ± 0.1) × 106 cells/mL and (2.6 ± 0.7) × 106 cells/mL, respectively, in serum-containing medium [18]. Cytodex 3, which is gelatin coated, is one of the most commonly used commercial microcarriers for the in vitro culture of hepatocytes and promotes their adhesion [19]. In addition, there have also been studies on the culture of hepatocytes on Cytopore 2 and CultiSpher-S [20, 21]. At present, there have been many studies on the large-scale culture of cells on microcarriers; however, previous hepatocyte microcarrier spinner cultures were mostly grown in serum-containing media, but there are many limitations to the clinical application of fetal bovine serum (FBS), and the culture of human hepatocytes, especially transdifferentiated cells, on microcarriers remains poorly reported [22].
In this study, to utilize microcarrier culture to achieve the high-density, large-scale expansion of preclinical transdifferentiated hiHeps in a self-developed, chemically defined SFM, we explored the types of microcarriers suitable for hiHeps and the basic culture conditions of microcarrier concentration, inoculation pattern, cell seeding density, and stirring speed in a spinner flask system. Furthermore, we investigated the effect of adding exogenous fibronectin (FN), an adhesion protein, on the attachment and proliferation of hiHeps on microcarriers and explored the signaling pathway involved. These studies provide a feasible methodology and design for hepatocyte serum-free microcarrier culture, which is important to expand a sufficient number of hiHeps for BAL treatment over a short time.
Materials and Methods
hiHeps Culture and Expansion in Static Conditions
hiHeps were obtained from the Shanghai Institutes for Biological Sciences of the Chinese Academy of Sciences. Upon thawing, cells were expanded in collagen coated plates, and passages 3–5 (P3–P5) of hiHeps were used for experiments. Adherent cells were cultured in a self-developed, chemically defined SFM, which consisted of DMEM/F12 (Gibco, USA) basal culture medium supplemented with 10 ng/mL TGF-α (PeproTech, USA), 40 ng/mL EGF (PeproTech), 10 μM DEX (Sigma-Aldrich, USA), 1.0 mg/L insulin (Sigma-Aldrich), 0.1 mg/L vitamin A acetate and 1.0 mg/L vitamin E acetate (Solarbio), which had previously been demonstrated to successfully support hiHep proliferation and function [15, 23]. Incubation at 37 °C in a humidified atmosphere contains 5% CO2.
hiHep Culture in a Stirred Spinner Flask
hiHep expansion was performed in 125-mL stirred spinner flask systems (Corning, NY, USA) with a magnetic, horizontal stirrer bar, and a vertical paddle (diam. D = 44 mm) was siliconized with silane. The working volume is 100 mL, and the spinner flasks were placed inside an incubator at 5% CO2 in air and 37 °C. Semi-continuous intermittent stirring for the first 8 h (25 min off, 5 min on, 30 rpm) was controlled by Cellroll Cellspin control unit (INTEGRA Biosciences AG, Switzerland). Three commercially available microcarriers: Cytodex 3 (GE Healthcare, Japan), Cytopore 2 (GE Healthcare, Japan), and CultiSpher-S (Sigma-Aldrich) were preconditioned in 50 mL SFM overnight before adding to the spinner flasks, and a 50% medium exchange was performed every 3 days in the first 6 days, then exchange every 2 days.
Attachment and Growth of hiHeps
To draw attachment and growth curves, 3 × 1 mL of evenly mixed microcarrier suspensions was collected and treated with a low-permeability crystal violet solution overnight. The cells were counted on a hemocytometer under an inverted microscope. The expansion and growth of hiHeps were calculated using the following equations:
where EFmax is the maximum increase in expansion, AEFmax is the actual maximum increase in expansion, ρ0 is the cell seeding density, and ρmax and ρ12h are the maximum cell density and the cell density at 12 h, respectively. μ is the specific growth rate (day−1), Cx(t) and Cx(0) are the cell numbers at the end and start of the exponential, respectively, and t is time (d).
Cell Viability
Cell viability was qualitative characterized with a live/dead assay. The collected cell-laden microcarriers were incubated with 5 μg/mL fluorogenic ester Calcein-AM (CAM) and 2 μg/mL propidium iodide (PI) (Sigma). Then, the microcarriers were viewed with an inverted fluorescence microscope (Nikon Eclipse Ti-S, Japan). Cell viability was quantitative characterized with a cell counting kit-8 (CCK-8, DOJINDO, Japan) according to the manufacturer’ s instructions.
Albumin and Urea Analyses
During cell culture, the culture supernatant was collected every 2 days. The urea concentration in the supernatant was measured using a BUN kit (Nanjing Jiancheng Bioengineering Institute, China), and albumin secreted by hiHeps was measured with a human albumin enzyme-linked immunosorbent assay (ELISA) kit (Bethyl, USA) according to the manufacturer’s instructions.
Measurement of LDH Release
The lactate dehydrogenase (LDH) levels in the supernatant were determined with a LDH assay kit-WST (Dojindo, Japan) according to the manufacturer’s instructions.
Selection of Stirring Speed Range
Stirring can improve the suspension of microcarriers, accelerate fluid mixing, and provide a uniform culture environment. However, excessive stirring may cause cell damage and poor cell growth.
Minimum Stirring Speed
The stirring speed at which the microcarriers usually leave the bottom of the reactor, with a residence time not more than 1–2 s, is called the minimum stirring speed, Njs [24]. The Zwietering formula is usually used to calculate Njs as follows [25]:
where S is a constant related to the type of stirring paddle, v is the kinetic viscosity, g is the acceleration due to gravity, p is the density, d is the diameter, and øs is the volume percentage of the microcarriers (Table 1).
Maximum Stirring Speed
During the suspension culture of microcarriers, cell damage mainly comes from turbulent eddies. The Kolmogorov turbulence theory is usually used to explain cell damage caused by a fluid [26]. The eddy size (η) and fluid shear stress (τs) were calculated as follows:
where ε is the energy dissipation rate, V is the culture volume, and Np is the power number (Table 2).
Detection of Glycogen and Lipids
Glycogen staining was performed with a Periodic Acid-Schiff kit (Sigma), and lipid staining was performed with an Oil Red O staining kit (Solarbio, G1262, China) according to the manufacturers’ instructions.
Uptake of Low-Density Lipoprotein (LDL)
hiHeps were incubated with 30 μg/mL Dil-ac-LDL (AngYuBio, China) for 5 h. Afterward, DAPI (1:100, Invitrogen, USA) was added to the cultures, which were incubated for 1 h. Images were taken with a fluorescence microscope.
Scanning Electron Microscopy (SEM)
The morphologies of cells on microcarriers were viewed by SEM (Hitachi S3400 N, Hitachi, Japan). Briefly, samples were fixed with 2.5% glutaraldehyde for 1 h and then dehydrated in a series of ethanol solutions (50%, 75%, 90%, 95%, and 100%). Afterward, the resultant samples were dried and sputter coated with gold, followed by SEM observation at a working voltage of 15 kV.
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
Total RNA was extracted from hiHeps with TRIzol reagent (Invitrogen, USA), and 1 mg of RNA was reverse transcribed with SuperScript III reverse transcriptase (Invitrogen) according to the manufacturer’s instructions. RT-PCR was performed as follows: 95 °C for 10 min and 40 cycles of PCR amplification. Each individual reaction was repeated three times, and the results were averaged. The primer sequences are shown in Table 3, in which GAPDH was used as a house keeping gene.
Western Blotting
hiHeps were lysed using RIPA buffer (Beyotime, China) containing 10 mM phenylmethylsulfonyl fluoride (Beyotime). Protein concentrations were determined using a BCA protein assay kit (Beyotime). Protein electrophoresis was performed with 10% SDS-PAGE gels, and proteins were transferred to PVDF membranes (Millipore) at 100 V for 100 min using a blotting apparatus. Membranes were blocked with 5% (w/v) nonfat dry milk in TBST buffer (20 mM Tris-HCl, pH 7.4, 150 mM NaCl, 0.05% Tween-20) for 30 min at room temperature and incubated with primary antibodies (Table 4) overnight at 4 °C, followed by the application of secondary antibodies conjugated with horseradish peroxidase (HRP) (1:10,000, Abcam) at room temperature for 1 h and visualization by enhanced chemiluminescence (Millipore). β-Actin served as the internal control, and the results of western blotting were analyzed by calculating the gray value.
Statistical Analysis
The results are shown as the mean ± standard deviation (SD), with at least three separate experiments for each issue addressed. Statistical significance was evaluated using one-way ANOVA and Student’s t test when data were determined to be normally distributed. A value of P < 0.05 indicated statistical significance.
Results
Effect of Microcarrier Type and Concentration on hiHep Attachment and Growth
In previous studies, hepatocytes were mainly cultured on three commercial microcarriers: Cytodex 3, Cytopore 2, and CultiSpher-S [19,20,21]. Therefore, we first determined which microcarrier is suitable for transdifferentiated hiHep culture and growth in SFM. The initial seeding regime (100 mL, 1 × 105 cells/mL, intermitted stirring) was identical for all three conditions. As determined by analysis of cell adherence, the attachment efficiencies of hiHeps on Cytodex 3, Cytopore 2, and CultiSpher-S at 12 h after inoculation were 72.81%, 35.00%, and 54.38%, respectively. These results indicated that hiHeps cultured on Cytodex 3 have better cell adhesion (Fig. 1a). All of the microcarriers were able to support hiHep expansion, and the maximal concentrations of cells cultured for 12 d with Cytodex 3, Cytopore 2, and CultiSpher-S were 12.10 × 105 cells/mL, 5.38 × 105 cells/mL, and 8.46 × 105 cells/mL, respectively. These concentrations corresponded to maximal increases in expansion (EFmax) values of 12.10, 5.38, and 8.46, respectively, indicating that hiHeps are cultured to higher cell concentrations on Cytodex 3 than on Cytopore 2 or CultiSpher-S (Fig. 1b, Table 5). The attachment efficiency of cells on Cytodex 3 can typically reach more than 80%, and the use of half-volume high-density inoculation is beneficial for increasing the cell adherence rate [27]. In addition, increasing the microcarrier concentration may increase the area of cell attachment and growth, which in turn increases the final cell concentration. Next, we explored the effect of inoculation method and Cytodex 3 microcarrier concentration on cell attachment and growth. When a half-volume high-density inoculation strategy was used within the first 8 h, the attachment efficiency of hiHeps cultured with different microcarrier concentrations for 12 h reached approximately 85% (Fig. 1c). However, the concentration of hiHeps decreased as the microcarrier concentration increased. (Fig. 1d, Table 6). Furthermore, hiHeps cultured on Cytodex 3 exhibited an epithelial morphology, and the cells were attached to each microcarrier for the first 2 days, but more empty spheres and unevenly attached cells gradually appeared beginning from the fourth day (Fig. 1e). It is possible that the hiHep adhesion ability on microcarriers in serum-free culture is weak and that hiHeps on microcarriers are susceptible to external factors and detachment, leading to death. Taken together, these results suggest that the Cytodex 3 microcarrier at a concentration of 3 mg/mL is more suitable for transdifferentiated hiHep attachment and growth than the other microcarriers tested or Cytodex 3 at different concentrations and that the half-volume high-density inoculation method is beneficial for improving the attachment efficiency of hiHeps.

Effect of microcarrier type and concentration on hiHeps attachment and growth. a, b The attachment efficiency (%) within 12 h and the concentration (105 cells/mL) within 14D of hiHeps on Cytodex 3, Cytopore 2, and CultiSpher-S microcarriers. c, d The attachment efficiency of hiHeps (%) and the concentration of hiHeps on different microcarrier concentrations. e Cell distribution observed with a microscopy (Nikon Eclipse Ti-S) under bright field (scale bars, 50 μm and 100 μm). All data are presented as mean ± SD; n = 3 samples per experimental group. *P < 0.05, between the different microcarrier type and different microcarrier concentration
Effect of Cell Seeding Density on hiHep Growth and Morphology
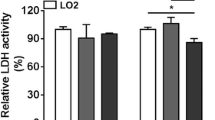
Because an increase in cell number per microcarrier may promote cell–cell interactions and increase the final hiHep concentration, we explored the effect of increasing cell seeding density on hiHep attachment and cell growth. The concentration of hiHeps at 12 d was highest when the cell seeding density was 2.0 × 105 cells/mL, and the maximal cell concentration was 17.81 × 105 cells/mL (Fig. 2a). The EFmax values at 12 d were 13.83, 9.79, 8.91, and 5.46 when cell seeding densities of 1.0 × 105 cells/mL, 1.5 × 105 cells/mL, 2.0 × 105 cells/mL, and 3.0 × 105 cells/mL, respectively, were used (Fig. 2b, Table 7). Similarly, the specific growth rate of hiHeps was significantly higher with a cell seeding density of 2.0 × 105 cells/mL than with the other cell seeding densities tested (Fig. 2c). Furthermore, the concentrations of LDH throughout the culture period were not significantly different when different cell seeding densities were used (Fig. 2d). SEM images still showed the presence of empty spheres, but hiHeps attached to the microcarriers exhibited a well-spread morphology with an elongated, polygonal shape (Fig. 2e). Together, these results suggest that the optimal cell seeding density in a 100-mL serum-free stirred spinner flask system is 2.0 × 105 cells/mL, which means it is best to inoculate each microcarrier with approximately 20 cells. In addition, increasing the cell seeding density did not eliminate the presence of empty spheres.

Effect of cell seeding density on hiHeps growth and morphology. a The concentration of hiHeps (105 cells/mL) on Cytodex 3. b The expansion fold at D12. c hiHeps growth kinetics. d LDH activity. e Cell morphology, scale bars are 200 μm, 100 μm, and 50 μm for SEM images, respectively. All data are presented as mean ± SD; n = 3 samples per experimental group. *P < 0.05, between the different cell seeding densities
Fibronectin Promotes hiHep Attachment and Proliferation on Microcarriers Through the Integrin-β1-Mediated FAK/ERK/CyclinD1 Pathway
A large number of glycoproteins have been found to mediate cell attachment, and fibronectin (FN) enhances hepatocyte spreading and attachment more than collagen does [28]. Therefore, we next explored the effect of FN addition on hiHep attachment and proliferation on microcarriers in SFM. Phase-contrast microscopy showed that the number of cells attached to microcarriers in SFM with a uniform distribution after 6 d was significantly higher with the addition of FN and that the epithelial morphology was not altered with the addition of FN (Fig. 3a). Then, the cell growth and viability of hiHeps in the presence of FN throughout the culture period were determined. FN significantly promoted hiHep growth and expansion in SFM (Fig. 3b, c), and similar results were observed for cell viability (Fig. 3d). Furthermore, to determine the signaling pathways by which FN promotes transdifferentiated hiHep attachment and proliferation, the effect of FN on the protein expression of integrin-β1, p-FAK, p-Akt, p-ERK, and CyclinD1 was detected by western blotting. All samples were collected on day 6 for protein expression. FN significantly increased the protein levels of integrin-β1, p-FAK, p-ERK, and CyclinD1, while the levels of p-Akt remained unchanged (Fig. 3e). Together, these results suggest that FN significantly promotes transdifferentiated hiHep attachment and proliferation and that this pro-proliferative effect is mediated by the integrin-β1/FAK/ERK/CyclinD1 pathway.

Fibronectin promotes hiHeps attachment and proliferation on microcarrier and this proliferation through the integrin-β1- mediated FAK/ERK/CyclinD1 signaling pathways. a Photomicrograph of hiHeps cultured in SFM without or with FN on Cytodex 3 at D6 (scale bars, 100 μm). b The concentration of hiHeps without or with FN within D14. c The expansion fold at D12. d Effect of FN on the vitality of hiHeps in SFM over 12 days. e The expression level of integrin-β1, p-FAK, p-Akt, p-ERK, and CyclinD1 was determined by western blotting at D6 (actin served as an internal control for protein loading). The bar graph shows the protein/β-actin ratio and is expressed as the fold change compared with the control, as evaluated by Image J. All data are presented as mean ± SD; n = 3 samples per experimental group. *P < 0.05, compared with control (SFM)
Effect of Stirring Speed on hiHep Growth, Viability and Function
In a previous experiment, we adopted a stirring speed of 60 rpm. Stirring can improve the suspension of microcarriers, accelerate fluid mixing, and provide a uniform culture environment; however, excessive stirring can cause cell damage and influence cell growth [29]. Studies have shown that the shearing stress generated by stirring can increase damage to cells under serum-free culture conditions, so the choice of stirring speed is crucial during microcarrier culture in SFM [30]. According to the Zwietering formula, the minimal stirring speed required for a suspension culture of 3 mg/mL Cytodex 3 in a 100-mL spinner system is approximately 30 rpm (Table 1) [24, 25]. During microcarrier suspension culture, the Kolmogorov turbulence theory is usually used to explain cell damage caused by fluid [26]; as the stirring speed increases, the fluid shear stress (τs) increases, but the eddy size (η) decreases (Table 2). The microcarrier particle size is approximately 175 µm, and cells are damaged when η is close to or smaller than the microcarrier particle size, so the maximum stirring speeding cannot exceed 75 rpm. According to the above theoretical calculation, the selected range of stirring speed was 30–60 rpm. The concentration of hiHeps in SFM at day 12 was highest when the stirring speed was 45 rpm (Fig. 4a). The EFmax values on day 12 were 8.63, 12.61, and 10.16 with stirring speeds of 30 rpm, 45 rpm, and 60 rpm, respectively (Fig. 4b). Phase-contrast microscopy showed that the microcarriers gradually gathered into larger aggregates, and more empty spheres appeared with a stirring speed of 30 rpm. The microcarriers were more dispersed with a stirring speed of 60 rpm, but an excessive stirring speed caused cells to detach from microcarriers and attach unevenly. With a stirring speed of 45 rpm, a small number of microcarriers aggregated, hiHeps attachment to microcarriers was relatively uniform, and there were few empty spheres (Fig. 4c). Furthermore, calcein-AM/PI fluorescent staining showed significantly larger areas of living cells (green) and smaller areas of dead cells (red) with a stirring speed of 45 rpm, suggesting that the cells had increased activity (Fig. 4d). The same result was found with the LDH release assay, and the LDH concentration in the culture supernatant was lowest with a stirring speed of 45 rpm for the first 10 days, but the concentration of LDH after this point was not significantly different with different stirring speeds (Fig. 4e). The urea synthesis function of hiHeps was highest with a stirring speed of 45 rpm throughout the culture period (Fig. 4f). These results indicated that hiHeps have increased growth and are more viable on Cytodex 3 in SFM with a stirring speed of 45 rpm than with other stirring speeds.

Effect of stirring speed on hiHeps growth, viability, and function. a The concentration of hiHeps (105 cells/mL) on Cytodex 3 at stirring speed of 30 rpm, 45 rpm, and 60 rpm. b The expansion fold at D12. c hiHeps distribution on Cytodex 3 observed with a microscopy under bright field at D3, D8 and D12 (scale bars, 100 μm). d hiHeps live-dead staining to characterize cell activity. e LDH activity. f Urea synthesis. All data are presented as mean ± SD; n = 3 samples per experimental group. *P < 0.05, between the different stirring speeds
Growth and Function of hiHeps on Microcarriers in SFM and HMM
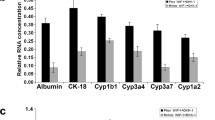
hiHeps cultured in hepatocyte maintenance medium (HMM) containing fetal bovine serum (FBS) maintain their growth and function for long periods of time in vitro, and hepatocyte microcarrier culture is mostly carried out in serum-containing medium. The above results revealed the optimal hiHep cultivation strategy in serum-free medium with microcarriers, which is beneficial for hiHep attachment and growth. Therefore, we next evaluated the growth and function of hiHeps on Cytodex 3 microcarriers in SFM and HMM. After 12 days of culture, the highest cell density in SFM was 25.22 × 105 cells/mL, and the maximum increase in expansion was 12.61-fold, while the highest cell density in HMM reached 28.18 × 105 cells/mL, and the maximum increase in expansion was 14.09-fold. When cultured in 2D serum-free culture conditions, the maximal cell concentration was achieved after 8 days culture, and the maximal expansion fold is 3.73, which much lower than 3D microcarrier culture, indicating the superiority of 3D culture in terms of the cell proliferation. (Fig. 5a,b). Then, calcein-AM/PI fluorescent staining showed that hiHeps in SFM and HMM both were well attached and exhibited a wide distribution of living cells (green), but there were fewer dead hiHeps (red) in SFM than in HMM (Fig. 5c). Similar results were observed for LDH secretion, and less LDH was released in SFM than in HMM, indicating a lower level of cell damage (Fig. 5d). Furthermore, analysis of liver-specific hiHep functions showed that as hiHep culture progressed, the levels of synthesized urea and secreted albumin were not significantly different from those at a later period of culture (Fig. 5e, f). In addition, the expression of hepatocyte function-associated genes was measured using fluorescence qRT-PCR, and cells cultured in 2D-SFM were used as controls. Compared with their expression in hiHeps cultured in 2D-SFM, expression of the ALB, TAT, TTR, HNF1A, and HNF4A genes was not significantly different, expression of the CK18, AAT, and CYP2A6 genes was increased, and expression of the TF gene was decreased in hiHeps cultured on microcarriers in 3D-SFM. Compared with those in hiHeps cultured in 3D-HMM, the gene expression levels of TAT and TTR were not significantly different, the gene expression levels of TF, CK18, AAT, HNF4A, and CYP2A6 were increased, and the gene expression levels of ALB and HNF1A were decreased in hiHeps cultured in 3D-SFM. Overall, 3D microcarrier culture increased the expression of only a small number of hepatocyte genes, and most hepatocyte genes showed no significant change in expression between 3D and 2D culture (Fig. 5g). Overall, the growth and function of hiHeps grown on microcarriers in SFM is close to those of hiHeps grown on microcarriers in HMM with the optimal cultivation strategy.

The growth and function of hiHeps on microcarrier in SFM and HMM. a The concentration of hiHeps (105 cells/mL) cultured on Cytodex 3 in 2D-SFM, 3D-SFM, and 3D-HMM. b The expansion fold at D12. c Cells live-dead staining. d LDH activity. e Urea synthesis. f Albumin secretion. g qRT-PCR was performed to quantitatively detect the relative hepatocyte gene expression in 2D-SFM, 3D-SFM, and 3D-HMM. All data are presented as mean ± SD; n = 3 samples per experimental group. *P < 0.05, ns is no significant difference, compared with control (2D-SFM, 3D-HMM)
Effects of Microcarrier Culture on hiHep Morphology and Function after Harvest in SFM
hiHeps cultured on Cytodex 3 were seeded in 24-well plates after 3 days, and cell morphology and glycogen and lipid staining were observed and photographed with an inverted microscope. In terms of cell morphology, the cells harvested on microcarriers exhibited an epithelial morphology similar to that of cells cultured directly in plates (Fig. 6a). Glycogen staining showed that hiHeps cultured on Cytodex 3 had the ability to synthesize glycogen (Fig. 6b). Similar results were observed by lipid staining, and the staining and quantitative analysis of lipids showed that hiHeps also had the ability to synthesize lipids to the same extent as those cultured in 2D (Fig. 6c). Furthermore, the ability of hiHeps to transport Dil-labeled ac-LDL was observed by fluorescence microscopy, and harvested hiHeps were able to transport LDL (red) (Fig. 6d). These results suggest that hiHeps maintain their morphology and function after expansion on microcarriers in an SFM spinner flask system.

Effects of microcarrier culture on hiHeps morphology and function after harvest in SFM. a Photomicrograph of hiHeps after harvest on Cytodex 3 in SFM, compared with cultured in 2D SFM. b Glycogen staining. c Lipid staining and quantification. d Dil-ac-LDL transport, LDL intake as indicated by red fluorescence, and the blue staining is DAPI. Scale bars: (a–d) 100 μm. All data are presented as mean ± SD; n = 3 samples per experimental group. ns is no significant difference, compared with control (2D-SFM)
Discussion
Achieving the rapid and large-scale preclinical expansion of hepatocytes in serum-free conditions for bioartificial livers is essential for the treatment of liver failure, and the microcarrier culture system is currently an important and effective approach. Previously, we demonstrated that transdifferentiated hiHep proliferation and function can be successfully supported during long-term passage in our self-developed, chemically defined SFM in 2D Petri dishes [15]. In this study, we found that the Cytodex 3 microcarrier is more suitable than the other microcarriers tested for hiHep culture and that the optimal culture conditions are the use of half-volume high-density inoculation during the adherence period, a Cytodex 3 concentration of 3 mg/mL, a cell seeding density of 2.0 × 105 cells/mL, and a stirring speed of 45 rpm; these conditions are beneficial to hiHep attachment and growth. In addition, the addition of fibronectin (FN) can promote hiHep attachment and proliferation on Cytodex 3 in SFM through the integrin-β1/FAK/ERK/CyclinD1 pathway. Finally, the cells harvested on microcarriers have a normal hepatocyte morphology and exhibit normal hepatocyte synthetic and uptake functions.
Due to the large specific surface area and the advantages of suspension culture over adherent culture, microcarrier technology for the large-scale culture of cells is currently recognized as a promising means of cell expansion [16]. Previous studies have shown that transdifferentiated hiHeps represent an important in vitro cell source for BAL treatment, but it is difficult to achieve a high cell density and large-scale cell expansion in self-developed SFM over a short time period [13, 15]. Therefore, we studied the culture of hiHeps on microcarriers under serum-free conditions. Over the past few decades, a variety of commercial microcarriers have been used to expand hepatocytes; these commercial microcarriers include Cytodex 3, Cytopore 2, and CultiSpher, among which Cytodex 3 is the most common microcarrier for hepatocyte culture [19,20,21]. Three kinds of microcarriers provide different surface area; Cytodex 3 is a solid microcarrier coated with a layer of denatured collagen, which is beneficial to the adhesion of epithelial cells and increases the success of hepatocyte culture [31]. However, Cytopore 2 and CultiSpher are macroporous microcarriers with a larger surface area. Consistent with most reports, our experimental results showed that Cytodex 3 is more suitable than the other microcarriers examined for hiHep culture. Furthermore, Cytodex 3 is the only microcarrier that has been used in experimental animals and clinical research on BAL, which forms a solid basis for the clinical application of hiHep-BALs.
Previous research on cell attachment in microcarrier suspension cultures showed that adopting intermittent stirring and inoculation in 50% of the final culture volume led to a notable increase in the attachment efficiency of rabbit mesenchymal stem cells in a Cytodex 3 microcarrier culture system [27]. This finding is consistent with our experimental results, which showed that the half-volume high-density inoculation method significantly increased the attachment efficiency of hiHeps on microcarriers by up to approximately 85% with different microcarrier concentrations. The maximum density of hepatocytes cultured using microcarriers is limited by the microcarrier concentration, which can affect the growth of hepatocytes and even cause cell death [32]. Therefore, it is important to choose the appropriate microcarrier concentration, and the concentration of Cytodex 3 microcarriers used is generally 0.5–5 mg/mL [33]. Our results showed that the maximum number of hiHeps can be harvested with a Cytodex 3 concentration of 3 mg/mL in a 100-mL spinner system. In addition, one of the key parameters affecting inoculation is the cell seeding density, that is, the number of hepatocytes inoculated per microcarrier, which directly affected the growth of hepatocytes on microcarriers and the final concentration of hepatocytes [31, 34]. A previous report showed that inoculation of 26.7 hepatocytes per microcarrier showed the highest viability [35, 36]. Our study revealed that the optimal number of hepatocytes per microcarrier is approximately 20 cells, which is beneficial to hiHep growth and increases the final hiHep concentration.
During microcarrier-based stirred hiHep culture in SFM, empty spheres gradually appeared beginning on the fourth day. This finding may be due to weaker hiHep adhesion on Cytodex 3 microcarriers under serum-free culture conditions compared to serum-containing conditions as the culture progresses, with cells more easily detached from the microcarriers, leading to empty spheres. The adhesion of cells to the culture surface is usually increased by the addition of extracellular matrix to the medium. One study showed that it is necessary to add FN or FBS, which can promote cell adhesion to the culture surface, to medium [37, 38]. The medium supplemented with 10% FBS contained nearly 2–3 μg FN/mL, and a large amount of FN adsorbed onto the microcarrier surface within a few minutes [39]. FN is usually added to SFM to allow most cells to adhere to the culture surface, which is consistent with our results showing a significant decrease in empty spheres and increase in the final hiHep concentration in SFM after the addition of 10 μg/mL FN. FN can affect cell attachment and proliferation through a variety of signaling pathways. As an example, Matsuo M et al. showed that FN induces the activation of extracellular signal-regulated kinase (ERK), p38 and Akt as well as cell proliferation and at least in part via integrins [40]. IIIario M et al. demonstrated that integrin stimulated by FN activates two signaling pathways: the Ras/Raf/ERK and calcium Ca2+/calcium calmodulin-dependent kinase II (CaMKII) pathways, both of which are necessary to stimulate cell proliferation [41]. In this study, we showed that FN does not affect the phosphorylation of Akt and that the proliferation of transdifferentiated hiHeps is mediated by the integrin-β1/FAK/ERK/CyclinD1 pathway.
Because hepatocytes have no protective cell walls and are particularly sensitive to shear forces, studies have found that the shear force generated by agitation increases cell damage under serum-free culture conditions [30]. Therefore, stirring speed is an important factor in the serum-free microcarrier culture of hepatocytes [42]. The appropriate agitation speed is usually 50–70 rpm when using a conventional magnetic stirring vessel, but we used a range of stirring speeds of 30–60 rpm according a relative theoretical calculation and determined the optimal stirring speed to be 45 rpm [24,25,26]. Finally, the growth and function of hiHeps growth on microcarriers in SFM are close to those of hiHeps grown on microcarriers in HMM under the optimal cultivation conditions, but the expression of most hepatocyte-related genes was not significantly different between 2D culture and microcarrier culture. In fact, microcarrier culture was essentially similar to monolayer cell culture in plates and merely enables high-density cell culture and amplification [43]. However, the obtained cells must also have normal liver function. hiHeps are a transdifferentiated cell with weak liver function compared to that of primary hepatocytes, so it is important to improve the function of hiHeps grown in serum-free culture in the future, including their coculture with other cells or the use of three-dimensional (3D) perfusion culture, which can through mimic the in vivo microenvironment to promote the function of hiHeps [44, 45]. In summary, we have developed a chemically defined, serum-free microcarrier culture strategy for culturing hiHeps in SFM that achieved high-density hiHep expansion, which provides a solid basis for the clinical application of hiHep-BALs.
References
Martin P, Friedman LS. Assessment of liver function and diagnostic studies. In: Handbook of Liver Disease. 2018:1–17.
Hernaez R, Solà E, Moreau R, et al. Acute-on-chronic liver failure: an update. Gut. 2017;66:541–553.
Habib S, Shaikh OS. Drug-induced acute liver failure. Clin Liver Dis. 2017;21(1):151–162.
Bernal W, Wendon J. Acute liver failure. N Engl J Med. 2013;369:2525–2534.
Struecker B, Raschzok N, Sauer IM. Liver support strategies: cutting-edge technologies. Nat Rev Gastroenterol Hepatol. 2013;11:166–176.
Sussman NL, Kelly JH. Artificial liver. Clin Gastroenterol Hepatol. 2014;12:1439–1442.
Lee KCL, Stadlbauer V, Jalan R. Extracorporeal liver support devices for listed patients. Liver Transpl. 2016;22(6):839–48.
Yu CB, Pan XP, Li LJ. Progress in bioreactors of bioartificial livers. Hepatobiliary Pancreat Dis Int. 2009;8:134–140.
Gu J, Shi X, Ren H, et al. Systematic review: extracorporeal bio-artificial liver-support system for liver failure. Hep Intl. 2012;6:670–683.
Pan XP, Li LJ. Advances in cell sources of hepatocytes for bioartificial live. Hepatobiliary Pancreat Dis Int. 2012;11:594–605.
Huang P, He Z, Ji S, et al. Induction of functional hepatocyte-like cells from mouse fibroblasts by defined factors. Nature. 2011;475:386–389.
Huang P, Zhang L, Gao Y, et al. Direct reprogramming of human fibroblasts to functional and expandable hepatocytes. Cell Stem Cell. 2014;14:370–384.
Shi XL, Gao Y, Yan Y, et al. Improved survival of porcine acute liver failure by a bioartificial liver device implanted with induced human functional hepatocytes. Cell Res. 2016;26:206.
Valk JVD, Brunner D, Smet KD, et al. Optimization of chemically defined cell culture media—replacing fetal bovine serum in mammalian in vitro methods. Toxicol In Vitro. 2010;24:1053–1063.
Gu C, Li PP, Liu W, et al. The role of insulin in transdifferentiated hepatocyte proliferation and function in serum-free medium. J Cell Mol Med. 2019;23:4165–4178.
Blüml Gerald. Microcarrier cell culture technology. Methods Biotechnol. 2007;24:149–178.
by Vero Cells Grown on Cytodex 1 Microcarriers in a 2-Litre Stirred Tank Bioreactor. J Biomed Biotechnol. 2015;2010:586363.
Fernandes AM, Fernandes TG, Diogo MM, et al. Mouse embryonic stem cell expansion in a microcarrier-based stirred culture system. J Biotechnol. 2007;132:227–236.
Tao X, Shaolin L, Yaoting Y. Preparation and culture of hepatocyte on gelatin microcarriers. J Biomed Mater Res, Part A. 2010;65:306–310.
Schulz CM, Ruzicka J. Real-time determination of glucose consumption by live cells using a lab-on-valve system with an integrated microbioreactor. The Analyst. 2002;127:1293–1298.
Werner A, Duvar S, Müthing Johannes, et al. Cultivation of immortalized human hepatocytes HepZ on macroporous CultiSpher G microcarriers. Biotechnol Bioeng. 2000;68:59–70.
Gstraunthaler G, Lindl T, et al. A plea to reduce or replace fetal bovine serum in cell culture media. Cytotechnology. 2013;65:791–793.
Li PP, Gu C, Liang BY, et al. A serum-free medium suitable for maintaining cell morphology and liver-specific function in induced human hepatocytes. Cytotechnology. 2019;71:329–344.
Dohi N, Takahashi T, Minekawa K, et al. Power consumption and solid suspension performance of large-scale impellers in gas–liquid–solid three-phase stirred tank reactors. Chem Eng J. 2004;97:103–114.
Frijlink JJ, Bakker A, Smith JM. Suspension of solid particles with gassed impellers. Chem Eng Sci. 1990;45:1703–1718.
Croughan MS, Hamel JF, Wang DIC. Hydrodynamic effects on animal cells grown in microcarrier cultures. Biotechnol Bioeng. 2000;67:841–852.
Jiang D, Hu J, Zhou Y, et al. Optimization of attachment conditions for rabbit mesenchymal stem cells in cytodex 3 microcarrier culture systems. Shengwu yixue gongchengxue zazhi. 2007;24:884.
Shiojiri N, Sugiyama Y. Immunolocalization of extracellular matrix components and integrins during mouse liver development. Hepatology. 2004;40:346–355.
Nienow AW, Hewitt CJ, Heathman TRJ, et al. Agitation conditions for the culture and detachment of hMSCs from microcarriers in multiple bioreactor platforms. Biochem Eng J. 2016;108:24–29.
Burnouf T, Griffiths E, Padilla A, et al. Assessment of the viral safety of antivenoms fractionated from equine plasma. Biologicals. 2004;32:115–128.
Yi G, Huanzhang H, Ke C, et al. Primary porcine hepatocytes with portal vein serum cultured on microcarriers or in spheroidal aggregates. World J Gastroenterol. 2000;6:365–370.
Yiheng C, Shuyu T, Xuping L, et al. The effects of microcarrier concentration and cell density on the growth of swine testicle cells. Biotechnol Bull. 2016;32:242–250.
Demetriou AA, Reisner A, Sanchez J, et al. Transplantation of microcarrier-attached hepatocytes into 90% partially hepatectomized rats. Hepatology (Baltimore, Md.),. 1988;8:1006–1009.
Hewitt CJ, Lee K, Nienow AW, et al. Expansion of human mesenchymal stem cells on microcarriers[J]. Biotech Lett. 2011;33:2325–2335.
Shin WY, Lee KU, Lee HW, et al. Optimal number of hepatocytes per microcarrier in spheroid culture using cytodex 3 microcarrier. J Korean Surg Soc. 2007;73:235–241.
Liu ML, Mars WM, Zarnegar R, et al. Collagenase pretreatment and the mitogenic effects of hepatocyte growth factor and transforming growth factor-alpha in adult rat liver. Hepatology. 2010;19:1521–1527.
Grinnell F, Hays DG, Minter D. Cell adhesion and spreading factor: partial purification and properties. Exp Cell Res. 1977;110:175–190.
Hughes RC, Pena SDJ, Clark J, et al. Molecular requirements for the adhesion and spreading of hamster fibroblasts. Exp Cell Res. 1979;121:307–314.
Feinberg AW, Schumacher JF, Brennan AB. Engineering high-density endothelial cell monolayers on soft substrates. Acta Biomater. 2009;5:2013–2024.
Matsuo M, Sakurai H, Ueno Y, et al. Activation of MEK/ERK and PI3K/Akt pathways by fibronectin requires integrin αv-mediated ADAM activity in hepatocellular carcinoma: a novel functional target for gefitinib. Cancer Sci. 2006;97:155–162.
Illario M, Cavallo AL, Monaco S, et al. Fibronectin-induced proliferation in thyroid cells is mediated by αvβ3 integrin through Ras/Raf-1/MEK/ERK and calcium/CaMKII signals. J Clin Endocrinol Metab. 2005;90:2865–2873.
Gigout A, Buschmann MD, Jolicoeur M. Chondrocytes cultured in stirred suspension with serum-free medium containing pluronic-68 aggregate and proliferate while maintaining their differentiated phenotype. Tissue Eng Part A. 2009;15:2237–2248.
Huang L, Xiao L, Jung Poudel A, et al. Porous chitosan microspheres as microcarriers for 3D cell culture. Carbohyd Polym. 2018;202:611–620.
Rebelo SP, Costa R, Silva MM, et al. Three-dimensional co-culture of human hepatocytes and mesenchymal stem cells: improved functionality in long-term bioreactor cultures. J Tissue Eng Regen Med. 2017;11:2034–2045.
Wei G, Wang J, Lv Q, et al. Three-dimensional coculture of primary hepatocytes and stellate cells in silk scaffold improves hepatic morphology and functionality in vitro. J Biomed Mater Res, Part A. 2018;106:2171.
Acknowledgments
We thank the Shanghai Institutes for Biological Sciences of the Chinese Academy of Sciences for providing hiHeps and the cell culturing protocol. Ce Gu performed all the experiments and wrote the manuscript. Miaomiao Chai, Jiaxing Liu, Hui Wang, and Wenjing Du were involved in useful discussions during the development of this study. Yan Zhou and Wen-Song Tan contributed to the conception, design of the work or of parts of it, and its interpretation.
Funding
This research was supported by the Basic Research Project of Shanghai Science and Technology Commission (Grant No. 16JC1400203), the National Key Research and Development Program of China, 2018YFC1105801, and the National Natural Science Foundation of China (Grant No. 81671841).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have declared that no conflict of interest exists.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Gu, C., Chai, M., Liu, J. et al. Expansion of Transdifferentiated Human Hepatocytes in a Serum-Free Microcarrier Culture System. Dig Dis Sci 65, 2009–2023 (2020). https://doi.org/10.1007/s10620-019-05925-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10620-019-05925-8