
Abstract
The flavonoid naringenin and a family of naringenin derivative Cu(II) complexes having phenanthroline-based second ligands were selected to study alkaline phosphatase activation. This enzyme plays a critical role in tissue formation, increasing the inorganic phosphate formation, favoring mineralization, and being essential to producing bone mineralization. The effects of those compounds on the function and structure of the enzyme were evaluated by kinetic measurements, fluorescence, FTIR, and UV-Vis spectroscopies. The results showed that naringenin did not affect alkaline phosphatase activity, having a value of the Michaelis–Menten-constant close to the enzyme (Km = 3.07 × 10–6). The binary complex, Cu(II)-naringenin, and the ternary complex Cu(II)-naringenin-phenanthroline behaved as an enzyme activator in all the concentrations range used in this study. Those complexes increased in c.a. 1.9% the catalytic efficiency concerning enzyme and naringenin. The ternary complex Cu(II)-naringenin-bathophenanthroline, provokes an activator mixed effect, dependent on the substrate concentrations. The different kinetic behavior can be correlated with different conformational changes observed under the interaction with ALP. Fluorescence experiments showed a raising of the binding constant with temperature. FTIR determinations showed that the complex with bathophenanthroline modifies the ALP structure but maintains the helical structure. The other copper complexes provoked a structural unfolding, decreasing the α-helix content. None of them affect the dephosphorylation enzyme ability. Even though the interactions and structural modifications on ALP are different, it is evident that the presence of copper favors enzymatic activity. The observed electrostatic interactions probably benefit the dissociation of the bound phosphate. The results suggest potential biological applications for the studied compounds.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Biological processes like phosphorylation and dephosphorylation are part of the metabolic control systems. These essential reactions are catalyzed by phosphatase enzymes through hydrolytic chemical reactions involving a phosphate ester residue. Phosphatases can be classified as (i) low and high molecular weight acid phosphatases containing histidine or lysine in the active site, (ii) protein phosphatase with cysteine in the active site, (iii) purple acid phosphatase (metal-binuclear active center), and (iv) alkaline phosphatase (ALP) with serine in the active site (McLauchlan et al. 2015). In this work, the interest is focused on this last enzyme whose name is related to the pH involved during the enzymatic process. Mammalian alkaline phosphatases are Zn-containing enzymes encoded by a multigene family, forming a large family of dimeric enzymes usually located at the cell surface. Those structures have been completely elucidated. Each monomeric unit has an active site (Zalatan et al. 2008). The site involved the coordination of two Zn(II) ions bridged by the phosphate substrate and close to the nucleophilic residue of Ser 102 and an Mg(II) ion in the vicinity of the active center. They are necessary for the enzymatic activity, the conformation of the ALP monomer, and the control of subunit interactions (Sharma et al. 2014
This enzyme is associated with many physiological functions, and its level is taken as a routine diagnosis. The tissue-nonspecific isozyme of ALP is found strongly expressed in the cells of mineralized tissue participating in the formation of hard tissue (Vimalraj 2020). It is known that ALP increment inorganic phosphate concentration facilitating the mineralization process and lowering the presence of pyrophosphate ions which act as an inhibitor of the formation of minerals. Two main steps accompanied the mineralization process. The first is the formation of hydroxyapatite crystals in the matrix-vesicles, and their distribution to the extracellular matrix through the membrane. The second is the expansion into the extracellular area that needs adequate calcium and inorganic phosphate concentration to produce again more hydroxyapatite crystals. In this step ALP acts via the pyrophosphate hydrolysis to favor the necessary inorganic phosphate production (Vimalraj 2020). Thus, this enzyme is vital because the fully differentiated osteoblasts are distinguished by ALP coexpression and collagen type I, both crucial for the production of bone matrix and mineralization event.
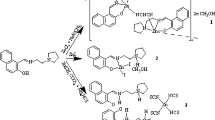
On the other side, several studies demonstrated great potency of flavonoids on the regulation of bone metabolism and in the reduction on bone loss (Ramesh et al. 2021; Bellavia et al 2021). Apigenin inhibited osteoblast (OB) and osteoclast (OC) differentiation and reduced bilateral ovariectomy (OVX) related to the bone loss. Formononetin increment bone regeneration of femoral drill hole injury in OVX mouse. Hesperidin induced OB differentiation. Baicalein reduced OC differentiation and promotes bone healing. Quercetin had a stimulatory effect on ALP and, like rutin, induced OB differentiation. Particularly, naringenin demonstrated to possess osteogenic effects. In bone marrow mesenchymal stem cells (BMSCs), upregulated the gene and expression levels of ALP, the transcription factor RUNX2, the C-X-C chemokine receptor type 4 (CXCR4), and stromal cell-derived factor 1 (SDF-1) via the SDF-1/CXCR4 signaling pathway (Ramesh et al. 2021; Bellavia et al 2021; Wang et al. 2021). Thus, flavonoids complement osteoblastogenesis and osteoclastogenesis assisting bone formation and avoiding bone resorption. In addition, flavonoids metal complexes have been also proved as promotors of bone formation (Vimalraj et al 2021, 2020, 2019; Vimalraj et al 2018a, b; Preeth et al. 2021; Rajalakshmi et al. 2018; Ferrer et al. 2006). Among several type of studies, ALP level was evaluated for all of them. Results showed remarkable up-regulation in ALP activity which is in all the treatment dose-dependent. This common behavior prompt us to study the effect on ALP activity by a family of copper(II) naringenin complexes. Previously, we studied the anticancer activity of those compounds on lung cancer cells (Caro-Ramírez et al. 2022). Here, we presented a kinetic profile of naringenin, Cu-naringenin (CuNar), Cu-naringenin-phenanthroline (CuNarPhen) and Cu-naringenin-bathophenanthroline (CuNarBatho) complexes (Fig. 1) analyzing phosphatase activity. In addition, structural modifications of the enzyme were assessed by UV-Vis, fluorescence and Raman spectroscopies.

Materials and methods
Reagents and instrumentation
Naringenin (95%), phenanthroline (99%), and bathophenanthroline (97%) were purchased from Sigma. Copper(II) chloride dihydrate (97%) from Anedra. Alkaline Phosphatase (ALP, EC 3.1.3.1) from bovine intestinal mucosa (Sigma Aldrich), lyophilized powder,10 DEA units/mg solid. For the determination of ALP activity, p-Nitrophenyl phosphate (p-NPP) bis(tris) salt 97% (Sigma Aldrich) was used. The binary CuNar complex ([CuNar2(H2O)2]) was prepared according to the method reported by Celiz et al (2019). CuNarPhen ([Cu(H2O)2(Nar)(phen)]Cl) was prepared according to Tamayo et al (2016) using copper(II) chloride dihydrate instead of Copper(II) perchlorate, and CuNarBatho ([Cu(Nar)(Batho)Cl]2.6H2O) according to our previous wok (Caro-Ramírez et al. 2022). Brief descriptions of their preparations are given in the supplementary material. All other chemicals used were of analytical grade.
UV-Vis determinations were recorded with a Shimadzu 260 0/270 0 spectrophotometer. Fluorescence spectra were obtained using a Shimadzu (RF60 0 0) luminescence spectrometer equipped with a pulsed xenon lamp.
Phosphatase activity
The assay was performed at 37 ºC and pH 10.4 in the incubation buffer (55 mM glycine + 0.55 mM MgSO4). The conversion of the substrate p-nitrophenyl phosphate (p-NPP) to p-nitrophenol (p-PNP) was monitored by the absorbance changes at 405 nm (ε = 18,500 M−1 cm−1). The experimental conditions for ALP-specific activity measurement were bovine intestinal ALP (7.96 µM) and p-NPP (with final concentrations ranging from 0 to 3.25 mM). The spectral data were recorded at 1-seg intervals for 3600 s. At first, a mixture of a solution of ALP enzyme (7.96 µM) and the studied compounds at 75 µM concentrations of naringenin, CuNar, CuNarPhen, and CuNarBatho were pre-incubated for 5 min at 37 °C. After that, the enzymatic reaction was started with the addition of the substrate solution. The effect of each concentration was repeated five times in three different experiments (Martini et al. 2021).
For the percentage calculation of the enzyme activity a control sample without the compounds was taken as 100%. It is worth mentioning that the percentage values for the reported metal-flavonoid complexes mentioned at the results and discussion section were estimated based on the graphs shown on the references papers (see below).
Evaluation of the catalytic parameters
A treatment based on the Michaelis–Menten model for enzyme kinetics studies was performed. The values of the Michaelis binding constant (Km), maximum velocity (Vmax), and rate constant for dissociation of substrates (i.e., turnover frequency, kcat) were calculated from the Lineweaver–Burk graph (double reciprocal plot) of 1/V versus 1/[S], using the Eq. 1/V = (Km /Vmax) (1/[S]) + 1/Vmax. Control reactions were assayed checking no changes in the absorbance at 405 nm. The data represent determinations in triplicate and were performed at least five times.
Alkaline phosphatase interaction
Fluorescence and UV–Vis spectroscopy
Alkaline phosphatase interaction experiments were performed by UV-Vis, and fluorescence spectroscopy at the same experimental conditions. For naringenin (Nar) and CuNar complex, the enzyme concentration was set at 6 µM and the solutions of the compounds were varied in the 0–75 µM range. The assay was performed at 298, 303, and 310 K. The exciting wavelength was set at 280 nm and recorded in the range of 310–400 nm.
For the CuNarPhen and CuNarBatho complexes, the fluorescence experiments were determined using a fixed complex concentration (75 µM) and varying ALP (0, 2, 4, 6, 8, and 10 µM). Fluorescence intensity was measured at 360 nm for CuNarPhen and 380 nm for CuNarBatho. The same excitation wavelength was used in all experiments.
The inner-filter effect was considered for the calculations, and the fluorescence intensities were corrected using the Eq. (1):
where Fcorr and Fobs are the fluorescence intensities corrected by inner-filter effect and recorded, respectively, Aex and Aem are the electronic absorbances of the solutions at excitation and emission wavelengths, respectively. For Nar and CuNar corrections, the UV-Vis spectra at each concentration of the compounds were used. For the corrections of CuNarPhen and CuNarBatho, the UV-Vis spectra of ALP were assessed.
The fluorescence quenching data were analyzed by the Stern–Volmer procedure involving the following Eq. (2):
where Fo is the steady-state fluorescence intensity of ALP alone while F is the observed intensity upon increasing the quencher concentration, Ksv is the Stern–Volmer quenching constant, and [Q] is the quencher concentration. Based on this equation the type of the fluorescence process can be established. If the curve Fo/F vs [Q] is linear, the type of quenching involves a unique process: static or dynamic. Static quenching is expected for a complex formation between the fluorophore and the quencher. It can be distinguished from collision effects when the Ksv value results higher than the value of the dynamic quenching constant (Kq). Kq could be calculated as the quotient between Ksv and the average lifetime of the biomolecule without quencher is assumed as τo = 10–8 s (Liu et al. 2012) being Kq = Ksv/τo. Then, this constant can be compared with the maximum diffusion collision quenching rate constant (reference value from the literature is 2 × 10–10 M−1 s−1). When Ksv value results higher than Kq, a mechanism of interaction involving the complex formation can be proposed. If not, it would be a collisional quenching (Roy et al. 2010).
When the quenching is static, specific binding sites are assumed. These binding sites and their association constants are estimated using the following mathematical relationship:
where Kb is the binding constant, and n is the average number of binding-site per protein molecule.
Thermodynamic parameters allowed to assess the type of interaction. Changes in Entropy (∆S), enthalpy (∆H), and Gibbs free energy (∆G), were calculated by the following equations:
where T (in K) is the experimental temperature, Kb is the binding constant at corresponding T, and R is the gas constant (8.314 J mol−1 K−1) (Roy et al. 2010).
FTIR spectroscopy
To improve signal-to-noise ratio in FTIR spectra, Tris–HCl buffer, ALP solution, naringenin-ALP solution, and copper(II) complexes solutions (ALP (6 µM), ALP(6 µM)-Nar(75 µM) and ALP(6 µM)-copper(II) compounds (75 µM)) were lyophilized. The vibrational spectra of proteins solutions and the freeze-dried powders are similar. The freeze-drying process does not affect protein conformation (Tu 1982). The FTIR spectra of the freeze-dried powdered samples were assessed with a Bruker IFS 66 FT-IR-spectrophotometer from 4000 to 400 cm−1 in the form of pressed KBr pellets. The buffer spectra were collected under the same experimental conditions and then its absorbance was subtracted from the spectra of the sample to get the FTIR spectra of the modified enzyme. The studied region involved the amide I band and some amino acids side chain contributions (1700–1535 cm−1).
The frequencies, the number of peaks to be fitted, and the half-width of each peak.to start a least squares iterative curve-fitting procedure were those obtained from the second derivative of the original spectra. The areas of the bands were calculated by integration of the corresponding fitted band. The curve-fitting process was performed by step-by-step iterative procedure towards a minimum root mean-square error of the different parameters determining the shape and position of the absorption peaks. It was effectuated by assuming an initial mixed Lorentzian-Gaussian line-shape function, with a full width at half height (FWHH) of 8–15 cm−1 and a maximum resolution factor. Baseline corrections, normalization, derivation, curve fitting and area calculation were carried out by means of Grams/32 (Galactic Industries Corporation, USA) software, OPUS 3.1, and PerkinElmer software. Amide I band components were analyzed according to the following assignments: β-sheets, 1637–1613 cm−1; solvated helix, 1625–1637 cm−1; random coil, 1645–1637 cm−1; α-helix, 1658–1650 cm−1; turns, 1673–1666 cm−1 and β-antiparallel, 1695–1675 cm−1. For the side chains contributions were considered Tyr-O− 1599–1602 cm−1; TyrOH 1594–1602 cm−1, Gln δNH2 1586–1610 cm−1, Asp νCOO− 1574–1579 cm−1, Glu νCOO− 1556–1560 cm−1. To calculate the percentage contribution of the different types of conformations to the area of all the components, bands assigned to a given conformation were summed and divided by the total area corresponding to each region (Barth 2000, 2007; Ferrer et al. 2008).
Results and discussion
Effect of Nar and Nar-metal complexes on alkaline phosphatase activity (ALP)
It was previously mentioned the role of flavonoids in bone metabolism. More interesting is the enhancement of pharmacological effects of their metal-flavonoid complexes in this newly studied area. The group of Vimalraj et al. have been researching the impacts of some copper(II) and zinc(II) complexes. A family of Cu(II)-quercetin complexes demonstrated osteogenic and angiogenic properties (Vimalraj et al. 2018a, b). Cu(II)-silibinin complexes behaved as promotors of osteoblast differentiation and calcium deposition (Rajalakshmi et al. 2018). Zn(II)-silibinin regulated miR 590/Smad7 signaling pathway and osteoblast differentiation (Vimalraj et al. 2018a, b). Zn(II)-Morin incremented Runx2, type 1-collagen, mRNAs expression and promoted osteocalcin and osteonectin proteins (Vimalraj et al. 2019). Zn(II)-kaempferol promoted bone growth (Vimalraj et al. 2020). Zn(II)-rutin incremented the osteoblast differentiation marker genes (Vimalraj et al. 2021). There was a common factor in all those studies, alkaline phosphatase was studied as a marker for osteoblastic differentiation and in all the studies ALP is activated in presence of the metal complexes. In view of these results, to understand the nature of ALP activation, the kinetic behavior of ALP with a series of Cu-naringenin (CuNar), Cu-naringenin-phenanthroline (CuNarPhen), Cu-naringenin-bathophenanthroline (CuNarBatho) complexes, and for naringenin (Nar), were evaluated. A comparison and analysis of the effects of these compounds on enzyme activity were assessed. To deal with those studies the enzyme (6 µM) was incubated with those compounds at pH = 10.4. The p-NPP concentrations were varied from 0.5 to 3.25 µM. For the majority of the compounds, the same concentrations were used. But, for some of them, concentrations that allowed kinetic differentiation were selected. For these measurements, the effect of Cu(II)-naringenin complexes and naringenin amount was evaluated using a concentration range from 25 to 100 µM. The previous experiments suggested that 75 µM concentration is adequate for the kinetic experiments. At that condition, a first order reaction was determined at lower substrate concentrations (linear plot), and, at higher concentrations, it became saturated. Fig. S1 (A) shows the graph of absorbance vs time for ALP in the absence of the compounds, while Fig. S1 (B, C, D, and E) correspond to the activities of ALP in the presence of Nar, CuNar, CuNarPhen, and CuNarBatho, respectively. Fig. S1(F) shows the comparison with the curves of Abs vs time for all the compounds and ALP alone at 0.75 µM of p-NPP concentration value. It can be observed that naringenin does not affect to a great extent the Abs vs time curve up to 20 min. Immediately after, the Nar curve increases its absorbance value. In the linear zone, the curve for CuNar behaves similarly to CuNarPhen up to the 7 min, after that CuNar decays and a significant difference can be observed. On the contrary, CuNarPhen clearly shows a significant increment in the product amount. None of them show a decrease in the number of products. To compare the effect on ALP activity at this experimental condition, the percentage of the enzyme activity for the selected compounds at different time intervals is shown in Fig. 2. The ALP activity without the complexes was considered 100%. It can be observed that at 5, 10, and 15 min of the kinetic reaction, there was no significant difference between ALP alone or in the presence of Nar. Similar increment percentages (~ 50%) were observed for the CuNar and CuNarPhen at 5 min. Note that, at the 10 and 15 min, the difference in the increment is 59% and 73% for CuNar and 89% and 126% for CuNarPhen, respectively, with respect to ALP alone. Remarkably different is the behavior in presence of CuNarBatho. The activity of the enzyme increased with time. This complex enhanced enzyme activity in 152%, 365% and 389 at 5, 10, and 15 min, respectively. The results indicate that under our experimental conditions, this complex was more effective than the others in the stimulation of ALP. As it was mentioned, many studies conducted in literature emphasized that flavonoids and its metal complexes were able to enhance ALP activity (Vimalraj et al 2021, 2020; Vimalraj et al 2018a, b; Preeth et al. 2021; Rajalakshmi et al. 2018; Ferrer et al. 2006). In previous studies we demonstrated that a magnesium complex ([Mg(DMA)2(H2O)4]Cl2.2H2O, 5 mM) was able to stimulate ALP in a similar experimental conditions of this work addressing 53% of increment in PNP concentration (Martini et al. 2021).

Percentage of activity of ALP in the presence of Nar (75 µM), CuNar (75 µM), CuNarPhen (75 µM) and CuNarBatho (75 µM) at different time intervals (5, 10, and 15 min). Errors bars are expressed as x ± SD
Unfortunately, data for other metal-flavonoids complexes were not in the same experimental conditions, but the assay was based on p-nitrophenyl phosphate substrate similar to this work. In that case, the following comments can be drawn. ALP determinations at day seven for rutin (10 µM) and Zn(II)-rutin (10 µM) complex in human dental pulp stem cells (hDPSCs) and immortalized human osteoblast MG-63 cells afforded approximately 35% and 70% of increment in the ALP activity (Vimalraj et al. 2021).
For Kaempferol-zinc(II) complex and kaempferol alone, three days of incubation at 25 µM on MG-63 cells gave an increase of 100% and 80%, respectively, showing up-regulation on ALP activity (Vimalraj et al 2020). As it was revealed, Rajalakshmi et al. showed the enhancement of ALP activity when studied a series of zinc and copper-silibinin complexes. Particularly the ternaries [Zn(sil)(phen)] and [Cu(sil)(phen)] augmented 42% and 150% the enzyme activity after up to 7 days of incubation on MG-33 cells with a 60 µM of concentration of the complexes (Vimalraj et al. 2018a, b; Rajalakshmi et al. 2018). With similar objective, zinc and copper quercetin complexes have been proved and compared with the control quercetin. It was shown that [Zn(quer)(H2O)] was not as effective as [Zn(quer)(phenanthroline)] which after seven days of treatment in the MG-63 cells with 60 µM of concentration reached at 142% of activity enhancement (Preeth et al. 2021). Copper quercetin was also tested at the same concentration value and cells lines as the zinc complexes. For a series, Cu(II)-quercetin, Copper-Quercetin-phenanthroline, and Copper-Quercetin-neocuproine, the presence of phenanthroline increased ALP activity by about 100% (Vimalraj et al. 2018a, b). In the same trend to evaluate osteogenic potential, ALP activity assessment of Morin and Morin zinc (60 μM, MG-63 cells lines, 7 days) showed stimulation being most potent the [Zn(Morin)2] complex with an 73% of increment in enzyme activity (Vimalraj et al. 2019). Even so, the experimental conditions used in those research works were not the same of the present investigation, resulting evident that the coordination of phenanthroline to copper(II) ion caused greater effect than the binary flavonoid metal complexes, in a similar trend observed for our results. In view of the results, it can be suggested that the presence of the methyl groups in the 2 and 9 position of the phenanthroline ring (neocuproine) did not favor ALP stimulation. On the contrary, as it was observed in this work, the presence of phenyl groups at the 4 and 7 positions of bathophenanthroline molecule have a promissory effect on the ALP improvement activity.
To understand the nature of ALP activation, an analysis of the kinetic experiments was performed. The reactions follow a typical Michaelis–Menten behavior (Fig. S1). The Lineweaver–Burk graphs were obtained by plotting 1/V versus 1/[p-NPP] (Fig. 3). According to this model, the kinetic parameters Km, Kcat, and Vmax were calculated to characterize the effect of the naringenin and the complexes on enzymatic reaction.

A Lineweaver–Burk plots for the kinetic experiments ALP ( ) in absence and in presence of Nar (
) in absence and in presence of Nar ( 75 µM), CuNar (
75 µM), CuNar ( 75 µM), CuNarPhen (
75 µM), CuNarPhen ( 75 µM) and CuNarBatho (
75 µM) and CuNarBatho ( 75 µM). Substrate concentrations in the 0–3.25 mM range. B Expanded region
75 µM). Substrate concentrations in the 0–3.25 mM range. B Expanded region
As shown in Table 1, the Michaelis-binding constants decrease with the presence of bathophenanthroline in the copper complex, but there is no significant change for the other compounds. Since Km is indicative of the enzyme affinity towards the substrate, a lower value suggested a stronger association. Therefore, the results showed that CuNarBatho is the complex with the highest affinity. On the other side, the increment in Vmax, and the catalytic constant kcat are significant for the CuNarBatho complex being the values fivefold higher than that of the enzyme without the complex. It is known that the specificity constant kcat/Km is frequently used as a measure of catalytic performance. So, the comparison of the catalytic efficiency, shows that the best catalytic power is displayed by the copper complex having Batho in the coordination sphere around the metal center.
It can also be inferred that the presence of Cu(II) improved catalytic efficiency in comparison with naringenin. CuNar complex presented an octahedral geometry formed by two Nar molecules coordinating through deprotonate 5-alkoxide (C5-OH, A ring) and 4-keto (C = O, C ring) groups and two water molecules completing the coordination sphere CuO4(Ow2) (Caro-Ramírez et al. 2022). Phenanthroline derivative complexes are predominantly monomeric species in solution with an equatorial CuO2N2 environment in which the coordination of Nar is assumed to be the same that in the binary complex (Caro-Ramírez et al. 2022; Celiz et al. 2019; Tamayo et al. 2016). Thus, it can be deduced that there is a marked influence of the co-ligand environment. N-donor ligands influence is evidenced in some catalytic experiments in which this type of ligand enhances the rate of reactions and in other pharmacological activities such as anticancer one (Masuri et al. 2022). The presence of Batho ligand remarkable changes the kinetic parameters and the type of activation mechanism. This result probably may be explained by the chemical nature of the substituents on the phenanthroline core. In bathophenanthroline, there is a more extended π-conjugated system and π-resonance communication in addition to the σ induction (Ferreira et al. 2017).Nevertheless, more experiments are needed to confirm this hypothesis.
Consistent with the general theory for enzyme activation or inhibition of Saboury (2009), a moderator could change the rate of an enzymatic reaction. When the effect of the moderator is the reduction of the reaction rate, the process is described as inhibition. Conversely, when the rate of reaction increases, the process is called activation. For the activation process, the following possibilities are considered: (i) the moderator activates the enzyme when the Km value decreases and the Vmax value increases (either at high substrate concentrations or in the entire concentration range), (ii) the moderator is called a non-essential activator when it activates the enzyme by increasing the catalytic constant without any effect on the binding affinity of the substrate to the enzyme, or when it decreases the binding affinity of the substrate to the enzyme without affecting the catalytic constant.
Based on this theory, the observed behavior of Nar and their metal complexes was discussed. The type of activation can be suggested by analyzing the position of the intersection of the Lineweaver–Burk graphs in the presence or absence of an activator (Saboury 2009). The behavior of Nar resulted similar to that of the enzyme at least at the concentration range used in this study (Fig. 3; Table 1). There are no significant differences between Km, Vmax, kcat, and the catalytic efficiency values for ALP and ALP-Nar systems. The ALP-CuNar and ALP-CuNarPhen systems behaved quite similar. There is a notorious increment in Vmax and in the catalytic constant, but Km values have no significant variation concerning ALP, so it can be considered that CuNar and CuNarPhen behaved like a non-essential activator. It means that the moderator activated the enzyme incrementing the catalytic constant without any effect on Km. Very different results were obtained for the CuNarBatho complex. It seems to be a mixed effect. When the ratio of 1/[p-NPP] is greater than 1, it behaved like the other copper complexes. But at 1/[p-NPP] ratios less than 1 there is decrease of the Km value (Fig. 3; Table 1) implying higher substrate affinity to ALP and a remarkable increment in the value of the Vmax.
Fluorescence spectroscopy measurements-Mechanisms of action
Enzyme activity is dependent on structural integrity (Das et al. 2018, Salehian and Shareghi 2019, Liu et al. 2012). Some conformational modifications result in moderation, activation, or inhibition of its biological activity. Hence, fluorescence spectroscopy was used to examine the interaction of Nar and Nar-metal complexes looking for conformational modifications. It is known that the aromatic residues tryptophan, tyrosine, and phenylalanine are the origin of the intrinsic fluorescence in proteins. ALP is a homodimeric enzyme having four tryptophan residues in a polypeptide chain that are the main fluorescence emitters. Any change in their surrounding environment due to structural protein modifications has consequences for the fluorophores-properties. Therefore, the endogenous protein fluorescence can be quenched by compounds. This quenching process can be applied to analyze the conformational protein changes. Native ALP showed a fluorescence-maximum emission at 332 nm (Figs. 4A, S2A). At fixed ALP concentration (6 µM), the fluorescence spectra were gradually quenched by the increasing Nar concentrations (0–75 µM). In addition, a red-shift in the maximum wavelength of the band is observed, and it moves to 342 nm. This change is observable at the three selected temperatures. The fluorescence behavior of CuNar is similar but with a more pronounced quenching (Figs. 4B, S2B). The band moved towards 351 nm. It is suggested that the concomitant progressive reduction of the fluorescence intensity and the bathochromic shift in the maximum emission wavelength are caused for an increment of the polarity in the ALP environment compared to the control (Das et al. 2018, Sarkar and Ghosh 1996). As result of the conformational changes, Trp residues are more exposed to a polar environment. Clearly, the interaction with the binary metal complex causes a 75% decrease in the intensity of the ALP fluorescence band against 55% caused by naringenin.

Fluorescence emission spectra of ALP (6 µM) and ALP (6 µM) with incrementing concentrations in the 0, 12.5, 25, 37.5, 50, 62.5 and 75 µM range at 298 K of A Nar and B CuNar (λexc = 280 nm, λem = 310–400 nm)
The partial structural loss was more evident in CuNar. Neither Nar nor CuNar causes a loss of conformational stability since the activity of ALP was not modified for Nar and it was activated by CuNar. It is possible that both compounds stick to the ALP molecule and provoke rearrangements around the tryptophan environment that does not inhibit ALP activity. In addition, the fact that both Nar and CuNar quench the Trp-fluorescence, but Nar does not affect enzyme activity while CuNar behaves as an activator suggest that the fluorophores do not directly interact with the active site.
The quenching experiments for the phenanthroline-derived complexes were very different. It is acknowledged that some compounds can enhance their emission after protein interaction or interference. Fig. S3A and S3B show this phenomenon for CuNarPhen and CuNarBatho complexes. Protein quenching is also revealed. It can be seen that at 75 µM of complex concentrations a complete quenching process occurs. The superposition of the bands affected the measurements. In those cases, it is convenient to use the compound emission instead ALP emission to study the interaction (Mukherjee et al. 2013; Zhou et al. 2019). Thus, the experiment was performed using a fixed complex concentration of 75 µM and varying ALP concentrations from 0 to 10 µM. The recorded fluorescence spectra are shown in Fig. 5.

Fluorescence emission spectra of A CuNarPhen and B CuNarBatho at 75 µM with incrementing concentrations of ALP from 0, 2, 4, 6, 8, and 10 µM range at 310 K (λexc = 280 nm, λem = 330–420 nm)
For both complexes, the fluorescence intensity decreases with the successive addition of ALP suggesting protein-metal complex interaction. The quenching effect of ALP on the complex with phenanthroline is not as stronger as that with bathophenanthroline. The maximum effect is observed at 10 µM ALP concentration with a reduction of no more than 30.2% of the initial fluorescence value at 310 K. It can also be noted that the typical fluorescence bands of the complex CuNarPhen do not modify their maximum emission values which remain constant at 360 nm and 375 nm (Figs. 5A, S4A). Conversely, the interaction is stronger at the three temperatures for CuNarBatho complex (Figs. 5B, S4B). At 10 µM of ALP concentration the quenching effect is about 85.5% and the maximum of the compound emission is shifted towards blue moving the fluorescence band of CuNarBatho at 374 nm. Thus, a different type of protein-compound binding can be inferred, according to the dissimilar kinetic behavior.
Binding parameters
The possible quenching mechanisms of the protein can be static, dynamic, or a combination of both. Static refers to the formation of a ground state complex. The dynamic is caused by a collisional effect between the quencher and protein. They can be distinguished analyzing the temperature dependence of the Stern–Volmer constant. When the quenching constants increase with temperature, there is an increase in the collisional effect. On the contrary, there is a lowering in the constant value for the static quenching process. Based on this concept, the Stern–Volmer graphs were analyzed after the inner-filter effect corrections were performed (Das et al. 2018, Salehian and Shareghi 2019). Figure 6 shows the results obtained for Nar and CuNar complexes.

Stern–Volmer plots and log[(Fo-F)/F] vs log[Q] graphs for the fluorescence data of Nar and CuNar in the 0, 12.5, 25, 37.5, 50, 62.5 and 75 µM concentrations range at (•) 298 K, (■)303 K, and (▲) 310 K. [ALP] = 6 µM, λexc = 280 nm, λem = 310–400 nm
As it is shown, for the three temperatures, a linearity in the curves was obtained. From those graphs, the Stern–Volmer constants were evaluated. The quenching rate constants, Kq, were also calculated considering the equation Ksv = Kq x τo and taking the fluorescence biopolymer lifetime value of 10–8 s (Table 2) (Liu et al. 2012). The Kq values, are larger than the known value of 2 × 10–10 M−1 s−1 for the bimolecular quenching constant in an aqueous medium (Liu et al. 2012). This result suggests that the collisional quenching mechanism can be discarded. On the other side, the values obtained for the Ksv are close and fall into the experimental error suggesting that the affinity of the compounds for the enzyme does not change substantially with the increase of temperature (Roy et al. 2012; Naso et al 2016).
From the observed changes, it is assumed that Nar and CuNar can interact with ALP. The binding constant Kb and the number of binding sites (n) were evaluated. The Kb values suggest magnitude of the distribution and interaction of a biomolecule. Strong binding implies strong interaction and attachment. According to the proposed formula, graphs of log[(Fo-F)/F] vs log[Q] at different temperatures were plotted (Fig. 6). The calculated values are disclosed in Table 3. Both compounds presented a binding site of ~ 1, which imply the presence of one binding site for a molecule.
The binding-constant values are in the expected order for the binding of bioactive molecules, 104–106 M−1. For Nar, the Kb values have little change with temperature increment. For CuNar, the Kb values resulted higher by metal complexation, and there is a slight increment of the binding constant with the temperature. This tendency may explain the different kinetic behavior of Nar in comparison with CuNar.
As stated above, the interaction of the phenanthroline-derived complexes was studied measuring their ALP quenching. With the same methodology, the binding constants were calculated (Fig. 7; Table 3).

Stern–Volmer plots and log[(Fo-F)/F] vs log[Q] graphs for the fluorescence data of A CuNarPhen (75 µM) and B CuNarBatho (75 µM) with incrementing concentrations of ALP from 0, 2, 4, 6, 8, and 10 µM range at (•) 298 K, (■)303 K, and (▲) 310 K. λexc = 280 nm, λem = 310–400 nm
An increase in the binding constant with the temperature is observed. This increment is more notorious for the CuNarBatho complex. The Kb value at 310 K is at least forty times greater than the one obtained at 298 K. A higher constant value implies strong binding and less free compound (Cacita and Nikolaou 2016). This Kb tendency has been found in other studies with flavonoid-albumin interactions like naringin, baicalin, and their metal complexes (Roy et al. 2010; Masuri et al. 2022; Xiao et al. 2007). The large Kb increments could be associated with the strong quenching effect and the blueshift of the fluorescence band of the CuNarBatho complex.
Analysis of thermodynamic parameters and binding forces
To examine the interaction forces occurring during the interaction between ALP and the compounds, the thermodynamic parameters were analyzed. Ross and Subramanian (1981) proposed a conceptual model of protein association based on the magnitude and sign of the thermodynamic parameters. In this model, the main force is hydrophobic attraction when ΔH > 0 and ΔS > 0, the hydrogen bonds and van der Waals are relevant when ΔH < 0 and ΔS < 0, and electrostatic interactions prevail when ΔH < 0 or displays a very low positive value and ΔS > 0. Table 3 showed the data obtained for the studied systems. Naringenin and CuNar produce electrostatic forces together with hydrophobic interactions. The slight positive ΔH values and the positive ΔS support this assumption together with the proposed rule that a stable association cannot be based on hydrophobic interactions alone. The same interaction of naringenin albumin was found (Bi et al. 2012). Assuming that naringenin at pH = 10.4 possesses the 4´-OH (ring B) group deprotonated (pK2 = 9.1) (Farajtabar and Gharib 2013), the negative charge may be responsible of the ionic type of interactions. In both compounds, the ΔG values are negative suggesting spontaneous interaction and entropically driven association. The phenanthroline-derived compounds (Table 3) showed a remarkable and thermodynamically favorable process with higher negative ΔG values. Both parameters, ΔH negative and a positive ΔS also suggested electrostatic and hydrophobic interactions. Possibly, the presence of the bulky substituents (phenanthroline and bathophenanthroline) contributes to the addition of extra hydrophobic interactions and favoring the association process. The binding interaction resulted spontaneous, exothermically and enthalpy favored.
UV–Vis spectroscopy
The interaction with ALP also can be visualized by UV–Vis spectroscopy. The ALP concentrations resulted in accord with the fluorescence experiments. Figure 8(A,B) displays the UV–Vis spectra of Nar (A) and CuNar (B) (75 µM) without (solid line) and with (dotted of line) ALP (6 µM). In Nar, the presence of the enzyme lowers ~ 11% the absorbance value of the band at 321 nm and produces hypsochromic shift of the 233 nm band. In CuNar, the interaction causes a splitting of the band at 321 nm to 308 nm and 330 nm (shoulder). For the 234 nm band an additional shoulder appeared at 230 nm. In both compounds the band at 242 nm modify the intensity. Figure 8C, D show the UV–Vis spectra for copper complexes at 75 µM without (solid line) and with ALP (10 µM, dotted line). It can be observed remarkable and distinctive changes in both of them.

Normalized UV absorption spectra of A Nar and B CuNar (75 µM) without (solid line, red) and with ALP 6 µM (dotted line, blue) and C CuNarPhen and D CuNarBatho (75 µM), without (solid line, red) and with ALP (10 µM, dotted line, blue). The ALP concentrations are in accord with the fluorescence experiments. (Color figure online)
After ALP addition, the UV–Vis of CuNarPhen lowers the intensities of the bands. The broad band with maximum peaks located at 238, 248, and 263 nm diminished the absorbance value by 11.8%, 15.3%, and 33.1%, respectively, and changed the whole shape. The peak at 319 nm lowers 7% of its intensity. On the contrary, the spectrum of CuNarBatho increases its intensity. The band at 319 nm of the metal complex becomes broadly with a maximum at 303 nm influenced by the enzyme interaction.
The most intense band in the 280–200 nm region belongs to the electronic transitions of the A and C rings of the naringenin moiety. The low intensity band at ~ 320 nm is assigned to the π → π* electronic transitions over the A and C rings. Other bands are related to the formation of copper complex or the presence of the second ligand (phenanthroline or bathophenanthroline) (Caro-Ramírez et al. 2022; Tamayo et al. 2016). Those bands are clearly modified in presence of ALP.
The modifications and shift of the characteristic bands of Nar and of the metal complexes, and the fact that the absorption spectra difference (Nar-Nar/ALP, CuNar-CuNar/ALP, CuNarPhen-CuNarPhen/ALP and CuNarBatho-CuNarBatho/ALP) resulted different from those of Nar or their copper complexes (Das et al. 2018), provide additional evidence for enzyme-interaction. The UV-Vis spectra are not supposed to change in the case of dynamical quenching in contrast to the case of static quenching (Das et al. 2018). Therefore, it is possible to assume that the UV-Vis results are in concordance with the fluorescence measurements, because of i) the observed modifications and shifts of the characteristic bands of Nar and the metal complexes and ii) the fact that the absorption spectra difference (Nar-Nar/ALP, CuNar-CuNar/ALP, CuNarPhen-CuNarPhen/ALP and CuNarBatho-CuNarBatho/ALP) resulted different from those of Nar or their copper complexes.
Similar modifications were observed for quercetin-copper(II)-albumin systems (Roy et al. 2012). A high content of hydrophobic interactions is proposed. These interactions may arise from an energy gap between the π → π* levels leading to the observed blue shifts or the bands splitting.
FTIR spectroscopy
Infrared spectroscopy is a great tool to investigate protein structures. Amide I band is a characteristic studied band including all the contributions as α-helix, β-sheet, solvate helix, turn, and random coil structures. This band centered between 1650–1658 cm−1 comes from the νC = O stretching vibrational mode and other contributions in a minor proportion of the CCN deformation, out-of-phase CN stretching vibration, and the NH in-plane bend. This band is strongly influenced when the protein interacts with different compounds that provoke conformational changes over the native form. Thus, the position and the shape are sensitive to the conformation of the protein (Tu 1985, Barth 2000, 2007; Ferrer et al. 2008). To test whether the interaction with the studied set of compounds caused the mentioned changes, an analysis of the amide I band, and some bands connected with the side chain amino acids were performed. Fig. S5 and Table 4 displays the obtained results for the quantification procedure.
The results obtained for the control sample (ALP, 6 µM, pH = 10.4) were in concordance with those previously obtained (Table 4) (Parente et al. 2019). Upon incubation of the compounds with ALP, considerable changes in the α -helix structure were observed, except for the CuNarBatho complex. The percentage of α-helix structure showed a significant decrease after binding with Nar, CuNar, and CuNarPhen. The changes are consistent with structural perturbations leading to the partial unfolding of α-helical domains into looser conformation. Although Nar did not affect the kinetic behavior of ALP, the structural changes in ALP support the interaction of the ligand with the enzyme. For this compound, the lowering of the helical contents is concomitant with the increment in the percentage of the solvated helix (69.2%) and turns structures (only 4%). It is known that changes of the protonation state of the side chains are relevant for catalytic mechanisms (Barth and Zscherp 2002). Therefore, the percentage of those contributions can modify the activity. For Nar, there is an increment in the contribution of the deprotonate Tyr residues along with an increase in the percentage of the carboxylate contribution associated with Asp and Glu residues.
The modifications caused by the addition of CuNar show different changes in different conformational components. The unfolding of the ALP is evident. There is a decrease in the corresponding value of α-helix (-70%) while increasing trends were observed for the percentage of β-antiparallel, solvated helix, and turns. Concerning the side chains absorptions there is a notable increment of the contribution of the aspartate residue. This observed change is consistent with the ionic interactions deduced from the thermodynamic parameters. Nar and CuNar denote approximately similar changes, but the differences in the percentages of the affected residues may explain the differences in the kinetic behavior. The aspartate side chains seem to be more involved in the interaction of CuNar with ALP. Upon addition of CuNarPhen, the quantitative analysis of the secondary structure of ALP reveals the lowering of the α-helix structure and solvated helix going to turns and β-sheet increase and the appearance of some random coil component. Like the others, there are changes in the side chain components like tyrosine and aspartate residues but, differently, glutamine residues appear to be exposed. The presence of positive charged NH3 residues (pK = 13) together the higher β sheet contents (39.3%) compared with the ALP control may explain the difference in the degree of interaction. In comparison with Nar and the other copper(II) complexes, the presence of CuNarBatho does not modify the percentage of the α-helix structure, and the changes are produced mainly in the turns and the β-antiparallel components. This compound also evidences the exposition of Tyr-O− and aspartate residues.
The observed modifications were different for all the compounds. Some points can be addressed considering the aforementioned.
-
(i)
Since the enzymatic activity was not modified and, in some cases, it was increased, it is possible to suggest that the conformational changes did not affect the active site of the enzyme (Bortolato et al. 2002). This assumption is supported by the observed side chain perturbations and the fluorescence experiments.
-
(ii)
The interaction with ALP seems quite different for the most active copper complex, CuNarBatho. The structural modifications do not involve the helical structure. Considering the high Kb and the low Km values, and the calculated catalytic efficiency, it can be concluded that this side chain attachment favors substrate affinity.
-
(iii)
Despite CuNar and CuNarPhen showed a similar kinetic activity, their attachment to ALP seems to be different. The presence of phenanthroline forces ALP to unfold with a higher β-sheet contents and exposes the side chain-charged components. This change is evidenced by the Kb constant, which is also higher at the optimum enzyme temperature.
Conclusions
In this study, we investigated the effect of the family of copper-naringenin complexes on alkaline phosphatase activity and compared those with naringenin. Different patterns of reactivity emerge when the kinetic activities were evaluated. Naringenin did not affect enzyme activity. The copper complexes behaved as an enzyme activator. The chemical structures of the complexes may explain the different interactions. The fluorescence quenching measurements of ALP in presence of naringenin and CuNar complex suggested that triptophan residues are exposed to a polar environment. The fact that Nar does not affect the enzymatic activity while CuNar behaves as an activator suggests that the fluorophores are not at the active site. The quenching measurements of the ternary complexes showed binding constant values higher than those of the binary complex and naringenin. All the compounds showed a thermodynamically favorable process including electrostatic and hydrophobic interactions being higher for the phenanthroline derivatives. The FTIR results showed structural ALP modifications. CuNarBatho complex did not affect a great extent the α-helix content. Nar and CuNar showed similar secondary structural modifications, but CuNar showed a higher lowering of α-helix contents and a higher increment of β-antiparallel and aspartate side chains. In summary, copper naringenin derivatives could be suitable compounds to activate ALP. Further studies of the involvement of bone tissue engineering applications are needed to determine the potential uses of these compounds.
References
Barth A (2000) The infrared absorption of amino acid side chains. Prog Biophys Mol Biol 74:141–173. https://doi.org/10.1016/S0079-6107(00)00021-3
Barth A (2007) Infrared spectroscopy of proteins Biochim. Biophys Acta 1767:1073–1101. https://doi.org/10.1016/j.bbabio.2007.06.004
Barth A, Zscherp C (2002) What vibrations tell us about proteins. Q Rev Biophys 35:369–430. https://doi.org/10.1017/S0033583502003815
Bellavia D, Dimarco E, Costa V, Carina V, De Luca A, Raimondi L, Giavaresi G (2021) Flavonoids in bone erosive diseases: perspectives in osteoporosis treatment. Trends Endocrinol Metab 32:76–94. https://doi.org/10.1016/j.tem.2020.11.007
Bi S, Yan L, Pang B, Wang Y (2012) Investigation of three flavonoids binding to bovine serum albumin using molecular fluorescence technique. J Lumin 132:132–140. https://doi.org/10.1016/j.jlumin.2011.08.014
Bortolato M, Besson F, Roux B (2002) An infrared study of the thermal and pH stabilities of the GPI-alkaline phosphatase from bovine intestine. Biochem Biophys Res Commun 292:874–879. https://doi.org/10.1006/bbrc.2002.6735
Cacita N, Nikolaou S (2016) Studying the interaction between trinuclear ruthenium complexes and human serum albumin by means of fluorescence quenching. J Lumin 169:115–120. https://doi.org/10.1016/j.jlumin.2015.08.066
Caro-Ramírez JY, Rivas MG, Gonzalez PJ, Williams PAM, Naso LG, Ferrer EG (2022) Copper (II) cation and bathophenanthroline coordination enhance therapeutic effects of naringenin against lung tumor cells. Biometals 35:1059–1076. https://doi.org/10.1007/s10534-022-00422-4
Celiz G, Suarez SA, Arias A, Molina J, Brondino CD, Doctorovich F (2019) Synthesis, structural elucidation and antiradical activity of a copper (II) naringenin complex. BioMetals 32(4):595–610. https://doi.org/10.1007/s10534-019-00187-3
Das S, Ghosh P, Koley S, Roy AS (2018) Binding of naringin and naringenin with hen egg white lysozyme: a spectroscopic investigation and molecular docking study. Spectrochim Acta Part a: Mol Biomol Spectros 192:211–221. https://doi.org/10.1016/j.saa.2017.11.015
Farajtabar A, Gharib F (2013) Spectral analysis of naringenin deprotonation in aqueous ethanol solutions. Chem Pap 67:538–545. https://doi.org/10.2478/s11696-013-0309-9
Ferreira H, Conradie MM, Von Eschwege KG, Conradie J (2017) Electrochemical and DFT study of the reduction of substituted phenanthrolines. Polyhedron 122:147–154. https://doi.org/10.1016/j.poly.2016.11.018
Ferrer EG, Salinas MV, Correa MJ, Naso LG, Barrio DA, Etcheverry SB, Williams PA (2006) Synthesis, characterization, antitumoral and osteogenic activities of quercetin vanadyl (IV) complexes. J Biol Inorg Chem 11:791–801. https://doi.org/10.1007/s00775-006-0122-9
Ferrer EG, Bosch A, Yantorno O, Baran EJ (2008) A spectroscopy approach for the study of the interactions of bioactive vanadium species with bovine serum albumin. Bioorg Med Chem 16:3878–3886. https://doi.org/10.1016/j.bmc.2008.01.060
Liu Q, Zhou P, Chen Y (2012) In vitro investigation of the interaction between pentachlorophenol and alkaline phosphatase by spectroscopic methods. Spectrochim Acta Part A 87:298–302. https://doi.org/10.1016/j.saa.2011.11.059
Martini N, Parente JE, Restrepo-Guerrero G, Franca CA, Piro OE, Echeverría GA, Ferrer EG (2021) Tetraacuo-bis-(N, N-dimethylacetamide-O) magnesium (II) chloride dehydrate. An option to improve magnesium effect on phosphatase stimulation and albumin binding. J Mol Struct 1223:129240. https://doi.org/10.1016/j.molstruc.2020.129240
Masuri S, Vaňhara P, Cabiddu MG, Moráň L, Havel J, Cadoni E, PivettaT. Copper(II), (2022) Phenanthroline-based complexes as potential anticancer drugs: a walkthrough on the mechanisms of action. Molecules 27:49. https://doi.org/10.3390/molecules27010049
McLauchlan CC, Peters BJ, Willsky GR, Crans DC (2015) Vanadium–phosphatase complexes: phosphatase inhibitors favor the trigonal bipyramidal transition state geometries. Coord Chem Rev 301:163–199. https://doi.org/10.1016/j.ccr.2014.12.012
Mukherjee M, Sardar PS, Ghorai SK, Samanta SK, Roy AS, Dasgupta S, Ghosh S (2013) (2013) A comparative study of interaction of tetracycline with several proteins using time resolved anisotropy, phosphorescence, docking and FRET. PLoS One 8:e60940. https://doi.org/10.1371/journal.pone.0060940.Print
Naso LG, Lezama L, Valcarcel M, Salado C, Villacé P, Kortazar D, Ferrer EG, Williams PAM (2016) Bovine serum albumin binding, antioxidant and anticancer properties of an oxidovanadium(IV) complex with luteolin. J Inorg Biochem 157:80–93. https://doi.org/10.1016/j.jlumin.2012.05.018
Parente JE, Naso LG, Jori K, Franca CA, da Costa Ferreira AM, Williams PA, Ferrer EG (2019) In vitro experiments and infrared spectroscopy analysis of acid and alkaline phosphatase inhibition by vanadium complexes. New J Chem 43:17603–17619. https://doi.org/10.1039/C9NJ01638D
Preeth DR, Saravanan S, Shairam M, Selvakumar N, Raja IS, Dhanasekaran A, Rajalakshmi S (2021) Bioactive Zinc (II) complex incorporated PCL/gelatin electrospun nanofiber enhanced bone tissue regeneration. Eur J Pharm Sci 160:105768. https://doi.org/10.1016/j.ejps.2021.105768
Rajalakshmi S, Vimalraj S, Saravanan S, Raj Preeth D, Shairam M, Anuradha D (2018) Synthesis and characterization ofsilibinin/phenanthroline/neocuproine copper (II) complexes for augmenting bone tissue regeneration: an in vitro analysis. J Biol Inorg Chem 23:753–762. https://doi.org/10.1007/s00775-018-1566-4
Ramesh P, Jagadeesan R, Sekaran S, Dhanasekaran A, Vimalraj S (2021) Flavonoids: classification, function, and molecular mechanisms involved in bone remodelling. Front Endocrinol 12:779638. https://doi.org/10.3389/fendo.2021.779638
Ross PD, Subramanian S (1981) Thermodynamics of protein association reactions: forces contributing to stability. Biochem 20:3096–3102. https://doi.org/10.1021/bi00514a017
Roy AS, Tripathy DR, Chatterjee A, Dasgupta S (2010) A spectroscopic study of the interaction of the antioxidant naringin with bovine serum albumin. J Biophys Chem 1:141–152. https://doi.org/10.4236/jbpc.2010.13017
Roy AS, Tripathy DR, Ghosh AK, Dasgupta S (2012) An alternate mode of binding of the polyphenol quercetin with serum albumins when complexed with Cu (II). J Lumin 132:2943–2951. https://doi.org/10.1016/j.jlumin.2012.05.018
Saboury AA (2009) Enzyme inhibition and activation: a general theory. J Iranian Chem Soc 6:219–229. https://doi.org/10.1007/BF03245829
Salehian P, Shareghi B (2019) Hosseini-Koupaei M (2019) Comparative studies on the interaction between biogenic polyamines and bovine intestinal alkaline phosphatases: spectroscopic and theoretical approaches. J Biol Phys 45:89–106. https://doi.org/10.1007/s10867-018-9517-4
Sarkar SN, Ghosh N (1996) Reversible unfolding of Escherichia coli alkaline phosphatase: active site can be reconstituted by a number of pathways. Arch Biochem Biophys 330:174–180. https://doi.org/10.1006/abbi.1996.0239
Sharma U, Pal D, Prasad R (2014) Alkaline phosphatase: an overview. Indian J Clinical Biochem 29:269–278. https://doi.org/10.1007/s12291-013-0408-y
Tamayo LV, Gouvea LR, Sousa AC, Albuquerque RM, Teixeira SF, de Azevedo RA, Beraldo H (2016) Copper (II) complexes with naringenin and hesperetin: cytotoxic activity against A 549 human lung adenocarcinoma cells and investigation on the mode of action. Biometals 29:39–52. https://doi.org/10.1007/s10534-015-9894-0
Tu AT (1982) Raman spectroscopy in biology Principles and Applications, 1st edn. Willey, New York, pp 65–116
Vimalraj S (2020) Alkaline phosphatase: structure, expression and its function in bone mineralization. Gene 754:144855. https://doi.org/10.1016/j.gene.2020.144855
Vimalraj S, Rajalakshmi S, Saravanan S, Preeth DR, Vasanthi RL, Shairam M, Chatterjee S (2018a) Synthesis and characterization of zinc-silibinin complexes: a potential bioactive compound with angiogenic, and antibacterial activity for bone tissue engineering. Colloids Surf B: Biointerfaces 167:134–143. https://doi.org/10.1016/j.colsurfb.2018.04.007
Vimalraj S, Rajalakshm S, Preeth DR, Kumar SV, DeepakT GV, Murugan K, Chatterjee S (2018b) Mixed-ligand copper(II) complex of quercetin regulate osteogenesis and angiogenesis. Mater Sci Eng C 83:187–194. https://doi.org/10.1016/j.msec.2017.09.005
Vimalraj S, Rajalakshmi S, Saravanan S, Deepak T, Murugan K, Vasanthi Rajkumare A, Anuradha D (2019) Zinc chelated morin promotes osteoblast differentiation over its uncomplexed counterpart. Process Biochem 82:167–172. https://doi.org/10.1016/j.procbio.2019.04.008
Vimalraj S, Saravanan S, Hariprabu G, Yuvashree R, Kanna SKA, Sujoy K, Anjali D (2020) Kaempferol-zinc (II) complex synthesis and evaluation of bone formation using zebrafish model. Life Sci 256:117993. https://doi.org/10.1016/j.lfs.2020.117993
Vimalraj S, Saravanan S, Subramanian R (2021) Rutin-Zn (II) complex promotes bone formation-A concise assessment in human dental pulp stem cells and zebrafish. Chem-Biol Interact 349:109674. https://doi.org/10.1016/j.cbi.2021.109674
Wang Y, Bai S, Cheng Q, Zeng Y, Xu X, Guan G (2021) Naringenin promotes SDF-1/CXCR4 signaling pathway in BMSCs osteogenic differentiation. Folia Histochem Et Cytobiol 59:66–73. https://doi.org/10.5603/FHC.a2021.0008
Xiao JB, Chen JW, Cao H, Rena FL, Yang CS, Chen Y, Xu M (2007) Study of the interaction between baicalin and bovine serum albumin by multi-spectroscopic method. J Photochem Photobiol a: Chem 191:222–227. https://doi.org/10.1016/j.jphotochem.2007.04.027
Zalatan JG, Fenn TD, Herschlag D (2008) Comparative enzymology in the alkaline phosphatase superfamily to determine the catalytic role of an active-site metal ion. J Mol Biol 384:1174–1189. https://doi.org/10.1016/j.jmb.2008.09.059
Zhou XQ, Wang BL, Kou SB, Lin ZY, Lou YY, Zhou KL, Shi JH (2019) Multi-spectroscopic approaches combined with theoretical calculation to explore the intermolecular interaction of telmisartan with bovine serum albumin. Chem Phys 522:285–293. https://doi.org/10.1016/j.chemphys.2019.03.019
Acknowledgements
This work was supported by Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT-PICT-2019-00945), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET-PIP1999-2021-2023), and Universidad Nacional de La Plata (UNLP-X929), Argentina. EGF is member of the Research Career, CONICET, Argentina. PAMW is member of the Research Career, CICPBA. JCR and GMG are CONICET fellowship holders.
Author information
Authors and Affiliations
Contributions
Conceptualization: PAMW, EGF Methodology: GMG, JYC-R, JEP Validation: PAMW, EGF Formal analysis: GMG, EGF Investigation: GMG, JYC-R, JEP Resources: PAMW, EGF Writing e original draft: EGF Writing e review and editing: PAMW, EGF Visualization: EGF Supervision: PAMW, EGF Project administration: PAMW, EGF Funding acquisition: PAMW, EGF.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no competing interests to declare that are relevant to the content of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Gaddi, G.M., Caro-Ramírez, J.Y., Parente, J.E. et al. Copper-flavonoid family of complexes involved in alkaline phosphatase activation. Biometals 36, 1221–1239 (2023). https://doi.org/10.1007/s10534-023-00511-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10534-023-00511-y