
Abstract
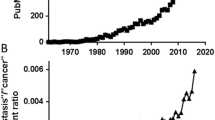
Brain metastasis, characterized by poor clinical outcomes, is a devastating disease. Despite significant mechanistic and therapeutic advances in recent years, pivotal improvements in clinical interventions have remained elusive. The heterogeneous nature of the primary tumor of origin, complications in drug delivery across the blood–brain barrier, and the distinct microenvironment collectively pose formidable clinical challenges in developing new treatments for patients with brain metastasis. Although current preclinical models have deepened our basic understanding of the disease, much of the existing research on brain metastasis has employed a reductionist approach. This approach, which often relies on either in vitro systems or in vivo injection models in young and treatment-naive mouse models, does not give sufficient consideration to the clinical context. Given the translational importance of brain metastasis research, we advocate for the design of preclinical experimental models that take into account these unique clinical challenges and align more closely with current clinical practices. We anticipate that aligning and simulating real-world patient conditions will facilitate the development of more translatable treatment regimens. This brief review outlines the most pressing clinical challenges, the current state of research in addressing them, and offers perspectives on innovative metastasis models and tools aimed at identifying novel strategies for more effective management of clinical brain metastasis.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Brain metastasis: a clinical challenge
Introduction
Brain metastases (BrMs) are generally considered secondary metastatic tumors, derived from the dissemination of a primary tumor. Brain metastatic relapse occurs in 20–30% of patients with advanced cancer. Most BrMs arise from lung (40–50%), breast (30%), melanoma (20%), colorectal cancer (CRC, 1–2%), and renal cell carcinoma (RCC, 7–10%) [1,2,3]. Lung cancer BrM is the most common primary tumor of origin observed in male patients diagnosed with BrM, while breast cancer BrM accounts for the majority of cases in female patients [1]. BrMs, although derived from single primary tumors of origin, may still exhibit different molecular subtypes. Even in a single patient harboring one primary tumor, numerous distinct, clonally derived brain lesions may be present, each potentially differing from the others. All these factors together contribute to BrM heterogeneity. Furthermore, systemic and lifestyle factors, such as age and microbiome status, might also influence the disease trajectory [4, 5]. Despite poor clinical outcomes and patient survival, progress in advancing treatment options for BrM patients has been marginal. This is partly attributed to the low inclusion of BrM patients in clinical trials and limited funding for BrM research, compared to other CNS cancers [6].
In current clinical practice, surgery and radiotherapy, such as stereotaxic radiosurgery (SRS) and whole brain radiotherapy (WBRT), are primary treatment regimens for brain metastases (BrMs) (Table 1). WBRT is used in a subset of patients, despite its significant side effects, including impaired cognitive function, which can be slightly alleviated through hippocampal avoidance [7]. Systemic treatment regimens tailored to the specific molecular subtype of the primary tumor of origin are also used for treating/preventing brain metastasis. For example, HER2 + breast cancer BrM patients receive anti-HER2 targeted therapy such as trastuzumab, while more general systemic chemotherapies are indicated in triple negative breast cancer (TNBC) BrMs [8, 9]. A single-arm phase III clinical trial was concluded in 2020 for trastuzumab emtansine (T-DM1) for patents with HER2 + metastatic breast cancer (KAMILLA, NCT01702571). T-DM1 exhibited a low toxicity profile and showed reduction of BrM lesion size in 42.9% of patients [10]. Another commonly utilized molecular therapy involves treating asymptomatic BrM in lung cancer patients with anaplastic lymphoma kinase (ALK) rearrangement, using receptor tyrosine inhibitors (TKIs) such as the third generation ALK/ROS1 inhibitor lorlatinib. In one single-arm trial, lorlatinib treatment led to a measurable intracranial response in 64% of patients which was sustained for 12.4 months (median) [11].
Immunotherapy, particularly immune checkpoint therapy (ICT) targeting the PD-1/PD-L1 and CTLA-4 axes, shows promise as a cancer treatment for certain patient subsets with metastatic melanoma, non-small cell lung cancer, RCC, and head and neck cancers. ICT modalities generally have better side effect profiles compared to traditional chemotherapies [12]. Thus far, immunotherapies have only been used in a subset of BrM patients such as a recent phase II study utilizing combinatorial ICT in asymptomatic BrMs derived from melanoma. Although the treatment regimen achieved efficacy in 57% of melanoma patients in some studies, ranging from stable disease for 6 months to complete response, such targeted approaches have been ineffective in treating other BrMs [13,14,15,16,17].
Aside from direct actions on the tumor cells, radiotherapy is known to induce immunogenic cell death (ICD) and abscopal effects, a phenomenon whereby radiotherapy treatment also affects lesions outside the radiation field [18, 19]. Consequently, recent efforts have been directed towards exploring combinatorial radioimmunotherapy in clinical settings. Melanoma patients who were treated with immune checkpoint therapy (ICT) ipilimumab in combination with WBRT achieved longer median survival compared to standard of care WBRT (21.3 versus 4.9 months) and an increased 2-year OS (47.2% versus 19.7%). In addition, SRS plus ipilimumab has also been shown to yield an increased OS (18.3 versus 5.3 months) in favor of the ICT + WBRT combination [20]. While promising, the current clinical practice of combining ICT with radiotherapy requires more informative guidance regarding treatment timing and dosing schedules. As the ideal schedule of each treatment modality is likely to vary between different BrM subtypes, there is a critical need for preclinical models to identify the optimal regimen with respect to immunotherapy and primary tumor/subtype of origin.
Brain metastases of unknown origin
Although BrMs are usually identified in patients later during the clinical treatment course of the diagnosed primary tumor, a subset of patients develop BrMs as the first symptom of previously undiagnosed cancer [1, 21]. A recent estimate is that up to 15% of all BrM patients present as BrMs with cancer of unknown primary (BM-CUP) and these patients are often treated with SRS. He diagnosis procedure for BM-CUP includes a physical examination, blood and biochemical tests, immunohistochemistry (IHC), mammography (for breast cancer), whole body computed tomography (CT), and/or magnetic resonance imaging (MRI). If a primary tumor is still not identified through these efforts, immunohistochemistry (IHC) on BrM biopsies using common primary tumor markers could provide clues on the BrM tissue of origin [22]. Despite these efforts, the primary tumor of origin can often remain elusive even after autopsy due to the lack of specific prognostic or predictive tumor markers and complete lack of molecular mechanistic insight into the disease [22]. Next generation sequencing (NGS) is the most valuable and feasible tool to guide identification of molecular targets in BM-CUP, in a similar fashion to primary tumors. A 2018 NGS study of formalin-fixed paraffin embedded (FFPE) tissue sections of 21 BM-CUP patients noted 76 gene variants, some of which are targetable. The commonly detected genomic variants in BM-CUP were TP53, KRAS, MET, and MYC [23]. Beyond correlative genetic studies, the enigmatic manifestation of BM-CUP, characterized by early metastatic spread to the brain and simultaneous regression of the primary tumor site, is difficult to model pre-clinically. Recently, a study proposed “agnospheres”, stem-like cells originating from human CUPs, as a novel disease model that recapitulates both early cell dissemination and multi-organ metastasis [24]. This study showed that agnosphere properties, namely proliferation and self-renewal while maintaining stemness, rely on the constitutive activation of the MAPK pathway and the MYC oncogene. Targeted inhibition of the MAPK pathway component MEK by trametinib caused cancer cell death and necrosis. The gene expression signature responsive to trametinib is a feature of most patient CUPs, suggesting that MAPK amplification is a common mechanism, potentially contributing to BM-CUPs [24].
Unique challenges in drug delivery to brain metastases
Separate from the features of originating primary tumor stroma, the brain environment poses unique obstacles for the delivery of treatments. The central nervous system (CNS) is protected by two main functional neurovascular barriers: (1) the blood–brain barrier (BBB), and (2) the blood–cerebrospinal fluid barrier [1]. The BBB consists of endothelial cells with high rates of efflux that are connected by tight junctions as well as pericytes and astrocytic end-feet (basement membrane). When intact, the BBB is mostly permeable to only small uncharged compounds that can diffuse passively. Although the mechanisms are not fully understood, egress of cells such as macrophages and disseminated tumor cells into the CNS occurs during BrM, while most chemotherapy agents and targeted- or immuno-therapies, such as ICT cross the BBB poorly with reduced therapeutic effects [1, 25]. The blood-cerebrospinal fluid barrier on the other hand is formed by choroid plexus epithelial cells. The breaching of this barrier is thought to be permissive for leptomeningeal metastasis [1, 26].
There is now both pre-clinical and clinical evidence that the BBB becomes more permeable in the later outgrowth stages of BrM, owning to the formation of the blood tumor barrier (BTB) [1, 27]. The BTB however is only partially permeable, and the degree of permeability can vary from lesion to lesion within the same patient [1, 25, 27, 28]. There are currently many clinical trials underway investigating various (molecular, biological, and physical) approaches to increase BBB permeability for brain cancers, as well as recent pre-clinical advances in non-invasive approaches [29]. Recent studies in mice and rat models have shown that a combination of low-intensity focused ultrasound (FUS) pulses and circulating microbubbles provide a physical/mechanical approach to transiently disrupt and permeabilize the BBB [30] and to deliver doxorubicin for the treatment of glioblastoma [31]. A more recent 2023 preclinical study aimed specifically at BrM examined FUS combined with paclitaxel and Doxil in metastatic TNBC. They showed that the FUS + chemo combinatorial regimen increased median survival and led to the slowest brain tumor burden progression compared to other groups [32]. More pre-clinical and translational studies are needed to assess the efficacy of such approaches in treating BrM.
Mechanisms of brain metastasis: an update
Our current understanding of key brain metastasis drivers from primary tumors of various origins has been extensively reviewed by Campbell et al. [33] and Achrol et al. [1]. Here, we only highlight some of the most recent advances in our understanding of BrM.
Novel insights gained from pre-clinical models
Cancer cell dormancy, therapy resistance, and metabolic alterations
Most disseminated tumor cells (DTCs) do not survive the process of extravasation. Among the surviving DTC seeds, some may enter an extended period of dormancy, characterized by a slow or non-cell-cycling state and activation of quiescence effectors downstream of p38 [34]. These cells can persist for long periods of time and later reactivate the cell cycle to induce overt metastases [35]. Moreover, due to their slow cycling state and high stemness/phenotypic plasticity, dormant cells that later lead to disease relapse are often therapy resistant [36], highlighting the importance of successful identification and eradication of these rare cells. General molecular drivers of dormancy along with current strategies for treating latent disease have been reviewed in detail [34] and several dormancy markers have been identified specifically in glioblastoma: H2BK, IFGBP5, and EPHA5. Despite clinical evidence that these markers are present in certain BrM patient populations [35], detailed mechanistic studies in pre-clinical models have not been performed to assess their functional relevance and equivalence in BrM. Recent work in a pre-clinical mouse model of TNBC BrM, however, demonstrated that astrocyte-deposited lamanin-211 drives DTC quiescence within the perivascular niche. Stripping of astrocyte end feet is required for activation of nuclear YAP-mediated signaling and BrM outgrowth [37]. This study not only identified a brain-specific mechanism of DTC dormancy, the rate limiting step of BrM outgrowth, but also highlighted the importance of conducting more studies exploring both the cancer cell intrinsic and extrinsic mechanisms governing entry and exit out of quiescence.
Therapy resistance in the context of brain metastasis is closely linked to resistance to radiotherapy and targeted therapy. Although radiotherapy is the primary treatment for most patients, the molecular mechanisms underlying resistance acquisition have been insufficiently studied. Melanoma and renal cell carcinoma (RCC) BrMs are largely resistant to radiotherapy [38]. Thus, there is critical need to better understand underlying drivers of resistance. A recent study employed radiotherapy (WBRT)-resistant experimental models of lung adenocarcinoma and breast cancer BrMs to delineate the molecular characteristics of resistance. They treated oncospheres derived from lung and breast cancer cell lines with radiation and conducted RNA sequencing to identify transcriptional differences between radioresistant and radiosensitive oncospheres. This study revealed that the S100A9-RAGE-NF-kB-JunB pathway mediates radioresistance in these models and established a correlation between S100A9 expression levels in BrM patient datasets and clinical responses to WBRT. These findings not only elucidated a mechanism of resistance acquisition but also point to a potential biomarker for assessing WBRT efficacy in clinical settings [39]. Elucidating the molecular mechanisms of resistance to targeted therapies presents a separate challenge, as these therapies are specific to different primary tumor subtypes. Recent studies in lung cancer mouse models have investigated brain lesions resistant to the EGFR inhibitor osimertinib. One such study linked S100A9 overexpression, via a distinct pathway involving retinoic acid signaling, to BrM relapse following osimertinib treatment [40]. Another study identified a RhoA/SRF signaling axis that activates gene programs associated with drug resistance. Importantly, combining RhoA inhibition with osimertinib resulted in a substantial reduction of BrM burden in this preclinical model [41].
The role of altered cellular metabolism in cancer in a wider context is well established, yet metabolic reprogramming in BrM has been less extensively examined and is an emerging area of exploration [42, 43]. Studies by Parida et al. demonstrated that metabolic diversity and flexibility within BrM cells is critical for successful BrM outgrowth in a HER2 + breast cancer model. Moreover, latent BrM cells utilize dynamin-related protein 1 (DRP1) to sustain fatty acid oxidation and survival, thus pharmacological inhibition, or genetic depletion of DRP1 reduces BrM burden [44, 45].
The CNS cellular microenvironment
Aside from the BBB, resident brain cells engage in dynamic crosstalk and together shape normal CNS function at steady state. Neurons, oligodendrocytes, astrocytes, microglia, and pericytes comprise the CNS-native cellular compartment which can additionally interact with peripheral immune cells [46].
CNS-Native cells
During the evolution of BrM, the brain environment undergoes dramatic remodeling [46]. It has been clinically observed that activated microglia and astrocytes encircle BrM lesions in a process called gliosis [47, 48]. There have been numerous pre-clinical studies in mouse models investigating the roles of microglia and astrocytes in BrM. It is now well established that astrocytes play pro- and anti-metastatic roles depending on the stage and specific astrocyte subtype involved. Astrocytes can directly induce apoptotic cell death in cancer cells via plasminogen secretion, while a subtype of reactive astrocytes marked with pSTAT3 can deliver cGAS and miRNAs to promote BrM progression [49, 50]. Moreover, astrocytes have been shown to activate proliferator-activated receptor γ (PPARγ) in cancer cells, inducing their proliferation and outgrowth [51]. Microglia, the main CNS-native immune macrophage, have similarly dichotomous roles in BrM. Recent work has highlighted the tumor-promoting roles of this cell population. In our recent study, Guldner et al. utilized scRNA-seq in a BrM mouse model and showed that microglia downregulate homeostatic genes such as Cx3cr1 in favor of a pro-inflammatory, interferon driven gene program. Excessive Cxcl10 release cultivated the microglia-dependent immunosuppressive niche. Moreover, microglial expressed a high level of Vista, a novel checkpoint molecule that can be targeted in combination with ICT for BrM treatment [52].
Peripheral immune cell infiltration
Immune cells, such as cytotoxic CD8 + T lymphocytes, are present in brain metastases (BrM), and immune surveillance occurs via the meningeal lymphatic system. However, the tumor-induced reprogramming of the microenvironment favors the accumulation of anti-inflammatory regulatory T cells (Tregs) [20, 46]. Currently, therapies such as anti-PD-1/PD-L1 and anti-CTLA-4, including nivolumab and ipilimumab, are clinically administered to treat melanoma and non-small cell lung cancer (NSCLC) derived BrMs but are not standard for BrMs from other primary tumors [20].
In our recent work, we detailed the role of the enhancer of zeste homolog 2 (EZH2), a well-documented histone methyltransferase, in the advancement of BrM. We discovered an upregulation of EZH2 in human BrM specimens which facilitates the recruitment of immunosuppressive neutrophils, characterized by high arginase1 (Arg1) and PD-L1 expression, into the brain metastatic niche, thereby aiding BrM proliferation. Pharmacological interventions that impede neutrophil recruitment, ICT, and the inhibition of EZH2 phosphorylation through Src inhibitors have shown significant reductions in BrM across various mouse models [53]. More recently, research has indicated that persistent type I interferon (IFN) signaling in astrocytes and cancer cells instigates the secretion of C–C motif chemokine receptor 2 (CCR2) by astrocytes, which in turn attracts pro-tumor monocytes from the periphery. Both genetic and pharmacological disruption of the CCR2/CCL2 pathway consequently decreased the BrM load in breast and melanoma mouse models [54].
Novel insights from profiling clinical brain metastases
Despite extensive research efforts, current experimental preclinical models of brain metastasis have been limited by the utilization of largely homogeneous cancer cell lines and immunocompromised mouse hosts when using human-derived tumor cells. Thus, the translational potential of these discoveries has varied. Recent omics profiling efforts, however, have been extremely valuable in building a more comprehensive unbiased view of the patient brain metastasis ecosystem containing heterogenous populations of cancer and stromal cells.
Genomic profiling
A 2022 study led by Nguyen et al. established the Memorial Sloan Kettering—Metastatic Events and Tropisms (MSK-MET) pan-cancer database resource containing clinical and genomic alteration data across 50 different tumor types [55]. Metastatic burden for some of the cancer types was strongly correlated with chromosomal instability, but not for others, highlighting both the intrinsic cancer and target organ differences. For BrM specifically, it was found that BrMs derived from lung adenocarcinoma had a higher incidence of TP53, EGFR mutations, and TERT amplification, while PTEN mutations and PI3K pathway alterations were common in melanoma derived BrMs [55]. Another recent study conducted whole-exome sequencing on patient primary tumors and matched brain metastases deriving from lung (n = 38), breast (n = 21) and RCCs (n = 10). They detected genetic vulnerabilities in pathways such as PI3K/AKT/mTOR, CDK, and HER2/EGFR that could be exploited for targeted treatments. Importantly, many of the BrM alterations were not present in matched primary tumor samples, indicating that the primary tumors and metastases had very divergent developmental trajectories [56]. Similarly, another recent study found MYC, YAP1, and MMP13 to be important genomic alterations in BrM in a cohort of 105 patients with lung adenocarcinoma [57].
scRNA-sequencing
Aimed to discover universal BrM characteristics, independent of the primary tumor of origin, Gonzalez et al. performed single cell RNA sequencing (scRNA-seq) in a large-scale study on more than 100,000 malignant and non-malignant cells harvested from patients’ BrMs. This comprehensive study on 15 human parenchymal BrMs described two distinct and functional BrM archetypes: one proliferative and one inflammatory, as well as characterized the immune infiltrate [58]. Another study utilized scRNA-seq to examine patient breast (n = 3) and lung (n = 2) cancer derived BrMs. They uncovered tumor-associated fibroblasts that produce high levels of collagen and facilitate TME remodeling. Furthermore, they identified tumor-specific collagen receptors that have the potential to become biomarkers of BrM patient survival [59]. These relatively large scale BrM profiling serve as valuable resources to guide further research. However, such studies are still largely under sampled. Additionally, it is currently not possible to examine the impact of different clinical treatments or predict therapy response. Similarly, there is a great need for further pre-clinical studies exploring the role of systemic factors on BrM development.
Multi-omics profiling
The molecular profiles of melanoma-derived BrM have been examined in 88 patient-derived samples and 42 matched extracranial lesions through a combination of RNA, whole exome, T cell receptor (TCR) sequencings, and IHC [60]. It was found that melanoma-derived BrMs had high oxidative phosphorylation (OXPHOS) levels and utilizing an OXPHOS inhibitor could abrogate BrM in mouse xenografts [60]. For lung (n = 21) and breast (n = 9) cancer derived BrM patients specifically, a different study performed whole-exome sequencing, RNA sequencing, flow cytometry, immunofluorescence, and tissue imaging on surgically collected tissue. This study revealed that the immune infiltrate was unique even among BrMs derived from within a single primary tumor of origin group and correlated TP53 mutations with overall infiltration level across all tumors [61]. Another multiome profiling study conducted RNA, exome, low-pass whole-genome sequencing, and global DNA microarrays on metastases from 55 breast cancer patients, of whom 11 were diagnosed with BrM and 28 had liver metastases. In these patient cohorts, patterns of DNA hypermethylation around the genes of major histocompatibility complex (MHC), Class I A, HLA-A, were associated with lower expression of MHC molecules and reduced immune cell infiltration [62]. Yet another valuable profiling study focusing on the immune compartment utilized RNA-seq (n = 48), flow cytometry (n = 100), IF, and ELISA assays to directly compare the immune infiltrate between patient gliomas and BrMs originating from melanoma, breast, and lung cancer. They found that globally, compared to gliomas BrMs exhibited greater immune cell diversity as well as distinct transcriptional programs in the myeloid compartment. Specifically, they observed upregulation of neuroinflammation mediators CSF2 and IL23A as well as the receptor MARCO in BrM microglia and infiltrated macrophages [63].
Cerebrospinal fluid liquid biopsy
Although the characteristics of the immune infiltrate are often used to predict immune checkpoint treatment response, it is often difficult to obtain biopsies from BrM lesions. Thus, a recent study explored the immune composition of cerebrospinal fluid (CSF) of six BrM patients with matched BrM lesions and found that the degree of CD8 T cell infiltration into the tumor can be predicted from T cell abundance in the CSF. Interestingly, TCR clonotypes expanded in BrM lesions can also be detected in the CSF, pointing to active cellular exchange between these two tissue compartments [64]. Another study examined the immune composition of melanoma BrM and leptomeningeal melanoma metastases (LMM). In total, they examined 8 skin samples, 14 melanoma BrMs, and 19 LMM specimens. It was shown that the LMM immune landscape was more immuno-suppressive compared to BrM or primary melanoma and that response to anti-PD-1 therapy was correlated with increased levels of T cells and dendritic cells in the CSF of patients [65].
Perspectives in translational brain metastasis research
A recent viewpoint article by the experts in the field has provided informative insights on the pressing needs in translational BrM research [66]. Moreover, a recent virtual National Cancer Institute (NCI) workshop consisting of BrM patents, advocates, scientists, and clinicians convened in 2022. Together, they outlined unmet needs in translational BrM research and offered guidelines for how to better align translational research with patient-defined meaningful advancements [6]. Here, we will highlight some key points from the workshop and provide additional viewpoints for future BrM research.
Immunotherapy beyond immune checkpoint blockade therapy
Although ICT has been extremely successful in treating certain types of cancer, for patients who do not respond to ICT there are other emerging modalities for enhancing the immune response against metastasized cancer cells [12]. For example, engineered chimeric antigen receptor (CAR) T cells targeting CD19 has been revolutionary in treating hematologic malignancies, such as acute lymphoblastic leukemia (ALL) [15]. More recently, CAR-T cells have been applied to CNS cancer such as glioblastoma. In a 2020 study, Wang et al. exploited a previously described scorpion venom protein chlorotoxin (CLTX) that binds specifically to glioblastoma cells to generate their CAR construct. It was found that CLTX CAR-T therapy results in GBM regression with no observable off target effects in mouse GBM models [67]. Following this study, a Phase I clinical trial was initiated to assess the clinical efficacy of CLTX CAR-T in patients with recurrent glioblastoma, malignant glioma, and WHO grade II and III gliomas (NCT04214392). For CAR T therapy to be effective in BrM, the specific tumor antigen to be targeted needs to be considered carefully, as off target effects in the brain could lead to devastating side effects. A 2018 study in an orthotopic xenograft model of breast cancer-derived BrM demonstrated that local, intracranial delivery of HER2-CAR T cells had potent in vivo effects on reducing BrM burden [68]. Thus, the ability to identify the specific molecular subtype of the primary tumor of origin is of paramount importance for further CAR T developments in BrM treatment.
The lack of clinical efficacy for ICT in certain solid tumors is at least partially attributed to an immunosuppressive microenvironment fueled by tumor-associated macrophages [69]. Indeed, CNS-native microglia and infiltrated macrophages are known to influence BrM outgrowth, consequently, blockade of the macrophage colony-stimulating factor 1 receptor (CSF1R) in a murine breast cancer BrM model led to reduced tumor burden. This response is more durable with dual targeting of CSF1R and microglial STAT5 signaling [70]. Therefore, in addition to CAR-T cell therapy, another immunotherapy approach is to focus efforts on reprogramming the myeloid cell compartment to elicit more efficient cancer cell phagocytosis (directly) or boost the cytotoxic T and natural killer (NK) cell response (indirectly), and potentially serve as drug delivery conduits [69]. The therapeutic utility of myeloid cell engineering has been demonstrated recently. Ex vivo engineered IL-2 secreting myeloid cells can readily cross the BBB after i.v. administration and prolong survival in a mouse model of glioma [71]. As bone marrow derived myeloid cells (BMDMs) are known to infiltrate the brain during BrM [72, 73], studies exploring myeloid cell-based therapies could prove to be very valuable.
Translation-oriented pre-clinical models and experimental designs
Historically, the most widely used models to study BrM rely on the utilization of brain-seeking cell lines generated through multiple rounds of in vivo selection of the original parental cell line with lesser metastatic propensity. Cancer cells are typically delivered via intracardiac (left ventricle) or intracarotid artery (ICA) injections [74]. Although the models provided novel insights into basic BrM biology, the limited genetic repertoire of cancer cell lines, common utilization of immunocompromised hosts, and treatment-naïve conditions have reduced the clinical relevance and translational impact of these findings. In this section, we will highlight some emerging pre-clinical models and future study design priorities for addressing the historical BrM pre-clinical research limitations.
Patient-derived xenografts (PDXs) from BrM are valuable models typically generated by transplanting patient tumors subcutaneously or orthotopically into immune-deficient mice to expand the tumors, followed by systemic or local injection for BrM establishment or generation of patient-derived cultures that can be cultured at low passages. PDX models have the advantage of more faithfully recapitulating the diverse and heterogenous genomic features of patient tumors [74]. However, in vivo PDX models in immune-deficient hosts lack immune elements which are critical for BrM success and immunotherapy testing. The recent progress in creating humanized mouse models holds potential for overcoming the limitation of current PDX models [75].
The prevailing BrM models based on tumor cell injection only recapitulate the later steps of the metastatic cascade. The ideal mouse model of BrM would rely on spontaneous metastasis, originating from either a syngeneic cell line implanted orthotopically or naturally developed in genetically engineered mouse models (GEMs). These model systems would recapitulate the entire metastatic cascade as well as allow for the examination of the pre-metastatic niche. Unfortunately, very few cell lines can efficiently metastasize to the brain in an immunocompetent host, limiting their practicality in a preclinical setting [74]. Development of primary tumor models that have more reliable and reproducible spontaneous BrM capability would significantly enhance both mechanistic and translational BrM studies.
Very few pre-clinical studies at present include clinically relevant neurosurgical resection, radiation therapy and/or concurrent systemic treatments. Most pre-clinical BrM research is conducted in treatment-naïve conditions, yet patients diagnosed with BrM have often undergone several rounds of systemic therapy for the primary tumor, followed by neurosurgery, radiotherapy, chemotherapy, targeted therapy, or even ICT, upon BrM diagnosis [74]. As radiotherapy (SRS or WBRT) is the standard of care in the clinical management of brain metastasis patients regardless of primary tumor of origin [3, 8, 9, 76,77,78,79,80,81] (Table 1), future preclinical studies under the context of radiation or systemic treatment would reveal valuable mechanisms that could be more relevant to clinical trial settings. A recent 2019 review article outlined the utility and pitfalls of various mouse models and currently available small animal radiotherapy platforms. This review can serve as a resource for pre-clinical researchers interested in incorporating radiotherapy modalities most applicable to the current clinical practice of tumor radiotherapy treatment [82].
Finally, as outlined in the 2020 NCI workshop patient-guided priorities, there is a critical need to improve health span metric data collection in BrM clinical trials [6]. The BrM intracranial mass leads to the development of a unique and devastating set of neurological symptoms and cognitive decline which vary depending on tumor size and anatomical location within the brain. The aggressive treatments used in BrM management such as radiotherapy and surgical resection can further exacerbate neurocognitive symptoms [83]. As this aspect of the disease can be a significant concern for the patient and their family, a high priority for developing novel pre-clinical animal models is incorporating assessments of overall health span metrics and neurological function, as compared to only assessing gross tumor burden.
Novel technologies
High throughput transcriptomic techniques such as scRNA-seq have revolutionized our understanding of cellular heterogeneity of tissues in steady state and various diseases. However, as these techniques are not performed in situ, they do not provide information about the spatial relationship among different cells within the critical niche of the metastasis. The advent of next-generation sequencing (NGS)-based and imaging-based spatial transcriptomics technologies provides a novel avenue for exploring the tumor and metastatic microenvironment [84]. Although NGS-based spatial transcriptomics platforms such as Visium have advanced our understanding of tumor microenvironment architecture [85, 86], they are not yet able to capture transcriptomes at a single cell level. To circumvent inherent limitations of both single cell and spatial methods, recent efforts have explored ways to integrate scRNA-seq, single-nucleus RNA-seq, and spatial information. Given the unique tissue architecture of the BrM microenvironment, jointed analysis of single cell transcriptome and spatial transcriptomic analyses could provide unique mechanistic insights into the spatial relationship between BrM cancer cells and the niche, which could potentially infer novel therapeutic targets. For example, a 2022 study led by Biermann et al. conducted single cell/nucleus RNA-seq, spatial single-cell transcriptomics and TCR sequencing in treatment naïve melanoma BrM patients [87]. They made key discoveries in both metastasized cancer cells and immune infiltrates. In cancer cells they found high levels of chromosomal instability, neuronal-like programs, and spatially variable metabolic alterations. In the BrM TME, lymphoid aggregates harboring B cells actively differentiating into plasma cells, infiltration of monocyte-derived macrophages, and dysfunctional T cells [87]. In addition to these transcriptomic analysis advancements, on the protein level, advances in tissue sequential immunofluorescence (IF) approaches such as the COMET™ (Lunaphore) platform will allow researchers to multiplex many more proteins on the same slide.
CRISPR/Cas9 based genetic screening technology is another emerging technology that could significantly reduce the time required to identify novel druggable targets in BrM and could work synergistically with profiling approaches described above. In the classical CRISPR “drop-out” genetic screen, depletion of specific guide RNAs (gRNAs) identifies genes which are essential for survival under the specified selection conditions [88]. Such in vitro screens conducted on various cell lines elucidated novel synthetic lethality and drug resistance pathways [89, 90]. Although valuable, this simplified 2D in vitro screening approach cannot completely model the heterogenous in vivo metastatic microenvironment. Thus, more recent studies are exploring organoid [91] and in vivo CRISPR screening approaches alongside scRNA-seq or using patient-derived xenografts (PDXs) [88, 92,93,94,95], further broadening the scope of information that can be obtained.
Pushing the boundary of clinical diagnosis
Clinical presentation of an intracranial BrM lesion includes symptoms such as headache (50% patients), papilledema (15–25%), focal neurological deficits (40%), and seizures (15–20%). Currently, the gold standard for diagnosing BrM is histopathological analysis of brain tissue involved. Neuroimaging techniques such as brain contrast-enhanced magnetic resonance imaging (MRI) can also be used to assess the global BrM burden [1]. Moreover, MRI imaging techniques are used throughout the treatment process to monitor therapeutic effects in a less invasive manner. Novel imaging modalities such as quantitative magnetic transfer (qMT) and magnetic resonance spectroscopy (MRS) are currently being evaluated for their sensitivity in distinguishing between different types of brain tumors and detection thresholds and is an emerging area with several clinical trials underway [96].
Pushing the sensitivity of imaging modalities is highly impactful for BrM early detection. One promising approach to increase the sensitivity of existing MRIs is increasing image contrast with BrM-targeted nanoparticles. A phase 1 clinical trial was conducted where 15 patients were injected with a single intravenous dose of gadolinium(Gd)-based AGuIX nanoparticles [97]. Notably, these BrM patients represented four different primary tumors of origin: melanoma, lung, colon, and breast cancer. Upon administration, the nanoparticles were able to home to the brain and increase MRI image contrast quality [97]. Similarly, a combination of neuroimaging and nanoparticles can be used to assess the status of the BBB for both integrity and heterogeneity. Currently, contrast-enhanced MRI is the gold standard for BBB assessment, however positron emission tomography (PET) and intravital microscopy can also be utilized. These approaches are an emerging clinical priority, not just for the detection of BrM lesions, but additionally for assessing BBB/BTB heterogeneity before and after implementing therapeutic interventions [28]. Perspectives in preclinical BrM imaging have been surveyed in a valuable 2021 review, which discussed recent MRI and PET strategies currently being applied in animal BrM models and provided recommendations for clinical translation [98].
Conclusion
In this article, we reviewed recent research efforts related to the translational perspectives of brain metastasis. The rise of single-cell omics technologies has opened a new era of unbiased studies for clinical BrMs and provided much-needed resources and insights for dissecting the extremely heterogeneous BrM ecosystem. However, the gap between preclinical studies and clinical profiling studies has widened. Given that BrM is a clinical challenge, there is a pressing need to develop preclinical models and conduct preclinical research under more clinically relevant treatment contexts. Here, we have summarized the potential high priority areas for future pre-clinical studies (Table 2). Continuing to expand preclinical research efforts on basic cellular mechanisms with more considerations to the current clinical practices will be pivotal to reduce the gap between basic discoveries and novel therapeutics. We believe this can be achieved through leveraging novel technologies adopted from different fields with emerging pre-clinical models.
References
Achrol AS, Rennert RC, Anders C, Soffietti R, Ahluwalia MS, Nayak L, Peters S, Arvold ND, Harsh GR, Steeg PS et al (2019) Brain metastases. Nat Rev Dis Primer 5:1–26. https://doi.org/10.1038/s41572-018-0055-y
Deshpande K, Buchanan I, Martirosian V, Neman J (2020) Clinical perspectives in brain metastasis. Cold Spring Harb Perspect Med. https://doi.org/10.1101/cshperspect.a037051
Imaizumi J, Shida D, Narita Y, Miyakita Y, Tanabe T, Takashima A, Boku N, Igaki H, Itami J, Kanemitsu Y (2019) Prognostic factors of brain metastases from colorectal cancer. BMC Cancer 19:755. https://doi.org/10.1186/s12885-019-5973-x
Singh R, Stoltzfus KC, Chen H, Louie AV, Lehrer EJ, Horn SR, Palmer JD, Trifiletti DM, Brown PD, Zaorsky NG (2020) Epidemiology of synchronous brain metastases. Neuro-Oncol Adv. https://doi.org/10.1093/noajnl/vdaa041
Mehrian-Shai R, Reichardt JKV, Harris CC, Toren A (2019) The gut-brain axis, paving the way to brain cancer. Trends Cancer 5:200–207. https://doi.org/10.1016/j.trecan.2019.02.008
Kim MM, Mehta MP, Smart DK, Steeg PS, Hong JA, Espey MG, Prasanna PG, Crandon L, Hodgdon C, Kozak N et al (2023) National cancer institute collaborative workshop on shaping the landscape of brain metastases research: challenges and recommended priorities. Lancet Oncol 24:e344–e354. https://doi.org/10.1016/S1470-2045(23)00297-8
Brown PD, Gondi V, Pugh S, Tome WA, Wefel JS, Armstrong TS, Bovi JA, Robinson C, Konski A, Khuntia D et al (2020) Hippocampal avoidance during whole-brain radiotherapy plus memantine for patients with brain metastases: phase III Trial NRG oncology CC001. J Clin Oncol Off J Am Soc Clin Oncol 38:1019–1029. https://doi.org/10.1200/JCO.19.02767
Stavrou E, Winer EP, Lin NU (2021) How we treat HER2-positive brain metastases. ESMO Open 6:100256. https://doi.org/10.1016/j.esmoop.2021.100256
Bailleux C, Eberst L, Bachelot T (2021) Treatment strategies for breast cancer brain metastases. Br J Cancer 124:142–155. https://doi.org/10.1038/s41416-020-01175-y
Montemurro F, Delaloge S, Barrios CH, Wuerstlein R, Anton A, Brain E, Hatschek T, Kelly CM, Peña-Murillo C, Yilmaz M et al (2020) Trastuzumab emtansine (T-DM1) in patients with HER2-positive metastatic breast cancer and brain metastases: exploratory final analysis of cohort 1 from KAMILLA, a single-arm phase IIIb clinical trial☆. Ann Oncol 31:1350–1358. https://doi.org/10.1016/j.annonc.2020.06.020
Nelson TA, Wang N (2023) Targeting lung cancer brain metastases: a narrative review of emerging insights for anaplastic lymphoma kinase (ALK)-positive disease. Transl Lung Cancer Res 12:379–392. https://doi.org/10.21037/tlcr-22-638
Shiravand Y, Khodadadi F, Kashani SMA, Hosseini-Fard SR, Hosseini S, Sadeghirad H, Ladwa R, O’Byrne K, Kulasinghe A (2022) Immune checkpoint inhibitors in cancer therapy. Curr Oncol 29:3044–3060. https://doi.org/10.3390/curroncol29050247
Di Lorenzo R, Ahluwalia MS (2017) Targeted therapy of brain metastases: latest evidence and clinical implications. Ther Adv Med Oncol 9:781–796. https://doi.org/10.1177/1758834017736252
Wang Y, Zhang Q, Chen C, Hu Y, Miao L, Zhou Y (2021) Association of brain metastases with immune checkpoint inhibitors efficacy in advanced lung cancer: a systematic review and meta-analysis. Front Oncol 11:721760. https://doi.org/10.3389/fonc.2021.721760
Nieblas-Bedolla E, Nayyar N, Singh M, Sullivan RJ, Brastianos PK (2021) Emerging immunotherapies in the treatment of brain metastases. Oncologist 26:231–241. https://doi.org/10.1002/onco.13575
Tawbi HA, Forsyth PA, Algazi A, Hamid O, Hodi FS, Moschos SJ, Khushalani NI, Lewis K, Lao CD, Postow MA et al (2018) Combined nivolumab and ipilimumab in melanoma metastatic to the brain. N Engl J Med 379:722–730. https://doi.org/10.1056/NEJMoa1805453
Rausch MP, Hastings KT (2017) Immune checkpoint inhibitors in the treatment of melanoma: from basic science to clinical application. In: Ward WH, Farma JM (eds) Cutaneous melanoma: etiology and therapy. Codon Publications, Singapore
Zhu S, Wang Y, Tang J, Cao M (2022) Radiotherapy induced immunogenic cell death by remodeling tumor immune microenvironment. Front Immunol 13:1074477. https://doi.org/10.3389/fimmu.2022.1074477
Nabrinsky E, Macklis J, Bitran J (2022) A review of the abscopal effect in the era of immunotherapy. Cureus 14:e29620. https://doi.org/10.7759/cureus.29620
Sevenich L (2019) Turning “cold” into “hot” tumors-opportunities and challenges for radio-immunotherapy against primary and metastatic brain cancers. Front Oncol 9:163. https://doi.org/10.3389/fonc.2019.00163
Füreder LM, Widhalm G, Gatterbauer B, Dieckmann K, Hainfellner JA, Bartsch R, Zielinski CC, Preusser M, Berghoff AS (2018) Brain metastases as first manifestation of advanced cancer: exploratory analysis of 459 patients at a tertiary care center. Clin Exp Metastasis 35:727–738. https://doi.org/10.1007/s10585-018-9947-1
Balestrino R, Rudà R, Soffietti R (2020) Brain metastasis from unknown primary tumour: moving from old retrospective studies to clinical trials on targeted agents. Cancers 12:3350. https://doi.org/10.3390/cancers12113350
Clynick B, Dessauvagie B, Sterrett G, Harvey NT, Allcock RJN, Saunders C, Erber W, Meehan K (2018) Genetic characterisation of molecular targets in carcinoma of unknown primary. J Transl Med 16:185. https://doi.org/10.1186/s12967-018-1564-x
Verginelli F, Pisacane A, Gambardella G, D’Ambrosio A, Candiello E, Ferrio M, Panero M, Casorzo L, Benvenuti S, Cascardi E et al (2021) Cancer of unknown primary stem-like cells model multi-organ metastasis and unveil liability to MEK inhibition. Nat Commun 12:2498. https://doi.org/10.1038/s41467-021-22643-w
Angeli E, Nguyen TT, Janin A, Bousquet G (2019) How to make anticancer drugs cross the blood-brain barrier to treat brain metastases. Int J Mol Sci 21:22. https://doi.org/10.3390/ijms21010022
Bönig L, Möhn N, Ahlbrecht J, Wurster U, Raab P, Puppe W, Sühs K-W, Stangel M, Skripuletz T, Schwenkenbecher P (2019) Leptomeningeal metastasis: the role of cerebrospinal fluid diagnostics. Front Neurol 10:839
Steeg PS (2021) The blood–tumour barrier in cancer biology and therapy. Nat Rev Clin Oncol 18:696–714. https://doi.org/10.1038/s41571-021-00529-6
Arvanitis CD, Ferraro GB, Jain RK (2020) The blood–brain barrier and blood–tumour barrier in brain tumours and metastases. Nat Rev Cancer 20:26–41. https://doi.org/10.1038/s41568-019-0205-x
Upton DH, Ung C, George SM, Tsoli M, Kavallaris M, Ziegler DS (2022) Challenges and opportunities to penetrate the blood-brain barrier for brain cancer therapy. Theranostics 12:4734–4752. https://doi.org/10.7150/thno.69682
Arvanitis CD, Askoxylakis V, Guo Y, Datta M, Kloepper J, Ferraro GB, Bernabeu MO, Fukumura D, McDannold N, Jain RK (2018) Mechanisms of enhanced drug delivery in brain metastases with focused ultrasound-induced blood–tumor barrier disruption. Proc Natl Acad Sci USA 115:E8717–E8726. https://doi.org/10.1073/pnas.1807105115
Park J, Aryal M, Vykhodtseva N, Zhang Y-Z, McDannold N (2017) Evaluation of permeability, doxorubicin delivery, and drug retention in a rat brain tumor model after ultrasound-induced blood-tumor barrier disruption. J. Control Release Off J Control Release Soc 250:77–85. https://doi.org/10.1016/j.jconrel.2016.10.011
Arsiwala TA, Blethen KE, Wolford CP, Panchal DM, Sprowls SA, Fladeland RA, Kielkowski BN, Pritt TA, Wang P, Wilson O et al (2023) Blood-tumor barrier opening by MRI-guided transcranial focused ultrasound in a preclinical breast cancer brain metastasis model improves efficacy of combinatorial chemotherapy. Front Oncol 13:1104594. https://doi.org/10.3389/fonc.2023.1104594
Campbell BK, Gao Z, Corcoran NM, Stylli SS, Hovens CM (2022) Molecular mechanisms driving the formation of brain metastases. Cancers 14:4963. https://doi.org/10.3390/cancers14194963
Kim K, Marquez-Palencia M, Malladi S (2019) Metastatic latency, a veiled threat. Front Immunol 10:1836
Flüh C, Mafael V, Adamski V, Synowitz M, Held-Feindt J (2020) Dormancy and NKG2D system in brain metastases: analysis of immunogenicity. Int J Mol Med 45:298–314. https://doi.org/10.3892/ijmm.2019.4449
Blasco MT, Espuny I, Gomis RR (2022) Ecology and evolution of dormant metastasis. Trends Cancer 8:570–582. https://doi.org/10.1016/j.trecan.2022.03.002
Dai J, Cimino PJ, Gouin KH, Grzelak CA, Barrett A, Lim AR, Long A, Weaver S, Saldin LT, Uzamere A et al (2022) Astrocytic laminin-211 drives disseminated breast tumor cell dormancy in brain. Nat Cancer 3:25–42. https://doi.org/10.1038/s43018-021-00297-3
Sayan M, Zoto Mustafayev T, Sahin B, Kefelioglu ESS, Wang S-J, Kurup V, Balmuk A, Gungor G, Ohri N, Weiner J et al (2019) Evaluation of response to stereotactic radiosurgery in patients with radioresistant brain metastases. Radiat Oncol J 37:265–270. https://doi.org/10.3857/roj.2019.00409
Monteiro C, Miarka L, Perea-García M, Priego N, García-Gómez P, Álvaro-Espinosa L, de Pablos-Aragoneses A, Yebra N, Retana D, Baena P et al (2022) Stratification of radiosensitive brain metastases based on an actionable S100A9/RAGE resistance mechanism. Nat Med 28:752–765. https://doi.org/10.1038/s41591-022-01749-8
Biswas AK, Han S, Tai Y, Ma W, Coker C, Quinn SA, Shakri AR, Zhong TJ, Scholze H, Lagos GG et al (2022) Targeting S100A9-ALDH1A1-retinoic acid signaling to suppress brain relapse in EGFR-mutant lung cancer. Cancer Discov 12:1002–1021. https://doi.org/10.1158/2159-8290.CD-21-0910
Adua SJ, Arnal-Estapé A, Zhao M, Qi B, Liu ZZ, Kravitz C, Hulme H, Strittmatter N, López-Giráldez F, Chande S et al (2022) Brain metastatic outgrowth and osimertinib resistance are potentiated by RhoA in EGFR-mutant lung cancer. Nat Commun 13:7690. https://doi.org/10.1038/s41467-022-34889-z
Martínez-Reyes I, Chandel NS (2021) Cancer metabolism: looking forward. Nat Rev Cancer 21:669–680. https://doi.org/10.1038/s41568-021-00378-6
Tyagi A, Wu S-Y, Watabe K (2022) Metabolism in the progression and metastasis of brain tumors. Cancer Lett 539:215713. https://doi.org/10.1016/j.canlet.2022.215713
Parida PK, Marquez-Palencia M, Ghosh S, Khandelwal N, Kim K, Nair V, Liu X-Z, Vu HS, Zacharias LG, Gonzalez-Ericsson PI et al (2023) Limiting mitochondrial plasticity by targeting DRP1 induces metabolic reprogramming and reduces breast cancer brain metastases. Nat Cancer 4:893–907. https://doi.org/10.1038/s43018-023-00563-6
Parida PK, Marquez-Palencia M, Nair V, Kaushik AK, Kim K, Sudderth J, Quesada-Diaz E, Cajigas A, Vemireddy V, Gonzalez-Ericsson PI et al (2022) Metabolic diversity within breast cancer brain-tropic cells determines metastatic fitness. Cell Metab 34:90-105.e7. https://doi.org/10.1016/j.cmet.2021.12.001
Srinivasan ES, Deshpande K, Neman J, Winkler F, Khasraw M (2021) The microenvironment of brain metastases from solid tumors. Neuro-Oncol Adv 3:v121–v132. https://doi.org/10.1093/noajnl/vdab121
Chuang H-N, van Rossum D, Sieger D, Siam L, Klemm F, Bleckmann A, Bayerlová M, Farhat K, Scheffel J, Schulz M et al (2013) Carcinoma cells misuse the host tissue damage response to invade the brain. Glia 61:1331–1346. https://doi.org/10.1002/glia.22518
Caffarel MM, Braza MS (2022) Microglia and metastases to the central nervous system: victim, ravager, or something else? J Exp Clin Cancer Res 41:327. https://doi.org/10.1186/s13046-022-02535-7
Shen M, Kang Y (2018) pSTAT3+ reactive astrocytes promote brain metastasis. Trends Mol Med 24:733–735. https://doi.org/10.1016/j.molmed.2018.07.002
Zhang L, Zhang S, Yao J, Lowery FJ, Zhang Q, Huang W-C, Li P, Li M, Wang X, Zhang C et al (2015) Microenvironment-induced PTEN loss by exosomal microRNA primes brain metastasis outgrowth. Nature 527:100–104. https://doi.org/10.1038/nature15376
Zou Y, Watters A, Cheng N, Perry CE, Xu K, Alicea GM, Parris JLD, Baraban E, Ray P, Nayak A et al (2019) Polyunsaturated fatty acids from astrocytes activate PPARγ signaling in cancer cells to promote brain metastasis. Cancer Discov 9:1720–1735. https://doi.org/10.1158/2159-8290.CD-19-0270
Guldner IH, Wang Q, Yang L, Golomb SM, Zhao Z, Lopez JA, Brunory A, Howe EN, Zhang Y, Palakurthi B et al (2020) CNS-native myeloid cells drive immune suppression in the brain metastatic niche through Cxcl10. Cell 183:1234-1248.e25. https://doi.org/10.1016/j.cell.2020.09.064
Zhang L, Yao J, Wei Y, Zhou Z, Li P, Qu J, Badu-Nkansah A, Yuan X, Huang Y-W, Fukumura K et al (2020) Blocking immunosuppressive neutrophils deters pY696-EZH2–driven brain metastases. Sci Transl Med. https://doi.org/10.1126/scitranslmed.aaz5387
Ma W, Oliveira-Nunes MC, Xu K, Kossenkov A, Reiner BC, Crist RC, Hayden J, Chen Q (2023) Type I interferon response in astrocytes promotes brain metastasis by enhancing monocytic myeloid cell recruitment. Nat Commun 14:2632. https://doi.org/10.1038/s41467-023-38252-8
Nguyen B, Fong C, Luthra A, Smith SA, DiNatale RG, Nandakumar S, Walch H, Chatila WK, Madupuri R, Kundra R et al (2022) Genomic characterization of metastatic patterns from prospective clinical sequencing of 25,000 patients. Cell 185:563-575.e11. https://doi.org/10.1016/j.cell.2022.01.003
Brastianos PK, Carter SL, Santagata S, Cahill DP, Taylor-Weiner A, Jones RT, Van Allen EM, Lawrence MS, Horowitz PM, Cibulskis K et al (2015) Genomic characterization of brain metastases reveals branched evolution and potential therapeutic targets. Cancer Discov 5:1164–1177. https://doi.org/10.1158/2159-8290.CD-15-0369
Shih DJH, Nayyar N, Bihun I, Dagogo-Jack I, Gill CM, Aquilanti E, Bertalan M, Kaplan A, D’Andrea MR, Chukwueke U et al (2020) Genomic characterization of human brain metastases identifies drivers of metastatic lung adenocarcinoma. Nat Genet 52:371–377. https://doi.org/10.1038/s41588-020-0592-7
Gonzalez H, Mei W, Robles I, Hagerling C, Allen BM, Hauge Okholm TL, Nanjaraj A, Verbeek T, Kalavacherla S, van Gogh M et al (2022) Cellular architecture of human brain metastases. Cell 185:729-745.e20. https://doi.org/10.1016/j.cell.2021.12.043
Song Q, Ruiz J, Xing F, Lo H-W, Craddock L, Pullikuth AK, Miller LD, Soike MH, O’Neill SS, Watabe K et al (2023) Single-cell sequencing reveals the landscape of the human brain metastatic microenvironment. Commun Biol 6:1–13. https://doi.org/10.1038/s42003-023-05124-2
Fischer GM, Jalali A, Kircher DA, Lee W-C, McQuade JL, Haydu LE, Joon AY, Reuben A, de Macedo MP, Carapeto FCL et al (2019) Molecular profiling reveals unique immune and metabolic features of melanoma brain metastases. Cancer Discov 9:628–645. https://doi.org/10.1158/2159-8290.CD-18-1489
Álvarez-Prado ÁF, Maas RR, Soukup K, Klemm F, Kornete M, Krebs FS, Zoete V, Berezowska S, Brouland J-P, Hottinger AF et al (2023) Immunogenomic analysis of human brain metastases reveals diverse immune landscapes across genetically distinct tumors. Cell Rep Med. https://doi.org/10.1016/j.xcrm.2022.100900
Garcia-Recio S, Hinoue T, Wheeler GL, Kelly BJ, Garrido-Castro AC, Pascual T, De Cubas AA, Xia Y, Felsheim BM, McClure MB et al (2023) Multiomics in primary and metastatic breast tumors from the AURORA US network finds microenvironment and epigenetic drivers of metastasis. Nat Cancer 4:128–147. https://doi.org/10.1038/s43018-022-00491-x
Klemm F, Maas RR, Bowman RL, Kornete M, Soukup K, Nassiri S, Brouland J-P, Iacobuzio-Donahue CA, Brennan C, Tabar V et al (2020) Interrogation of the microenvironmental landscape in brain tumors reveals disease-specific alterations of immune cells. Cell 181:1643-1660.e17. https://doi.org/10.1016/j.cell.2020.05.007
Rubio-Perez C, Planas-Rigol E, Trincado JL, Bonfill-Teixidor E, Arias A, Marchese D, Moutinho C, Serna G, Pedrosa L, Iurlaro R et al (2021) Immune cell profiling of the cerebrospinal fluid enables the characterization of the brain metastasis microenvironment. Nat Commun 12:1503. https://doi.org/10.1038/s41467-021-21789-x
Smalley I, Chen Z, Phadke M, Li J, Yu X, Wyatt C, Evernden B, Messina JL, Sarnaik A, Sondak VK et al (2021) Single-cell characterization of the immune microenvironment of melanoma brain and leptomeningeal metastases. Clin Cancer Res 27:4109–4125. https://doi.org/10.1158/1078-0432.CCR-21-1694
Boire A, Brastianos PK, Garzia L, Valiente M (2020) Brain metastasis. Nat Rev Cancer 20:4–11. https://doi.org/10.1038/s41568-019-0220-y
Wang D, Starr R, Chang W-C, Aguilar B, Alizadeh D, Wright SL, Yang X, Brito A, Sarkissian A, Ostberg JR et al (2020) Chlorotoxin-directed CAR T cells for specific and effective targeting of glioblastoma. Sci Transl Med. https://doi.org/10.1126/scitranslmed.aaw2672
Priceman SJ, Tilakawardane D, Jeang B, Aguilar B, Murad JP, Park AK, Chang W-C, Ostberg JR, Neman J, Jandial R et al (2018) Regional delivery of chimeric antigen receptor-engineered T cells effectively targets HER2+ breast cancer metastasis to the brain. Clin Cancer Res 24:95–105. https://doi.org/10.1158/1078-0432.CCR-17-2041
Canella A, Rajappa P (2023) Therapeutic utility of engineered myeloid cells in the tumor microenvironment. Cancer Gene Ther 30:964–972. https://doi.org/10.1038/s41417-023-00600-7
Klemm F, Möckl A, Salamero-Boix A, Alekseeva T, Schäffer A, Schulz M, Niesel K, Maas RR, Groth M, Elie BT et al (2021) Compensatory CSF2-driven macrophage activation promotes adaptive resistance to CSF1R inhibition in breast-to-brain metastasis. Nat Cancer 2:1086–1101. https://doi.org/10.1038/s43018-021-00254-0
Canella A, Nazzaro M, Rajendran S, Schmitt C, Haffey A, Nigita G, Thomas D, Lyberger JM, Behbehani GK, Amankulor NM et al (2023) Genetically modified IL2 bone-marrow-derived myeloid cells reprogram the glioma immunosuppressive tumor microenvironment. Cell Rep. https://doi.org/10.1016/j.celrep.2023.112891
You H, Baluszek S, Kaminska B (2019) Immune microenvironment of brain metastases—are microglia and other brain macrophages little helpers? Front Immunol 10:1941
Bowman RL, Klemm F, Akkari L, Pyonteck SM, Sevenich L, Quail DF, Dhara S, Simpson K, Gardner EE, Iacobuzio-Donahue CA et al (2016) Macrophage ontogeny underlies differences in tumor-specific education in brain malignancies. Cell Rep 17:2445–2459. https://doi.org/10.1016/j.celrep.2016.10.052
Miarka L, Valiente M (2021) Animal models of brain metastasis. Neuro-Oncol Adv 3:v144–v156. https://doi.org/10.1093/noajnl/vdab115
Chuprin J, Buettner H, Seedhom MO, Greiner DL, Keck JG, Ishikawa F, Shultz LD, Brehm MA (2023) Humanized mouse models for immuno-oncology research. Nat Rev Clin Oncol 20:192–206. https://doi.org/10.1038/s41571-022-00721-2
Ernani V, Stinchcombe TE (2019) Management of brain metastases in non-small-cell lung cancer. J Oncol Pract 15:563–570. https://doi.org/10.1200/JOP.19.00357
Kepka L, Socha J, Sas-Korczynska B (2021) Radiotherapy for brain metastases from small-cell lung cancer in distinct clinical indications and scenarios. J Thorac Dis 13:3269–3278. https://doi.org/10.21037/jtd.2019.10.51
Tan X-L, Le A, Lam FC, Scherrer E, Kerr RG, Lau AC, Han J, Jiang R, Diede SJ, Shui IM (2022) Current treatment approaches and global consensus guidelines for brain metastases in melanoma. Front Oncol 12:885472. https://doi.org/10.3389/fonc.2022.885472
Matsui Y (2020) Current multimodality treatments against brain metastases from renal cell carcinoma. Cancers 12:2875. https://doi.org/10.3390/cancers12102875
Guo L, Shao W, Zhou C, Yang H, Yang L, Cai Q, Wang J, Shi Y, Huang L, Zhang J (2023) Neratinib for HER2-positive breast cancer with an overlooked option. Mol Med 29:134. https://doi.org/10.1186/s10020-023-00736-0
Sidaway P (2023) T-DXd is effective after T-DM1. Nat Rev Clin Oncol 20:426–426. https://doi.org/10.1038/s41571-023-00779-6
Butterworth KT (2019) Evolution of the supermodel: progress in modelling radiotherapy response in mice. Clin Oncol 31:272–282. https://doi.org/10.1016/j.clon.2019.02.008
Wefel JS, Parsons MW, Gondi V, Brown PD (2018) Neurocognitive aspects of brain metastasis. Handb Clin Neurol 149:155–165. https://doi.org/10.1016/B978-0-12-811161-1.00012-8
Rao A, Barkley D, França GS, Yanai I (2021) Exploring tissue architecture using spatial transcriptomics. Nature 596:211–220. https://doi.org/10.1038/s41586-021-03634-9
Ji AL, Rubin AJ, Thrane K, Jiang S, Reynolds DL, Meyers RM, Guo MG, George BM, Mollbrink A, Bergenstråhle J et al (2020) Multimodal analysis of composition and spatial architecture in human squamous cell carcinoma. Cell 182:497-514.e22. https://doi.org/10.1016/j.cell.2020.05.039
Yu Q, Jiang M, Wu L (2022) Spatial transcriptomics technology in cancer research. Front Oncol 12:1019111
Biermann J, Melms JC, Amin AD, Wang Y, Caprio LA, Karz A, Tagore S, Barrera I, Ibarra-Arellano MA, Andreatta M et al (2022) Dissecting the treatment-naive ecosystem of human melanoma brain metastasis. Cell 185:2591-2608.e30. https://doi.org/10.1016/j.cell.2022.06.007
Bock C, Datlinger P, Chardon F, Coelho MA, Dong MB, Lawson KA, Lu T, Maroc L, Norman TM, Song B et al (2022) High-content CRISPR screening. Nat Rev Methods Primer 2:1–23. https://doi.org/10.1038/s43586-021-00093-4
Wang C, Wang G, Feng X, Shepherd P, Zhang J, Tang M, Chen Z, Srivastava M, McLaughlin ME, Navone NM et al (2019) Genome-wide CRISPR screens reveal synthetic lethality of RNASEH2 deficiency and ATR inhibition. Oncogene 38:2451–2463. https://doi.org/10.1038/s41388-018-0606-4
Alyateem G, Wade HM, Bickert AA, Lipsey CC, Mondal P, Smith MD, Labib RM, Mock BA, Robey RW, Gottesman MM (2023) Use of CRISPR-based screens to identify mechanisms of chemotherapy resistance. Cancer Gene Ther 30:1043–1050. https://doi.org/10.1038/s41417-023-00608-z
Mircetic J, Camgöz A, Abohawya M, Ding L, Dietzel J, Tobar SG, Paszkowski-Rogacz M, Seidlitz T, Schmäche T, Mehnert M-C et al (2023) CRISPR/Cas9 screen in gastric cancer patient-derived organoids reveals KDM1A-NDRG1 axis as a targetable vulnerability. Small Methods 7:e2201605. https://doi.org/10.1002/smtd.202201605
Dubrot J, Du PP, Lane-Reticker SK, Kessler EA, Muscato AJ, Mehta A, Freeman SS, Allen PM, Olander KE, Ockerman KM et al (2022) In vivo CRISPR screens reveal the landscape of immune evasion pathways across cancer. Nat Immunol 23:1495–1506. https://doi.org/10.1038/s41590-022-01315-x
Kuhn M, Santinha AJ, Platt RJ (2021) Moving from in vitro to in vivo CRISPR screens. Gene Genome Ed 2:100008. https://doi.org/10.1016/j.ggedit.2021.100008
Dixit A, Parnas O, Li B, Chen J, Fulco CP, Jerby-Arnon L, Marjanovic ND, Dionne D, Burks T, Raychowdhury R et al (2016) Perturb-seq: dissecting molecular circuits with scalable single-cell RNA profiling of pooled genetic screens. Cell 167:1853-1866.e17. https://doi.org/10.1016/j.cell.2016.11.038
Lin S, Larrue C, Scheidegger NK, Seong BKA, Dharia NV, Kuljanin M, Wechsler CS, Kugener G, Robichaud AL, Conway AS et al (2022) An in vivo CRISPR screening platform for prioritizing therapeutic targets in AML. Cancer Discov 12:432–449. https://doi.org/10.1158/2159-8290.CD-20-1851
Tong E, McCullagh KL, Iv M (2020) Advanced imaging of brain metastases: from augmenting visualization and improving diagnosis to evaluating treatment response. Front Neurol 11:270
Verry C, Dufort S, Lemasson B, Grand S, Pietras J, Troprès I, Crémillieux Y, Lux F, Mériaux S, Larrat B et al (2020) Targeting brain metastases with ultrasmall theranostic nanoparticles, a first-in-human trial from an MRI perspective. Sci Adv. https://doi.org/10.1126/sciadv.aay5279
Aasen SN, Espedal H, Keunen O, Adamsen TCH, Bjerkvig R, Thorsen F (2021) Current landscape and future perspectives in preclinical MR and PET imaging of brain metastasis. Neuro-Oncol Adv. https://doi.org/10.1093/noajnl/vdab151
Acknowledgements
We thank both members in both Zhang and Yu laboratories for providing valuable discussions on the concepts presented in this article.
Funding
This work was partially supported by National Institutes of Health (NIH) grants R01CA222405 (SZ), R01CA255064 (SZ), RR220024 (SZ), R01CA266099 (DY), R01CA208213 (DY), R01CA231149 (DY), METAvivor research grants #56675 and #58284 (DY). D.Y. is the Hubert L. & Olive Stringer Distinguished Chair in Basic Science at MD Anderson Cancer Center.
Author information
Authors and Affiliations
Contributions
E.A., S.Z. and D.Y conceived the concepts and wrote the main manuscript text.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Aleksandrovic, E., Zhang, S. & Yu, D. From pre-clinical to translational brain metastasis research: current challenges and emerging opportunities. Clin Exp Metastasis 41, 187–198 (2024). https://doi.org/10.1007/s10585-024-10271-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10585-024-10271-9