
Abstract
In migratory birds, increasing temperatures have been linked to earlier arrival to breeding sites, enabling an earlier start of breeding and leading to changes in abundance. Long-term population trends may thus reflect a species capacity to respond to climate change. However, when a species is more abundant, it is also more easily detectable by observers, leading to an earlier detection of its arrival. Therefore, investigations of the drivers of shifts in apparent arrival dates to breeding sites and population trends remain challenging. Here, we formulate predictions aiming to disentangle the different drivers, analysing spring arrival dates and population changes of 52 migratory birds in a Central European country, the Czech Republic, from 1994 to 2017. If shifts in arrival dates are driven by increasing spring temperatures, migrants should arrive earlier in a warmer year and their abundance should increase in the subsequent year due to earlier breeding. If earlier migrant arrival results from their increased detectability caused by higher abundance, then migrants should arrive earlier in the same years when their abundance is high. We found clear support for the former prediction, indicating that climate change drives the earlier arrival of migrants irrespective of changes in their detectability. Moreover, species advancing their arrival to a greater degree had more positive population trends, and responses to rising spring temperatures in the Czech Republic became weaker with increasing migration distance. Therefore, climate change drives population trends of migratory species according to their capacity to adjust their arrival date to variations in spring temperatures.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
1 Introduction
Recent climate change has serious consequences for various taxa (reviewed by Parmesan and Yohe 2003; Parmesan 2006; Radchuk et al. 2019), including impacts on animal populations and phenology (Bellard et al. 2012; Thackeray et al. 2016; Pacifici et al. 2017; Cohen et al. 2018). In migratory birds, increases in average global temperatures have led to the advancement of their arrivals to breeding grounds (Gienapp et al. 2007; Lehikoinen and Sparks 2010; Pearce-Higgins et al. 2014). However, species migrating over longer distances that arrive later in the spring have advanced their arrivals less than species with shorter migration distances that arrive earlier (Lehikoinen et al. 2004; Rubolini et al. 2007; Miller-Rushing et al. 2008; Usui et al. 2017; but see Stervander et al. 2005; Jonzén et al. 2006; Zalakevicius et al. 2006). This is likely because the ability to optimise departures from more distant non-breeding areas is naturally lower and might be more tightly related to rigid endogenous controls or fuelling rates (Berthold 1996; Gwinner 1996; Lindström et al. 2019). Consequently, migrants that have not been able to sufficiently advance their spring migration and adjust their timing of breeding are more prone to population declines than those with more flexible arrivals (Møller et al. 2008; Salido et al. 2012).
Nevertheless, relationships between the timing of arrival and population changes are not as straightforward as the studies cited above indicate. Apparent bird arrivals to breeding grounds might be governed not only by climatic factors but also by species abundance. In years when a species is abundant, individuals are more easily detected by observers and its arrival might be thus detected earlier than in years with low abundance (Tryjanowski and Sparks 2001). As population abundance is driven by various factors independent of climate, e.g. land use change, hunting pressure and interspecific competition, shifts in apparent arrival date may be the consequence, rather than the cause of population abundance (Tryjanowski et al. 2005). To the best of our knowledge, we are not aware of any previous studies that aimed to disentangle these different drivers.
To fill this major knowledge gap, we investigated the two alternative mechanisms described above by formulating predictions enabling the assessment of their relative importance. If shifts in arrival dates are driven by increasing spring temperatures, migrants should arrive earlier in a warmer year. Consequently, their abundance should increase in the subsequent year due to earlier breeding, because the earlier-breeding birds have better opportunities to acquire higher-quality territories and to exploit food resources and, in the case of multiple clutches, raise more than one brood (Salido et al. 2012; Visser et al. 2012; Halupka and Halupka 2017). Therefore, the change in arrival date between two consecutive years (t–1 and t) should be positively related to the change in abundance between the next 2 years (i.e. t and t+1). In contrast, if the earlier arrival of migrants results from their increased detectability caused by higher abundance, then migrants should arrive earlier in the same years when their abundance is high. Therefore, the change in arrival date between two consecutive years (t–1 and t) should be positively related to the change in abundance between the same years (i.e. t–1 and t). Although these predictions are not mutually exclusive, they should provide important insights into which mechanism is dominant in a particular system.
In addition to analyses on an annual basis, we studied the consequences of shifts in migrant arrivals for their long-term population trends. In this respect, we predicted that the species with a more advanced spring arrival over the long term will have more positive population trends than species with smaller shifts in arrival dates. However, migrant arrival dates, abundances and their relationships can be affected by the distance over which the species migrate (Both et al. 2006; Møller et al. 2008; Jones and Cresswell 2010): (i) the longer the distance a given species migrates, the smaller should be the advancement in its arrival date; and (ii) increasing migration distance is associated with increasingly negative population trends, as species that migrate over longer distances will be less able to respond to climatic changes.
In this study, we test these relationships, controlling for the effects of various other traits known to affect bird populations using a unique long-term dataset containing both spring arrival dates (hereafter ‘arrival dates’) and population changes of 52 migratory bird species in a Central European country, the Czech Republic, collected over a 24-year period (1994–2017).
2 Material and methods
2.1 Arrival data
Bird phenology data were based on yearly mean first arrival dates (MFAD) of migratory bird species to the Czech Republic between 1994 and 2017. The data were mainly collected by hundreds of volunteers through a citizen science program called ‘Phenological observations’ organised by the Czech Society for Ornithology. The observers were instructed to record the first arrival dates at their focal sites. The bulk of these data was collected with the same observers over multiple years, and these were supplemented with additional long-term first arrival dates from additional several experienced ornithologists. In 2010, the recording of bird arrivals became part of a national portal for bird observations (Czech Society for Ornithology 2019). From 2014 onwards, we processed all MFAD from the national portal, but changes in the way of data collection over the study period were not accompanied by changes in methodology. We restricted the dataset to 52 common species where 1–191 (median = 33) records for each year were available, except for the Red-breasted Flycatcher (Ficedula parva), for which arrival data are lacking for 2000 and 2001 (Online Resource 1), thereby gathering a total of 48,535 observations of first arrival dates from hundreds of sites across the Czech Republic. From these local observations, we calculated the MFAD of each species for every year. All dates of observations are expressed as days of the year where January 1 = day 1, and leap years were taken into account. In the next step, we expressed the shift in MFAD between 1994 and 2017 for each species as the slope of the weighted regression of the MFAD versus years. In these regressions, we used the number of all arrival date observations of a given species in a given year as weights. This gave more weight to years with a better record of a given species. Finally, we calculated one grand mean first arrival date (GMFAD) for each species as the weighted mean of its annual MFADs, again using the number of observations of a given species in a given year as weights.
2.2 Population changes
Data on the abundance of individual species were collected within the Breeding Bird Monitoring Programme (BBMP) in the Czech Republic on an annual basis since 1982. BBMP is a large-scale generic monitoring scheme based on voluntary fieldwork using standardised point counts (for more details see e.g. Reif et al. 2010). For each species, we used its population index in a given year as a measure of its relative abundance (in %). The index value for the first year of the focal time series (1994) was set to 100%. Population indices were estimated using log-linear models with Poisson error distribution in TRIM software (Pannekoek and van Strien 2001), which takes serial correlation and overdispersion into account (see Jørgensen et al. 2016 for more details). To estimate the mean population growth rate (i.e. population trend) of each species from 1994 to 2017, we fitted a linear regression model across its annual population indices (%) weighted by 1/standard error of mean (SE). Each species population trend was expressed as the slope of this regression line.
2.3 Migration distance
Although previous studies have often sorted species according to their ‘migration strategy’ into groups of long- and short-distance migrants, we expressed migration distance as a continuous variable because the distinction between short- and long-distance migrants is not always clear and the division line between these groups is sometimes rather arbitrary (BirdLife International and NatureServe 2014; Cepák et al. 2008). We calculated the distance between the centroid of the Czech Republic (49.74 N, 15.34 E) and the centroid of a species non-breeding range using R (R Core Team 2017), packages ‘geosphere’, ‘maps’, ‘maptools’, ‘raster’ and ‘rgeos’ (Bivand and Lewin-Koh 2013; Hijmans 2013, 2014; Bivand and Rundel 2014), and based on maps of species non-breeding ranges provided by BirdLife International and NatureServe (2014).
2.4 Species traits
Arrival dates and population changes of birds might be affected by numerous factors whose influence can be assessed by focusing on species traits. Controlling for the effects of such traits enables investigations of the focal drivers not biased by confounding factors. We selected an assortment of 13 predictors from seven broader categories that included habitat, diet, climate niches, nest type and location, life history strategy, detectability and legal status. Most of these predictors have been successfully used in various studies (e.g. Böhning-Gaese and Oberrath 2003; Reif et al. 2011; Koleček et al. 2018). Below, we provide a short rationale and description of each of them. Unless otherwise stated, the measures were obtained from previous studies (Koleček and Reif 2011; Koleček et al. 2014, 2018), which extracted the data from Cramp (1977-1994), Hudec (1983, 1994) and Hudec and Šťastný (2005).
Habitat niche
Species breeding in open habitats, arid habitats and habitat specialists are known to have declined more than forest species, wetland species and habitat generalists (Reif 2013). Three measures of habitat niche were quantified. The first two were based on a classification of the habitat requirements of species along a gradient of decreasing vegetation structural complexity and density, from (1) closed forest to (7) open country without trees or shrubs. Species were allocated up to three different values, and their habitat niche position was calculated as the mean of these values. Habitat niche breadth was determined as the difference between the extreme values (i.e. the first and last occupied habitat; ranging between zero for species occupying a single habitat class to 6 for those utilising the whole range of habitat classes). We also calculated a third measure, habitat requirements along a gradient of increasing habitat humidity, from (1) dry (e.g. rocks, dry forests) and (2) humid (without open water surfaces—e.g. wet meadows and marshlands) to (3) aquatic habitats (with prevailing open water bodies—e.g. rivers and water reservoirs; for details, see Koleček et al. 2018).
Diet niche
Feeding on insects has recently been linked to population declines in contrast to species with herbivorous diet (Bowler et al. 2019). Diet niche position was based on a classification of species into four different trophic levels based on the dominant part of their diet, as either (1) herbivorous, (2) herbivorous and insectivorous, i.e. omnivorous, (3) insectivorous or (4) carnivorous. Diet niche breadth was based on a classification of species into (1) obligatory herbivorous (i.e. feed on plant sources only) or obligatory insectivorous/carnivorous (i.e. feed on animal sources only), (2) predominantly herbivorous with a minor proportion of insectivorous/carnivorous diet (or vice versa), or (3) omnivorous (i.e. plant, animal or other sources without clear preferences).
Climatic niche
Species breeding in colder climate and within a narrow range of climatic conditions have had more negative population trends than species breeding in warmer climate and climatic generalists (Jiguet et al. 2006; Stephens et al. 2016). We calculated climatic niche from maps of mean temperatures in the main 3-month species-specific breeding season (mostly April, May and June) for the period 1961–1990 based on temperature data obtained from Haylock et al. (2008), following Jiguet et al. (2007). The climatic niche position was the mean breeding season temperature over the breeding range of each species within Europe (taken from Hagemeijer and Blair 1997). Climatic niche breadth was classified as the difference between maximum and minimum temperatures across the European breeding range.
Nest type and location
Species breeding on the ground and in cavities are considered more prone to declines than species breeding higher above ground and in cavities (Gregory et al. 2007). To express the nest type, each species was scored along a gradient of three levels of increasing nest concealment: (1) open cup nests outside cavities, (2) semi-closed nests either outside cavities or open nests in semi-cavities and (3) cavity nests. Nest location described the height of the nest above the ground, from (1) on or close to the ground, (2) intermediate—i.e. shrubs and lower trees, to (3) high trees.
Life history strategy
Species with a slower pace of life (K-selected species) have been found to be declining less than species with a faster pace of life (r-selected species; Koleček et al. 2014). We chose six life history traits: body mass, egg mass, incubation period, clutch size, laying date (day of the year of the beginning of egg laying in the first breeding attempt) and number of broods per season, using mean values from published data (Cramp 1977-1994; Hudec 1983, 1994; Hudec and Šťastný 2005). As these six life history traits are highly correlated, we reduced their number by principal component analysis (PCA) into two independent axes (see also Koleček et al. 2014). Each species was positioned along the two most important ordination axes, and these scores were used for further analyses (Online Resource 2). We interpret the first ordination axis (PC 1, explaining 55.4% of the variability among species, eigenvalue = 3.33) as a gradient from ‘slower strategy’ (K-selected) species (i.e. those having larger eggs, larger body mass, longer incubation period and smaller clutch sizes) to ‘faster strategy’ (r-selected) species (hereafter the ‘first life history axis’). The second axis (PC 2, explaining 21.8% of the variability, eigenvalue = 1.31) expressed a gradient from species allocating most of their energy to just one breeding attempt per season (i.e. late breeding species having a single brood and larger clutch sizes) to species spreading their investments across multiple breeding attempts per season—i.e. early breeding species having multiple broods and smaller clutch sizes (hereafter the ‘second life history axis’). Each variable was log transformed before entering the PCA to improve normality.
Detectability
Species with a loud and well-known song, marked plumage colouration or behaviour might be observed earlier regardless of their abundance, temperature or other conditions (Tryjanowski and Sparks 2001). Therefore, we took the detectability of species at their spring arrival into account. Since obtaining reliable field measurements of this variable was hardly feasible, we used the expert classification of 22 skilled Czech ornithologists as a proxy. Each expert was asked to assign each species to one class of detection probability during the first days after arrival: 1, very low; 2, moderately low; 3, moderately high; 4, very high. From these classifications, we calculated the mean detectability of each species (see Online Resource 1), and these values entered the statistical analysis.
Legal protection
Species benefit from being listed as protected (Koleček et al. 2014; Koschová et al. 2018). We recognised four levels of increasing protection according to the protected species list of the Czech Republic (Anonymous 1992): species without any special legal protection (0), endangered (i.e. least protected; 1), highly endangered (2) and critically endangered species (i.e. most protected; 3). Please note that these terms do not describe the actual level of threat, but they reflect the labelling of official categories of legal protection used in the Czech Nature and Landscape Protection Act (Anonymous 1992).
2.5 Spring temperatures
Mean daily temperatures were calculated as average values from ten locations considered as representative for the Czech Republic (Czech Hydrometeorological Institute 2019). For further analyses relating MFAD to temperature for each species, we used mean daily temperatures over a period of 15 days before its MFAD in the focal year. We believe this length is a reasonable trade-off between selecting longer periods (such as 1 month) that might largely overlap some individuals’ stays in tropical regions (see e.g. Koleček et al. 2016) and shorter periods (such as several days) that ignore the conditions the individuals faced before arrival, which might be more important than the actual weather (e.g. Blackburn et al. 2019).
2.6 Data analyses
2.6.1 Analysis of annual variation
To identify predictors of species MFADs (Table 1), we defined a basic model relating a species MFAD in individual years (a response variable) to spring temperature before their arrival and their population indices in individual years as a linear mixed-effects model (LMM; R package lme4; Bates et al. 2015). The identity of species nested in genus and the latter nested in family nested in order (based on taxonomy taken from Gill and Donsker 2019) were included as a random effect to account for species taxonomy (see Lockwood et al. 2002; Jiguet et al. 2010), as recommended in comparative analyses across species (see e.g. Thomas 2008; Diniz-Filho et al. 2013). Before this analysis, we removed long-term trends in MFAD and population indices by detrending. For this purpose, we used residuals from the regressions of MFAD on year and population indices on year, respectively (see the “Arrival data” and “Population changes” sections and Table 1).
Responses of species MFADs to temperatures before arrival can vary with migration distances (temperature effects on arrival should be weaker in species migrating over longer distances). In the same way, the relationship between a species MFAD and abundance can be affected by the species detectability (Tryjanowski and Sparks 2001). Therefore, we developed two variants of the basic model. Firstly, we included a two-way interaction between temperature before arrival and migration distance into the basic model together with population indices. Secondly, we included a two-way interaction between a species population index and detectability into the basic model together with temperature before arrival. In both of these variants, we scaled and centred the explanatory variables to zero mean and unit variance, thereby allowing a correct interpretation of the interaction effects (Schielzeth 2010).
Finally, we expressed annual changes (where t signifies a given year, t–1 the previous year and t+1 the subsequent year) in MFAD and population indices, respectively, to perform further tests of possible causality in their relationships (Table 1). Firstly, we ran an LMM with population index(t+1)/population index(t) related to population index(t) and MFAD(t)/MFAD(t–1). Secondly, we ran an LMM with MFAD(t)/MFAD(t–1) related to population index(t)/population index(t–1). The significance of the first relationship would suggest an important statistical effect of the arrival date on bird populations, whereas the second relationship would support the opposite. In addition, both models contained species nested in genus nested in family and order as a random effect.
2.6.2 Analysis of long-term trends
We also assessed the relationship of a species population trend (response variable) with the shift in MFAD using an LMM, where 1/SE of the population trend was used as a weight. In this test, we accounted for GMFAD, migration distance, climatic niche position and breadth, habitat and diet niches, nest type, nest location, life history strategies (expressed as the two PC axes) and legal protection status. Genus nested in family and the latter nested in order were included as a random effect.
All variables would be ideally included into a single global linear model with a full set of explanatory variables on which the subsequent analysis was performed. However, migration distance was strongly collinear with PC 2 (rs = 0.75) and with GMFAD (rs = 0.77). Therefore, after the main global model that contained migration distance, we ran two additional global models consecutively replacing migration distance by the GMFAD and PC 2 to evaluate the effect of all variables. Using the Akaike information criterion corrected for small sample sizes (AICc; Burnham and Anderson 2002), we evaluated the performance of candidate models with particular combinations of explanatory variables from each global model set. With respect to our sample size (n = 52 species), we restricted the maximum number of parameters (k) in each candidate model to five, following the recommendation of n/k = 10 to prevent overfitting and to obtain conclusive inference (Burnham and Anderson 2002). We considered the best-supported models to be those with ΔAICc < 2. Model averaging was applied across all these models, taking model weight into account (Johnson and Omland 2004) in order to reveal parameter estimates for the explanatory variables (Bartoń 2016).
3 Results
3.1 Annual variation in MFAD
The basic model investigating the contributions of temperature before spring arrival and species abundance (controlling for detectability upon arrival) on arrival date showed that birds arrived earlier in years with higher temperatures (beta = − 0.298 ± 0.129 SE, Wald χ2 = 5.35, df = 1, P < 0.021), but not in years with higher abundance (beta = − 0.086 ± 0.121 SE, Wald χ2 = 0.50, df = 1, P = 0.477). Simultaneously, species detectability after arrival was not related to MFAD (beta = − 0.119 ± 0.128 SE, Wald χ2 = 0.86, df = 1, P = 0.354).
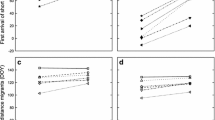
By introducing an interaction between temperature and migration distance into this basic model a striking result emerged—as temperatures increased from 1994 to 2017, birds migrating shorter distances advanced their arrival more than species that migrate longer distances (beta for the interaction = 0.562 ± 0.134 SE, Wald χ2 = 17.72, df = 1, P < 0.0001; Fig. 1). In contrast, including the interaction between species detectability and population indices into the basic model did not find any support for this interaction (beta = 0.156 ± 0.165 SE, Wald χ2 = 0.89, df = 1, P = 0.347).

The interactive effect of mean temperature at breeding grounds and migration distance on detrended mean first arrival dates (FADs) of species in individual years as assessed by a linear mixed-effects model (LMM). The regression lines illustrate the effect of mean temperature before spring arrival on detrended species mean FADs in individual years for different migration distances: green line = mean distance (4264 km), red line = − 1.5SD (944 km), blue line = + 1.5SD (7584 km)
The effect of arrival date on species abundance also emerged in the model evaluating the changes in abundance between consecutive years. Specifically, the negative change of population index in a subsequent year was related with the MFAD delayed relatively to the previous year (beta = − 0.328 ± 0.162 SE, Wald χ2 = 4.10, df = 1, P = 0.043). In contrast, the change in population index in a given year relative to the previous year was not related with the change of arrival date between these years (beta = 0.001 ± 0.005 SE, Wald χ2 = 0.07, df = 1, P = 0.796).
Finally, taxonomy explained only a minor proportion of the annual variation in all models (Online Resource 3).
3.2 Long-term population trends
In the analysis relating bird population trends to their predictors, 8 out of 2380 candidate models (i.e. models with possible combinations of up to five variables) met the ΔAICc < 2 threshold. The shift in MFAD and position on the slow/fast continuum (PC 1) were retained in all of the best models (Table 2), and these were the only variables related to the population trends (Table 3). As predicted, population trends were more negative in those species that had either delayed or not advanced their spring arrivals, while positive population trends were found in species with more advancing arrivals (Fig. 2). In addition, r-selected species had more negative population trends than K-selected species (Table 3). Except for orders, taxonomy explained only a marginal part of the variability in the eight top-ranked candidate models (Online Resource 3).

Relationship between population trends and shift in mean first arrival date (MFAD) of 52 migratory bird species in the Czech Republic, 1994–2017, based on linear regression (solid line). Dashed lines display the zero population trend (i.e. stable population) and no change in phenology
4 Discussion
We provide an important contribution to disentangling the complex relationships between spring temperature, arrival date and population trends of migratory birds with respect to migration distance, controlling for various ecological and life-history traits and phylogenetic relatedness. In respective years, arrival date was related to temperature (earlier arrival in warmer springs), but not to species abundance. The positive population change between two consecutive years followed an earlier arrival in the preceding year implying that the earlier arrival might have promoted more successful breeding which may translate into higher abundance in the subsequent year. Migration distance played a major role in the observed shifts of arrival dates: the shorter the distance, the larger was the arrival date advancement. Finally, species with more advanced spring arrivals had more positive population trends, whereas those species that did not advance arrivals declined.
The relationship between population trends and shifts in arrival date was in accord with previous studies reporting more positive trends in species advancing their arrivals to breeding grounds (Tryjanowski et al. 2005; Møller et al. 2008; Salido et al. 2012), as well as with our expectations. Here we provide deeper insights into this relationship. Theoretically, two different mechanisms are possible: a population increase (caused by any factor) results in a higher reporting rate of a species by observers, which means that there is a higher probability of reporting early individuals. This ultimately leads to the advancement of FAD (Tryjanowski et al. 2005). This relationship may be unrelated to any climatic effects on species arrival dates, and indeed corresponds to the equivocal support for climate change as a main driver of bird population changes found in several recent studies (Jones and Cresswell 2010; Currie and Venne 2017; Scridel et al. 2018).
The alternative (but not mutually exclusive) mechanism is based on climatically induced cascading impacts across trophic levels and has been more widely considered in the literature (Renner and Zohner 2018). Under this scenario, the advancement of arrival date is an evolutionary plastic response to a warming climate by which species track optimal conditions for breeding (Salido et al. 2012). An earlier arrival in a warmer spring enables migrants to start breeding earlier. This provides opportunities to better exploit resources available for breeding such as nesting sites and food (Halupka and Halupka 2017). Species relying on a peak of food supply can match the timing of breeding to the optimum food supply (Visser et al. 2012), and multi-brooded species can prolong the breeding season and have more breeding attempts (Halupka and Halupka 2017). Taken together, earlier arrival results in an eventual increase of breeding productivity and consequently of population size in the subsequent breeding season (Møller et al. 2010; Halupka and Halupka 2017). In contrast, species unable to advance their arrival date sufficiently may face the consequences of trophic mismatches or a reduced availability of nesting sites (Ahola et al. 2007; Samplonius and Both 2019), and their breeding productivity would be low (Both et al. 2006; Saino et al. 2011; but see Vatka et al. 2011). Thus, over the long-term, populations of more advancing species increase, while the least advancing species decline (Møller et al. 2008; Salido et al. 2012). Our results indicate clear support for the latter alternative, i.e. spring temperatures govern the timing of species arrival, and these shifts then translate into population changes. We infer this support from the fact that arrival date was tightly related to temperature, but not to species abundance in respective years. Moreover, when the species advanced their arrival compared with the previous year, their abundance increased in the following year, whereas the delayed arrival was accompanied by a subsequent population decline. In contrast, if arriving individuals did not follow the year-to-year increase in average spring temperatures and delay arrival relatively to the previous year, the probability of experiencing suboptimal temperature conditions and trophic mismatches would grow and may result in a decrease of productivity (Visser et al. 2006). Taken together, these findings indicate that an adjustment of arrival date is important for shaping migrant population sizes.
We found that the relationship between arrival date and temperature was dependent on the distance the species migrates. The shorter the distance, the earlier the arrival in years with higher temperature. We explain this variation by differences in the mechanisms responsible for controlling the migratory behaviour of birds. A shorter migratory route enables the species to respond rapidly to rising temperatures, which considerably advances the arrival probably due to the possibility of fine-tuned migration timing from nearby wintering sites (Møller et al. 2008). This ability decreases with the increasing length of the migratory route, when mechanisms of endogenous migration controls or the need to refuel at stopover sites possibly play a more important role (Berthold 1996; Gwinner 1996; Lindström et al. 2019). Although such differences in arrivals with respect to migration distance and their population consequences have been frequently studied (e.g. Gienapp et al. 2007; MacMynowski and Root 2007; Miller-Rushing et al. 2008), empirical evidence for the response of the timing of arrival to temperature in the interaction with migration distance is surprisingly lacking. As far as we know, this study is first to elucidate this pattern.
Previous studies have highlighted relationships between phenological and life-history traits (e.g. Jenni and Kéry 2003; Rubolini et al. 2007; Moussus et al. 2010). Namely, a pronounced advancement in arrivals has been documented in generalist species with broad climatic niches, great genetic variability and phenotypic plasticity (Hurlbert and Liang 2012), multiple broods and variable diets (Møller et al. 2008; Vegvari et al. 2010; Dunn and Møller 2014) compared with more specialised species. In contrast, evidence is lacking that shifts in arrivals are related to clutch size, laying date (Dunn and Møller 2014) or body mass and population size (Møller et al. 2008). Surprisingly, in our study, the life-history and ecological traits were not significantly related with the population phenology of species. This can be partly explained by our selection of traits, because we did not focus on genetic variability and phenotypic plasticity. Also, the broad scale of our study could have played a role. We used relatively crude measures of niche traits (i.e. habitat categories) that may have been too inaccurate to reflect subtle relationships with phenology, since these relationships were not the core focus of our study.
We show that r-selected species had less positive population trends than K-selected ones. These results are in line with relationships that have already been observed in the Czech Republic (Reif et al. 2010; Koleček et al. 2014), as well as in Western Europe (Jiguet et al. 2007). K-selected species may benefit from their ability to postpone reproduction longer until conditions are optimal, which may be crucial for long-term survival in suboptimal environments (Sol et al. 2012).
Apart from the correlative nature of our study, which was inevitable given its spatial and temporal extent, there are additional limitations to be aware of when interpreting the results. First, the collection of all data on bird abundance and arrival dates was based on the work of citizen scientists, which may vary in skills and effort. However, the identification skills of these voluntary observers are considerably high, since both the collection of phenological observations and the BBMP were organised by the Czech Society for Ornithology, and these programs did not allow the participation of unskilled people.
Second, the FAD is considered as a less biologically relevant estimator of arrival date than the mean arrival date (MAD), because the FAD is more prone to the effects of extreme events (Moussus et al. 2010). We acknowledge that this limitation may be present in our data, but MAD could not be collected within the citizen science programs we were working with. At the same time, we attempted to overcome the disadvantages of FAD through the spatiotemporal design of our study. While single-site FADs are known to be quite variable, this was not the case here because our dataset comprises records collected and averaged over multiple sites at a national level, and we control for the number of observations during the analysis. In addition, we deliberately focused on the MFAD as an early phenological indicator, because our targets are the early arriving individuals that are likely the bearers of change. In such cases, this indicator is preferable to other estimators (Moussus et al. 2010). Finally, the use of FAD alongside other measures of arrival does not have to lead to inconsistent results (Gienapp et al. 2007, but see Miller-Rushing et al. 2008), and Sparks et al. (2001) argued that FADs can be used cautiously to reveal the impacts of climate change on migration phenology, as we do here.
Third, regardless of the measure of arrival used, our data by its nature cannot reliably distinguish local Czech breeders and individuals on passage to northern breeding sites. However, we think that passage individuals likely represent a minor part of the sample. Recent individual-tracking studies have shown that there are distinct migration times among northern and southern breeding populations (e.g. Briedis et al. 2016; Gow et al. 2019). Birds from northern sites pass through central Europe later than local breeders, and thus, the majority of early arriving birds to Czechia must have been local breeders.
We have built on previous evidence relating population trends to phenology (e.g. Møller et al. 2008), shedding new light on these relationships. However, we still know little about the mechanisms linking species’ arrival to changes in their populations. The existence of a trophic mismatch, one of the key drivers in this framework, is often not supported by data on food abundance (e.g. Saino et al. 2011). As climate change is in progress, and may even accelerate in the forthcoming decades, further shifts in species phenology with profound population consequences are likely to be observed. From this perspective, comprehensive analyses simultaneously assessing bird phenology, food supply and population changes at various sites are highly desirable and will likely advance our understanding of animal responses to rapidly changing climatic conditions.
References
Ahola MP, Laaksonen T, Eeva T, Lehikoinen E (2007) Climate change can alter competitive relationships between resident and migratory birds. J Anim Ecol 76:1045–1052. https://doi.org/10.1111/j.1365-2656.2007.01294.x
Anonymous (1992) Decree of the Ministry of the Environment of 11 Jun 1992, implementing selected provisions of Czech National Council Act No. 114/1992 Coll. on the protection of the environment and the natural landscape
Bartoń K (2016) MuMIn: multi-model inference. R package ver. 1.15.6. https://CRAN.R-project.org/package=MuMIn. Accessed 1 June 2019
Bates DM, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Bellard C, Bertelsmeier C, Leadley P, Thuiller W, Courchamp F (2012) Impacts of climate change on the future of biodiversity. Ecol Lett 15:365–377. https://doi.org/10.1111/j.1461-0248.2011.01736.x
Berthold P (1996) Control of bird migration. Chapman and Hall, London
BirdLife International and NatureServe (2014) Bird species distribution maps of the world. BirdLife International and NatureServe
Bivand R, Lewin-Koh N (2013) maptools: tools for reading and handling spatial objects. R package ver. 0.8-27. http://CRAN.R-project.org/package=maptools. Accessed 1 June 2019
Bivand R, Rundel C (2014) rgeos: Interface to Geometry Engine – Open Source (GEOS). R package ver. 0.3–8. http://CRAN.R-project.org/package=rgeos. Accessed 1 June 2019
Blackburn E, Burgess M, Freeman B, Risely A, Izang A, Ivande S, Hewson C, Cresswell W (2019) Spring migration strategies of Whinchat Saxicola rubetra when successfully crossing potential barriers of the Sahara and the Mediterranean Sea. Ibis 161:131–146. https://doi.org/10.1111/ibi.12610
Böhning-Gaese K, Oberrath R (2003) Macroecology of habitat choice in long-distance migratory birds. Oecologia 137:296–303. https://doi.org/10.1007/s00442-003-1346-9
Both C, Bouwhuis S, Lessells CM, Visser ME (2006) Climate change and population declines in a long-distance migratory bird. Nature 441:81. https://doi.org/10.1038/nature04539
Bowler DE, Heldbjerg H, Fox AD, de Jong M, Böhning-Gaese K (2019) Long-term declines of European insectivorous bird populations and potential causes. Conserv Biol 33:1120–1130. https://doi.org/10.1111/cobi.13307
Briedis M, Hahn S, Gustafsson L, Henshaw I, Träff J, Král M, Adamík P (2016) Breeding latitude leads to different temporal but not spatial organization of the annual cycle in a long-distance migrant. J Avian Biol 47:743–748. https://doi.org/10.1111/jav.01002
Burnham KP, Anderson DM (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Cepák J, Formánek J, Hořák D, Jelínek M, Klvaňa P, Schröpfer L, Škopek J, Zárybnický J (2008) Czech and Slovak bird migration atlas. Aventinum, Prague
Cohen JM, Lajeunesse MJ, Rohr JR (2018) A global synthesis of animal phenological responses to climate change. Nat Clim Chang 8:224. https://doi.org/10.1038/s41558-018-0067-3
R Core Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing. https://www.R-project.org. Accessed 1 June 2019
Cramp SE (1977–1994) The birds of the Western Palearctic, vol I–IX. Oxford Univ. Press, Oxford
Currie DJ, Venne S (2017) Climate change is not a major driver of shifts in the geographical distributions of North American birds. Glob Ecol Biogeogr 26:333–346. https://doi.org/10.1111/geb.12538
Czech Hydrometeorological Institute (2019) Territorial air temperature. http://portal.chmi.cz/historicka-data/pocasi/denni-data. Accessed 1 June 2019
Czech Society for Ornithology (2019) Faunistic database of the Czech Society for Ornithology. http://birds.cz/avif. Accessed 1 June 2019
Diniz-Filho JAF, Loyola RD, Raia P, Mooers AO, Bini LM (2013) Darwinian shortfalls in biodiversity conservation. Trends Ecol Evol 28:689–695. https://doi.org/10.1016/j.tree.2013.09.003
Dunn PO, Møller AP (2014) Changes in breeding phenology and population size of birds. J Anim Ecol 83:729–739. https://doi.org/10.1111/1365-2656.12162
Gienapp P, Leimu R, Merilä J (2007) Responses to climate change in avian migration time—microevolution versus phenotypic plasticity. Clim Res 35:25–35. https://doi.org/10.3354/cr00712
Gill F, Donsker D (2019) IOC World Bird List (v9.1). https://doi.org/10.14344/IOC.ML.9.1
Gow EA, Burke L, Winkler DW, Knight SM, Bradley DW, Clark RG, Bélisle M, Berzins LL, Blake T, Bridge ES, Dawson RD, Dunn PO, Garant D, Holroyd G, Horn AG, Hussell DJT, Lansdorp O, Laughlin AJ, Leonard ML, Pelletier F, Shutler D, Siefferman L, Taylor CM, Trefry H, Vleck CM, Vleck D, Whittingham LA, Norris DR (2019) A range-wide domino effect and resetting of the annual cycle in a migratory songbird. Proc R Soc Lond B Biol Sci 286:20181916. https://doi.org/10.1098/rspb.2018.1916
Gregory RD, Vorisek P, Van Strien A, Gmelig Meyling AW, Jiguet F, Fornasari L, Reif J, Chylarecki P, Burfield IJ (2007). Population trends of widespread woodland birds in Europe. Ibis 149:78-97. https://doi.org/10.1111/j.1474-919X.2007.00698.x
Gwinner E (1996) Circadian and circannual programmes in avian migration. J Exp Biol 199:39–48
Hagemeijer WJM, Blair MJ (1997) The EBCC atlas of European breeding birds. Their distribution and abundance. T and AD Poyser, London
Halupka L, Halupka K (2017) The effect of climate change on the duration of avian breeding seasons: a meta-analysis. Proc R Soc Lond B Biol Sci 284:20171710. https://doi.org/10.1098/rspb.2017.1710
Haylock MR, Hofstra N, Klein Tank AMG, Klok EJ, Jones PD, New M (2008) A European daily high-resolution gridded dataset of surface temperature and precipitation. J Geophys Res Atmos. https://doi.org/10.1029/2008JD010201
Hijmans RJ (2013) raster: geographic data analysis and modeling. R package ver. 2.1–66. http://CRAN.R-project.org/package=raster. Accessed 1 June 2019
Hijmans RJ (2014) geosphere: spherical trigonometry. R package ver. 1.3–11. http://CRAN.R-project.org/package=geosphere. Accessed 1 June 2019
Hudec K (1983) Fauna of Czechoslovakia: birds III. Academia, Prague
Hudec K (1994) Fauna of the Czech and Slovak Republic: birds I. Academia, Prague
Hudec K, Šťastný K (2005) Fauna of the Czech Republic: birds II. Academia, Prague
Hurlbert AH, Liang Z (2012) Spatiotemporal variation in avian migration phenology: citizen science reveals effects of climate change. PLoS One 7:e31662. https://doi.org/10.1371/journal.pone.0031662
Jenni L, Kéry M (2003) Timing of autumn bird migration under climate change: advances in long-distance migrants, delays in short-distance migrants. Proc R Soc Lond B Biol Sci 270:1467–1471. https://doi.org/10.1098/rspb.2003.2394
Jiguet F, Julliard R, Thomas CD, Dehorter O, Newson SE, Couvet D (2006) Thermal range predicts bird population resilience to extreme high temperatures. Ecol Lett 9:1321–1330. https://doi.org/10.1111/j.1461-0248.2006.00986.x
Jiguet F, Gadot AS, Julliard R, Newson SE, Couvet D (2007) Climate envelope, life history traits and the resilience of birds facing global change. Glob Chang Biol 13:1672–1684. https://doi.org/10.1111/j.1365-2486.2007.01386.x
Jiguet F, Devictor V, Ottvall R, Van Turnhout C, Van der Jeugd H, Lindström Å (2010) Bird population trends are linearly affected by climate change along species thermal ranges. Proc R Soc Lond B Biol Sci 277:3601–3608. https://doi.org/10.1098/rspb.2010.0796
Johnson JB, Omland KS (2004) Model selection in ecology and evolution. Trends Ecol Evol 19:101–108. https://doi.org/10.1016/j.tree.2003.10.013
Jones T, Cresswell W (2010) The phenology mismatch hypothesis: are declines of migrant birds linked to uneven global climate change? J Anim Ecol 79:98–108. https://doi.org/10.1111/j.1365-2656.2009.01610.x
Jonzén N, Lindén A, Ergon T, Knudsen E, Vik OJ, Rubolini D, Piacentini D, Brinch C, Spina F, Karlsson L, Stervander M, Andersson A, Waldenström J, Lehikoinen A, Edvardsen E, Solvang R, Stenseth NC (2006) Rapid advance of spring arrival dates in long-distance migratory birds. Science 312:1959–1961. https://doi.org/10.1126/science.1126119
Jørgensen PS, Böhning-Gaese K, Thorup K, Tottrup AP, Chylarecki P, Jiguet F, Lehikoinen A, Noble DG, Reif J, Schmid H, van Turnhout C, Burfield IJ, Foppen R, Voříšek P, van Strien A, Gregory RD, Rahbek K (2016) Continent-scale global change attribution in European birds – combining annual and decadal time scales. Glob Chang Biol 22:530–534. https://doi.org/10.1111/gcb.13097
Koleček J, Reif J (2011) Differences between the predictors of abundance, trend and distribution as three measures of avian population change. Acta Ornithol 46:143–153. https://doi.org/10.3161/000164511X625919
Koleček J, Schleuning M, Burfield IJ, Báldi A, Böhning-Gaese K, Devictor V, Fernández-García JM, Hořák D, Van Turnhout CAM, Hnatyna O, Reif J (2014) Birds protected by national legislation show improved population trends in Eastern Europe. Biol Conserv 172:109–116. https://doi.org/10.1016/j.biocon.2014.02.029
Koleček J, Procházka P, El-Arabany N, Tarka M, Ilieva M, Hahn S, Honza M, de la Puente J, Bermejo A, Gürsoy A, Bensch S, Zehtindjiev P, Hasselquist D, Hansson B (2016) Cross-continental migratory connectivity and spatiotemporal migratory patterns in the great reed warbler. J Avian Biol 47:756–767. https://doi.org/10.1111/jav.00929
Koleček J, Procházka P, Ieronymidou C, Burfield IJ, Reif J (2018) Non-breeding range size predicts the magnitude of population trends in trans-Saharan migratory passerine birds. Oikos 127:599–606. https://doi.org/10.1111/oik.04549
Koschová M, Rivas-Salvador J, Reif J (2018) Continent-wide test of the efficiency of the European Union’s conservation legislation in delivering population benefits for bird species. Ecol Indic 85:563–569. https://doi.org/10.1016/j.ecolind.2017.11.019
Lehikoinen E, Sparks T (2010) Changes in migration. In: Møller AP, Fiedler W, Berthold P (eds) Effects of climate change on birds. Oxford Univ. Press, Oxford, pp 89–112
Lehikoinen E, Sparks TH, Zalakevicius M (2004) Arrival and departure dates. Adv Ecol Res 35:1–31. https://doi.org/10.1016/S0065-2504(04)35001-4
Lindström Å, Alerstam T, Hedenström A (2019) Faster fuelling is the key to faster migration. Nat Clim Chang 9:288. https://doi.org/10.1038/s41558-019-0443-7
Lockwood JL, Russell GJ, Gittleman JL, Daehler CC, McKinney ML, Purvis A (2002) A metric for analysing taxonomic patterns of extinction risk. Conserv Biol 16:1137–1142. https://doi.org/10.1046/j.1523-1739.2002.01152.x
MacMynowski DP, Root TL (2007) Climate and the complexity of migratory phenology: sexes, migratory distance, and arrival distributions. Int J Biometeorol 51:361–373. https://doi.org/10.1007/s00484-006-0084-1
Miller-Rushing AJ, Lloyd-Evans TL, Primack RB, Satzinger P (2008) Bird migration times, climate change, and changing population sizes. Glob Chang Biol 14:1959–1972. https://doi.org/10.1111/j.1365-2486.2008.01619.x
Møller AP, Rubolini D, Lehikoinen E (2008) Populations of migratory bird species that did not show a phenological response to climate change are declining. Proc Natl Acad Sci U S A 105:16195–16200. https://doi.org/10.1073/pnas.0803825105
Møller AP, Flensted-Jensen E, Klarborg K, Mardal W, Nielsen JT (2010) Climate change affects the duration of the reproductive season in birds. J Anim Ecol 79:777–784. https://doi.org/10.1111/j.1365-2656.2010.01677.x
Moussus JP, Julliard R, Jiguet F (2010) Featuring 10 phenological estimators using simulated data. Methods Ecol Evol 1:140–150. https://doi.org/10.1111/j.2041-210X.2010.00020.x
Pacifici M, Visconti P, Butchart SH, Watson JE, Cassola FM, Rondinini C (2017) Species’ traits influenced their response to recent climate change. Nat Clim Chang 7:205. https://doi.org/10.1038/nclimate3223
Pannekoek J, van Strien A (2001) TRIM 3 manual. http://www.ebcc.info/art-13.
Parmesan C (2006) Ecological and evolutionary responses to recent climate change. Annu Rev Ecol Evol Syst 37:637–669. https://doi.org/10.1146/annurev.ecolsys.37.091305.110100
Parmesan C, Yohe G (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37. https://doi.org/10.1038/nature01286
Pearce-Higgins JW, Green RE, Green R (2014) Birds and climate change: impacts and conservation responses. Cambridge Univ. Press, Cambridge
Radchuk V, Reed T, Teplitsky C, van de Pol M, Charmantier A, Hassall C, Adamík P, Adriaensen F, Ahola MP, Arcese P, Avilés JM, Balbontin J, Berg KS, Borras A, Burthe S, Clobert J, Dehnhard N, de Lope F, Dhondt AA, Dingemanse NJ, Doi H, Eeva T, Fickel J, Filella I, Fossøy F, Goodenough AE, Hall SJG, Hansson B, Harris M, Hasselquist D, Hickler T, Joshi J, Kharouba H, Martínez JG, Mihoub J-B, Mills JA, Molina-Morales M, Moksnes A, Ozgul A, Parejo D, Pilard P, Poisbleau M, Rousset F, Rödel M-O, Scott D, Senar JC, Stefanescu C, Stokke BG, Kusano T, Tarka M, Tarwater CE, Thonicke K, Thorley J, Wilting A, Tryjanowski P, Merilä J, Sheldon BC, Møller AP, Matthysen E, Janzen F, Dobson FS, Visser ME, Beissinger SR, Courtiol A, Kramer-Schadt S (2019) Adaptive responses of animals to climate change are most likely insufficient. Nat Commun 10:3109. https://doi.org/10.1038/s41467-019-10924-4
Reif J (2013) Long-term trends in bird populations: a review of patterns and potential drivers in North America and Europe. Acta Ornithol 48:1–16. https://doi.org/10.3161/000164513X669955
Reif J, Vermouzek Z, Voříšek P, Šťastný K, Bejček V, Flousek J (2010) Population changes in Czech passerines are predicted by their life-history and ecological traits. Ibis 152:610–621. https://doi.org/10.1111/j.1474-919X.2010.01036.x
Reif J, Böhning-Gaese K, Flade M, Schwarz J, Schwager M (2011) Population trends of birds across the iron curtain: brain matters. Biol Conserv 144:2524–2533. https://doi.org/10.1016/j.biocon.2011.07.009
Renner SS, Zohner CM (2018) Climate change and phenological mismatch in trophic interactions among plants, insects, and vertebrates. Annu Rev Ecol Evol Syst 49:165–182. https://doi.org/10.1146/annurev-ecolsys-110617-062535
Rubolini D, Møller AP, Rainio K, Lehikoinen E (2007) Intraspecific consistency and geographic variability in temporal trends of spring migration phenology among European bird species. Clim Res 35:135–146. https://doi.org/10.3354/cr00720
Saino N, Ambrosini R, Rubolini D, von Hardenberg J, Provenzale A, Hüppop K, Hüppop O, Lehikoinen A, Lehikoinen E, Rainio K, Romano M, Sokolov L (2011) Climate warming, ecological mismatch at arrival and population decline in migratory birds. Proc R Soc Lond B Biol Sci 278:835–842. https://doi.org/10.1098/rspb.2010.1778
Salido L, Purse BV, Marrs R, Chamberlain DE, Shultz S (2012) Flexibility in phenology and habitat use act as buffers to long-term population declines in UK passerines. Ecography 35:604–613. https://doi.org/10.1111/j.1600-0587.2011.06797.x
Samplonius JM, Both C (2019) Climate change may affect fatal competition between two bird species. Curr Biol 29:327–331. https://doi.org/10.1016/j.cub.2018.11.063
Schielzeth H (2010) Simple means to improve the interpretability of regression coefficients. Methods Ecol Evol 1:103–113. https://doi.org/10.1111/j.2041-210X.2010.00012.x
Scridel D, Brambilla M, Martin K, Lehikoinen A, Iemma A, Matteo A, Jähnig S, Caprio E, Bogliani G, Pedrini P, Rolando A, Arlettaz R, Chamberlain D (2018) A review and meta-analysis of the effects of climate change on Holarctic mountain and upland bird populations. Ibis 160:489–515. https://doi.org/10.1111/ibi.12585
Sol D, Maspons J, Vall-llosera M, Bartomeus I, García-Peña GE, Piñol J, Freckleton RP (2012) Unraveling the life history of successful invaders. Science 337:580–583. https://doi.org/10.1126/science.1221523
Sparks TH, Roberts DR, Crick HQP (2001) What is the value of first arrival dates of spring migrants in phenology? Avian Ecol Behav 7:75–85
Stephens PA, Mason LR, Green RE, Gregory RD, Sauer JR, Alison J, Aunins A, Brotons L, Butchart SHM, Campedelli T, Chodkiewicz T, Chylarecki P, Crowe O, Elts J, Escandell V, Foppen RPB, Heldbjerg H, Herrando S, Husby M, Jiguet F, Lehikoinen A, Lindström Å, Noble DG, Paquet J-Y, Reif J, Sattler T, Szép T, Teufelbauer N, Trautmann S, van Strien AJ, van Turnhout CAM, Vorisek P, Willis SG (2016) Consistent response of bird populations to climate change on two continents. Science 352:84–87. https://doi.org/10.1126/science.aac4858
Stervander M, Lindström Å, Jonzén N, Andersson A (2005) Timing of spring migration in birds: long-term trends, North Atlantic Oscillation and the significance of different migration routes. J Avian Biol 36:210–221. https://doi.org/10.1111/j.0908-8857.2005.03360.x
Thackeray SJ, Henrys PA, Hemming D, Bell JR, Botham MS, Burthe S, Helaouet P, Johns DG, Jones ID, Leech DI, Mackay EB, Massimino D, Atkinson S, Bacon PJ, Brereton TM, Carvalho L, Clutton-Brock TH, Duck C, Edwards M, Elliott JM, Hall SJG, Harrington R, Pearce-Higgins JW, Høye TT, Kruuk LEB, Pemberton KM, Sparks TH, Thompson PM, White I, Winfield IJ, Wanless S (2016) Phenological sensitivity to climate across taxa and trophic levels. Nature 535:241. https://doi.org/10.1038/nature18608
Thomas GH (2008) Phylogenetic distributions of British birds of conservation concern. Proc R Soc Lond B Biol Sci 275:2077–2083. https://doi.org/10.1098/rspb.2008.0549
Tryjanowski PS, Sparks TH (2001) Is the detection of the first arrival date of migrating birds influenced by population size? A case study of the red-backed shrike Lanius collurio. Int J Biometeorol 45:217–219. https://doi.org/10.1007/s00484-001-0112-0
Tryjanowski P, Kuźniak S, Sparks TH (2005) What affects the magnitude of change in first arrival dates of migrant birds? J Ornithol 146:200–205. https://doi.org/10.1007/s10336-005-0079-4
Usui T, Butchart SH, Phillimore AB (2017) Temporal shifts and temperature sensitivity of avian spring migratory phenology: a phylogenetic metaanalysis. J Anim Ecol 86:250–261. https://doi.org/10.1111/1365-2656.12612
Vatka E, Orell M, Rytkönen S (2011) Warming climate advances breeding and improves synchrony of food demand and food availability in a boreal passerine. Glob Chang Biol 17:3002–3009. https://doi.org/10.1111/j.1365-2486.2011.02430.x
Vegvari Z, Bokony V, Barta Z, Kovacs G (2010) Life history predicts advancement of avian spring migration in response to climate change. Glob Chang Biol 16:1–11. https://doi.org/10.1111/j.1365-2486.2009.01876.x
Visser ME, Holleman LJM, Gienapp P (2006) Shifts in caterpillar biomass phenology due to climate change and its impact on the breeding biology of an insectivorous bird. Oecologia 147:164–172. https://doi.org/10.1007/s00442-005-0299-6
Visser ME, te Marvelde L, Lof ME (2012) Adaptive phenological mismatches of birds and their food in a warming world. J Ornithol 153:75–84. https://doi.org/10.1007/s10336-011-0770-6
Zalakevicius M, Bartkeviciene G, Raudonikis L, Janulaitis J (2006) Spring arrival response to climate change in birds: a case study from Eastern Europe. J Ornithol 147:326–343. https://doi.org/10.1007/s10336-005-0016-6
Acknowledgements
We thank P. Lumpe for curating the arrival data in the initial period and hundreds of collaborators for gathering data on FAD. We also thank the hundreds of voluntary ornithologists who monitor bird abundance within BBMP. Z. Vermouzek curated data in the BBMP database and part of the database on arrival dates. V. Beran, V. Brlík, J. Cepák, J. Chytil, M. Fejfar, J. Flousek, D. Křenek, V. Kubelka, R. Lučan, P. Lumpe, M. Paclík, P. Procházka, M. Pudil, P. Řepa, O. Sedláček, M. E. Šálek, K. Ševčíková, J. Šírek and P. Voříšek kindly provided their ranking of species’ detectability on arrival. We are grateful to three anonymous reviewers for their constructive comments. D. W. Hardekopf improved the English.
Funding
This study was supported through the Charles University (PRIMUS/17/SCI/16), the Institutional Research Plan (RVO: 68081766), Palacký University grant scheme (IGA PrF 2018_016, 2019_019) and the Czech Science Foundation (20-00648S).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOCX 51 kb)
Rights and permissions
About this article

Cite this article
Koleček, J., Adamík, P. & Reif, J. Shifts in migration phenology under climate change: temperature vs. abundance effects in birds. Climatic Change 159, 177–194 (2020). https://doi.org/10.1007/s10584-020-02668-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10584-020-02668-8